
Leaf Litter Breakdown and Soil Microbes in Catalpa bungei Plantations in Response to Various Fertilization Regimes
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Plot and Tree Species
2.2. Fertilization
2.3. Leaf Litter Collection and Decomposition
2.4. Soil Sampling
2.5. Elemental Analysis of Leaf Litter and Soil
2.6. Molecular Analysis of Soil Microbes
2.7. Data Analysis
3. Results
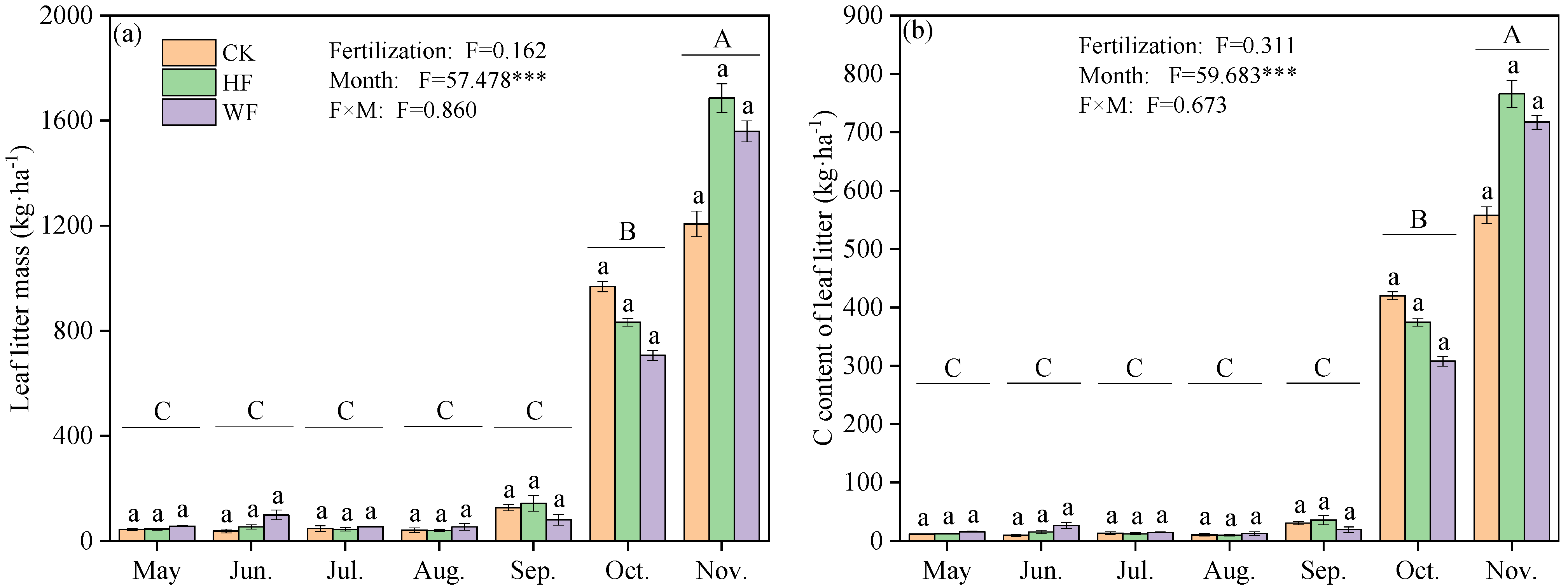
3.1. Biomass and C Content of Leaf Litter
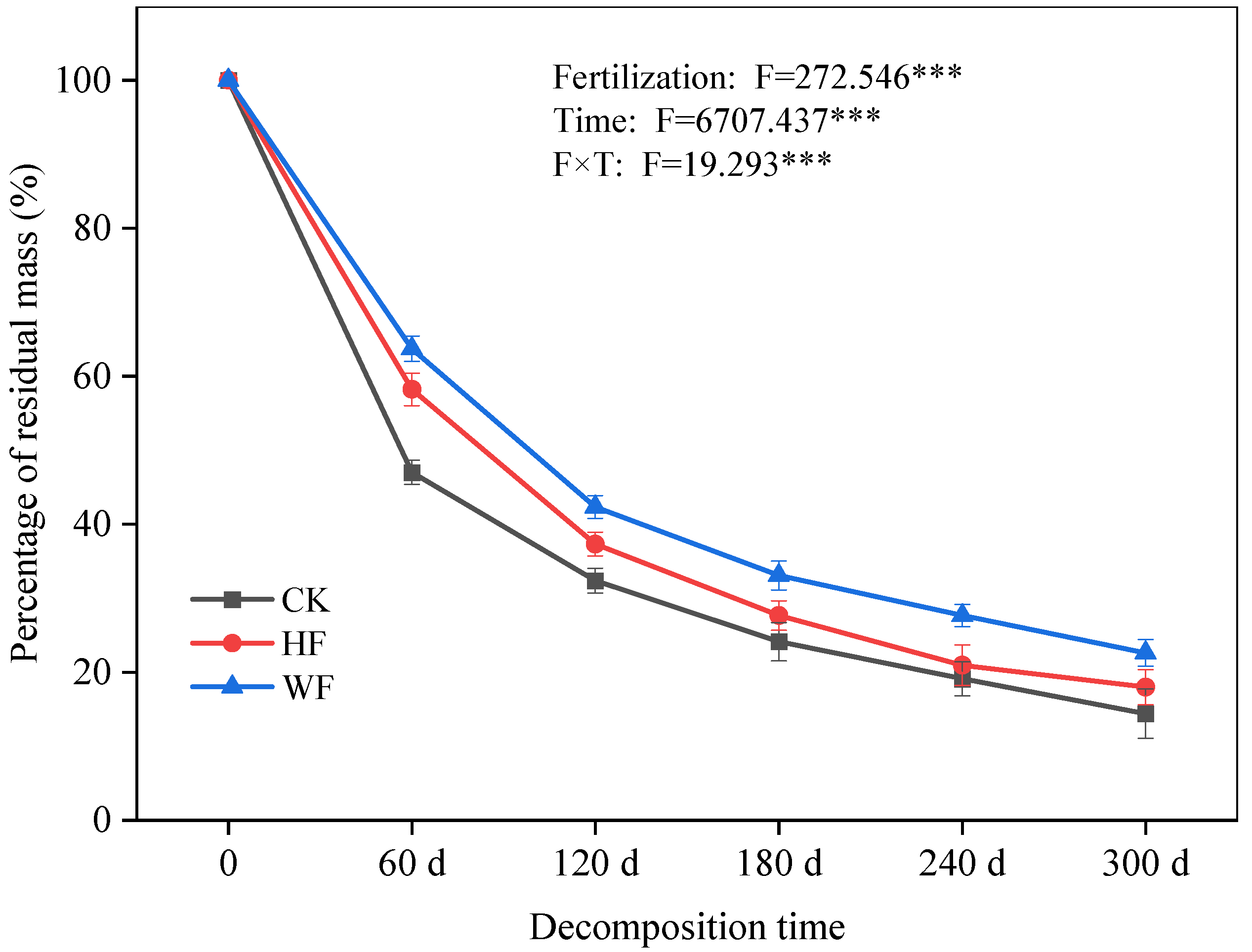
3.2. Decomposition of Leaf Litter
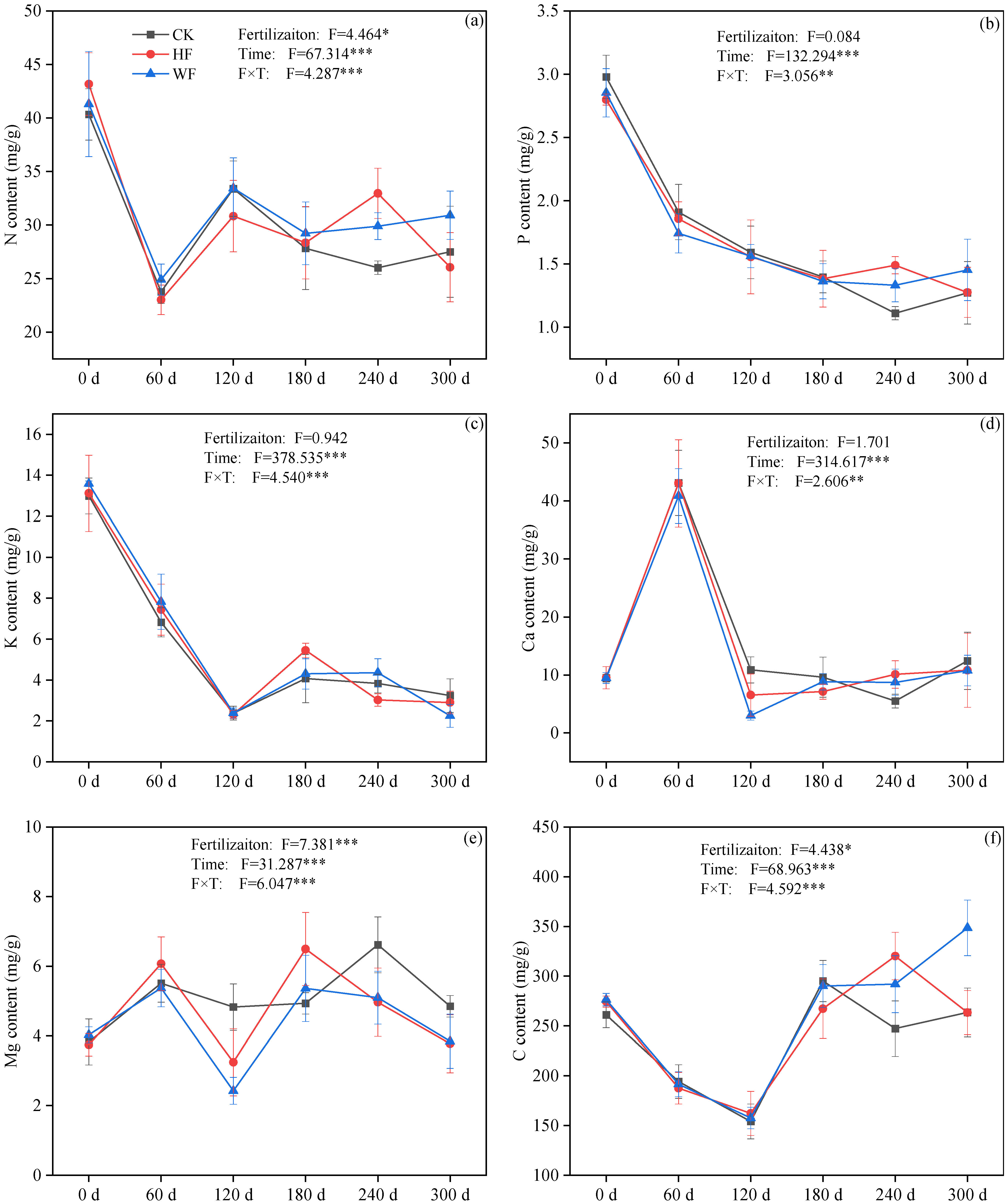
3.3. Stoichiometry of Leaf Litter
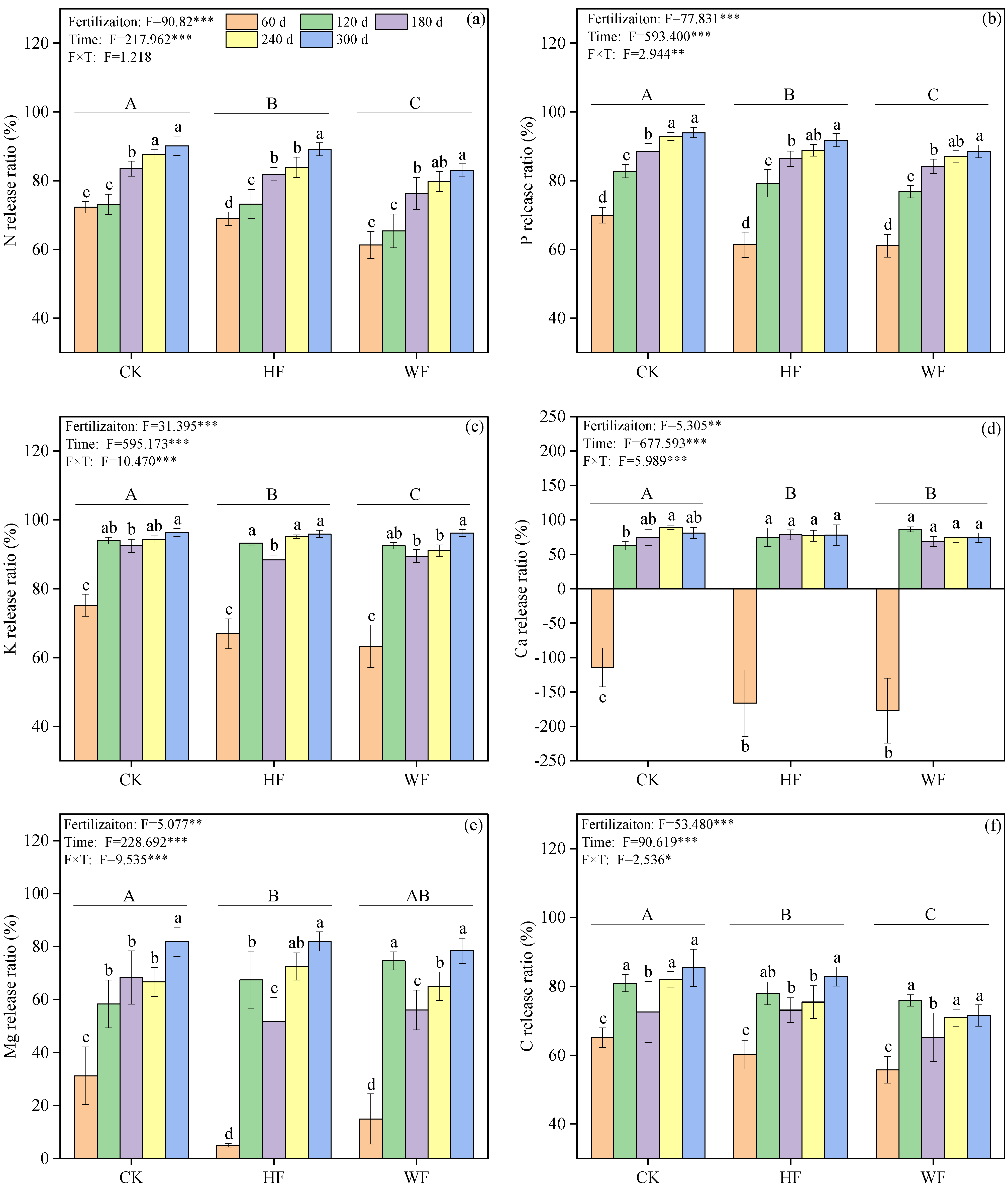
3.4. Elemental Release from Leaf Litter
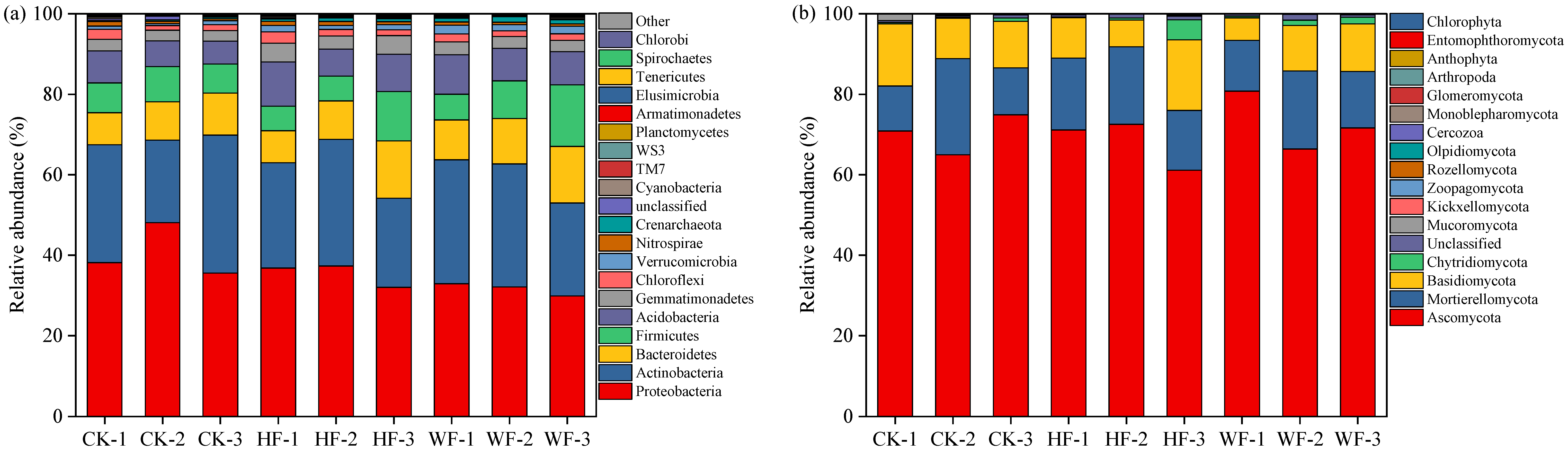
3.5. Microbial Composition
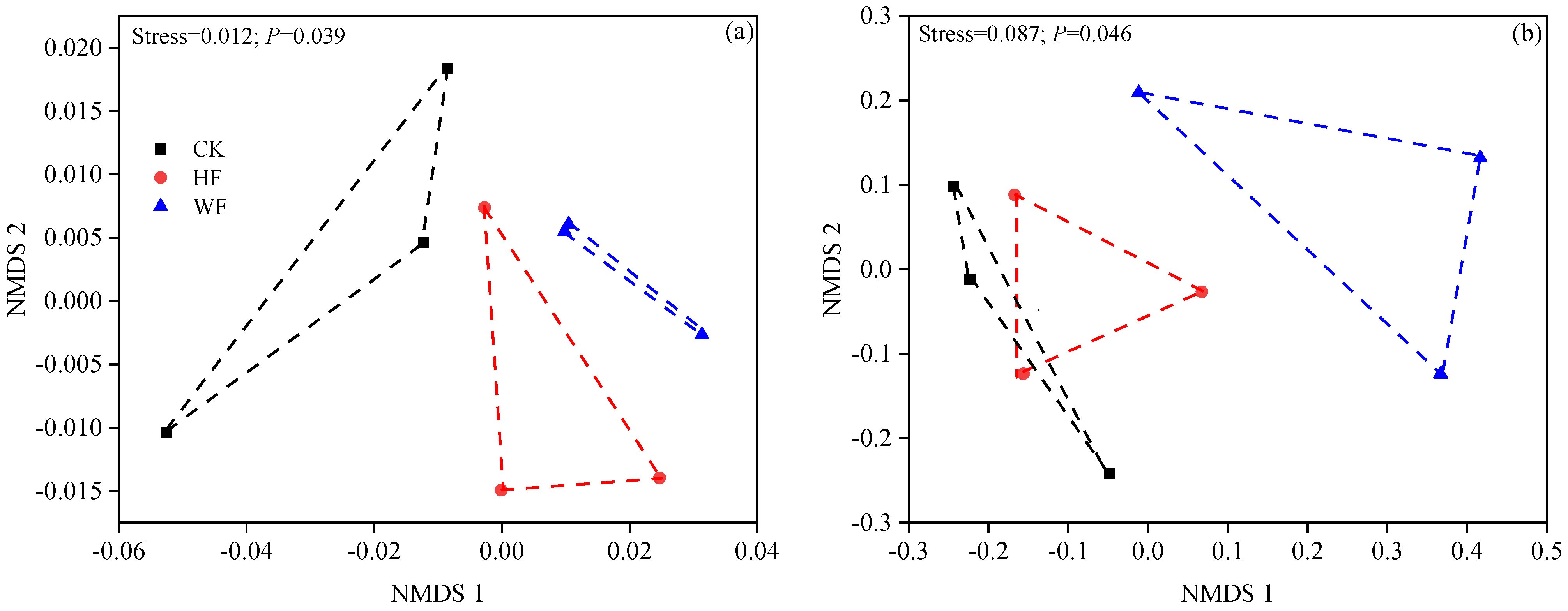
3.6. Microbial Diversity and Community
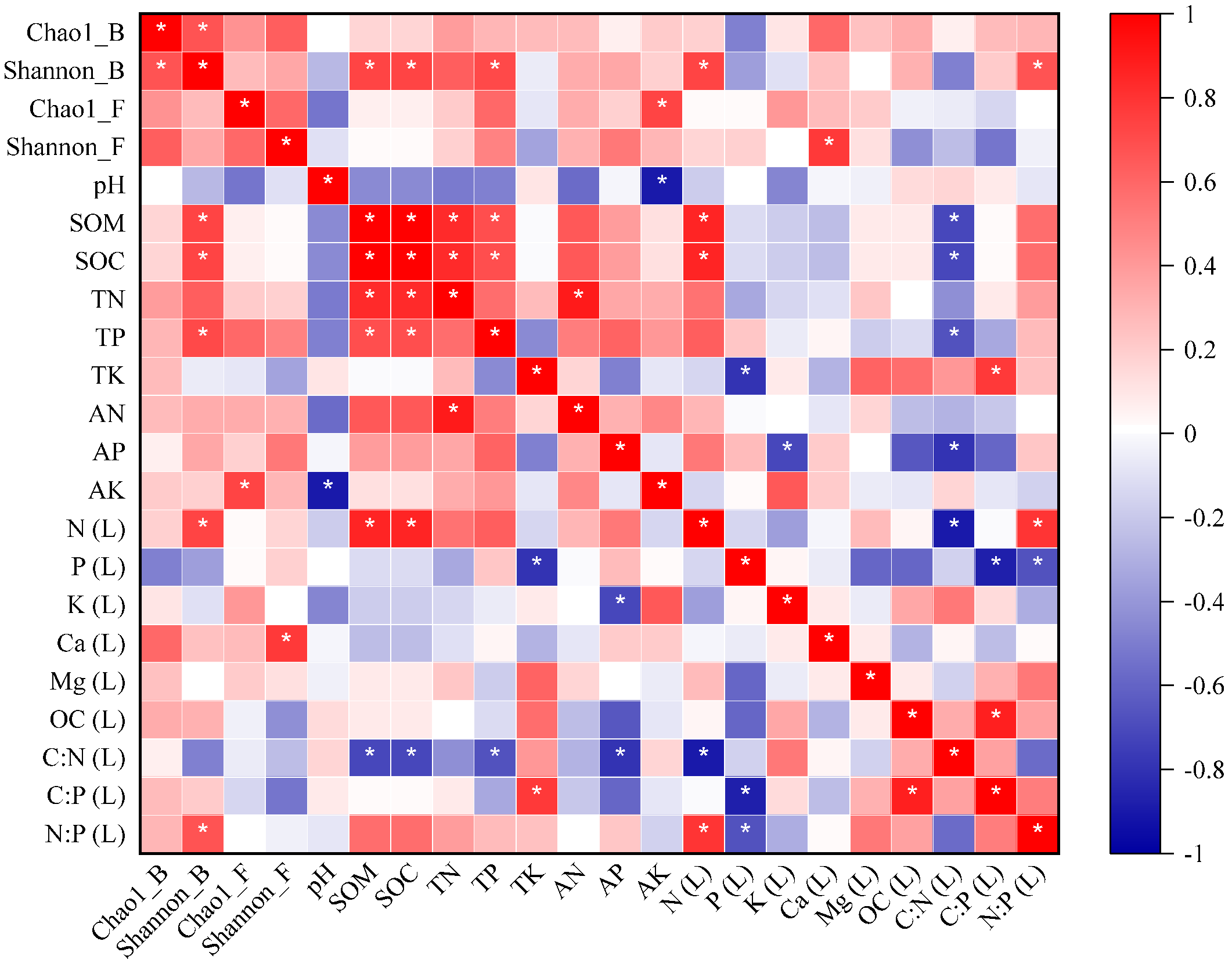
3.7. Relationships between Microbial Diversity and Community, Litter Nutrients, Decomposition Constant (k), and Soil Properties
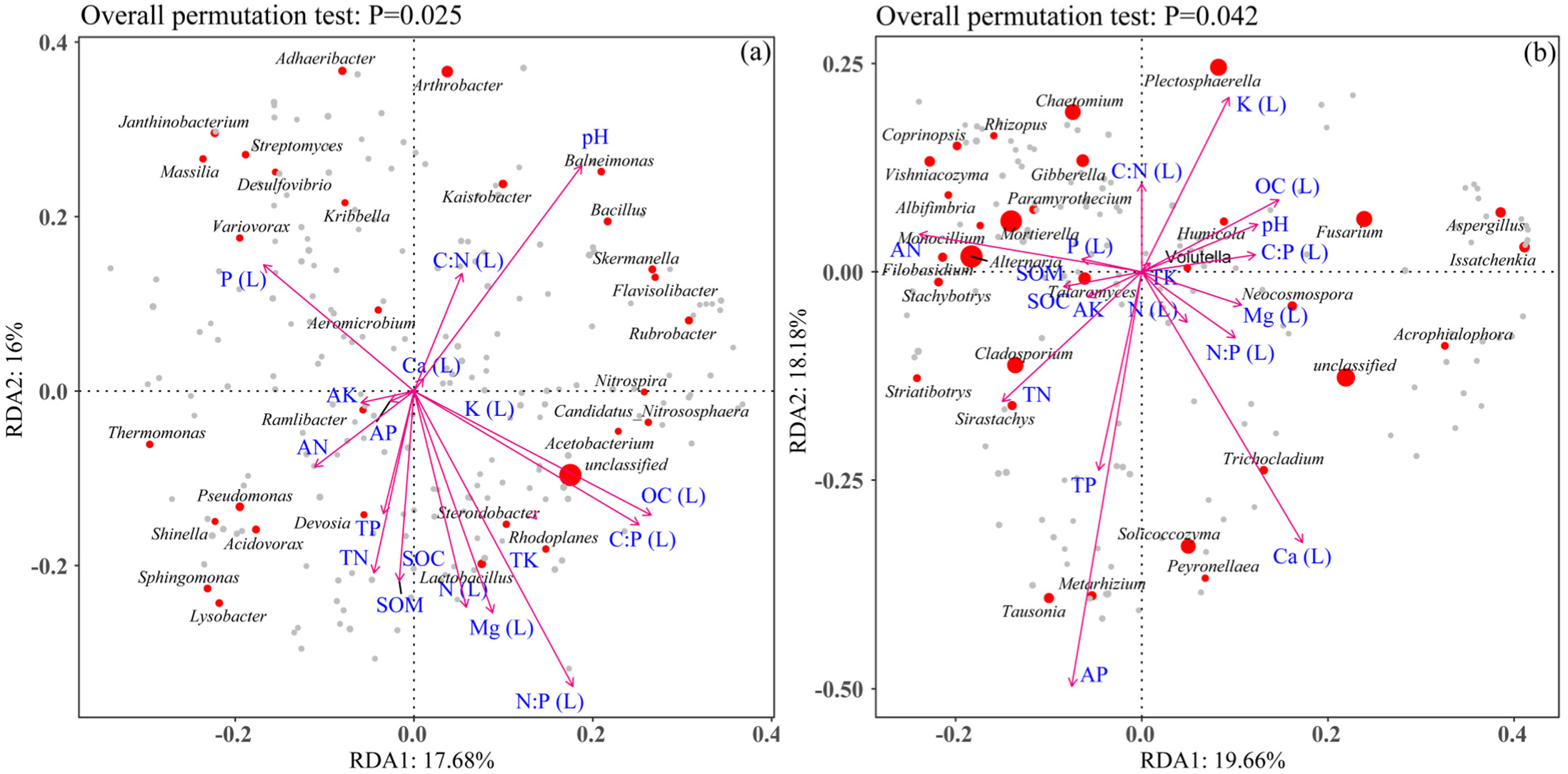
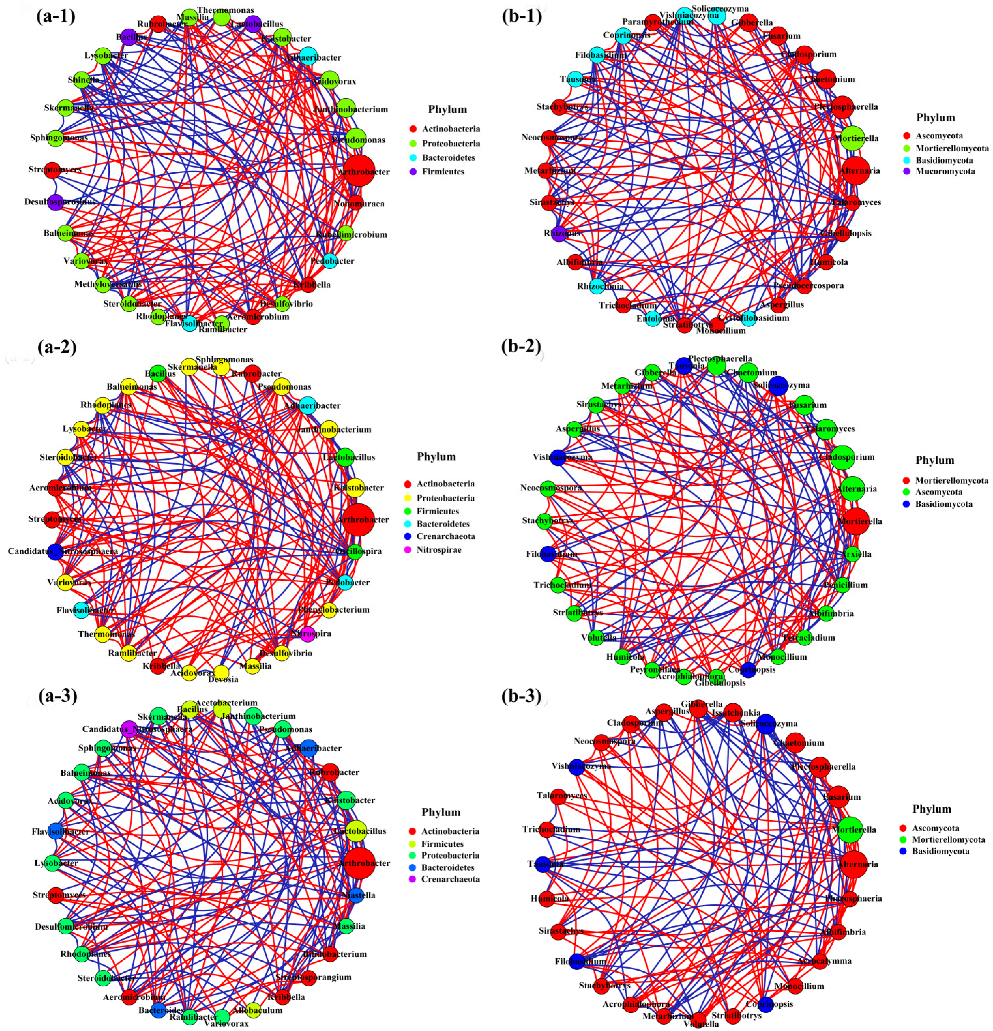
3.8. Microbial Network Analysis
3.9. Major Microbes Involved in the Breakdown of Leaf Litter
4. Discussion
4.1. Litter Biomass and C Content in Response to Fertilization
4.2. Litter Breakdown in Response to Fertilization
4.3. Microbial Community in Response to Fertilization
4.4. Key Microbes Involved in Litter Breakdown
4.5. Limitations and Prospects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, J. Review on the Influence of the Forest Litter Decomposition on Soil Microorganism. J. Anhui Agric. Sci. 2020, 48, 25–27, 31. [Google Scholar] [CrossRef]
- Portillo-Estrada, M.; Pihlatie, M.; Korhonen, J.F.J.; Levula, J.; Frumau, A.K.F.; Ibrom, A.; Lembrechts, J.J.; Morillas, L.; Horvath, L.; Jones, S.; et al. Climatic Controls on Leaf Litter Decomposition across European Forests and Grasslands Revealed by Reciprocal Litter Transplantation Experiments. Biogeosciences 2016, 13, 1621–1633. [Google Scholar] [CrossRef]
- Pinos, J.; Studholme, A.; Carabajo, A.; Gracia, C. Leaf Litterfall and Decomposition of Polylepis Reticulata in the Treeline of the Ecuadorian Andes. Mt. Res. Dev. 2017, 37, 87–96. [Google Scholar] [CrossRef]
- Wan, C.; Tao, C.; Yang, X.; Long, W.; Feng, D.; Zhou, W.; Yang, Q. Impact of Forest Community Species Composition on Litter Species Composition. Acta Ecol. Sin. 2015, 35, 7435–7443. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Higuchi, N.; Schimel, J.P.; Ferreira, L.V.; Melack, J.M. Decomposition and Carbon Cycling of Dead Trees in Tropical Forests of the Central Amazon. Oecologia 2000, 122, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Leroy, C.J.; Marks, J.C. Litter Quality, Stream Characteristics and Litter Diversity Influence Decomposition Rates and Macroinvertebrates. Freshw. Biol. 2006, 51, 605–617. [Google Scholar] [CrossRef]
- Bradford, M.A.; Warren, R.J.; Baldrian, P.; Crowther, T.W.; Maynard, D.S.; Oldfield, E.E.; Wieder, W.R.; Wood, S.A.; King, J.R. Climate Fails to Predict Wood Decomposition at Regional Scales. Nat. Clim. Chang. 2014, 4, 625–630. [Google Scholar] [CrossRef]
- Ulyshen, M.D. Wood Decomposition as Influenced by Invertebrates. Biol. Rev. Camb. Philos. Soc. 2016, 91, 70–85. [Google Scholar] [CrossRef]
- Huang, S.; Huang, Y.; Gao, W.; Nie, S.; Cai, B.; Lin, J. Dynamics of Litterfall and Nutrient Return in Three Typical Forests of Wuyi Mountain along Altitudinal Gradient. J. Trop. Subtrop. Bot. 2020, 28, 394–402. [Google Scholar] [CrossRef]
- Strickland, M.S.; Osburn, E.; Lauber, C.; Fierer, N.; Bradford, M.A. Litter Quality is in the Eye of the Beholder: Initial Decomposition Rates as a Function of Inoculum Characteristics. Funct. Ecol. 2009, 23, 627–636. [Google Scholar] [CrossRef]
- Kubartová, A.; Ranger, J.; Berthelin, J.; Beguiristain, T. Diversity and Decomposing Ability of Saprophytic Fungi from Temperate Forest Litter. Microb. Ecol. 2009, 58, 98–107. [Google Scholar] [CrossRef]
- Bray, S.R.; Kitajima, K.; Mack, M.C. Temporal Dynamics of Microbial Communities on Decomposing Leaf Litter of 10 Plant Species in Relation to Decomposition Rate. Soil Biol. Biochem. 2012, 49, 30–37. [Google Scholar] [CrossRef]
- Voriskova, J.; Baldrian, P. Fungal Community on Decomposing Leaf Litter Undergoes Rapid Successional Changes. ISME J. 2013, 7, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Purahong, W.; Kapturska, D.; Pecyna, M.J.; Jariyavidyanont, K.; Kaunzner, J.; Juncheed, K.; Uengwetwanit, T.; Rudloff, R.; Schulz, E.; Hofrichter, M.; et al. Effects of Forest Management Practices in Temperate Beech Forests on Bacterial and Fungal Communities Involved in Leaf Litter Degradation. Microb. Ecol. 2015, 69, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Gao, P.; Li, T.; Zhang, J.; Dong, J.; Xu, J.; Tun, X. Effects of Soil Microbial Community on the Litter Decomposition in Mixed Quercus acutissima Carruth. and Robinia pseudoacacia L. Forest. Acta Ecol. Sin. 2021, 41, 2315–2325. [Google Scholar] [CrossRef]
- Gao, J. Characteristics of Leaf Litter Decomposition and the Influencing Factors of Forest Ecosystem in Liaoheyuan; Beijing Forestry University: Beijing, China, 2016. [Google Scholar]
- Dong, X.; Gao, P.; Zhou, R.; Li, C.; Dun, X.; Niu, X. Changing Characteristics and Influencing Factors of the Soil Microbial Community during Litter Decomposition in a Mixed Quercus acutissima Carruth. and Robinia pseudoacacia L. Forest in Northern China. Catena 2021, 196, 104811. [Google Scholar] [CrossRef]
- Yang, G.; Zhang, Z.; Zhao, Y.; Shi, Y.; Hu, R. Litter Decomposition and Its Effects on Soil Microbial Community in Shapotou Area, China. Chin. J. Appl. Ecol. 2022, 33, 1810–1818. [Google Scholar] [CrossRef]
- Zeng, J.; Liu, X.; Song, L.; Lin, X.; Zhang, H.; Shen, C.; Chu, H. Nitrogen Fertilization Directly Affects Soil Bacterial Diversity and Indirectly Affects Bacterial Community Composition. Soil Biol. Biochem. 2016, 92, 41–49. [Google Scholar] [CrossRef]
- Liu, W.; Shen, F.; Xu, Z.; Wu, J.; Duan, H.; Ge, Y.; Fan, H. Impacts of Nitrogen Deposition on Nutrient Release during Leaf Litter Decomposition in Cunninghamia lanceolata Plantations. Ecol. Environ. Sci. 2019, 28, 695–701. [Google Scholar] [CrossRef]
- Xie, J.; Fang, H.; Zhang, Q.; Chen, M.; Xu, X.; Pan, J.; Gao, Y.; Fang, X.; Guo, X.; Zhang, L. Understory Plant Functional Types Alter Stoichiometry Correlations between Litter and Soil in Chinese Fir Plantations with N and P Addition. Forests 2019, 10, 742. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.; Xu, X.; Lei, J.; Sun, J. Effects of Fertilizer Addition on Surface Litter Decomposition in the Tarim Desert Highway Shelterbelt. Acta Ecol. Sin. 2017, 37, 1506–1514. [Google Scholar] [CrossRef]
- Zheng, X.; She, H.; Xue, L.; Cai, J. Effects of N and P Additions on Decomposition of Needle Litter and Soil Characteristics in Pinus massoniana Woodland. Ecol. Environ. Sci. 2017, 26, 1710–1718. [Google Scholar] [CrossRef]
- Zhang, X. The Effects of Nitrogen and Phosphorus Additions on the Quality of Chinese Fir Leaf Litter and Its Decomposition Processes; Jiangxi Agricultural University: Nanchang, China, 2019. [Google Scholar] [CrossRef]
- Chen, S.; Cai, J.; Lin, C.; Song, H.; Yang, Y. Response of Leaf Litter Decomposition of Different Tree Species to Nitrogen Addition in a Subtropical Forest. Chin. J. Plant Ecol. 2020, 44, 214–227. [Google Scholar] [CrossRef]
- Zhang, Y.; Jin, Y.; Xu, J.; He, H.; Tao, Y.; Yang, Z.; Bai, Y. Effects of Exogenous N and Endogenous Nutrients on Alpine tundra Litter Decomposition in an Area of High Nitrogen Deposition. Sci. Total Environ. 2022, 805, 150388. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Wang, G.G.; Fang, X.; Wan, S.; Zhang, Y.; Liang, C. Nitrogen Deposition Effect on Forest Litter Decomposition is Interactively Regulated by Endogenous Litter Quality and Exogenous Resource Supply. Plant Soil 2019, 437, 413–426. [Google Scholar] [CrossRef]
- Britton, A.J.; Mitchell, R.J.; Fisher, J.M.; Riach, D.J.; Taylor, A.F.S. Nitrogen Deposition Drives Loss of Moss Cover in Alpine Moss-Sedge Heath via Lowered C:N Ratio and Accelerated Decomposition. New Phytol. 2018, 218, 470–478. [Google Scholar] [CrossRef]
- Hou, S.; Hättenschwiler, S.; Yang, J.; Sistla, S.; Wei, H.; Zhang, Z.; Hu, Y.; Wang, R.; Cui, S.; Lv, X.; et al. Increasing Rates of Long-Term Nitrogen Deposition Consistently Increased Litter Decomposition in a Semi-arid Grassland. New Phytol. 2021, 229, 296–307. [Google Scholar] [CrossRef]
- Neff, J.C.; Townsend, A.R.; Gleixner, G.; Lehman, S.J.; Turnbull, J.; Bowman, W.D. Variable Effects of Nitrogen Additions on the Stability and Turnover of Soil Carbon. Nature 2002, 419, 915–917. [Google Scholar] [CrossRef]
- Song, C.; Liu, D.; Yang, G.; Song, Y.; Mao, R. Effect of Nitrogen Addition on Decomposition of Calamagrostis angustifolia Litters from Freshwater Marshes of Northeast China. Ecol. Eng. 2011, 37, 1578–1582. [Google Scholar] [CrossRef]
- Micks, P.; Downs, M.; Magill, A.; Nadelhoffer, K.; Aber, J. Decomposing Litter as a Sink for N-15-Enriched Additions to an Oak Forest and a Red Pine Plantation. For. Ecol. Manag. 2004, 196, 71–87. [Google Scholar] [CrossRef]
- Hobbie, S. Contrasting Effects of Substrate and Fertilizer Nitrogen on the Early Stages of Litter Decomposition. Ecosystems 2005, 8, 644–656. [Google Scholar] [CrossRef]
- Ma, F.; Liu, Y.; Cui, J.; Fan, H.; Lu, Y.; Li, M. Review on the Research Progress of Water and Fertilizer Integration. Xinjiang Agric. Sci. 2019, 56, 183–192. [Google Scholar] [CrossRef]
- Yan, X.; Dai, T.; Zhao, D.; Jia, L. Combined Surface Drip Irrigation and Fertigation Significantly Increase Biomass and Carbon Storage in a Populus × euramericana cv. Guariento Plantation. J. For. Res. 2016, 21, 280–290. [Google Scholar] [CrossRef]
- Bhat, R.; Sujatha, S.; Balasimha, D. Impact of Drip Fertigation on Productivity of Arecanut (Areca catechu L.). Arg. Water Manag. 2007, 90, 101–111. [Google Scholar] [CrossRef]
- Lu, R. Agricultural and Chemistry Analysis of Soil; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Wang, W.; Zhai, S.; Xia, Y.; Wang, H.; Ruan, D.; Zhou, T.; Zhu, Y.; Zhang, H.; Zhang, M.; Ye, H.; et al. Ochratoxin a Induces Liver Inflammation: Involvement of Intestinal Microbiota. Microbiome 2019, 7, 151. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Kang, F.; Li, T.; Song, X.; Zhao, W.; Yu, X.; Han, H. Assessing the Effect of Leaf Litter Diversity on the Decomposition and Associated Diversity of Fungal Assemblages. Forests 2015, 6, 2371–2386. [Google Scholar] [CrossRef]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Li, Y.; Bezemer, T.M.; Yang, J.; Lu, X.; Li, X.; Liang, W.; Han, X.; Li, Q. Changes in Litter Quality Induced by N Deposition Alter Soil Microbial Communities. Soil Biol. Biochem. 2018, 130, 33–42. [Google Scholar] [CrossRef]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A Tool for Visualizing High-Throughput Microbial Community Data. GigaScience 2013, 2, 16. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing Taxonomic Classification of Marker-Gene Amplicon Sequences with QIIME 2′s Q2-Feature-Classifier Plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An Open Annotation Tool for Parsing Fungal Community Datasets by Ecological Guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Gan, D.; Zeng, H.; Zhu, B. The Rhizosphere Effect on Soil Gross Nitrogen Mineralization: A Meta-Analysis. Soil Ecol. Lett. 2021, 4, 144–154. [Google Scholar] [CrossRef]
- Zhao, X.; Guan, M.; Sun, M.; Wang, Z.; Xu, X. Effects of Nitrogen and Phosphorus Additions on Litterfall Production and Nutrient Dynamics in Evergreen Broad-Leaved Forests. J. Nanjing For. Univ. 2020, 44, 55–62. [Google Scholar] [CrossRef]
- Apolinário, V.X.O.; Dubeux, J.C.B., Jr.; Mello, A.C.L.; Vendramini, J.M.B.; Lira, M.A.; Santos, M.V.F.; Muir, J.P. Deposition and Decomposition of Signal Grass Pasture Litter under Varying Nitrogen Fertilizer and Stocking Rates. Agron. J. 2013, 105, 999–1004. [Google Scholar] [CrossRef]
- Longhini, V.Z.; Cardoso, A.S.; Berca, A.S.; Carvalho, I.N.O.; Boddey, R.M.; Reis, R.A.; Dubeux, J.C.B., Jr.; Ruggieri, A.C. Nitrogen Fertilizer Increased Litter Deposition and Litter N in Warm-Climate Grasslands. Nutr. Cycl. Agroecosyst. 2021, 119, 247–258. [Google Scholar] [CrossRef]
- Zhang, H.; Yuan, W.; Dong, W.; Liu, S. Seasonal Patterns of Litterfall in Forest Ecosystem Worldwide. Ecol. Complex. 2014, 20, 240–247. [Google Scholar] [CrossRef]
- Andivia, E.; Bou, J.; Fernandez, M.; Caritat, A.; Alejano, R.; Vilar, L.; Vazquez-Pique, J. Assessing the Relative Role of Climate on Litterfall in Mediterranean Cork Oak Forests. Iforest 2018, 11, 786–793. [Google Scholar] [CrossRef]
- Wang, C.G.; Zheng, X.B.; Wang, A.Z.; Dai, G.H.; Zhu, B.K.; Zhao, Y.M.; Dong, S.J.; Zu, W.Z.; Wang, W.; Zheng, Y.G.; et al. Temperature and Precipitation Diversely Control Seasonal and Annual Dynamics of Litterfall in a Temperate Mixed Mature Forest, Revealed by Long-Term Data Analysis. JGR Biogeosci. 2021, 126, e2020JG006204. [Google Scholar] [CrossRef]
- Xiao, W.; Wang, K.; Song, Y.; An, W.; Zheng, X.; Pan, Y. Responses of Leaf Litter and Twig Decomposition Characteristics in Pinus armandii Forest to Nitrogen in Central Yunnan Plateau. J. Northwest A F Univ. (Nat. Sci. Ed.) 2023, 4, 2–12. [Google Scholar] [CrossRef]
- Ong, C.S.P.; Juan, J.C.; Yule, C.M. The Contribution of Leaching to Nutrient Release from Leaf Litter of Two Emergent Tree Species in a Malaysian Tropical Peat Swamp Forest. Hydrobiologia 2017, 794, 125–137. [Google Scholar] [CrossRef]
- Zhou, W.; Guo, Y.; Zhu, B.; Wang, X.; Zhou, L.; Yu, D.; Dai, L. Seasonal Variations of Nitrogen Flux and Composition in a Wet Deposition Forest Ecosystem on Changbai Mountain. Acta Ecol. Sin. 2015, 35, 158–164. [Google Scholar] [CrossRef]
- Spohn, M.; Pötsch, E.M.; Eichorst, S.A.; Woebken, D.; Wanek, W.; Richter, A. Soil Microbial Carbon Use Efficiency and Biomass Turnover in a Long-Term Fertilization Experiment in a Temperate Grassland. Soil Biol. Biochem. 2016, 97, 168–175. [Google Scholar] [CrossRef]
- Widdig, M.; Schleuss, P.; Biederman, L.A.; Borer, E.T.; Crawley, M.J.; Kirkman, K.P.; Seabloom, E.W.; Wragg, P.D.; Spohn, M. Microbial Carbon Use Efficiency in Grassland Soils Subjected to Nitrogen and Phosphorus Additions. Soil Biol. Biochem. 2020, 146, 107815. [Google Scholar] [CrossRef]
- Gill, A.L.; Schilling, J.; Hobbie, S.E. Experimental Nitrogen Fertilisation Globally Accelerates, then Slows Decomposition of Leaf Litter. Ecol. Lett. 2021, 24, 802–811. [Google Scholar] [CrossRef]
- Li, J.; Sang, C.; Yang, J.; Qu, L.; Xia, Z.; Sun, H.; Jiang, P.; Wang, X.; He, H.; Wang, C. Stoichiometric Imbalance and Microbial Community Regulate Microbial Elements Use Efficiencies under Nitrogen Addition. Soil Biol. Biochem. 2021, 156, 108207. [Google Scholar] [CrossRef]
- Salinas, N.; Malhi, Y.; Meir, P.; Silman, M.; Roman-Cuesta, R.; Huaman, J.; Salinas, D.; Huaman, V.; Gibaja, A.; Mamani, M.; et al. The Sensitivity of Tropical Leaf Litter Decomposition to Temperature: Results from a Large-Scale Leaf Translocation Experiment along an Elevation Gradient in Peruvian Forests. New Phytol. 2011, 189, 967–977. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Z. Responses of Litter Decomposition and Nutrient Release of Bothriochloa ischaemum to Soil Petroleum Contamination and Nitrogen Fertilization. Int. J. Environ. Sci. Technol. 2019, 16, 719–728. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, J.; Wang, L.; Mi, H.; Hui, H.; Li, L. Response Differences in Decomposition and Nutrient Release of Litter from Robinia pseudoacacia Plantations with Different Stand Ages to Nitrogen Deposition. J. Plant Resour. Environ. 2021, 30, 10–18. [Google Scholar] [CrossRef]
- Wardle, D.A. Communities and Ecosystems: Linking the Aboveground and Belowground Components; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Anderson, T.H.; Joergensen, R.G. Relationship between SIR and FE Estimates of Microbial Biomass C in Deciduous Forest Soils at Different pH. Soil Biol. Biochem. 1997, 29, 1033–1042. [Google Scholar] [CrossRef]
- Baath, E.; Anderson, T.H. Comparison of Soil Fungal/Bacterial Ratios in a pH Gradient Using Physiological and PLFA-based Techniques. Soil Biol. Biochem. 2003, 35, 955–963. [Google Scholar] [CrossRef]
- Fisk, M.C.; Fahey, T.J. Microbial Biomass and Nitrogen Cycling Responses to Fertilization and Litter Removal in Young Northern Hardwood Forests. Biogeochemistry 2001, 53, 201–223. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; McNulty, S.; Fernandez, I.J.; Boggs, J.; Schlesinger, W.H. Nitrogen Fertilization Decreases Forest Soil Fungal and Bacterial Biomass in Three Long-term Experiments. For. Ecol. Manag. 2006, 222, 459–468. [Google Scholar] [CrossRef]
- Schlesinger, W.H. Biogeochemistry: An Analysis of Global Change; Academic Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Fenn, M.E.; Poth, M.A.; Aber, J.D.; Baron, J.S.; Bormann, B.T.; Johnson, D.W.; Lemly, A.D.; McNulty, S.G.; Ryan, D.F.; Stottlemyer, R. Nitrogen Excess in North American Ecosystems: Predisposing Factors, Ecosystem Responses, and Management Strategies. Ecol. Appl. 1998, 8, 706–733. [Google Scholar] [CrossRef]
- Ghosh, A.; Bhattacharyya, R.; Dwivedi, B.S.; Meena, M.C.; Agarwal, B.K.; Mahapatra, P.; Shahi, D.K.; Salwani, R.; Agnihorti, R. Temperature Sensitivity of Soil Organic Carbon Decomposition as Affected by Long-Term Fertilization under a Soybean Based Cropping System in a Sub-tropical Alfisol. Agr. Ecosyst. Environ. 2016, 233, 202–213. [Google Scholar] [CrossRef]
- Dincă, L.C.; Grenni, P.; Onet, C.; Onet, A. Fertilization and Soil Microbial Community: A Review. Appl. Sci. 2022, 12, 1198. [Google Scholar] [CrossRef]
- Gao, R.; Duan, Y.; Zhang, J.; Ren, Y.; Liang, J.; Jing, Y.; Zhao, P. Effects of Long-Term Fertilization on Soil Microbial Diversity and Community Structure in the Agro-pastoral Ecotone. Environ. Sci. 2022, 44, 1063–1073. [Google Scholar] [CrossRef]
- Guan, Z.; Lu, Q.; Lin, Y.; Lin, D.; Lu, Y.; Han, Q.; Li, N.; Ma, W.; Wang, J.; Su, Y.; et al. Spatial Variations and Pools of Non-structural Carbohydrates in Young Catalpa bungei Undergoing Different Fertilization Regimes. Front. Plant Sci. 2022, 13, 1010178. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, J.; Yu, Z.; Yao, Q.; Li, Y.; Liang, A.; Zhang, W.; Mi, G.; Jin, J.; Liu, X.; et al. Long-Term Continuous Cropping of Soybean is Comparable to Crop Rotation in Mediating Microbial Abundance, Diversity and Community Composition. Soil Tillage Res. 2020, 197, 104503. [Google Scholar] [CrossRef]
- Andlar, M.; Rezic, T.; Mardetko, N.; Kracher, D.; Ludwig, R.; Santek, B. Lignocellulose Degradation: An Overview of Fungi and Fungal Enzymes Involved in Lignocellulose Degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef]
- Tsegaye, B.; Balomajumder, C.; Roy, P. Microbial Delignification and Hydrolysis of Lignocellulosic Biomass to Enhance Biofuel Production: An Overview and Future Prospect. Bull. Natl. Res. Cent. 2019, 43, 51. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Yanga, Y. Phylogenetic Molecular Ecological Network of Soil Microbial Communities in Response to Elevated CO2. mBio 2011, 2, e00122-11. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kou, D.; Li, F.; Ding, J.; Yang, G.; Fang, K.; Yang, Y. Linkage of Plant and Abiotic Properties to the Abundance and Activity of N-cycling Microbial Communities in Tibetan Permafrost-affected Regions. Plant Soil 2019, 434, 453–466. [Google Scholar] [CrossRef]
- Gao, C.; Shi, N.; Chen, L.; Ji, N.; Wu, B.; Wang, Y.; Xu, Y.; Zheng, Y.; Mi, X.; Ma, K.; et al. Relationships between Soil Fungal and Woody Plant Assemblages Differ between Ridge and Valley Habitats in a Subtropical Mountain Forest. New Phytol. 2016, 213, 1874–1885. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gao, S.; Chen, Y.; He, P.; He, Q.; Qiu, Q.; Li, J. Effects of Short-Term Fertilization on Soil Fungal Community Structure and Functional Group in Eucalyptus Artificial Forest. Acta Ecol. Sin. 2020, 40, 3813–3821. [Google Scholar] [CrossRef]
- Guan, Z.; Lin, D.; Chen, D.; Guo, Y.; Lu, Y.; Han, Q.; Li, N.; Su, Y.; Li, J.; Wang, J.; et al. Soil Microbial Communities Response to Different Fertilization Regimes in Young Catalpa bungei Plantation. Front. Microbiol. 2022, 13, 948875. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fertilization | Litter Number | a | k | 95% Confidence Interval | Equation | R2 | t50% (a) | t95% (a) | |
|---|---|---|---|---|---|---|---|---|---|
| Lower Bound | Upper Bound | ||||||||
| CK | CK-1 | 94.014 | 2.862 | 2.363 | 3.362 | Y = 94.014 × EXP(−2.862 × t) | 0.935 | 0.242 | 1.047 |
| CK-2 | 95.058 | 2.984 | 2.545 | 3.422 | Y = 95.058 × EXP(−2.984 × t) | 0.955 | 0.232 | 1.004 | |
| CK-3 | 95.392 | 3.176 | 2.712 | 3.640 | Y = 95.392 × EXP(−3.176 × t) | 0.957 | 0.218 | 0.943 | |
| HF | HF-1 | 97.497 | 2.630 | 2.375 | 2.886 | Y = 97.497 × EXP(−2.630 × t) | 0.979 | 0.264 | 1.139 |
| HF-2 | 96.181 | 2.498 | 2.230 | 2.766 | Y = 96.181 × EXP(−2.498 × t) | 0.973 | 0.277 | 1.199 | |
| HF-3 | 95.935 | 2.500 | 2.222 | 2.777 | Y = 95.935 × EXP(−2.500 × t) | 0.971 | 0.277 | 1.198 | |
| WF | WF-1 | 95.526 | 2.061 | 1.810 | 2.312 | Y = 95.526 × EXP(−2.061 × t) | 0.961 | 0.336 | 1.454 |
| WF-2 | 96.688 | 2.096 | 1.895 | 2.297 | Y = 96.688 × EXP(−2.096 × t) | 0.976 | 0.331 | 1.429 | |
| WF-3 | 96.304 | 2.135 | 1.928 | 2.343 | Y = 96.304 × EXP(−2.135 × t) | 0.976 | 0.325 | 1.403 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, Z.; Chen, T.; Chen, D.; Lu, Y.; Han, Q.; Li, N.; Ma, W.; Wang, J.; Su, Y.; Li, J.; et al. Leaf Litter Breakdown and Soil Microbes in Catalpa bungei Plantations in Response to Various Fertilization Regimes. Forests 2023, 14, 699. https://doi.org/10.3390/f14040699
Guan Z, Chen T, Chen D, Lu Y, Han Q, Li N, Ma W, Wang J, Su Y, Li J, et al. Leaf Litter Breakdown and Soil Microbes in Catalpa bungei Plantations in Response to Various Fertilization Regimes. Forests. 2023; 14(4):699. https://doi.org/10.3390/f14040699
Chicago/Turabian StyleGuan, Zhuizhui, Tianxiao Chen, Dong Chen, Yizeng Lu, Qingjun Han, Ningning Li, Wenjun Ma, Junhui Wang, Yan Su, Jiyue Li, and et al. 2023. "Leaf Litter Breakdown and Soil Microbes in Catalpa bungei Plantations in Response to Various Fertilization Regimes" Forests 14, no. 4: 699. https://doi.org/10.3390/f14040699