
Difference in Response of Caragana intermedia Photosynthesis to Soil Water Content in Different Afforestation Years and Related Threshold Effects in Alpine Sandy Lands
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Materials
2.3. Experimental Methods
2.4. Data Analysis
3. Results
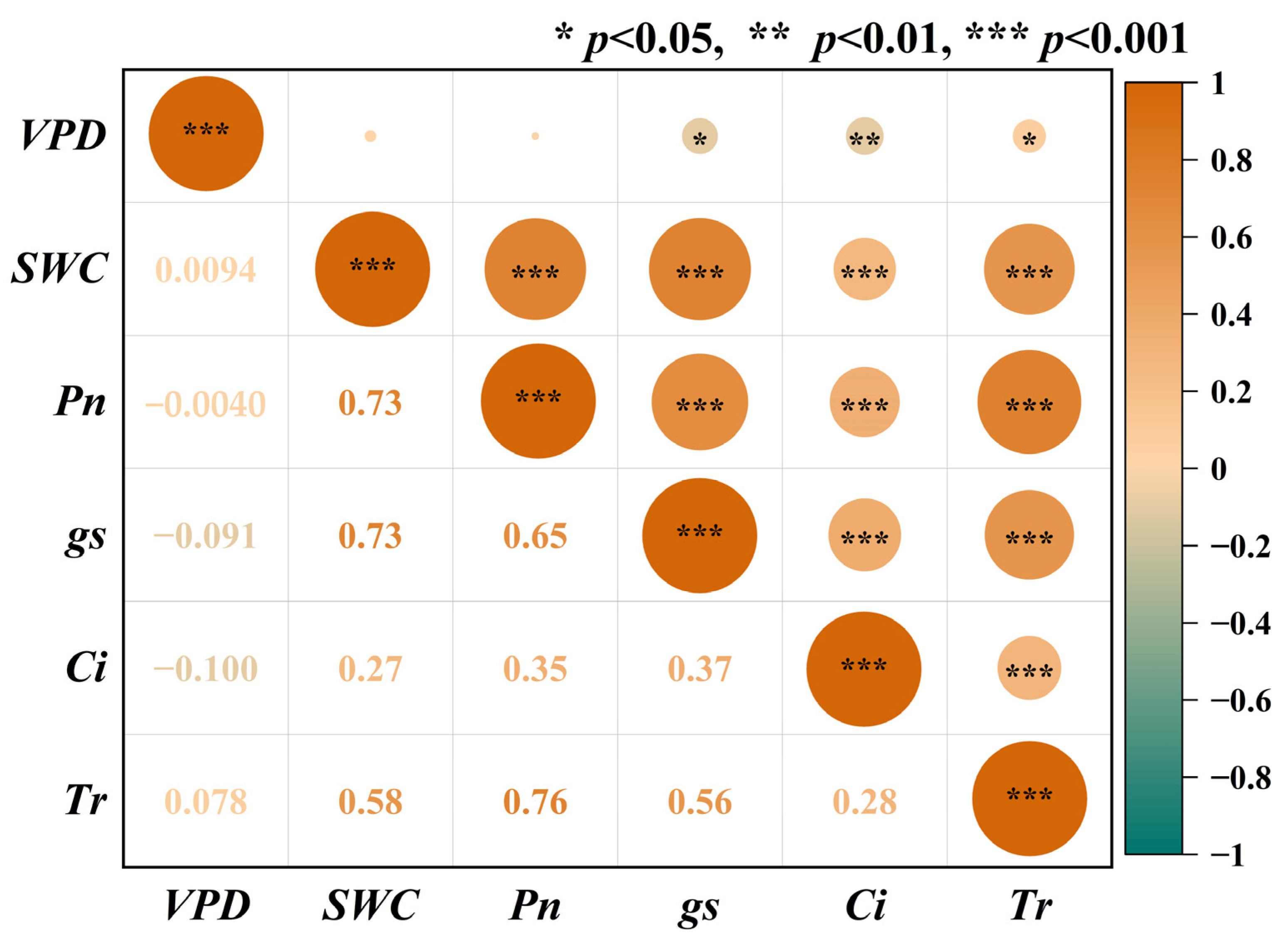
3.1. Response of Photosynthetic Parameters to SWC and VPD
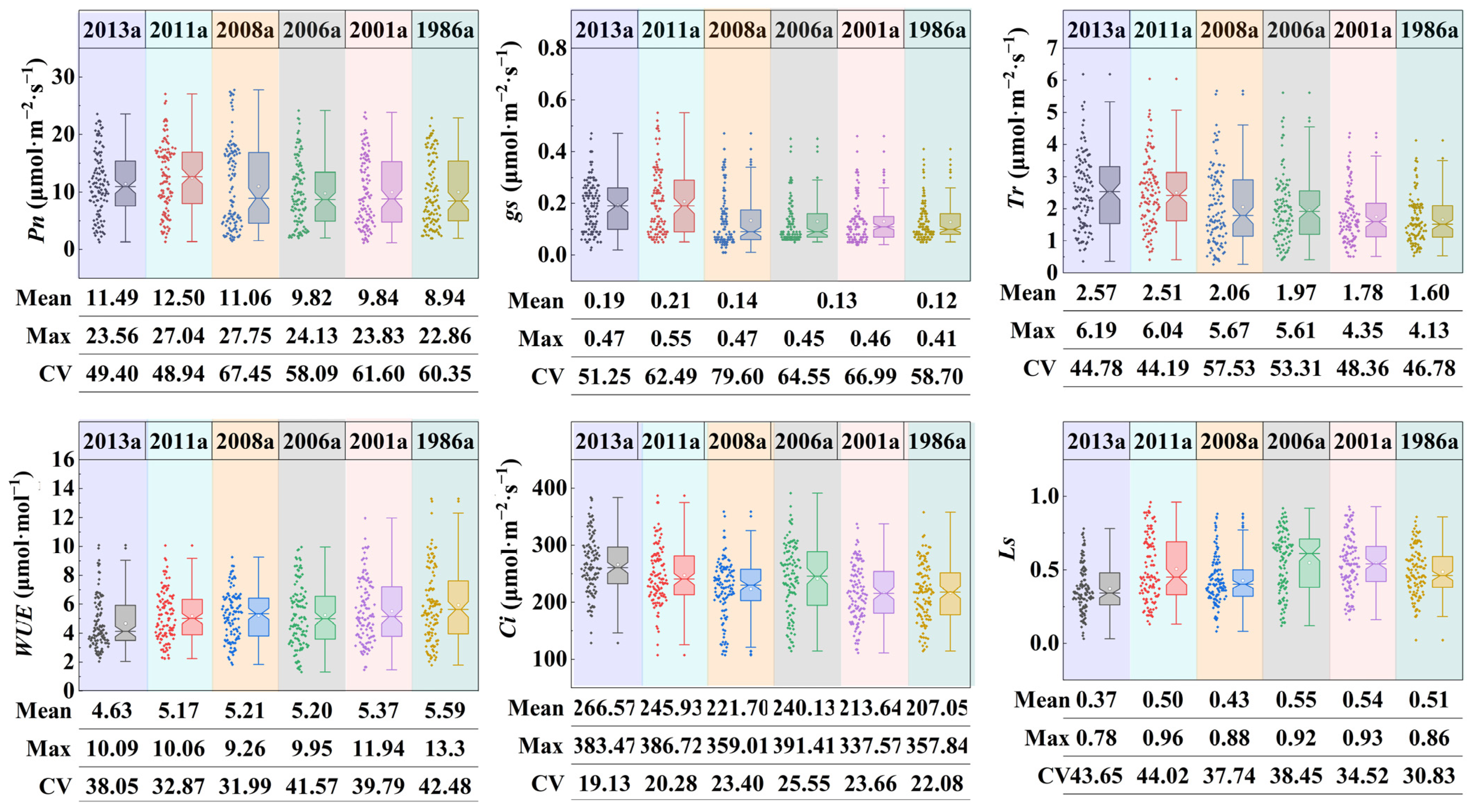
3.2. Characteristics of Photosynthetic Parameters of Plantations in Different Afforestation Years
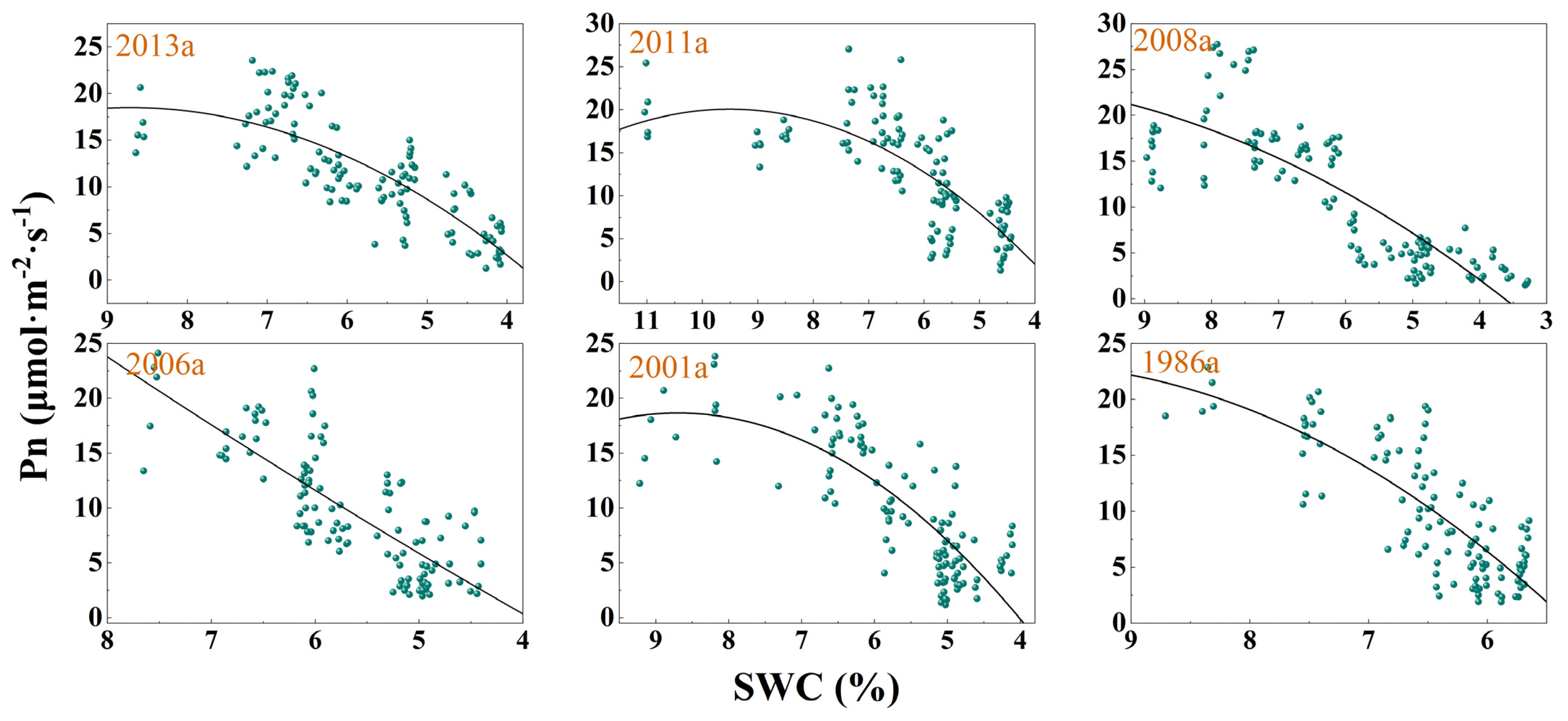
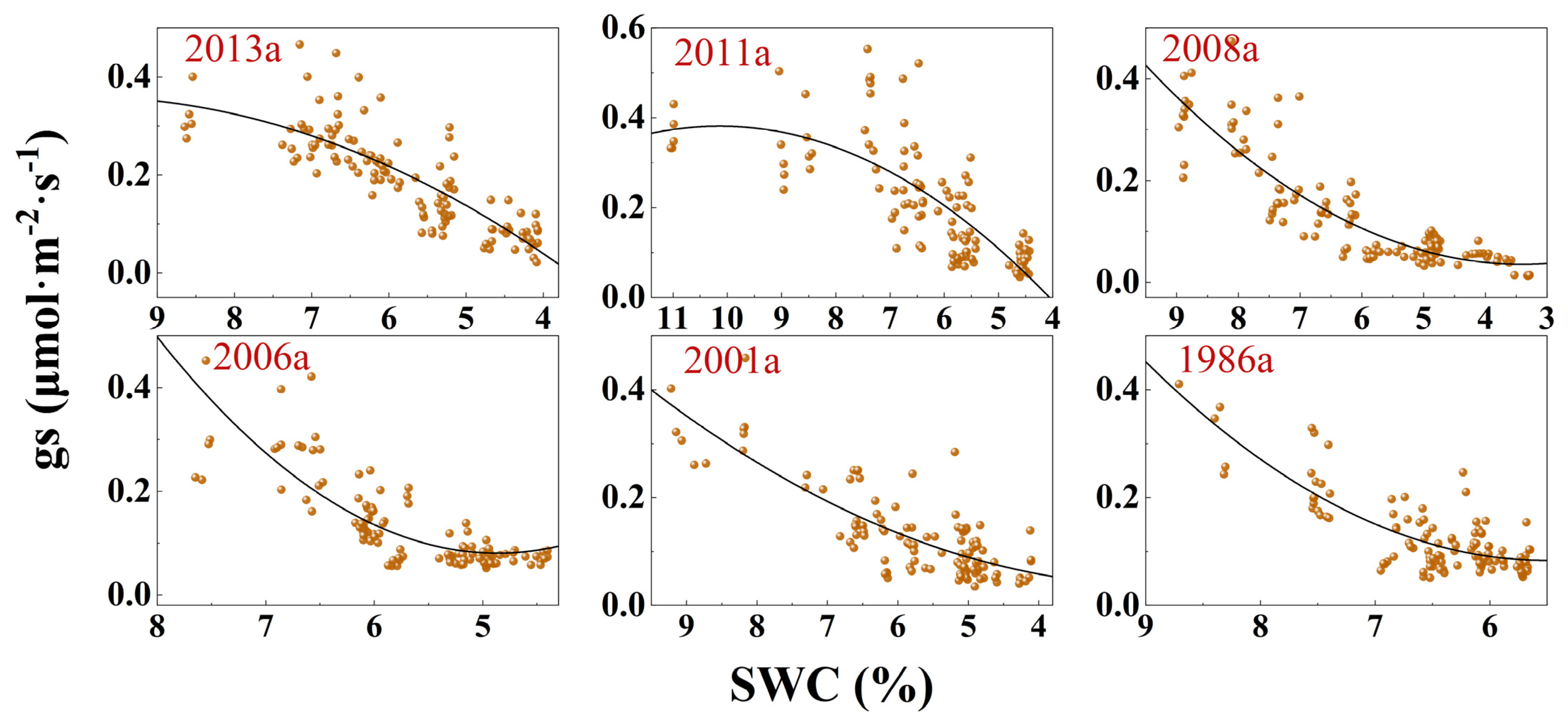
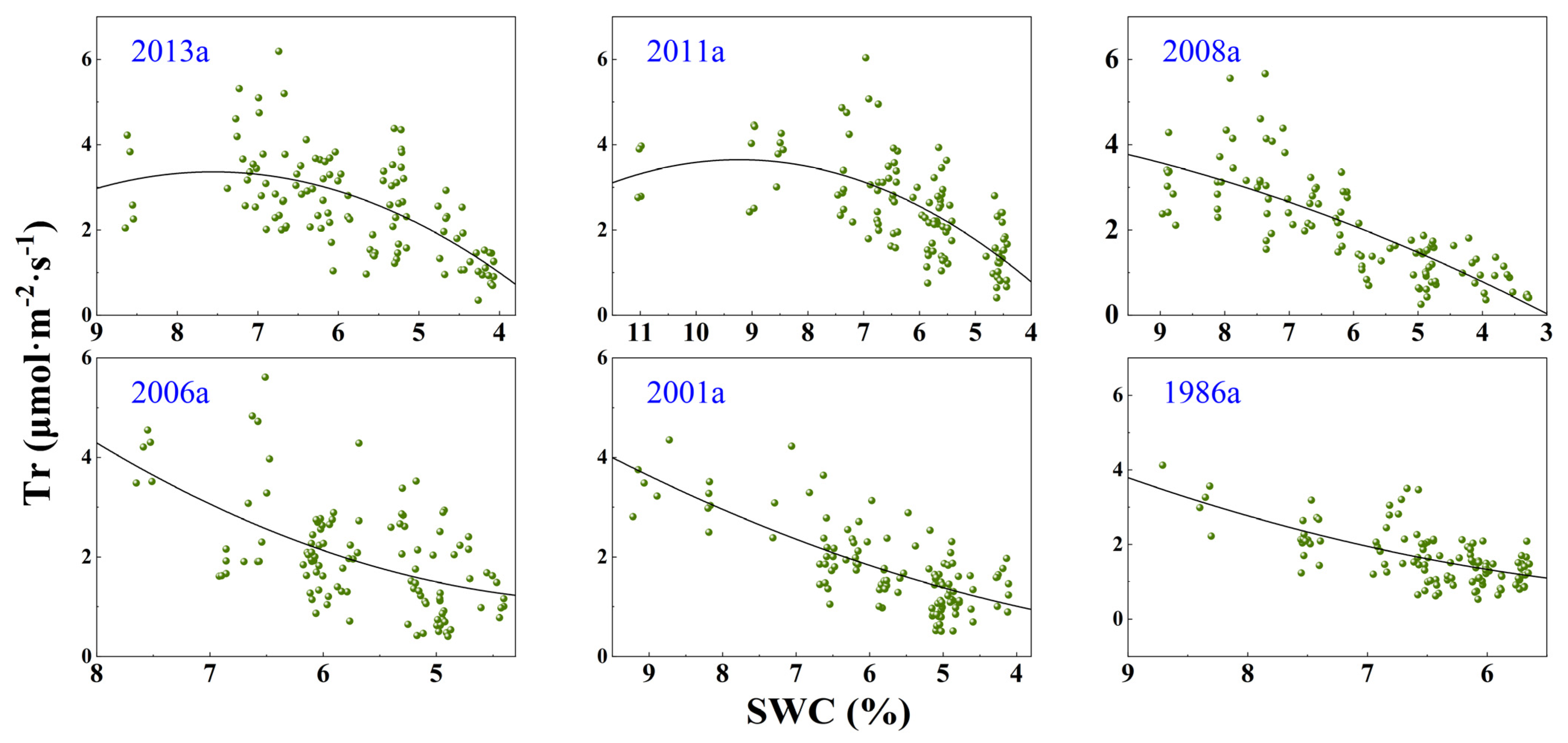
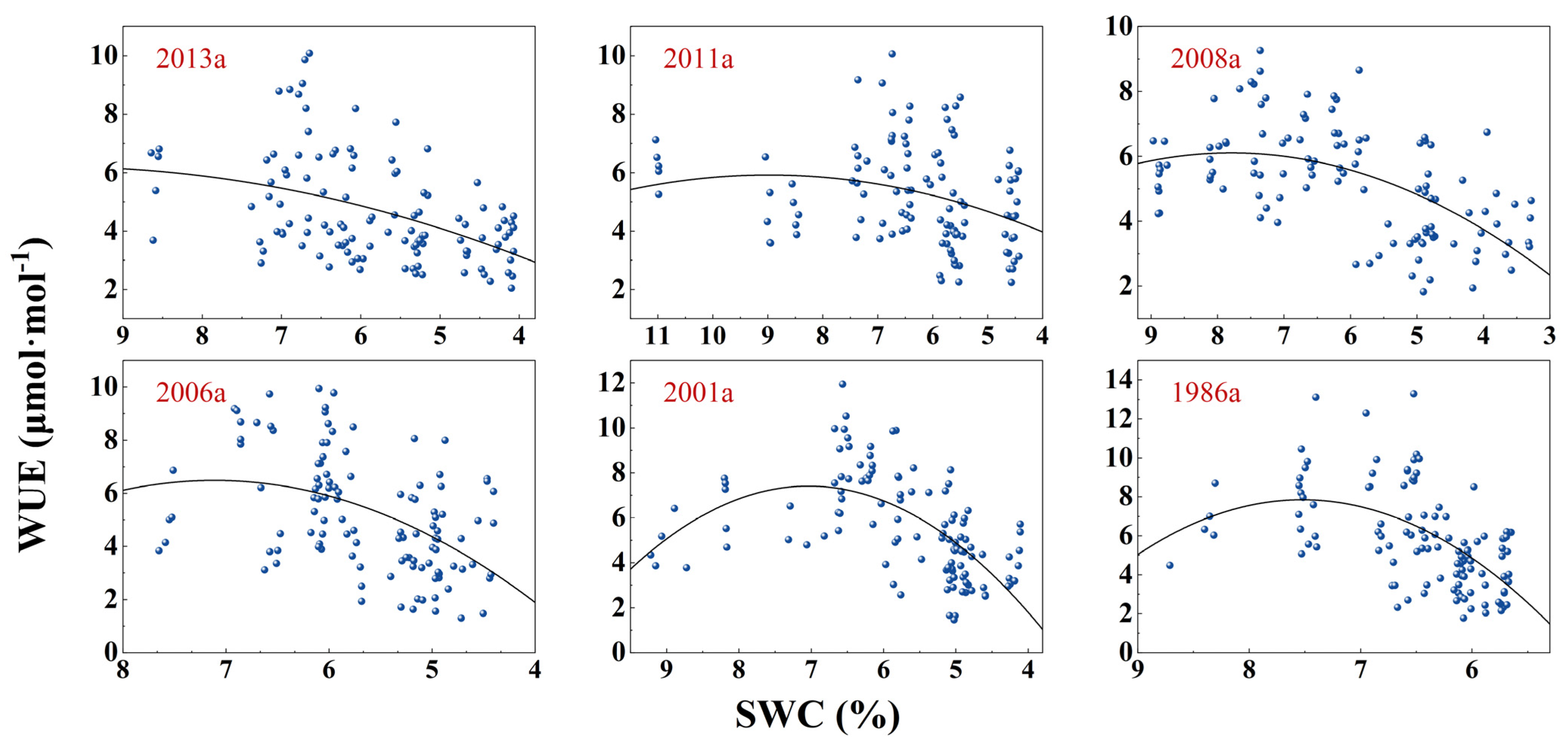
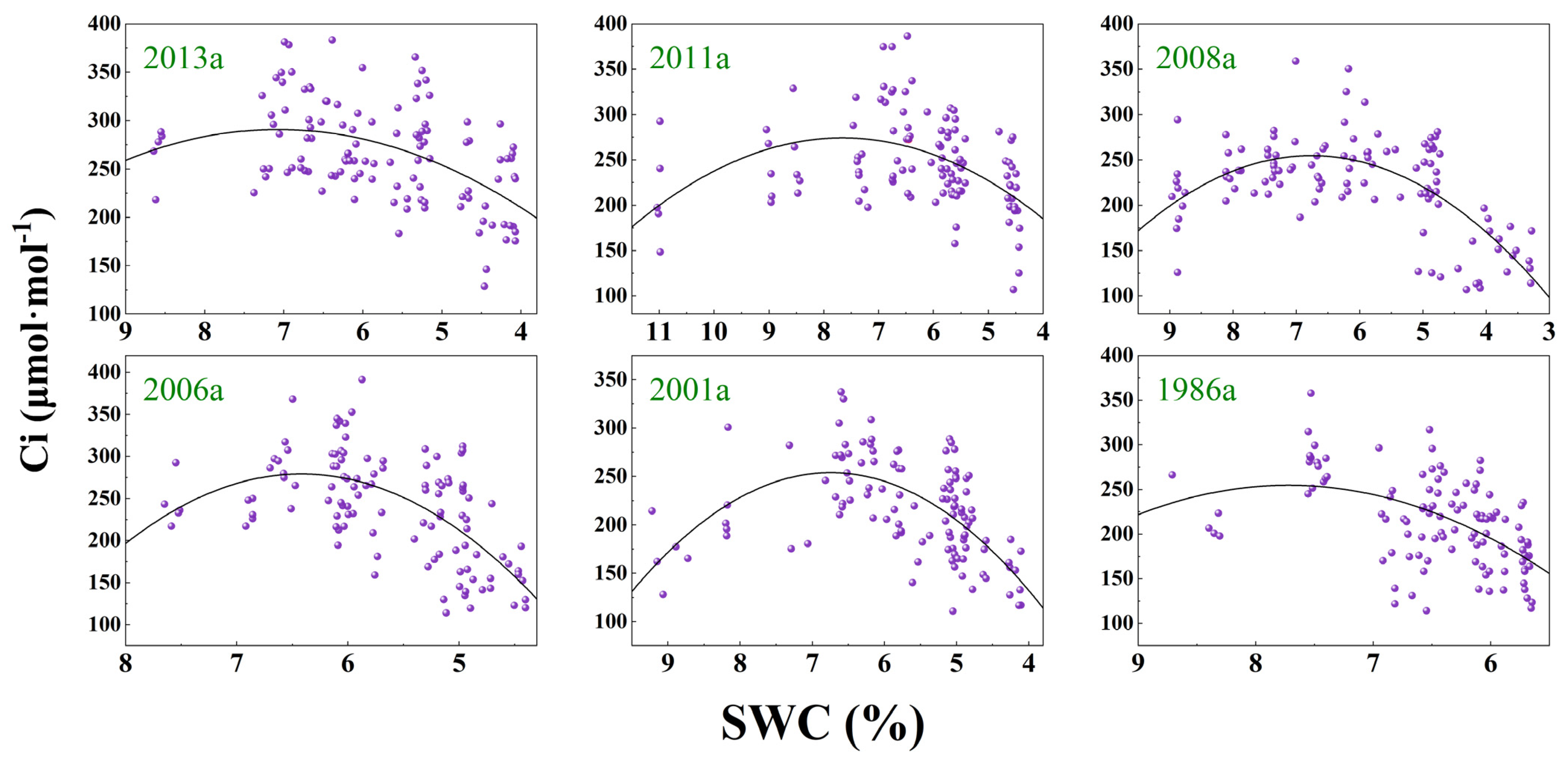
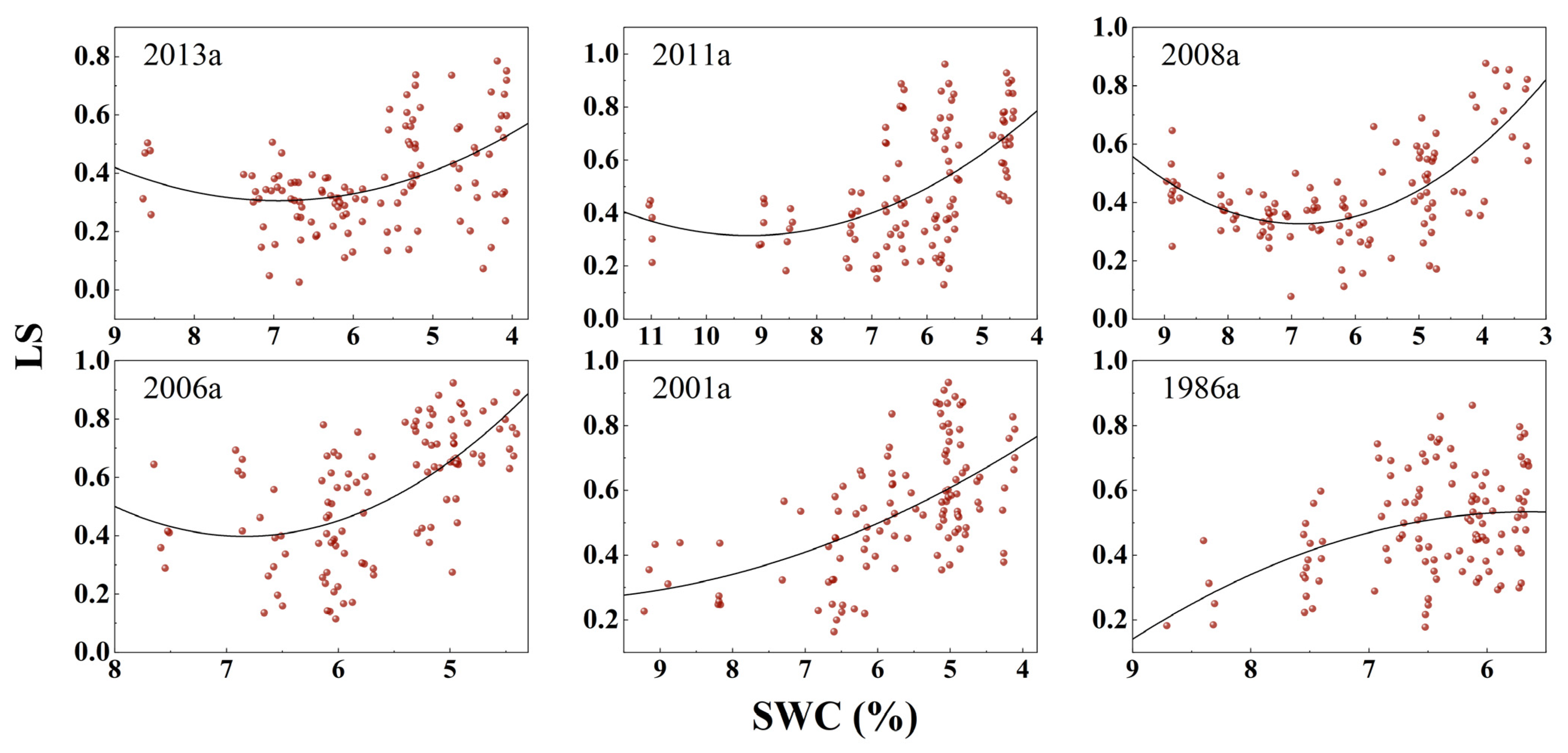
3.3. Response of Photosynthesis to SWC Changes
3.4. SWC Threshold Classification of Photosynthetic Parameters of C. intermedia
4. Discussion
4.1. Effect of SWC on Plant Gas Exchange Parameters
4.2. Difference in the SWC Threshold of C. Intermedia in Different Afforestation Years
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burrell, A.L.; Evans, J.P.; De Kauwe, M.G. Anthropogenic climate change has driven over 5 million km2 of drylands towards desertification. Nat. Commun. 2020, 11, 3853. [Google Scholar] [CrossRef] [PubMed]
- Bouras, E.; Jarlan, L.; Khabba, S.; Er-Raki, S.; Dezetter, A.; Sghir, F.; Tramblay, Y. Assessing the impact of global climate changes on irrigated wheat yields and water requirements in a semi-arid environment of Morocco. Sci. Rep. 2019, 9, 19142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.; He, Z.; Du, J.; Chen, L.; Zhu, X.; Li, J. Recent changes in daily climate extremes in an arid mountain region, a case study in northwestern China’s Qilian Mountains. Sci. Rep. 2017, 7, 2245. [Google Scholar] [CrossRef] [Green Version]
- Franz, T.E.; Caylor, K.K.; King, E.G.; Nordbotten, J.M.; Celia, M.A.; Rodriguez-Iturbel, I. An ecohydrological approach to predicting hillslope-scale vegetation patterns in dryland ecosystems. Water Resour. Res. 2012, 48, 1515. [Google Scholar] [CrossRef] [Green Version]
- Manzoni, S.; Vico, G.; Katul, G.; Palmroth, S.; Jackson, R.B.; Porporato, A. Hydraulic limits on maximum plant transpiration and the emergence of the safety-efficiency trade-off. New Phytol. 2013, 198, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Sendall, K.M.; Stefanski, A.; Rich, R.L.; Hobbie, S.E.; Montgomery, R.A. Effects of climate warming on photosynthesis in boreal tree species depend on soil moisture. Nature 2018, 562, 263–267. [Google Scholar] [CrossRef]
- Fu, Z.; Ciais, P.; Prentice, I.C.; Gentine, P.; Makowski, D.; Bastos, A.; Luo, X.; Green, J.K.; Stoy, P.C.; Yang, H.; et al. Atmospheric dryness reduces photosynthesis along a large range of soil water deficits. Nat. Commun. 2022, 13, 989. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Tian, F.P.; Wu, G.L. Soil water availability threshold indicator was determined by using plant physiological responses under drought conditions. Ecol Indic. 2020, 118, 106740. [Google Scholar] [CrossRef]
- Qing, Y.; Wang, S.; Ancell, B.C.; Yang, Z.L. Accelerating flash droughts induced by the joint influence of soil moisture depletion and atmospheric aridity. Nat. Commun. 2022, 13, 1139. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Migliavacca, M.; Forkel, M.; Denissen, J.M.C.; Reichstein, M.; Yang, H.; Duveiller, G.; Weber, U.; Orth, R. Widespread increasing vegetation sensitivity to soil moisture. Nat. Commun. 2022, 13, 3959. [Google Scholar] [CrossRef]
- Hicke, J.A.; Zeppel, M.J.B. Climate-driven tree mortality: Insights from the pinon pine die-off in the United States. New Phytol. 2013, 200, 301–303. [Google Scholar] [CrossRef]
- Zhou, S.; Williams, A.P.; Lintner, B.R.; Berg, A.M.; Zhang, Y.; Keenan, T.F.; Cook, B.I.; Hagemann, S.; Seneviratne, S.I.; Gentine, P. Soil moisture–atmosphere feedbacks mitigate declining water availability in drylands. Nat. Clim. Change 2021, 11, 38–44. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Cui, Z.; Fang, Y.; He, H.; Liu, B.-R.; Wu, G.-L. Soil water storage deficit of alfalfa (Medicago sativa) grasslands along ages in arid area (China). Field Crops Res. 2018, 221, 1–6. [Google Scholar] [CrossRef]
- Humphrey, V.; Berg, A.; Ciais, P.; Gentine, P.; Jung, M.; Reichstein, M.; Seneviratne, S.I.; Frankenberg, C. Soil moisture–atmosphere feedback dominates land carbon uptake variability. Nature 2021, 592, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Kumarathunge, D.P.; Drake, J.E.; Tjoelker, M.G.; Lopez, R.; Pfautsch, S.; Varhammar, A.; Medlyn, B.E. The temperature optima for tree seedling photosynthesis and growth depend on water inputs. Global. Change Biol. 2020, 26, 2544–2560. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Zhong, Y.; Shangguan, Z. Responses of different physiological parameter thresholds to soil water availability in four plant species during prolonged drought. Agric. For. Meteorol. 2017, 247, 311–319. [Google Scholar] [CrossRef]
- Zhang, G.C.; Xia, J.B.; Shao, H.B.; Zhang, S.Y. Grading Woodland Soil Water Productivity and Soil Bioavailability in the Semi-Arid Loess Plateau of China. Clean-Soil Air Water 2012, 40, 148–153. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Zhang, G.C.; Gu, S.Y.; Xia, J.B.; Zhao, J.K. Critical responses of photosynthetic efficiency of goldspur apple tree to soil water variation in semiarid loess hilly area. Photosynthetica 2010, 48, 589–595. [Google Scholar] [CrossRef]
- Li, Q.; Jia, Z.; Zhu, Y.; Wang, Y.; Li, H.; Yang, D.; Zhao, X. Spatial Heterogeneity of Soil Nutrients after the Establishment of Caragana intermedia Plantation on Sand Dunes in Alpine Sandy Land of the Tibet Plateau. PLoS ONE 2019, 14, e0218503. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Wang, Y.; Li, Y.; Cheng, H. Influences of alpine ecosystem responses to climatic change on soil properties on the Qinghai-Tibet Plateau, China. Catena 2007, 70, 506–514. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Fu, B.; Wang, S.; Stringer, L.C.; Wang, Y.; Li, Z.; Liu, Y.; Zhou, W. Drivers and impacts of changes in China’s drylands. Nat. Rev. Earth. Environ. 2021, 2, 858–873. [Google Scholar] [CrossRef]
- Yan, W.; Deng, L.; Zhong, Y.; Shangguan, Z. The Characters of Dry Soil Layer on the Loess Plateau in China and Their Influencing Factors. PLoS ONE 2015, 10, e0134902. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Xiao, J.; Zhang, Z.; Ollinger, S.V.; Hollinger, D.Y.; Pan, Y.; Wan, J. Canopy photosynthetic capacity drives contrasting age dynamics of resource use efficiencies between mature temperate evergreen and deciduous forests. Global. Change Biol. 2020, 26, 6156–6167. [Google Scholar] [CrossRef]
- Yu, Y.; Zheng, W.; Zhong, X.; Ying, B. Stoichiometric characteristics in Zanthoxylum planispinum var. dintanensis plantation of different ages. Agron. J. 2021, 113, 685–695. [Google Scholar]
- Au, T.F.; Maxwell, J.T.; Robeson, S.M.; Li, J.; Siani, S.M.; Novick, K.A.; Dannenberg, M.P.; Phillips, R.P.; Li, T.; Chen, Z. Younger trees in the upper canopy are more sensitive but also more resilient to drought. Nat. Clim. Change 2022, 12, 1168–1174. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, H.; Jin, H.; Chen, F. Holocene Moisture Variation Recorded by Aeolian Sand-Palaeosol Sequences of the Gonghe Basin, Northeastern Qinghai-Tibetan Plateau, China. Acta Geol. Sin. Engl. 2020, 94, 668–681. [Google Scholar] [CrossRef]
- Gao, Y.; He, L.X.Z.; Jia, Z.Q.; Li, Q.X.; Dai, J. Effects of precipitation on water use characteristics of Caragana intermedia plantations with different stand ages in alpine sandy land. Chin. J. Appl. Ecol. 2021, 32, 1935–1942. (In Chinese) [Google Scholar]
- Liu, L.; Gudmundsson, L.; Hauser, M.; Qin, D.; Li, S.; Seneviratne, S.I. Soil moisture dominates dryness stress on ecosystem production globally. Nat. Commun. 2020, 11, 4892. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.Y.; Pang, X.Y.; Peuke, A.D.; Wang, X.; Chen, K.; Gong, R.G. Growth and photosynthetic responses in Jatropha curcas L. seedlings of different provenances to watering regimes. Photosynthetica 2016, 54, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Gong, C.M.; Bai, J.; Wang, J.H.; Zhou, Y.L.; Kang, T.; Wang, J.J.; Hu, C.X.; Guo, H.B.; Chen, P.L.; Xie, P.; et al. Carbon Storage Patterns of Caragana korshinskii in Areas of Reduced Environmental Moisture on the Loess Plateau, China. Sci. Rep. 2016, 6, 28883. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Zhong, Y.; Shangguan, Z. A meta-analysis of leaf gas exchange and water status responses to drought. Sci. Rep. 2016, 6, 20917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Miguez-Macho, G.; Jobbagy, E.G.; Jackson, R.B.; Otero-Casal, C. Hydrologic regulation of plant rooting depth. Proc. Natl. Acad. Sci. USA 2017, 114, 10572–10577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcausland, L.; Vialet-Chabrand, S.; Davey, P.; Baker, N.R.; Brendel, O.; Lawson, T. Effects of kinetics of light-induced stomatal responses on photosynthesis and water-use efficiency. New Phytol. 2016, 211, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Bertolino, L.T.; Caine, R.S.; Gray, J.E. Impact of Stomatal Density and Morphology on Water-Use Efficiency in a Changing World. Front. Plant Sci. 2019, 10, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miner, G.L.; Bauerle, W.L.; Baldocchi, D.D. Estimating the sensitivity of stomatal conductance to photosynthesis: A review. Plant Cell Environ. 2017, 40, 1214–1238. [Google Scholar] [CrossRef] [PubMed]
- Drake, J.E.; Raetz, L.; Davis, S.C.; Delucia, E.H. Hydraulic limitation not declining nitrogen availability causes the age-related photosynthetic decline in loblolly pine (Pinus taeda L.). Plant Cell Environ. 2010, 33, 1756–1766. [Google Scholar] [CrossRef] [PubMed]
- Steppe, K.; Niinemets, Ü.; Teskey, R.O. Tree size-and age-related changes in leaf physiology and their influence on carbon gain. In Size-and Age-Related Changes in Tree Structure and Function; Springer: Dordrecht, The Netherlands, 2011; pp. 235–253. [Google Scholar]
- Bao, J.T.; Wang, J.; Li, X.R.; Zhang, Z.S.; Su, J.Q. Age-related changes in photosynthesis and water relations of revegetated Caragana korshinskii in the Tengger desert, Northern China. Trees 2015, 29, 1749–1760. [Google Scholar] [CrossRef]
- Garg, A.; Bordoloi, S.; Ganesan, S.P.; Sekharan, S.; Sahoo, L. A relook into plant wilting: Observational evidence based on unsaturated soil–plant-photosynthesis interaction. Sci. Rep. 2020, 10, 22064. [Google Scholar] [CrossRef]
- Drake, J.E.; Davis, S.C.; Raetz, L.M.; DeLucia, E.H. Mechanisms of age-related changes in forest production: The influence of physiological and successional changes. Global. Change Biol. 2011, 17, 1522–1535. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Afforestation Years | Habitat | Soil Type | Altitud/m | Plant Height/cm | Base Diameter/cm | Crown Width/% | Vegetation Coverage/% | Row Spacing/m |
|---|---|---|---|---|---|---|---|---|
| 2013a | Semi-fixed dune | Aeolian soil | 2873 | 84.27 ± 31.06 | 0.51 ± 0.01 | 92.15 ± 20.38 | 33 | 1 |
| 2011a | Semi-fixed dune | Aeolian soil | 2877 | 95.33 + 12.45 | 0.71 + 0.02 | 120.51 ± 7.29 | 50 | 2 |
| 2008a | Fixed dune | Aeolian soil | 2886 | 135.20 ± 10.04 | 0.74 ± 0.01 | 124.28 ± 11.93 | 62 | 2 |
| 2006a | Fixed dune | Aeolian soil | 2884 | 145.00 + 5.77 | 1.46 + 0.03 | 132.17 + 6.83 | 70 | 2 |
| 2001a | Fixed dune | Aeolian soil | 2882 | 165.60 ± 12.65 | 1.27 ± 0.01 | 153.67 ± 24.30 | 83 | 2 |
| 1986a | Fixed dune | Aeolian soil | 2883 | 180.67 ± 42.43 | 1.50 ± 0.03 | 178.33 ± 59.28 | 71 | 2 |
| T (°C) | RH (%) | SR (mv/km·m−2) | VPD (kPa) | WS (m/s) | |
|---|---|---|---|---|---|
| Mean | 17.89 | 52.58 | 0.57 | 1.06 | 2.12 |
| Stdev.p | 3.80 | 17.22 | 0.23 | 0.57 | 0.73 |
| CV | 21.26 | 32.76 | 40.30 | 53.93 | 34.20 |
| Pn | Gs | Ci | Tr | Wue | Ls | |
|---|---|---|---|---|---|---|
| Afforestation year | −0.836 * | −0.709 | −0.805 | −0.871 * | 0.820 * | 0.477 |
| WCP-Pn | WCP-gs | WCP-Tr | WSP-Pn | WSP-gs | WSP-Tr | |
|---|---|---|---|---|---|---|
| 2013a | 3.62 | 3.64 | 3.31 | 7.19 | 7.16 | 6.74 |
| 2011a | 3.70 | 4.06 | 3.33 | 7.36 | 7.42 | 6.96 |
| 2008a | 3.63 | 3.38 | 3.08 | 7.91 | 8.11 | 7.37 |
| 2006a | 3.85 | 3.90 | 3.36 | 7.51 | 7.55 | 6.51 |
| 2001a | 4.02 | 2.13 | 1.45 | 8.19 | 8.17 | 8.72 |
| 1986a | 5.31 | 5.50 | 3.39 | 8.35 | 8.71 | 8.71 |
| Afforestation Year | Model | Fitting Equation | F | DF | p | R2 |
|---|---|---|---|---|---|---|
| 2013a | y(Pn)~x(SWC) | y = −0.713x2 + 12.423x − 35.640 | 115.8 | 115 | <0.0001 | 0.6624 |
| y(gs)~x(SWC) | y = −0.009x2 + 0.179x − 0.532 | 119.9 | 113 | <0.0001 | 0.6741 | |
| y(Ci)~x(SWC) | y = −8.554x2 + 120.979x − 136.969 | 17.87 | 113 | <0.0001 | 0.2269 | |
| y(Tr)~x(SWC) | y = −0.187x2 + 2.825x − 7.3104 | 37.94 | 113 | <0.0001 | 0.3911 | |
| y(WUE)~x(SWC) | y = −0.088x2 + 1.743x − 2.418 | 14.31 | 113 | <0.0001 | 0.1879 | |
| y(Ls)~x(SWC) | y = 0.027x2 − 0.375x + 1.607 | 13.9 | 113 | <0.0001 | 0.1832 | |
| 2011a | y(Pn)~x(SWC) | y = −0.594x2 + 11.289x − 33.626 | 71.52 | 113 | <0.0001 | 0.5509 |
| y(gs)~x(SWC) | y = −0.103x2 + 0.209x − 0.677 | 77.85 | 109 | <0.0001 | 0.5806 | |
| y(Ci)~x(SWC) | y = −6.646x2 + 101.823x − 115.844 | 16.76 | 109 | <0.0001 | 0.2228 | |
| y(Tr)~x(SWC) | y = −0.105x2 + 1.936x − 5.284 | 42.28 | 109 | <0.0001 | 0.4266 | |
| y(WUE)~x(SWC) | y = −0.078x2 + 1.408x − 0.410 | 5.07 | 109 | <0.05 | 0.0684 | |
| y(Ls)~x(SWC) | y = 0.017x2 − 0.316x + 1.776 | 22.66 | 109 | <0.0001 | 0.2807 | |
| 2008a | y(Pn)~x(SWC) | y = −0.329x2 + 7.9913x − 24.493 | 132.7 | 114 | <0.0001 | 0.6943 |
| y(gs)~x(SWC) | y = 0.011x2 − 0.071x + 0.156 | 200.7 | 114 | <0.0001 | 0.775 | |
| y(Ci)~x(SWC) | y = −11.052x2 + 149.467x − 250.611 | 44.83 | 110 | <0.0001 | 0.439 | |
| y(Tr)~x(SWC) | y = −0.037x2 + 1.052x − 2.885 | 91.53 | 110 | <0.0001 | 0.6178 | |
| y(WUE)~x(SWC) | y = −0.149x2 + 2.360x − 3.238 | 23.07 | 110 | <0.0001 | 0.2827 | |
| y(Ls)~x(SWC) | y = 0.033x2 − 0.455x + 1.886 | 47.36 | 110 | <0.0001 | 0.4529 | |
| 2001a | y(Pn)~x(SWC) | y = 0.142x2 + 4.030x − 17.647 | 87.04 | 111 | <0.0001 | 0.6036 |
| y(gs)~x(SWC) | y = 0.022x2 − 0.1735x + 0.3873 | 114.7 | 111 | <0.0001 | 0.6681 | |
| y(Ci)~x(SWC) | y = −33.09x2 + 424.76x − 1084.06 | 35.5 | 111 | <0.0001 | 0.3791 | |
| y(Tr)~x(SWC) | y = 0.148x2 − 0.997x + 2.773 | 25.13 | 111 | <0.0001 | 0.2992 | |
| y(WUE)~x(SWC) | y = −0.475x2 + 6.753x − 17.527 | 15.23 | 111 | <0.0001 | 0.2012 | |
| y(Ls)~x(SWC) | y = 0.076x2 − 1.040x + 3.963 | 34.36 | 111 | <0.0001 | 0.3712 | |
| 2006a | y(Pn)~x(SWC) | y = −0.851x2 + 14.802x − 45.729 | 100.3 | 111 | <0.0001 | 0.6373 |
| y(gs)~x(SWC) | y = 0.007x2 − 0.029x + 0.065 | 114.4 | 111 | <0.0001 | 0.6675 | |
| y(Ci)~x(SWC) | y = −16.178x2 + 218.225x − 481.931 | 36.96 | 111 | <0.0001 | 0.389 | |
| y(Tr)~x(SWC) | y = 0.0215x2 + 0.2539x − 0.4117 | 67.69 | 111 | <0.0001 | 0.5708 | |
| y(WUE)~x(SWC) | y = −0.609x2 + 8.570x − 22.746 | 36.48 | 111 | <0.0001 | 0.3857 | |
| y(Ls)~x(SWC) | y = 0.012x2 − 0.245x + 1.467 | 28.51 | 111 | <0.0001 | 0.3275 | |
| 1986a | y(Pn)~x(SWC) | y = −1.077x2 + 21.437x − 83.468 | 101.4 | 112 | <0.0001 | 0.6379 |
| y(gs)~x(SWC) | y = 0.030x2 − 0.332x + 0.996 | 116.5 | 112 | <0.0001 | 0.6696 | |
| y(Ci)~x(SWC) | y = −20.116x2 + 310.891x − 946.404 | 22.82 | 112 | <0.0001 | 0.2769 | |
| y(Tr)~x(SWC) | y = 0.101x2 − 0.681x + 1.790 | 42.63 | 112 | <0.0001 | 0.4221 | |
| y(WUE)~x(SWC) | y = −1.289x2 + 19.379x − 64.998 | 25.12 | 112 | <0.0001 | 0.2974 | |
| y(Ls)~x(SWC) | y = −0.035x2 + 0.3965x − 0.593 | 9.94 | 112 | <0.0001 | 0.1356 |
| Afforestation Year | NPNEW | WSP-Pn | WSP-WUE | HPHAW |
|---|---|---|---|---|
| 2013a | 3.31–3.64% | 7.19% | 6.65% | 6.65–7.19% |
| 2011a | 3.33–4.06% | 7.36% | 6.74% | 6.74–7.36% |
| 2008a | 3.08–3.63% | 7.91% | 7.36% | 7.36–7.91% |
| 2006a | 3.36–3.85% | 7.51% | 6.10% | 6.10–7.51% |
| 2001a | 1.45–4.02% | 8.19% | 6.57% | 6.57–8.19% |
| 1986a | 3.39–5.50% | 8.35% | 6.52% | 6.52–8.35% |
| Afforestation Year | Min-SWC (%) | Max-SWC (%) | Mean-SWC (%) | Mode-SWC (%) |
|---|---|---|---|---|
| 2013a | 4.05 | 9.7 | 6.01 | 4–5 |
| 2011a | 4.4 | 12.35 | 6.87 | 5–7 |
| 2008a | 3.125 | 10.3 | 5.91 | 4–5 |
| 2006a | 3.95 | 10.5 | 5.88 | 4–5 |
| 2001a | 3.45 | 12.6 | 6.07 | 4–5 |
| 1986a | 5.15 | 10.55 | 6.75 | 6–7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Jia, Z.; Wu, R.; He, L.; Liu, T.; Li, Q.; Dai, J.; Zhang, J.; Wang, L. Difference in Response of Caragana intermedia Photosynthesis to Soil Water Content in Different Afforestation Years and Related Threshold Effects in Alpine Sandy Lands. Forests 2023, 14, 701. https://doi.org/10.3390/f14040701
Gao Y, Jia Z, Wu R, He L, Liu T, Li Q, Dai J, Zhang J, Wang L. Difference in Response of Caragana intermedia Photosynthesis to Soil Water Content in Different Afforestation Years and Related Threshold Effects in Alpine Sandy Lands. Forests. 2023; 14(4):701. https://doi.org/10.3390/f14040701
Chicago/Turabian StyleGao, Ya, Zhiqing Jia, Rina Wu, Lingxianzi He, Tao Liu, Qingxue Li, Jie Dai, Jiapeng Zhang, and Long Wang. 2023. "Difference in Response of Caragana intermedia Photosynthesis to Soil Water Content in Different Afforestation Years and Related Threshold Effects in Alpine Sandy Lands" Forests 14, no. 4: 701. https://doi.org/10.3390/f14040701