
A Contribution to Knowledge of Russula Section Ingratae (Russulales, Russulaceae) in China: Two New Taxa and Amended Descriptions of One Known Species
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. Molecular Procedures
2.3. Dataset Assembly
2.4. Phylogenetic Analyses
3. Results
3.1. Molecular Data
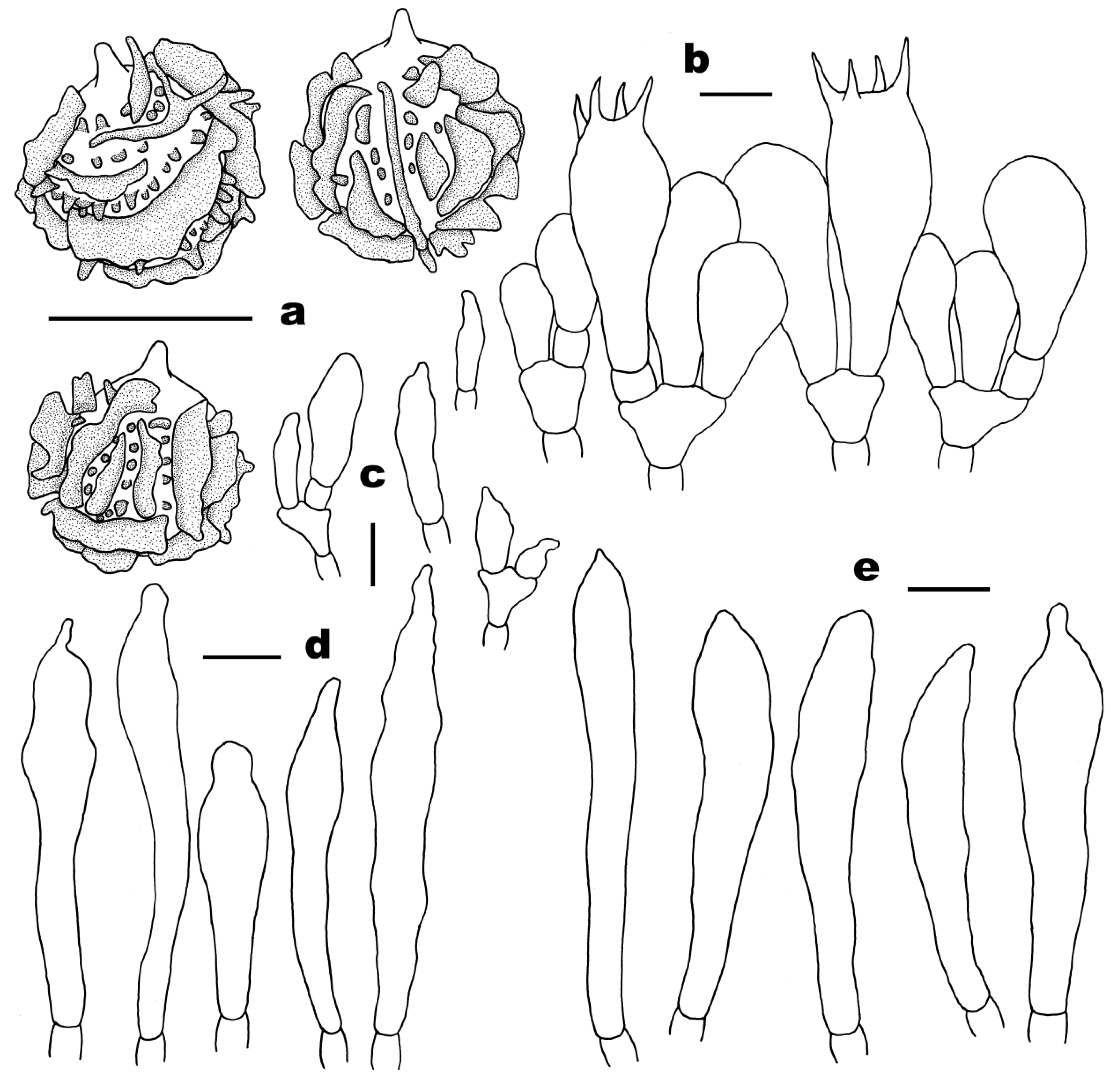
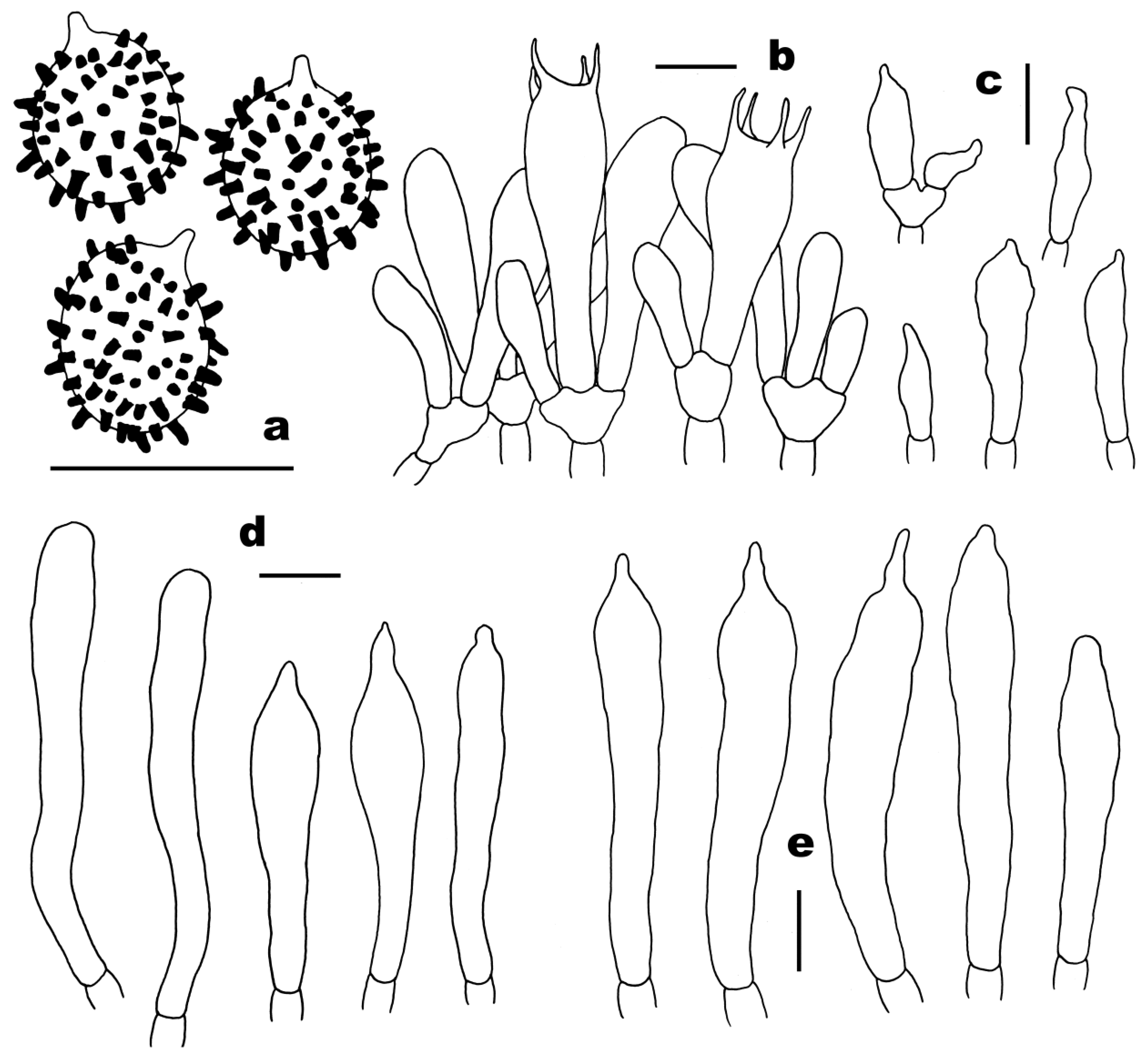
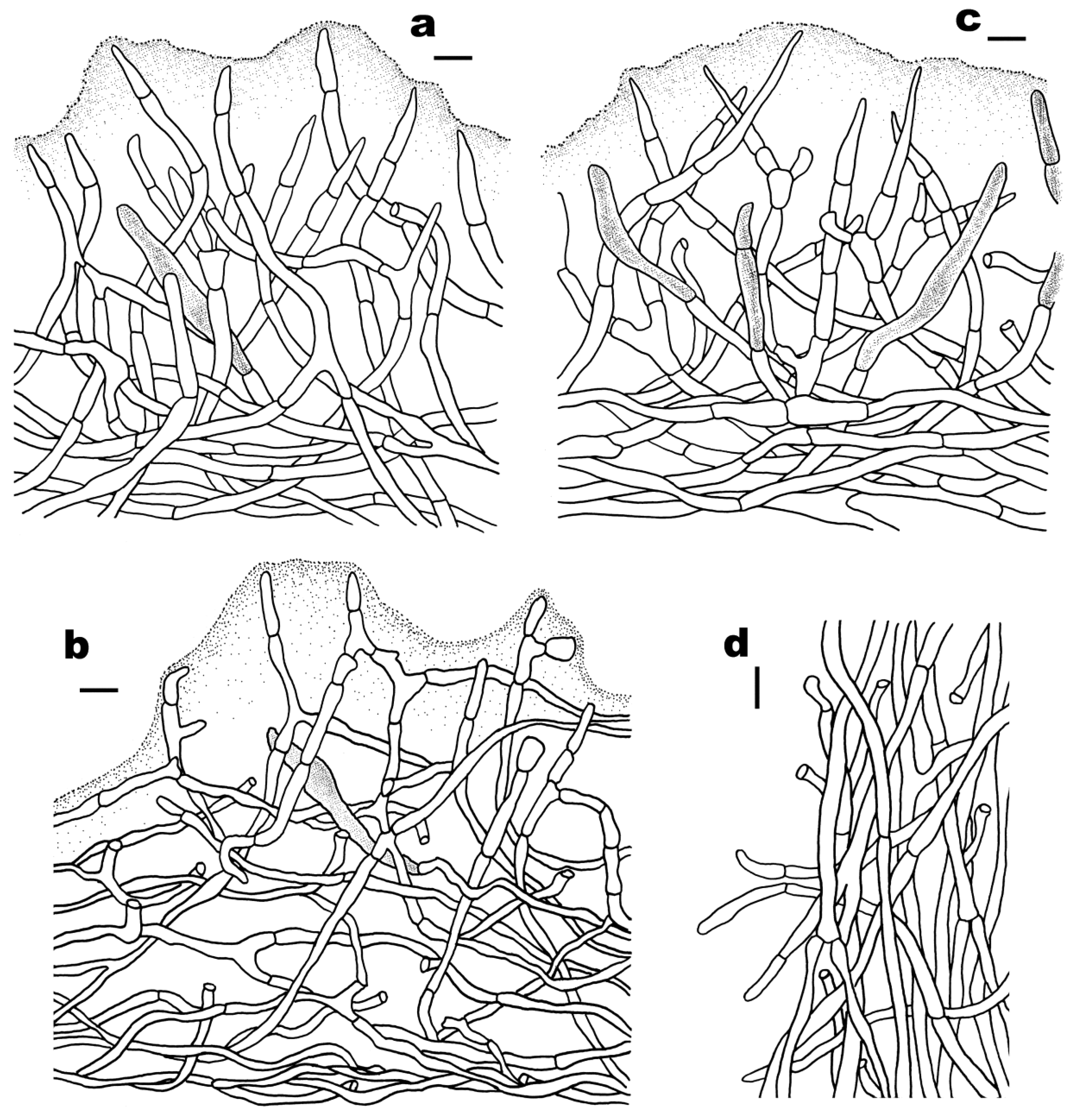
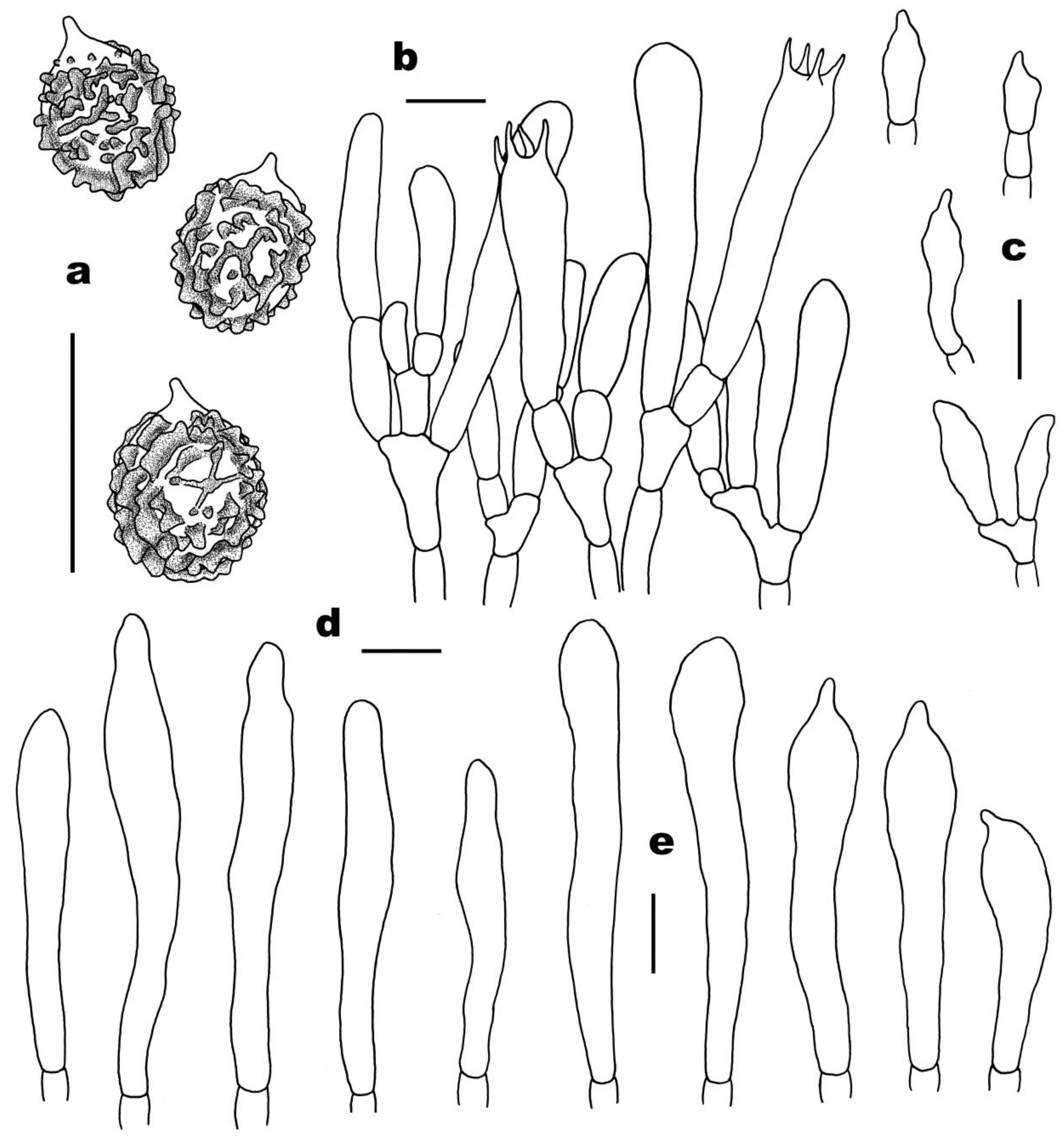
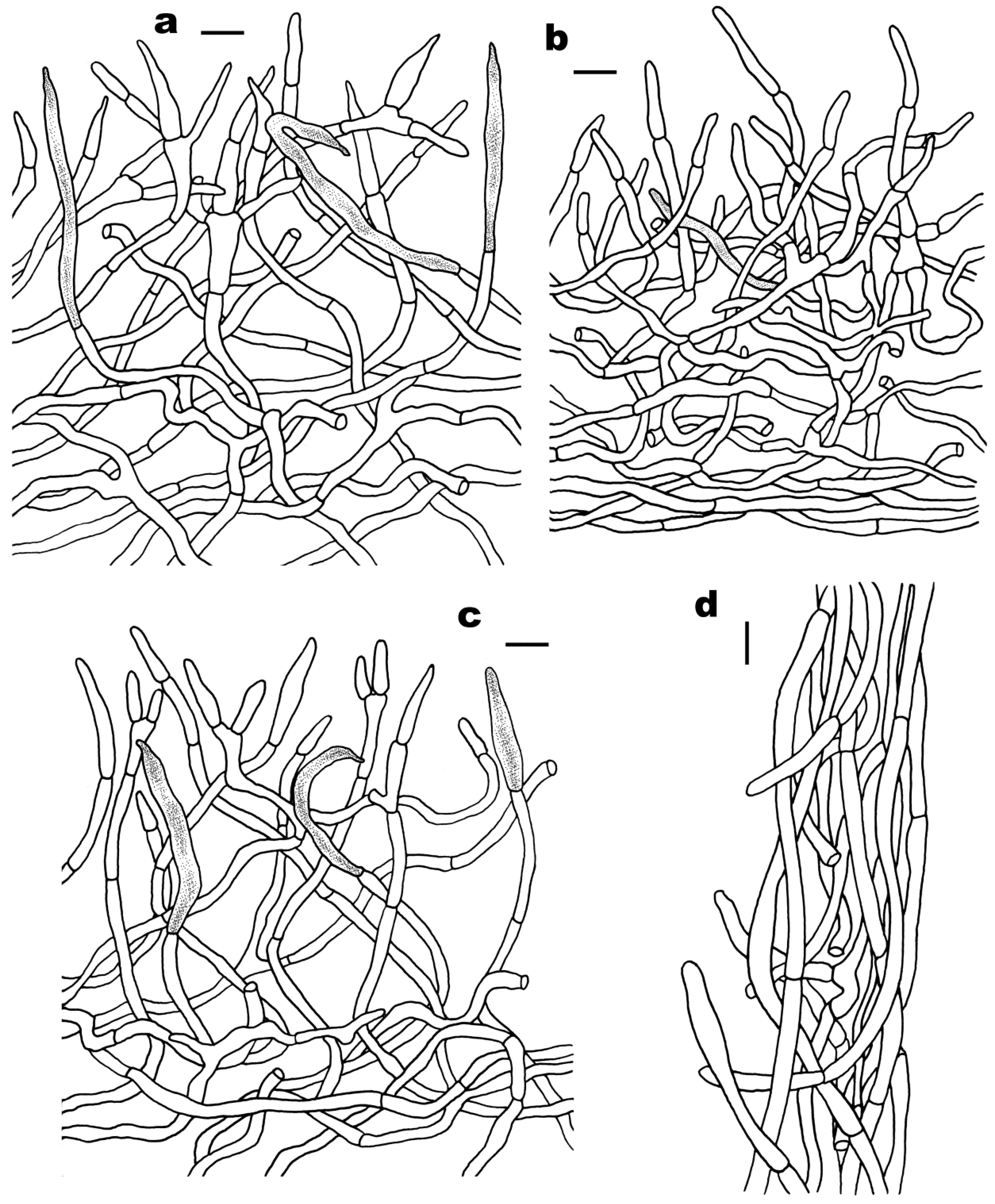
3.2. Taxonomy
4. Discussion
4.1. Species Diversity of Sect. Ingratae in China
4.2. Species Delimitation of Sect. Ingratae in China
4.3. Phylogenetic Relationships and Geographic Divergence of Chinese Sect. Ingratae
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buyck, B.; Thoen, D.; Watling, R. Ectomycorrhizal fungi of the Guinea-Congo region. Proceedings of the Royal Society of Edinburgh. Sect. B Biol. Sci. 1996, 104, 313–333. [Google Scholar] [CrossRef]
- Yamashita, S.; Hijii, N. The role of fungal taxa and developmental stage of mushrooms in determining the composition of the mycophagous insect community in a Japanese forest. Eur. J. Entomol. 2007, 104, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Looney, B.P.; Meidl, P.; Piatek, M.J.; Miettinen, O.; Martin, F.M.; Matheny, P.B.; Labbé, J.L. Russulaceae: A new genomic dataset to study ecosystem function and evolutionary diversifcation of ectomycorrhizal fungi with their tree associates. New Phytol. 2018, 218, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Buyck, B.; Zoller, S.; Hofstetter, V. Walking the thin line… ten years later: The dilemma of above- versus below-ground features to support phylogenies in the Russulaceae (Basidiomycota). Fungal Divers. 2018, 89, 267–292. [Google Scholar] [CrossRef]
- Buyck, B.; Wang, X.H.; Adamcíková, K.; Caboň, M.; Jančovičová, S.; Hofstetter, V.; Adamčík, S. One step closer to unravelling the origin of Russula: Subgenus Glutinosae subg. nov. Mycosphere 2020, 11, 285–305. [Google Scholar] [CrossRef]
- Shaffer, R.L. North American russulas of the subsection Foetentinae. Mycologia 1972, 64, 1008–1053. [Google Scholar] [CrossRef]
- Singer, R. The Agaricales in Modern Taxonomy, 4th ed.; Koeltz Scientific Books: Königstein, Germany, 1986; pp. 1–981. [Google Scholar]
- Sarnari, M. Monografia Illustrate de Genere Russula in Europa; Fondazione Centro Studi Micologici: Trento, Italy, 1998; pp. 1–800. [Google Scholar]
- Li, G.J.; Li, S.M.; Buyck, B.; Zhao, S.Y.; Xie, X.J.; Shi, L.Y.; Deng, C.Y.; Meng, Q.F.; Sun, Q.B.; Yan, J.Q.; et al. Three new Russula species in sect. Ingratae (Russulales, Basidiomycota) from southern China. MycoKeys 2021, 84, 103–139. [Google Scholar] [CrossRef]
- Bi, Z.S.; Li, T.H. A preliminary note on Russula species from Guangdong, with a new species and a new variety. Guihaia 1986, 6, 193–199. [Google Scholar]
- Li, G.J.; Zhao, D.; Li, S.F.; Wen, H.A. Russula chiui and R. pseudopectinatoides, two new species from southwestern China supported by morphological and molecular evidence. Mycol. Prog. 2015, 14, 1–14. [Google Scholar] [CrossRef]
- Song, Y.; Buyck, B.; Li, J.W.; Yuan, F.; Zhang, Z.W.; Qiu, L.H. Two novel and a forgotten Russula species in sect. Ingratae (Russulales) from Dinghushan Biosphere Reserve in southern China. Cryptogamie Mycol. 2018, 39, 341–357. [Google Scholar] [CrossRef]
- Song, J.; Chen, B.; Liang, J.F.; Li, H.J.; Wang, S.K.; Lu, J.K. Morphology and phylogeny reveal Russula subpunctipes sp. nov., from southern China. Phytotaxa 2020, 459, 16–24. [Google Scholar] [CrossRef]
- Yuan, F.; Song, Y.; Buyck, B.; Li, J.W.; Qiu, L.H. Russula viridicinnamomea F. Yuan & Y. Song, sp. nov. and R. pseudocatillus F. Yuan & Y. Song, sp. nov., two new species from southern China. Cryptogamie Mycol. 2019, 40, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Song, J.; Zhang, J.H.; Liang, J.F. Morphology and molecular phylogeny reveal two new species in Russula sect. Ingratae from China. Phytotaxa 2021, 525, 109–123. [Google Scholar] [CrossRef]
- Han, Y.X.; Liang, Z.Q.; Jiang, S.; Zeng, N.K. Russula hainanensis (Russulaceae, Russulales), a new species from tropical China. Phytotaxa 2022, 552, 035–050. [Google Scholar] [CrossRef]
- Mao, X.L. Poisonous mushrooms and their toxins in China. Mycosystema 2006, 25, 345–363. [Google Scholar]
- Chen, Z.H.; Zhang, P.; Zhang, Z.G. Investigation and analysis of 102 mushroom poisoning cases in Southern China from 1994 to 2012. Fungal Divers. 2013, 64, 123–131. [Google Scholar] [CrossRef]
- Tolgor, B.; Bao, H.Y.; Li, Y. A revised checklist of poisonous mushrooms in China. Mycosystema 2014, 33, 517–548. [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Taschenlexikon der Farben. 3. Aufl.; Muster-Schmidt Verlag: Göttingen, Germany, 1981; p. 242. [Google Scholar]
- Adamčík, S.; Looney, B.; Caboň, M.; Jančovičová, S.; Adamčíková, K.; Avis, P.G.; Barajas, M.; Bhatt, R.P.; Corrales, A.; Das, K.; et al. The quest for a globally comprehensible Russula language. Fungal Divers. 2019, 99, 369–449. [Google Scholar] [CrossRef]
- Han, Y.X.; Liang, Z.Q.; Zeng, N.K. Notes on four species of Russula subgenus Heterophyllidiae (Russulaceae, Russulales) from southern China. Front. Microbiol. 2023, 14, 1140127. [Google Scholar] [CrossRef]
- Adamčík, S.; Marhold, K. Taxonomy of the Russula xerampelina group. I. Morphometric study of the Russula xerampelina group in Slovakia. Mycotaxon 2000, 76, 463–480. [Google Scholar]
- Buyck, B. The study of microscopic features in Russula 2. Sterile cells of the hymenium. Russulales News 1991, 1, 62–85. [Google Scholar]
- Buyck, B. Valeur taxonomique du bleu de crésyl pour le genre Russula. Bull. Soc. Myc. Fr. 1989, 105, 1–6. [Google Scholar]
- Caboň, M.; Eberhardt, U.; Looney, B.; Hampe, F.; Kolařík, M.; Jančovičová, S.; Verbeken, A.; Adamčík, S. New insights in Russula subsect. Rubrinae: Phylogeny and the quest for synapomorphic characters. Mycol. Prog. 2017, 16, 877–892. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, T.Y.; Kauff, F.; Schoch, C.; Matheny, P.B.; Hofstetter, V.; Cox, C.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J.; et al. Reconstructing the early evolution of the fungi using a six gene phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenies. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analyses program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Liu, X.L.; Tolgor, B.; Wang, X.H. Species diversity of Russula from the Greater and Lesser Hinggan Mountains in Northeast China. Mycosystema 2017, 36, 1355–1368. [Google Scholar] [CrossRef]
- Jabeen, S.; Razaq, A.; Niazi, A.R.K.; Ahmad, I.; Grebenc, T.; Khalid, A.N. Russula ahmadii (Basidiomycota, Russulales), a new species in section Ingratae and its ectomycorrhiza from coniferous forests of Pakistan. Phytotaxa 2017, 321, 241–253. [Google Scholar] [CrossRef]
- Vidal, J.M.; Alvarado, P.; Loizides, M.; Konstantinidis, G.; Chachuła, P.; Mleczko, P.; Moreno, G.; Vizzini, A.; Krakhmalnyi, M.; Paz, A.; et al. A phylogenetic and taxonomic revision of sequestrate Russulaceae Mediterranean and temperate Europe. Persoonia 2019, 42, 127–185. [Google Scholar] [CrossRef] [Green Version]
- Eberhardt, U. Molecular kinship analyses of the agaricoid Russulaceae: Correspondence with mycorrhizal anatomy and sporocarp features in the genus Russula. Mycol. Prog. 2002, 1, 201–223. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.; Barber, P.A.; Alvarado, P.; Barnes, C.W.; Buchanan, P.K.; Heykoop, M.; Moreno, G.; et al. Fungal planet description sheets: 558–624. Persoonia 2017, 38, 240–384. [Google Scholar] [CrossRef]
- Wang, X.H.; Das, K.; Horman, J.; Antonin, V.; Baghela, A.; Chakraborty, D.; Hembrom, M.E.; Nakasone, K.; Ortiz-Santana, B.; Vizzini, A.; et al. Fungal biodiversity profiles 51–60. Cryptogam. Mycol. 2018, 39, 211–257. [Google Scholar] [CrossRef]
- Yuan, H.S.; Lu, X.; Dai, Y.C.; Hyde, K.D.; Kan, Y.H.; Kušan, I.; He, S.H.; Liu, N.G.; Sarma, V.V.; Zhao, C.L.; et al. Fungal diversity notes 1277–1386: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2020, 104, 1–266. [Google Scholar] [CrossRef]
- Lee, H.; Park, M.S.; Jung, P.E.; Eimes, J.A.; Seok, S.J.; Lim, Y.W. Re-evaluation of the taxonomy and diversity of Russula section Foetentinae (Russulales, Basidiomycota) in Korea. Mycoscience 2017, 58, 351–360. [Google Scholar] [CrossRef]
- Looney, B.P.; Ryberg, M.; Hampe, F.; Sánchez-García, M.; Matheny, P.B. Into and out of the tropics: Global diversification patterns in a hyperdiverse clade of ectomycorrhizal fungi. Mol. Ecol. 2016, 25, 630–647. [Google Scholar] [CrossRef]
- Buyck, B.; Eyssartier, G.; Armada, F.; Corrales, A.; Hembrom, M.E.; Rossi, W.; Bellanger, J.M.; Das, K.; Dima, B.; Ghosh, A.; et al. Fungal Biodiversity Profiles 111–120. Cryptogam. Mycol. 2022, 43, 23–61. [Google Scholar] [CrossRef]
- Razaq, A.; Ilyas, S.; Khalid, A.N.; Niazi, A.R. Russula foetentoides (Russulales, Russulaceae)—A new species from Pakistan. Sydowia 2014, 66, 289–298. [Google Scholar] [CrossRef]
- Melera, S.; Ostellari, C.; Roemer, N.; Avis, P.G.; Tonolla, M.; Barja, F.; Narduzzi-Wicht, B. Analysis of morphological, ecological and molecular characters of Russula pectinatoides Peck and Russula praetervisa Sarnari, with a description of the new taxon Russula recondita Melera & Ostellari. Mycol. Prog. 2017, 16, 117–134. [Google Scholar]
- Li, G.J. Taxonomy of Russula from China. Ph.D. Thesis, University of Chinese Academy of Sciences, Beijing, China, 2014. [Google Scholar]
- Buyck, B.; Hofstetter, V.; Eberhardt, U.; Verbeken, A.; Kauff, F. Walking the thin line between Russula and Lactarius: The dilemma of Russula subsect. Ochricompactae. Fungal Divers. 2008, 28, 15–40. [Google Scholar]
- Ghosh, A.; Das, K.; Bhatt, R.P.; Hembrom, M.E. Two new species of genus Russula from Western Himalaya with morphological details and phylogenetic estimations. Nova Hedwig. 2020, 111, 115–130. [Google Scholar] [CrossRef]
- Ghosh, A.; Buyck, B.; Das, K.; Chakraborty, D. Two new Asian species of Russula sect. Ingratae with unique basidiospore features for subg. Heterophyllidiae. Eur. J. Taxon. 2022, 847, 104–120. [Google Scholar] [CrossRef]
- Xie, X.D.; Liu, P.G.; Yu, F.Q. Species diversity of russuloid mycorrhizae forming fungi on Pinus yunnanensis seedlings and the mycorrhizal morphology. Acta Bot. Yunnanica 2010, 32, 211–220. [Google Scholar] [CrossRef]
- Wang, J.; Buyck, B.; Wang, X.H.; Tolgor, B. Visiting Russula (Russulaceae, Russulales) with samples from southwestern China finds one new subsection of R. subg. Heterophyllidia with two new species. Mycol. Prog. 2019, 18, 771–784. [Google Scholar] [CrossRef]
- Looney, B.P. Molecular annotation of type specimens of Russula species described by W.A. Murrill from the southeast United States. Mycotaxon 2014, 129, 255–268. [Google Scholar] [CrossRef]
- Haelewaters, D.; Dirks, A.C.; Kappler, L.A.; Mitchell, J.K.; Quijada, L.; Vandegrift, R.; Buyck, B.; Pfister, D.H. A preliminary checklist of fungi at the Boston Harbor Islands. Northeast. Nat. 2018, 25, 45–77. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Ghosh, A.; Chakraborty, D.; Li, J.W.; Qiu, L.H.; Baghela, A.; Halama, M.; Hembrom, M.E.; Mehmood, T.; Parihar, A.; et al. Fungal biodiversity profiles 31–40. Cryptogamie Mycol. 2017, 38, 353–406. [Google Scholar] [CrossRef]
- Lebel, T.; Tonkin, J.E. Australasian species of Macowanites are sequestrate species of Russula (Russulaceae, Basidiomycota). Aust. Syst. Bot. 2007, 20, 355–381. [Google Scholar] [CrossRef]
- Miller, S.L.; Buyck, B. Molecular phylogeny of the genus Russula in Europe with a comparison of modern infrageneric classifications. Mycol. Res. 2002, 106, 259–276. [Google Scholar] [CrossRef]
- Khatua, S.; Dutta, A.K.; Acharya, K. Prospecting Russula senecis: A delicacy among the tribes of West Bengal. PeerJ 2015, 3, e810. [Google Scholar] [CrossRef] [Green Version]
- Yomyart, S.; Piapukiew, J.; Watling, R.; Whalley, A.J.S.; Sihanonth, P. Russula siamensis: A new species of annulate Russula from Thailand. Mycotaxon 2006, 95, 247–254. [Google Scholar]
- Buyck, B.; Duhem, B.; Das, K.; Jayawardena, R.S.; Niveiro, N.; Pereira, O.L.; Prasher, I.B.; Adhikari, S.; Albertó, E.O.; Bulgakov, T.S.; et al. Fungal biodiversity profiles 21–30. Cryptogam. Mycol. 2017, 38, 101–146. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.A.; Dunn, C.W. Phyutility: A phyloinformatics tool for trees, alignments and molecular data. Bioinformation 2008, 24, 715–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. Bayesian analysis of molecular evolution using MrBayes. In Statistical Methods in Molecular Evolution; Nielsen, R., Ed.; Springer: New York, NY, USA, 2005; pp. 183–226. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest 2.3. Program Distributed by the Author; Evolutionary Biology Center, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Das, K.; Sharma, J.R.; Atri, N.S. Russula in Himalaya 3: A new species of subgenus Ingratula. Mycotaxon 2006, 95, 271–275. [Google Scholar]
- Chou, W.N.; Wang, Y.Z. Nine species of Russula (Basidiomycotina) new to Taiwan. Taiwania 2005, 50, 93–100. [Google Scholar]
- Wu, X.L.; Dai, Y.C.; Li, T.H.; Yang, Z.L.; Song, B. Fungi of Tropical China; Science Press: Beijing, China, 2010; pp. 1–477. [Google Scholar]
- Bazzicalupo, A.L.; Buyck, B.; Saar, I.; Vauras, J.; Carmean, D.; Berbee, M.L. Troubles with mycorrhizal mushroom identification where morphological differentiation lags behind barcode sequence divergence. Taxon 2017, 66, 791–810. [Google Scholar] [CrossRef]
- Tang, L.P.; Hao, Y.J.; Cai, Q.; Tolgor, B.; Yang, Z.L. Morphological and molecular evidence for a new species of Rhodotus from tropical and subtropical Yunnan, China. Mycol. Prog. 2013, 13, 45–53. [Google Scholar] [CrossRef]
- Xie, H.J.; Tang, L.P.; Mu, M.; Fan, Y.G.; Jiang, S.; Su, M.S.; Liang, Z.Q.; Zeng, N.K. A contribution to knowledge of Gyroporus (Gyroporaceae, Boletales) in China: Three new taxa, two previous species, and one ambiguous taxon. Mycol. Prog. 2022, 21, 71–92. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Lin, W.F.; Buyck, B.; Liang, Z.Q.; Su, M.S.; Chen, Z.H.; Zhang, P.; Jiang, S.; An, D.Y.; Zeng, N.K. Morphological and phylogenetic evidences reveal four new species of Cantharellus subgenus Cantharellus (Hydnaceae, Cantharellales) from China. Front. Microbiol. 2022, 13, 900329. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, M.; Liu, X.Y.; Wu, F.; Dai, Y.C. Phylogeny, divergence time estimation and biogeography of the genus Onnia (Basidiomycota, Hymenochaetaceae). Front. Microbiol. 2022, 13, 907961. [Google Scholar] [CrossRef] [PubMed]
- Aoki, W.; Bergius, N.; Kozlan, S.; Fukuzawa, F.; Okuda, H.; Murata, H.; Ishida, T.A.; Vaario, L.M.; Kobayashi, H.; Kalmiş, E.; et al. New findings on the fungal species Tricholoma matsutake from Ukraine, and revision of its taxonomy and biogeography based on multilocus phylogenetic analyses. Mycoscience 2022, 63, 197–214. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Voucher | Locality | GenBank Accession Nos. | Reference | |
|---|---|---|---|---|---|
| ITS | 28S | ||||
| Russula aff. amoenolens | HMJAU37318 | Heilongjiang, NE China | KY357332 | — | [31] |
| R. ahmadii | LAH 35004 | Pakistan | KT834638 | KX911834 | [32] |
| R. ahmadii | LAH 18081013 | Pakistan | KU535609 | — | [32] |
| R. amerorecondita | F (PGA17-017) | USA | MN130066 | — | [21] |
| R. amerorecondita | SH120828 | USA | MN130067 | — | [21] |
| R. amoenolens | 46372 (AH) | Spain | MK105626 | MK105707 | [33] |
| R. amoenolens | TUB nl27.9.95.6 | Germany | AF418615 | AF325295 | [34] |
| R. arunii | CUH:AM270 | India | KY450661 | KY946733 | [35] |
| R. arunii | CUH:AM261 (holotype) | India | KR872619 | KY946732 | [35] |
| R. blennia | 569/BB08.066 (holotype) | Madagascar | MH545687 | KU237556 | [36] |
| R. benghalensis | CUH AM654 (holotype) | India | MK414583 | MK496844 | [37] |
| R. catillus | SFC20120725-52 | Republic of Korea | KX574685 | — | [38] |
| R. catillus | SFC20120827-01 (holotype) | Republic of Korea | KX574686 | — | [38] |
| R. catillus | SFC20120919-35 | Republic of Korea | KX574688 | — | [38] |
| R. catillus | LHJ150915-19 | Guangdong, southern China | MK860690 | — | Unpublished |
| R. cerolens | F36 | USA | JN681168 | — | Unpublished |
| R. cf. pseudolepida | BPL247 | USA | KT933962 | KT933821 | [39] |
| R. clavulus | RITF3300 | Guizhou, SW China | MN317302 | MN325079 | [15] |
| R. clavulus | RITF3216 | Guizhou, SW China | MN317303 | MN325080 | [15] |
| R. clavulus | RITF3204 | Yunnan, SW China | MN317304 | MN325081 | [15] |
| R. clavulus | RITF3334 (holotype) | Yunnan, SW China | MN317301 | MN325078 | [15] |
| R. compacta | BPL242 | USA | KT933960 | KT933819 | [39] |
| R. compacta | HFJAU0301 | Jiangxi, SE China | MN258682 | — | Unpublished |
| R. echidna | HO 593336 | Australia | MN130079 | — | [21] |
| R. echidna | HO 593337 | Australia | MN130080 | — | [21] |
| R. elastica | 3/BB 06.009 | Madagascar | — | KU237451 | [40] |
| R. ferruginea subsp. ferruginea | HUA Corrales935 | Colombia | MZ604289 | MZ604284 | [40] |
| R. ferruginea subsp. ferruginea | HUA Corrales1019 | Colombia | MZ604291 | MZ604286 | [40] |
| R. ferruginea subsp. ferruginea | HUA Corrales944 (holotype) | Colombia | MZ604288 | MZ604283 | [40] |
| R. ferruginea subsp. panamensis | UCH A28 (holotype) | Panama | MZ604292 | MZ604287 | [40] |
| R. flavobrunnea var. violaceotincta | 71//BB 06.050 | Madagascar | — | KU237468 | [40] |
| R. fluvialis | KUO (JR8666) (holotype) | Finland | MN130084 | MN130125 | [21] |
| R. fluvialis | KUO (JR8313) | Finland | MN130085 | MN130126 | [21] |
| R. foetens | HMJAU 32234 | Heilongjiang, NE China | KX095018 | — | Unpublished |
| R. foetens | TUB hue124 | Germany | AF418613 | AF325299 | [34] |
| R. foetens | GENT FH-12-277 | Germany | KT934016 | KT933877 | [39] |
| R. foetentoides | LAH 04081023 | Pakistan | HE647707 | — | [41] |
| R. foetentoides | LAH 13081034 | Pakistan | HE647708 | — | [41] |
| R. foetentula | 156 | USA | KJ834623 | — | [42] |
| R. fragrantissima | 98 | Italy | KJ530751 | — | [42] |
| R. fujianensis | N.K. Zeng1449 (FHMU993) | Fujian, SE China | ON332048 | ON318391 | This study |
| R. fujianensis | N.K. Zeng1431 (FHMU975) (holotype) | Fujian, SE China | ON332049 | ON318392 | This study |
| R. garyensis | F (PGA17-008) | USA | MN130088 | — | [21] |
| R. garyensis | F (PGA1-910) | USA | MN130089 | — | [21] |
| R. gelatinosa | K15052626 (holotype) | Guangdong, southern China | MH168575 | — | [12] |
| R. gelatinosa | K16053119 | Guangdong, southern China | MH168574 | — | [12] |
| R. granulata | PC BB2004-226 | USA | EU598192 | — | Unpublished |
| R. granulata | PC BB2004-225 | USA | EU598190 | — | Unpublished |
| R. granulata | HMAS252604 | Jilin, NE China | KF850414 | — | [43] |
| R. granulata | BPL272 | USA | KT933971 | KT933832 | [39] |
| R. grata | HMJAU 38008 | Heilongjiang, NE China | KY681444 | — | [31] |
| R. griseobrunnea | JAC13201 | New Zealand | MW683824 | MW683661 | Unpublished |
| R. hainanensis | N.K. Zeng4888 (FHMU4860) | Hainan, southern China | — | OM280996 | [16] |
| R. hainanensis | N.K. Zeng4899 (FHMU4855) (holotype) | Hainan, southern China | OM280999 | OM280997 | [16] |
| R. heterophylla | UE20.08.2004-2 | Sweden | DQ422006 | — | [44] |
| R. hortensis | IB 1997/0787 (holotype) | Italy | HG798528 | — | Unpublished |
| R. illota | MICH73719 | France | KF245509 | — | Unpublished |
| R. illota | UE26.07.2002-3 | Sweden | DQ422024 | — | [34] |
| R. inamoena | 107 | Italy | KJ834597 | — | [42] |
| R. inamoena | 109 | Italy | KJ834595 | — | [42] |
| R. indocatillus | HGAS-MF009917 | Guizhou, SW China | MN649191 | — | [9] |
| R. indocatillus | HGAS-MF009903 | Guizhou, SW China | MN649192 | — | [9] |
| R. indocatillus | AG 17-1571 (holotype) | India | MN581483 | — | [45] |
| R. indocatillus | AG 18-1653 | India | MN581165 | — | [45] |
| R. indosenecis | AG-21-04A | India | OL701254 | — | [46] |
| R. indosenecis | AG-21-06A (holotype) | India | OL701269 | — | [46] |
| R. insignis | HMAS267740 | Heilongjiang, NE China | KF850404 | — | [9] |
| R. laurocerasi | FH12178 | Germany | KT933988 | KT933849 | [39] |
| R. laurocerasi | TUB nl1348 | Germany | AF418614 | — | [34] |
| R. laurocerasi | E00290534 | UK | KF245532 | — | Unpublished |
| R. livescens | F0177 | China | GU371295 | — | [47] |
| R. maguanensis | XHW4765 | Yunnan, SW China | MH724918 | MH714537 | [48] |
| R. mistiformis | JC170305NR | Spain | MK105677 | — | [33] |
| R. mistiformis | AMC H-69 | Spain | MK105680 | — | [33] |
| R. multilamellula | LHJ14092423 (holotype) | Guizhou, SW China | MT791347 | MT793120 | [15] |
| R. multilamellula | wuxingliang03 | Guizhou, SW China | MT791348 | MT793121 | [15] |
| R. mustelina | 1176/S. Adamcik 09.88 | Slovakia | — | KU237596 | [40] |
| R. mutabilis | DPL10654 | USA | KF810137 | — | [49] |
| R. mutabilis | BHI-F384a | USA | MF161239 | — | [50] |
| R. obscuricolor | KD 16-30 (holotype) | India | MF805816 | — | [51] |
| R. obscuricolor | KD 16-22 | India | MF805817 | — | [51] |
| R. oleifera | 254/BB 98.024 | Burundi | — | KU237490 | [40] |
| R. ombrophila | 86 | Spain | KF971694 | — | [42] |
| R. oraria | N.K. Zeng4829 (FHMU5374) | Hainan, southern China | ON332052 | OQ380695 | This study |
| R. oraria | N.K. Zeng4823 (FHMU5355) (holotype) | Hainan, southern China | ON332053 | OQ380696 | This study |
| R. pectinata | 2010BT02 | Germany | KF318081 | — | [42] |
| R. pectinata | 2010BT48 | Germany | KF318082 | — | [42] |
| R. pectinatoides | HMAS251202 | Yunnan, SW China | JX425405 | — | [43] |
| R. pectinatoides | NYS2303.1 | USA | KU640189 | — | [42] |
| R. pila | JMV800654 (BCN) | Spain | MK105691 | MK105736 | [33] |
| R. pilosella | BRI-H5974 | Australia | EU019941 | — | [52] |
| R. pseudocatillus | K16042406 (holotype) | Guangdong, southern China | MK049974 | — | [14] |
| R. pseudocatillus | K15060706 | Guangdong, southern China | MK049975 | — | [14] |
| R. pseudociliata | 545/BB 08.061 | Madagascar | MH545688 | KU237537 | [40] |
| R. pseudopectinatoides | HMAS265020 | Xizang, SW China | KM269079 | — | [11] |
| R. pseudopectinatoides | HMAS251523 (holotype) | Xizang, SW China | KM269077 | — | [11] |
| R. pseudopectinatoides | RITF1353 | China | MK860689 | — | [13] |
| R. pseudosenecis | AG 20-062 (holotype) | India | OL461233 | — | [46] |
| R. pseudosenecis | AG 21-073 | India | OL461234 | — | [46] |
| R. pulverulenta | 4-1144IS79 | USA | AY061736 | — | [53] |
| R. pulverulenta | PC BB2004-245 | USA | EU598186 | — | Unpublished |
| R. punctipes | K17052318 | Guangdong, southern China | MH168576 | — | [12] |
| R. punctipes | K16051001 | Guangdong, southern China | MH168577 | — | [12] |
| R. recondita | 92 | Switzerland | KJ530750 | — | [42] |
| R. rufobasalis | H17052204 (holotype) | Guangdong, southern China | MH168570 | — | [12] |
| R. rufobasalis | H15060622 | Guangdong, southern China | MH168567 | — | [12] |
| R. rufobasalis | N.K. Zeng1416 (FHMU961) | Fujian, SE China | ON332050 | ON318393 | This study |
| R. rufobasalis | N.K. Zeng1270 (FHMU827) | Fujian, SE China | ON332051 | ON318394 | This study |
| R. senecis | CUH AM102 | India | KP142981 | — | [54] |
| R. senecis | SFC20110921-18 | Republic of Korea | KX574698 | — | [38] |
| R. siamensis | Watling 28784 | Thailand | AB206535 | — | [55] |
| R. straminella | HGAS-MF009920 | Guizhou, SW China | MN649194 | — | [9] |
| R. straminella | HGAS-MF009922 (holotype) | Guizhou, SW China | MN649195 | — | [9] |
| R. straminella | HGAS-MF009925 | Guizhou, SW China | MN649189 | — | [9] |
| R. subfoetens | HMJAU 38006 | Heilongjiang, NE China | KY681430 | — | [31] |
| R. subpectinatoides | HBAU15023 (holotype) | Jiangsu, eastern China | MW041163 | — | [9] |
| R. subpectinatoides | HBAU15024 | Jiangsu, eastern China | MW041164 | — | [9] |
| R. subpectinatoides | HBAU15025 | Jiangsu, eastern China | MW041165 | — | [9] |
| R. subpectinatoides | HBAU15026 | Jiangsu, eastern China | MW041166 | — | [9] |
| R. subpunctipes | RITF2616 (holotype) | Hubei, central China | MK860692 | MK860695 | [13] |
| R. subpunctipes | RITF2617 | Hubei, central China | MK860693 | MK860696 | [13] |
| R. substriata | WJ292 | Yunnan, SW China | MH724919 | MH714538 | [48] |
| R. substriata | XHW4749 | Yunnan, SW China | MH724920 | MH714539 | [48] |
| R. substriata | XHW4766 (holotype) | Yunnan, SW China | MH724921 | MH714540 | [48] |
| R. substriata | XHW4767 | Yunnan, SW China | MH724922 | MH714541 | [48] |
| R. substriata | XHW4785 | Yunnan, SW China | MH724923 | MH714542 | [48] |
| R. succinea | HGAS-MF 009909 | Guizhou, SW China | MN649196 | — | [9] |
| R. succinea | HGAS-MF 009904 (holotype) | Guizhou, SW China | MN649188 | — | [9] |
| R. succinea | HGAS-MF 009906 | Guizhou, SW China | MN649198 | — | [9] |
| R. succinea | HGAS-MF 009915 | Guizhou, SW China | MN649190 | — | [9] |
| R. variata | BPL241 | USA | KT933959 | KT933818 | [39] |
| R. ventricosipes | PC 0142480 | USA | KY800364 | — | [56] |
| R. vinaceocuticulata | PDD 64246 | New Zealand | GU222258 | — | Unpublished |
| Russula sp. | LHJ170913-01 | Guangdong, southern China | MK860691 | MK860694 | [13] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.; Liang, Z.; Zeng, N. A Contribution to Knowledge of Russula Section Ingratae (Russulales, Russulaceae) in China: Two New Taxa and Amended Descriptions of One Known Species. Forests 2023, 14, 695. https://doi.org/10.3390/f14040695
Han Y, Liang Z, Zeng N. A Contribution to Knowledge of Russula Section Ingratae (Russulales, Russulaceae) in China: Two New Taxa and Amended Descriptions of One Known Species. Forests. 2023; 14(4):695. https://doi.org/10.3390/f14040695
Chicago/Turabian StyleHan, Yunxiao, Zhiqun Liang, and Niankai Zeng. 2023. "A Contribution to Knowledge of Russula Section Ingratae (Russulales, Russulaceae) in China: Two New Taxa and Amended Descriptions of One Known Species" Forests 14, no. 4: 695. https://doi.org/10.3390/f14040695