
Comparative Genomics Analysis of Ganoderma Orthologs Involved in Plant-Pathogenesis

Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia (UPM), Serdang 43400, Selangor, Malaysia
Forests 2023, 14(3), 653; https://doi.org/10.3390/f14030653
Submission received: 8 February 2023
/
Revised: 14 March 2023
/
Accepted: 20 March 2023
/
Published: 22 March 2023
(This article belongs to the Section Forest Health)
Abstract
:Ganoderma species are producers of bioactive secondary metabolites and lignin degraders. A few Ganoderma species are known to be plant pathogens that attack economically important trees. In this study, comparative genomics analysis was conducted on the proteome of ten Ganoderma species/strains, focusing on the proteins that have been reported to be involved in plant-pathogenesis in other fungi. Fungal trophic lifestyle prediction of these Ganoderma species/strains supported that G. boninense (a potent pathogen to oil palm) is a hemibiotrophic fungus while the other Ganoderma species/strains analyzed were predicted to be saprophytes or a symbiont based on their Carbohydrate-Active Enzyme (CAZyme) contents. Although these Ganoderma species/strains were demonstrated to share many CAZymes and secondary metabolite core gene clusters, individual species may produce unique CAZymes and secondary metabolite core genes that determine their lifestyles, host-specificity, and potential as a producer of bioactive secondary metabolites. Ortholog groups that are related to fungal virulence from seven Ganoderma species/strains including those involved in lignin degradation, mycotoxin, siderophore and ergosterol biosynthesis, and virulence were summarized. Potential effectors were predicted from the proteome of these Ganoderma species/strains, and putative effectors that were being expressed in G. boninense in oil palm roots but not found in other species were identified. The findings provide a useful resource to further analyze plant-pathogenesis and wood degradation activities of these Ganoderma species.
1. Introduction
The genus Ganoderma (Ganodermataceae; Polyporales) consists of more than 300 species of widely distributed wood-decaying fungi [1]. While some are saprophytes that feed on dead wood, others are hemibiotrophic pathogens that decompose living hardwood trees, such as conifers, rubber trees, oil palms, and coconut palms [2,3,4] by producing lignin- and cellulose-degrading enzymes. Ganoderma species are also featured as producers of secondary metabolites (mainly triterpenes such as ganoderic acids), oligosaccharides, polysaccharides, polysaccharide–peptide complex, phenols, and steroids that have potential bioactivities [1,5,6,7,8]. A few Ganoderma species, such as G. lucidum and G. sinense, have been used in traditional medicine [9,10]. The transcript sequences for enzymes related to the biosynthesis of terpenes [11,12,13] and ganoderic acids [14] have been identified. The expression of genes in the mevalonate pathway has also been profiled [15,16,17]. The wood-decaying capacity and pathogenicity of Ganoderma species is associated with the ability of these fungi to produce plant-cell-wall degrading enzymes. The gene sequences encoding carbohydrate-active enzymes (CAZymes) were identified from the genome assembly of G. boninense strain NJ3, G. boninense strain G3, G. australe, G. leucocontextum, G. lingzhi, G. sinense, and G. tsugae [11,13,18]. The transcript sequence and abundance of a few manganese peroxidases and laccases from G. boninense PER71 have also been reported [19], following the identification of these transcripts in the transcriptome of infected oil palm [20]. In addition, the purification and characterization of laccases from different Ganoderma strains [21,22,23,24,25], recombinant manganese peroxidases [26], and laccases from G. lucidum and G. australe [27,28,29] were reported. Other transcripts related to pathogenesis that have been characterized include the necrosis and ethylene inducing protein (NEP)-like protein and cyclophilin from G. boninense [30,31], while the sequences of hydrophobins from different Ganoderma species were reported by Sulaiman et al. [32].
As of March 2023, 17 genome resources from eight Ganoderma species were available in the National Center for Biotechnology Information (NCBI) database compared to 12 genome assemblies from six Ganoderma species in March 2022. However, only the full suite of annotated proteins from G. sinense [33] and G. leucocontextum Dai12418 [34] were available in NCBI, while the predicted proteins from the nuclear genomes of other Ganoderma species remained unpublished, unknown, or no longer accessible from the portals mentioned in publications [14,17,32]. In addition, a total of 5957 predicted protein sequences from G. boninense strain NJ3 were deposited by Ramzi et al. [18] in NCBI. Kües et al. [14] compared the genome sequence data of three Ganoderma species, i.e., Ganoderma sp. 10,597 [35], G. lucidum strain Xiangnong No. 1 [36], and G. lucidum strain G260125-1 [15], focusing on the sequence analysis of proteins belonging to the CYP450 family and in the mating loci matA and matB from these Ganoderma species/strains. Recently, the phylogenomic and comparative genomics of a few Ganoderma species focusing on genes involved in terpene metabolism and gene encoding secreted proteins were also reported [12,13,18]. In another recent paper, Khairi et al. [37] reported the predicted effector proteins from three G. boninense strains. Nevertheless, comparative genomics analyses of Ganoderma species, especially on proteins involved in plant-pathogenesis, are still limited.
Hence, the current study focused on the comparative genomics of putative orthologs that participate in plant-pathogenesis and wood degradation from ten Ganoderma species/strains. Orthologs that could potentially contribute to fungal virulence and pathogenesis of seven Ganoderma strains in five different species were also identified and analyzed. The findings could serve as an impetus for future molecular cloning and characterization of genes from these Ganoderma species.
2. Materials and Methods
2.1. Retrieval of Genome Assemblies and Protein Sequences
The genome assemblies of Ganoderma species (G. boninense strain G3 GCA_002900995.2; G. boninense strain NJ3 GCA_001855635.1; G. lucidum strain BCRC36111 GCA_012655175.1; G. lucidum strain BCRC37177 GCA_000338035.1; G. lucidum strain G260125-1 GCA_000262775.1; G. lucidum strain Xiangnong No.1 GCA_000271565.1; G. multipileum GCA_000338015.1; G. sinense ZZ0214-1 GCA_002760635.1; G. tsugae GCA_003057275.1; and Ganoderma sp. BRIUMSc GCA_008694245.1) and the annotated protein sequences of G. sinense ZZ0214-1 were retrieved from GenBank in December 2020. Table S1 shows the summary of the Ganoderma species, strains, hosts, and other information.
2.2. Gene Prediction from Ganoderma Species
The genome assembly and reference protein sequences of G. sinense ZZ0214-1 were submitted to WebAUGUSTUS Service (http://bioinf.uni-greifswald.de/webaugustus/index.gsp; accessed on 9 December 2020) as a training gene set to generate training species-specific parameters. Gene prediction from the other Ganoderma species was conducted in WebAUGUSTUS by using the training species-specific parameters obtained for G. sinense ZZ0214-1. The completeness of predicted proteome was evaluated by the Benchmarking Universal Single-Copy Orthologs (BUSCO) tool [38] with the polyporales_odb10 database.
2.3. Functional Annotation
BLAST2GO was used for the functional annotation of predicted proteins from Ganoderma species by assigning Gene Ontology identifier (GO ID), EC number, and pfam to the query sequences. Blast2GO [39] was conducted with a minimum cut-off of 10−5 for BlastP against the SwissProt database and default parameters set by OmicsBox (www.biobam.com accessed on 27 March 2021), i.e., annotation cut-off (55), GO-weight (5), HSP-hit coverage cut-off (0), and apply no filter GO by taxonomy. GO enrichment using Fisher’s exact test (p < 0.01, FDR <0.05) was performed in OmicsBox. A KEGG Orthology (KO) identifier (K number) was assigned to the sequence by GHOSTX searches in GhostKOALA (www.kegg.jp/ghostkoala/; accessed on 27 March 2021; [40]), respectively, against the “genus_prokaryotes+family_eukaryotes” database in Kyoto Encyclopedia of Genes and Genomes (KEGG).
2.4. Identification of Secondary Metabolite Gene Clusters, CAZyme and Classification of Fungal Lifestyles
The secondary metabolite gene clusters were identified by antiSMASH fungal version v6.0 [41] from each Ganoderma species/strain using the standard (recommended) parameters at the antiSMASH webserver (https://fungismash.secondarymetabolites.org/; accessed on 2 August 2022). CAZymes were identified using hmmscan in HMMER 3.0 [42] with the “-domtblout” parameter, followed by the shell/perl script “hmmscan-parser.sh” in dbCAN ver. 6 [43] with a minimum alignment length of 80 amino acids, E-value < 10−5, and >30% coverage of HMM and included in a prediction tool known as CAZyme-Assisted Training And Sorting of trophy (CATAStrophy) [44]. The trophic lifestyles of Ganoderma species/strains were inferred according to CAZyme content by CATAStrophy. Principal component analysis (PCA) of CAZyme contents in CATAStrophy allowed the separation of most of the species/strains with the first two principal components (PCs). Hence, PC1 and PC2, which separated most of the species according to lifestyles (biotrophs, hemibiotrophs, necrotrophs, saprotrophs, and symbionts), were plotted in Excel for visualization.
2.5. Ortholog Analysis
Orthologs were identified by OrthoMCL (https://orthomcl.org/; [45] in OrthoVenn (https://orthovenn2.bioinfotoolkits.net/home; accessed on 14 February 2021; [46]) at an E-value of 10−5. Ortholog clusters related to virulence and pathogenesis were identified by keyword search or GO IDs. The Venn diagrams were drawn with InteractiVenn (http://www.interactivenn.net/index.html; accessed on 18 February 2021; [47]).
2.6. Prediction of Effector Sequences
Phobius (http://phobius.sbc.su.se/instructions.html; accessed on 13 March 2021; [48]) was used to predict fungal proteins with trans-membrane topology and signal peptide. EffectorP ver 2.0 (http://effectorp.csiro.au/; accessed on 17 March 2021; [49]) was used for the prediction of putative effector sequences from the putative protein sequences of Ganoderma species with transmembrane topology and signal peptide. Protein sequences that have BlastP matches against the SwissProt database with an E-value of 10−5 or less were considered to be significant. Fisher’s exact test (p < 0.01, FDR < 0.05) was conducted to identify the GO terms that were enriched among the predicted effectors in each Ganoderma species.
Identification of putative effectors that were present in oil palm infected by G. boninense (G) and Trichoderma (T) was performed by mapping the RNAseq data (European Nucleotide Archive (ENA) accession number: PRJEB7252) of a previous transcriptome study [20] to the genes of G. boninense strain G3 in OmicsBox (www.biobam.com; accessed on 4 April 2021).
3. Results
3.1. Prediction of Proteins of Ganoderma Species/Strains
Since G. sinense is the only Ganoderma species with annotated proteins among the Ganoderma genomes retrieved from NCBI when this study was conducted, the proteins from other Ganoderma genome assemblies were predicted with the protein sequences of G. sinense as a training set. The number of predicted proteins ranged from 11,040 in G. lucidum G260125-1 to 21,487 from G. boninense strain NJ3. The numbers of predicted proteins from the two G. boninense genome assemblies, i.e., G. boninense strain G3 and G. boninense strain NJ3, were comparable to those reported by an independent group [37] on the same strains. Based on BUSCO analysis, the completeness of the predicted proteomes of G. lucidum strain BCRC37177, G. multipileum, and G. sinense ZZ0214-1 was more than 90% while the completeness of the predicted proteomes of G. boninense strain NJ3 and Ganoderma sp. BRIUMSc was only 66%–69% (Table 1), mainly due to the presence of a high number of fragmented and missing proteins (Table 1).
Table 1 also shows the number of proteins with significant matches to SwissProt database (≤10−5), GO, and KO annotations from each Ganoderma species or strain. The number of proteins with BlastP matches from each species/strain ranges from 39%–57%. Almost all the proteins with BlastP matches were assigned with at least one GO ID. The number of genes with KO from each species/strain ranges from 24%–32%. The four G. lucidum strains and G. multipileum were found to have a higher number of annotated proteins compared to other species/strains. G. boninense strain NJ3 has the lowest number of annotated proteins, possibly due to fragmented genes or incomplete genome sequencing data (Table 1).
3.2. CAZymes and Classification of Prediction of Fungal Lifestyles
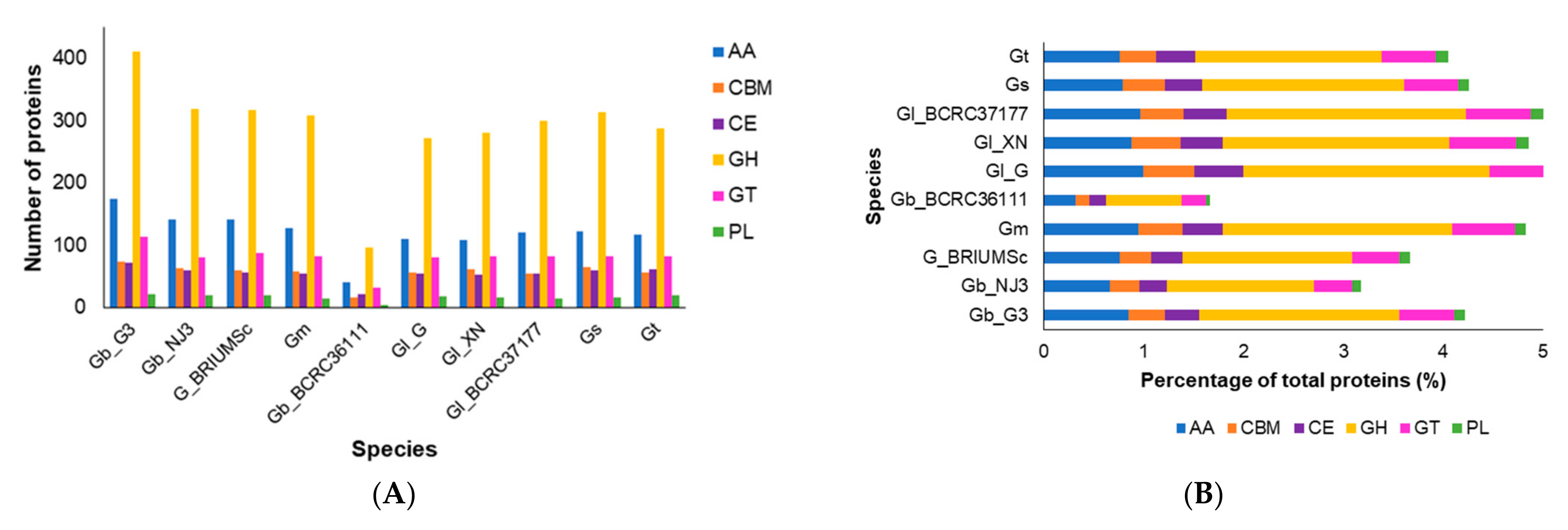
Figure 1 shows the CAZyme content in Ganoderma species/strains. The G. boninense strain G3 was found to have the highest number of CAZymes (Figure 1A), especially those in the glycosyl hydrolase (GH) family and glycosyl transferase (GT) family. The CAZymes in the GH family dominated the predicted CAZymes from all Ganoderma species/strains, followed by CAZymes with auxiliary activities (AA) and GT activities (Figure 1B). The three most prominent GH families among all Ganoderma species/strains were GH18 (including chitinases Classes III and V and xylanase inhibitor), GH16_1 (including xyloglucan:xyloglucosyltransferase), and GH3 (including β-glucosidase). A few enzymes seemed to be unique to one or a few Ganoderma species/strains, including GH13 enzyme (possibly with α-amylase or sucrose phosphorylase activity), which was only found in G. boninense strain G3, and GH5 (endo-β-1,4-glucanase/cellulase), which was only found in G. lucidum strain BCRC37177 and G. multipileum. A few CAZymes were found to be missing from G. boninense strains, including GH43-33 (α-L-arabinofuranosidase), GH45 (xyloglucan-specific endo-β-1,4-glucanase or endo-β-1,4-mannanase), and GH99 (mannan endo-1,2-α-mannanase).
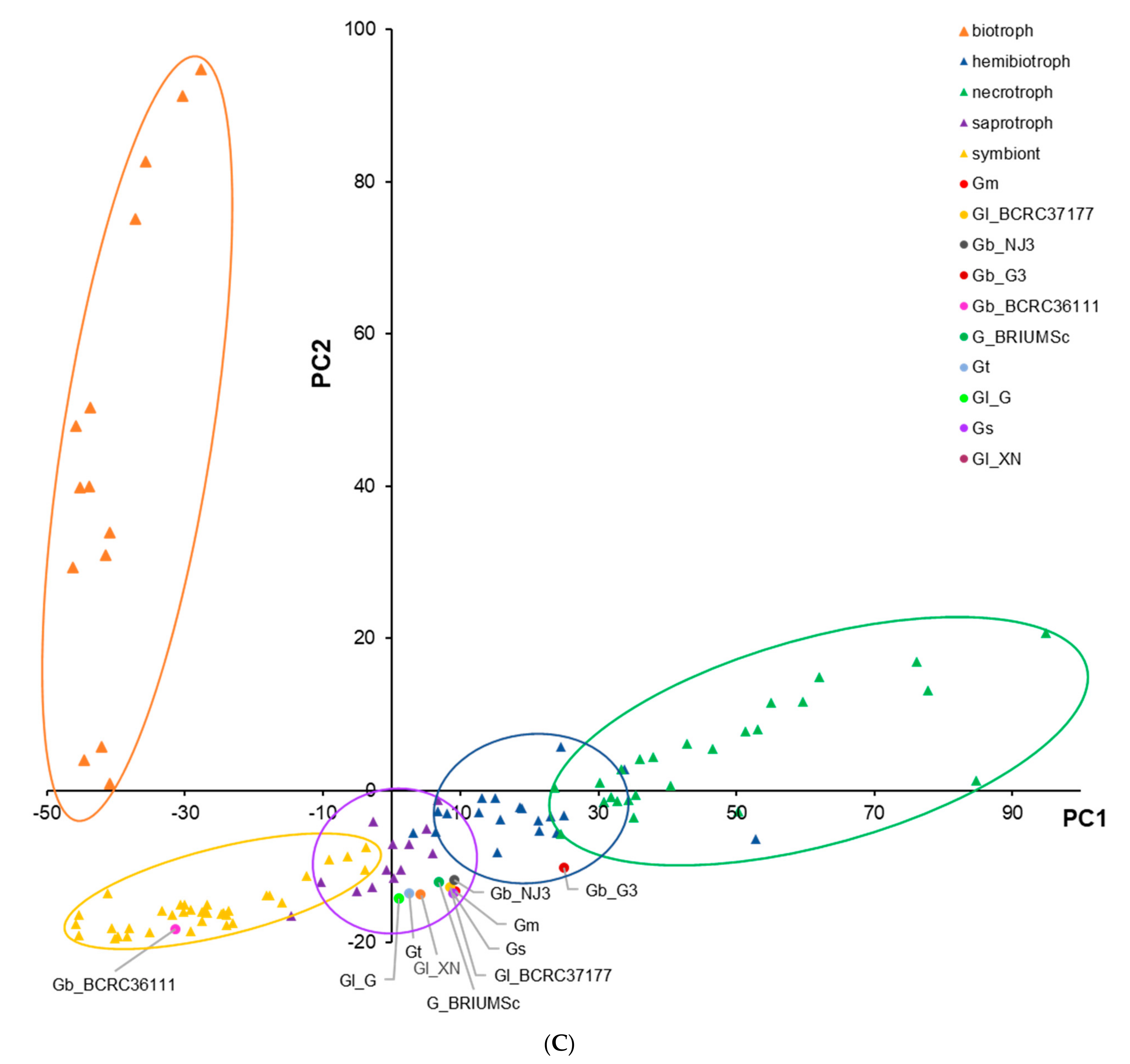
The AA class CAZymes were dominated by a few subclasses, such as the AA3_2 (including both aryl alcohol oxidase and glucose 1-oxidase in the glucose-methanol-choline oxidoreductases family with flavin-adenine dinucleotide (FAD)-binding domain), AA9 (copper-dependent lytic polysaccharide monooxygenases), AA1_1 (laccases), AA2 (class II lignin-modifying peroxidases), and AA7 (gluco- or chito- oligosaccharide oxidase, cellooligosaccharide dehydrogenase), while the GT class CAZymes have many enzymes belonging to subclasses GT2 (especially GT2_ Chitin_synth_2), GT1, and GT8. All Ganoderma species/strains were classified as saprotrophs by CATAStrophy according to their CAZyme contents (Figure 1C), except G. boninense strain G3, which was identified as a hemibiotroph and G. lucidum strain BCRC36111, which was predicted to be a symbiont. However, the PCA plot of PC1 and PC2 which separated most of the species according to lifestyles shows some overlap between the neighboring clusters, except for the biotroph cluster (Figure 1C).
3.3. Secondary Metabolite Production Potential of Ganoderma Species
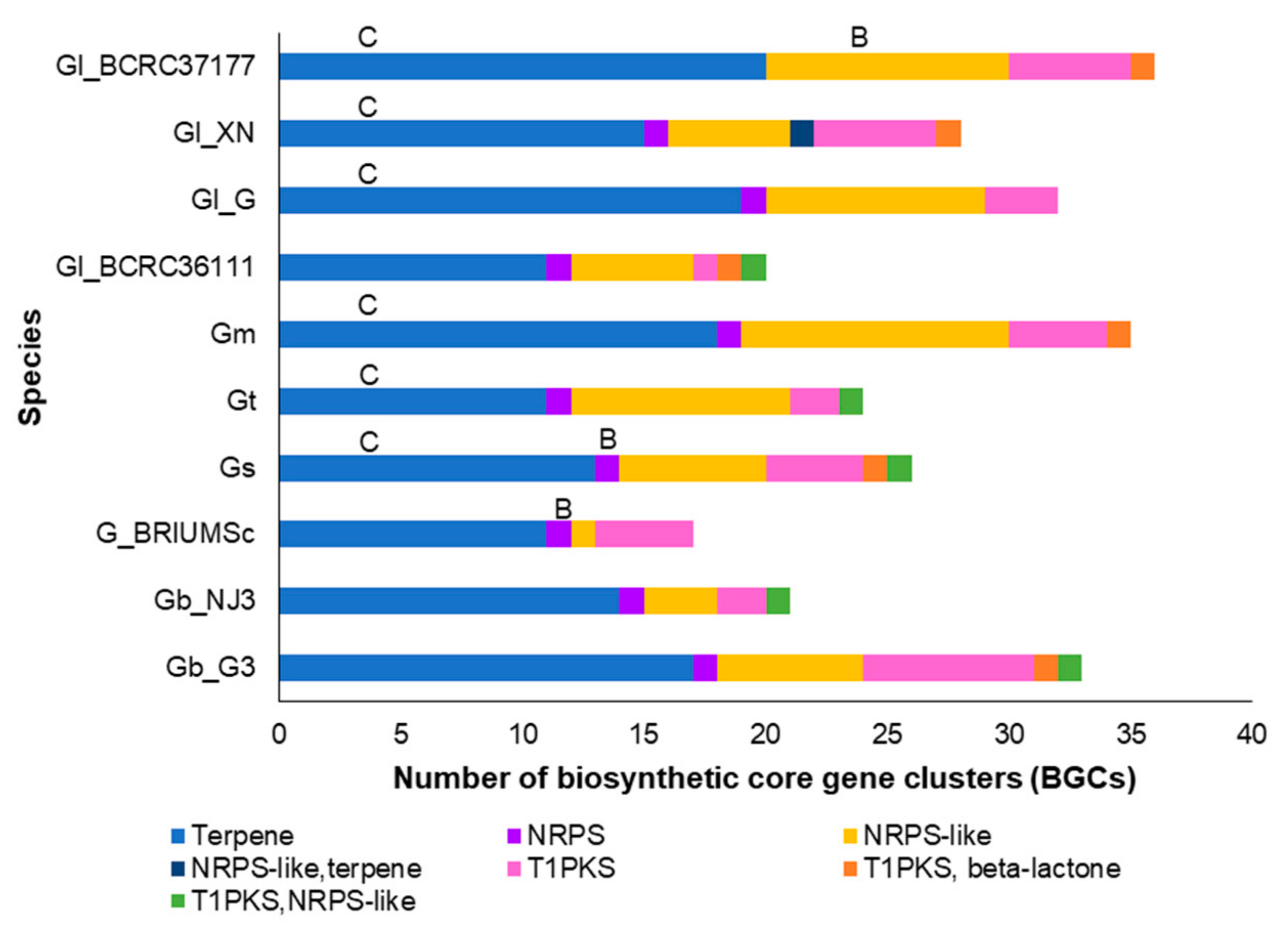
The secondary metabolite production potential of the Ganoderma species/strains was inferred from the putative biosynthetic core gene clusters (BGCs) for secondary metabolite production identified in each species/strain. The counts of BGCs vary among the species/strains, whereby G. lucidum strain BCRC37177, G. multipileum, G. lucidum strain G260125-1, and G. boninense strain G3 have more than 30 putative BGCs. Most of the Ganoderma species/strains were found to have the highest counts for BGCs related to terpene biosynthesis followed by the BGCs for nonribosomal peptide synthase (NRPS)-like while only one BGC for NRPS was detected in each species/strain (except for G. lucidum strain BCRC37177 which has none). The G. boninense strain G3 was found to have a slightly higher number of BGCs associated with type 1 polyketide synthase (T1PKS) (Figure 2). The BGC for β-lactone production was identified in six Ganoderma species/strains but absent in G. lucidum strain G260125-1, G. tsugae, G. boninense strain NJ3, and Ganoderma sp. BRIUMSc. The BGC for clavaric acid production was identified as the most similar known cluster for terpene clusters in most Ganoderma species/strains except for G. boninense strain G3, G. boninense strain NJ3, Ganoderma sp. BRIUMSc, and G. lucidum strain BCRC36111. Meanwhile, the BGC for the biosynthesis of basidioferrin (an iron chelator) was identified as the most similar known cluster for NRPS/NRPS-like clusters in G. lucidum strain BCRC37177, G. sinense, and Ganoderma sp. BRIUMSc.
3.4. Identification of Ganoderma Orthologs
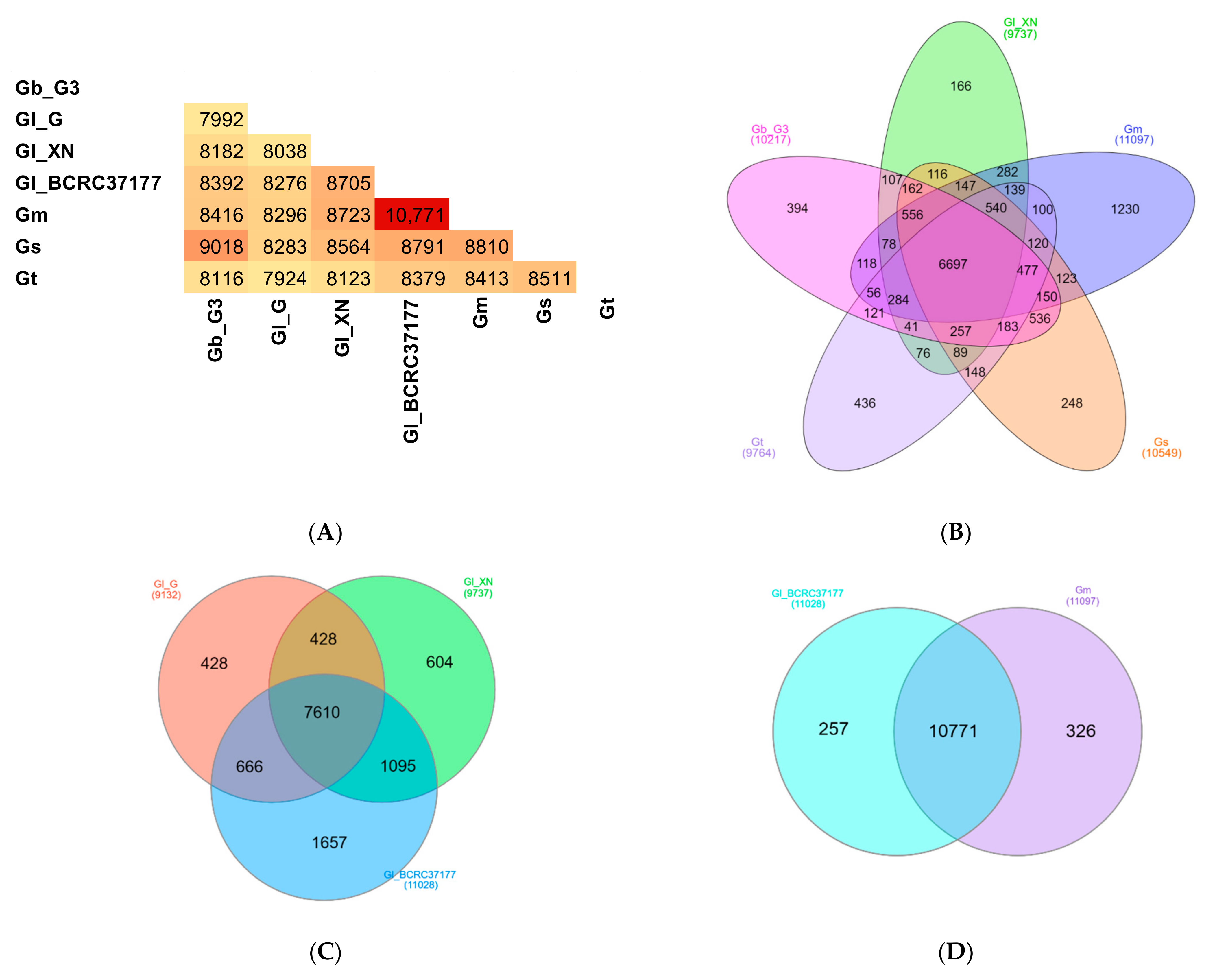
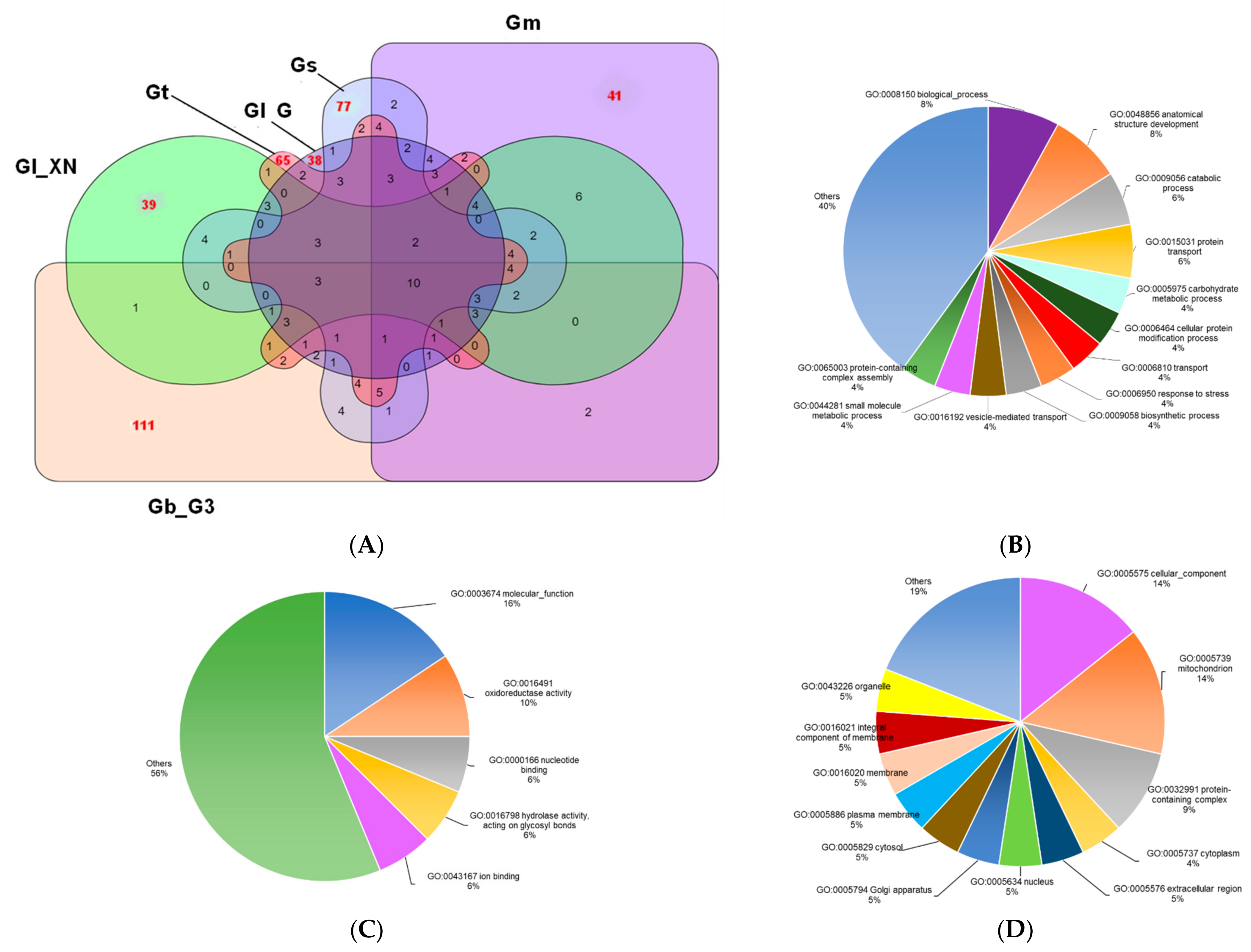
Since some of the predicted proteomes were of low quality, only the Ganoderma orthologs from seven Ganoderma proteomes which were more than 80% complete (Table 1) were further analyzed in this study. A total of 83,241 proteins were assigned to 14,238 ortholog clusters, with 6255 of them (50,153 proteins) shared by all seven Ganoderma species/strains whereas a total of 15,095 proteins were found in a single species/strain (singletons). A heatmap (similarity matrix of pairwise proteome comparisons) was plotted based on the ortholog clusters shared between a pair of proteomes (Figure 3A). G. lucidum strain BCRC37177 was found to share the highest number of ortholog clusters with G. multipileum.
The numbers of orthologous groups shared by representatives of five different Ganoderma species, i.e., G. boninense, G. lucidum, G. multipileum, G. sinense, and G. tsugae, are shown in a Venn diagram in Figure 3B. G. lucidum strain Xiangnong was chosen as the representative for G. lucidum instead of G. lucidum strain BCRC37177, although the latter has a more complete proteome because G. lucidum strain BCRC37177 was found to share a higher similarity with G. multipileum instead of other G. lucidum strains (Figure 3C). Figure 3B shows that 6697 clusters were shared by all five Ganoderma representative species, while the number of orthologous clusters shared by four, three, and two Ganoderma species were 2114, 1165, and 1727, respectively. A total of 2474 orthologous groups consisted of proteins from a single Ganoderma species (with the highest number of singletons in G. multipileum). Comparison of orthologous groups of the three G. lucidum strains (Figure 3C) revealed that 7610 orthologous groups were shared by all strains. The number of unique orthologous clusters which was only found in the proteome of a single Ganoderma species was 2689, mainly from G. lucidum strain BCRC37177. G. lucidum strain BCRC37177 shares the highest number of ortholog clusters with G. multipileum (previously mistaken as a tropical G. lucidum; [50]). A total of 10,771 orthologous groups were shared between G. lucidum strain BCRC37177 and G. multipileum (Figure 3D). Since the nomenclature of Ganoderma is confusing and no consensus has been achieved [5], it is possible that either of these Ganoderma spp. was incorrectly named.
3.5. Ganoderma Orthologous Groups Associated with Pathogenesis
The current study focuses on the ortholog groups with GO IDs related to biological and molecular functions that could possibly contribute to fungal virulence in plant-pathogenesis. A total of 98 ortholog clusters with GOs related to pathogenesis including mycotoxin biosynthesis, siderophore biosynthesis, transcription factors, transporter, fungal development, cell wall degradation, and others were found. Some of these ortholog groups are shown in Table 2.
3.5.1. Mycotoxin and Secondary Metabolite Biosynthesis
A total of 35 ortholog clusters related to terpenoid biosynthetic process were discovered, including many polyketide synthases, cytochrome P450 monooxygenases, FAD-dependent monooxygenase, oxidoreductases, methyltransferases, and dehydrogenases (Table 2). Based on a homology search, some of them were possibly involved in the biosynthesis of mycotoxins such as yanuthone D, melleolides, culmorin, botcinic acid and botcinin derivatives terretonin, and mycophenolic acid, etc. In addition, Ganoderma orthologs related to alkaloid metabolism were identified. The orthologs for isoflavone reductase (IFR) homolog A622-like (a putative NADPH-dependent oxidoreductase involved in the biosynthesis of tobacco alkaloids; [51]) and chanoclavine-I aldehyde reductase (one of the enzymes which mediate the biosynthesis of a fungal ergot alkaloids known as isofumigaclavines; [52]) were also found in the Ganoderma species (Table 2).
Among the Ganoderma orthologs related to peptide mycotoxin biosynthesis, putative enzymes involved in the biosynthesis of AF-toxins e.g., aldo-keto reductase AFTS1, and enoyl-CoA hydratase AFT3-1 were identified. The Ganoderma orthologous implicated in the biosynthesis of aflatoxin and other mycotoxins (e.g.; ustiloxin B, gliotoxin and bibenzoquinone oosporein) include norsolorinic acid ketoreductase, O-methylsterigmatocystin oxidoreductase, FAD-binding monooxygenase aflW, and O-methylsterigmatocystin oxidoreductase, oxidase ustYa, cytochrome P450 monooxygenase gliF, and FAD-dependent monooxygenase and hydroxylase OpS4. In addition, there are orthologs for glutathione S-transferases which were possibly involved in toxin catabolic process. Several Ganoderma orthologs related to the biosynthesis of siderophore ferrichrome A, such as hydroxymethylglutaryl (HMG)-CoA synthase, enoyl-CoA isomerase/hydratase fer4, Fe-regulated protein 8, and acyl-CoA ligase sidI [53,54,55], were also identified (Table 2).
3.5.2. Ergosterol Biosynthesis and Sterol Transport
Ortholog clusters related to ergosterol biosynthesis (e.g., 3-keto-steroid reductases, C-8 sterol isomerase, ergosterol biosynthetic protein 28, NADH-cytochrome b5 reductase, sterol 24-C-methyltransferase, sterol-4-α-carboxylate 3-dehydrogenase) and regulation of ergosterol biosynthetic process (damage response protein 1) were also identified (Table 2). In addition, protein clusters related to sterol transport such as oxysterol-binding protein, pathogen related in yeast (PRY)2, PRY3, phosphatidylglycerol/phosphatidylinositol transfer protein and non-specific lipid-transfer protein (sterol carrier protein 2) were also identified in the Ganoderma species.
3.5.3. Other Virulence Factors
The orthologs for transcription factors related to plant-pathogenesis in Ganoderma species include global transcription regulator sge1 [56], bZIP transcription factor hapX [57], and C2H2 finger domain transcription factor dvrA [58]. Other orthologs related to virulence in Ganoderma species include calcineurin subunit B [59], LysM domain-containing protein ARB_00327 [60], extracellular metalloproteinase, metallocarboxypeptidase A-like protein, aspartic-type endopeptidase, leucine aminopeptidase, tripeptidyl-peptidases [61], which play different roles in fungal virulence (Table 2).
3.5.4. Lignolytic Enzymes and Other Plant-Cell-Wall Degrading Enzymes
Many ortholog clusters related to lignin catabolic process, including 3-O-methyltransferases, 4-O-methyl-glucuronoyl methylesterases, 4-O-methyltransferases, and laccases that may play a role in modifying or degrading plant lignin, were found in Ganoderma species. The seven Ganoderma species were found to have 103 laccases in total (Table 2). Peroxidases including 16 lignin peroxidases, 19 manganese peroxidase, and 19 versatile peroxidases were found among the ortholog groups related to stresses from Ganoderma species. In addition, ortholog clusters related to pectin biosynthetic process, pectin catabolic process, and cellulose catabolic process were also found among the Ganoderma species.
3.6. Putative Effectors from Ganoderma Species
In this study, only proteins with a signal peptide (normally 20–30 amino acid residue in length and positively charged, followed by a hydrophobic region and a cleavage site) were used as inputs for EffectorP analysis because signal peptides were expected to be important for the secretion of effectors for plant infection. The predicted effector proteins ranged from 0.85%–0.98% of each proteome, with the lowest percentage in G. boninense strain G3 and the highest percentage in G. lucidum strain BCRC37177 (Table 3). The predicted effectors with functional annotation ranged from 45.7%–59.0%. Among these effectors were allergens (such as Asp F7 and Asp F15), thaumatin-like proteins, PRY2, hydrophobins, papain inhibitor, ustiloxin B biosynthesis protein UstYa, etc. The GO:0005576 (extracellular region) was found to be overrepresented among the GOs of the predicted effectors from four Ganoderma species (i.e., G. boninense strain G3, G. lucidum strain BCRC37177, G. sinense ZZ0214-1 and G. tsugae), while GO:0005615 (extracellular space) was found to be overrepresented among the GOs of the predicted effectors of G. lucidum BCRC37177. Other GO terms that were overrepresented among the GOs of the predicted effectors include GO:0016798 (hydrolase activity acting on glycosyl bonds; in G. tsugae), GO:0010466 (negative regulation of peptidase activity; in G. lucidum strain G260125-1 and G. multipileum), and GO:0004867 (serine-type endopeptidase inhibitor activity; in G. lucidum strain G260125-1 and G. multipileum). No GO terms were over-represented among the predicted effectors of G. lucidum Xiangnong (Table 3).
The predicted effectors of Ganoderma species/strains were grouped into ortholog groups whereby 10 effector ortholog clusters were shared by six Ganoderma species/strains (Figure 4A), including papain inhibitor and thaumatin-like protein. About 21.7% of these ortholog groups (68 from a total of 313 groups) were conserved in at least four Ganoderma species/strains. Meanwhile, there are also putative effectors that are unique to only a single species (Figure 4A). Among the 175 predicted effectors from G. boninense strain G3, 111 (63.5%) of them were not found in other Ganoderma species. Figure 4B–D shows the distribution of the unique effectors from G. boninense strain G3 in different GO categories. Meanwhile, 72 of the predicted effectors from G. boninense strain G3 were found to be present in the transcriptome oil palm infected by G. boninense PER71 [20], including papain inhibitor, allergen Asp F15, thaumatin-like protein 3.5 (allergen Cry j 3.5), protein PRY2, cytochrome P450 monooxygenase FSL4 (Fusarielin biosynthesis cluster protein 4), probable glycosidase C21B10.07 (EC 3.2.1.-), hydrophobin SC1, hydrophobin-1, etc. However, only 35 of these 72 predicted effectors were found to be unique to G. boninense strain G3 (not found in other Ganoderma species analyzed) (Figure 5). Almost all of these candidate effectors were found to be present only in oil palm roots treated with G. boninense PER71 but not in oil palm root sample treated with Trichoderma.
4. Discussion
Ganoderma species are known to be either saprophytes that degrade dead wood or hemibiotrophs that cause stem, butt, and root rot in living trees [14]. All the Ganoderma species/strains analyzed in this study were predicted to be saprophytes except for G. boninense strain G3, which was predicted to be a hemibiotroph, and G. lucidum strain BCRC36111, which was predicted to be a symbiont, according to their CAZyme contents. G. boninense is a known pathogen which causes basal stem rot in oil palms [2]. Although G. boninense strain G3 was predicted to be a hemibiotroph as expected, the G. boninense strain NJ3 was classified as a saprophyte by CATAStrophy. This could possibly be due to the inferior quality of proteome from G. boninense strain NJ3. G. lucidum was generally found to be non-invasive to oil palms [62]. Although G. lucidum, G. boninense, and a few other Ganoderma species have been previously implicated as pathogens associated with stem rot in coconut [3], Vinjusha and Arun Kumar [63] revised the Ganoderma species associated with stem rot in coconut palms as G. keralense and G. pseudoapplanatum. Meanwhile, the ability of other Ganoderma species analyzed in this study to infect living trees was not clear. The presence and absence of certain groups of CAZymes were possibly important in determining the host plant specificity of a Ganoderma species/strain or in the determination of its trophic lifestyle. G. boninense strain G3, which was predicted to have a hemibiotrophic lifestyle, was found to have a higher number of GHs and GTs compared to other Ganoderma species or G. bonienese strain NJ3.
Ganoderma species are white rot fungi that can degrade lignin; hence, it is logical that cellulose-, lignin-, xylan-, and chitin-degrading enzymes were found to be widespread in the Ganoderma species/strains. The same groups of CAZymes were also reported by Sun et al. [13] in their study on the CAZymes from seven Ganoderma species, including G. australe, G. leucocontextum, and G. lingzhi, which were not analyzed in this study. The Ganoderma species/strains were found to have a greater number of laccases than lignin peroxidases, manganese peroxidases, and versatile peroxidases. In addition, Ganoderma species/strains were also found to be rich in lytic polysaccharide monooxygenases, especially aryl alcohol oxidase, glucose 1-oxidase, copper-dependent lytic polysaccharide monooxygenases, gluco- or chito- oligosaccharide oxidases, and cellooligosaccharide dehydrogenases. These polysaccharide depolymerases may facilitate degradation of polysaccharides in plant cell walls. The ligninolytic enzymes and polysaccharide depolymerases are useful for the decomposition of wood, recycling of carbon, and lignin degradation for bioethanol production [64]. Pathogenic Ganoderma species may also use some of these CAZymes to degrade and modify plant cell walls during plant infection.
Ganoderma species were found to have a high content of biosynthetic core gene clusters associated with terpene biosynthesis and nonribosomal peptide synthase (NRPS)-like genes, in agreement with the findings of Jiang et al. [11] on the genomes of G. tsugae, G. lingzhi, G. sinense, and G. boninense. Most Ganoderma species/strains have the biosynthetic core gene for clavaric acid (a triterpenoid) except for G. boninense strain G3, G. boninense strain NJ3, and Ganoderma sp. BRIUMSc. Clavaric acid is found to be a potent inhibitor of the RAS farnesyltransferase, which plays a crucial role in the proliferation of tumor cells [65]. Its role in plant-pathogenesis has not been reported. The enzymes that participate in terpenoid biosynthesis were found to be present in all Ganoderma species/strains. Some of these proteins displayed high sequence similarities to enzymes in the biosynthesis of mycotoxins. Mycotoxins not only cause disease and death in the vertebrates that consume contaminated agricultural products [66] but may also be phytotoxic and play a role in plant pathogenesis [67,68]. In addition to terpenoids, peptide and ergot alkaloids form a large family of mycotoxins that consist of amino acid and isoprenoid precursors [66]. Ganoderma species were also found to have enzymes that are involved in the biosynthesis of AF-toxins, ustiloxin B, gliotoxin, bibenzoquinone oosporein, tryprostatins, ergovaline, and fumiclavanine C. The biosynthesis of basidioferrin siderophore is known to be catalyzed by an NRPS [69]. The gene cluster associated with the biosynthesis of basidioferrin was identified among the NRPS-like BGCs of G. lucidum strain BCRC37177 and G. sinense as well as the NRPS cluster of Ganoderma sp. BRIUMSc. However, all the Ganoderma species analyzed in this study were found to have a complete suite of enzymes in the biosynthesis of siderophore ferrichrome A, which contributes to organismal virulence. It is noteworthy that the gene expression of a NRPS gene of G. boninense strain UPMGB001 (belonging to the type VI siderophore-synthesizing NRPS family) was previously reported to be related to the basal stem rot disease severity of oil palm [70]. In addition, the bZIP transcription factor hapX, which represses genes related to iron-dependent and mitochondrial-localized activities during iron starvation [57], was identified in all Ganoderma species/strains. This transcription factor is critical for the coordination of the production of siderophores and their precursor ornithine [57] to ensure efficient iron uptake and inhibition of iron-consuming pathways [71]. Yasmin et al. [55] demonstrated that the interdependency of ergosterol and siderophore biosynthesis in Aspergillus fumigatus was governed by mevalonate (which is required for both biosynthetic pathways) through the acyl-CoA ligase SidI and the enoyl-CoA hydratase SidH. Iron-starvation induced the production of siderophore and decreased the ergosterol content and composition, which are critical for virulence. The biosynthetic core gene for β-lactone was identified in six of the Ganoderma species/strains analyzed in this study. It is a potent antibacterial and antifungal compound [72] which has been reported to inhibit the growth of fungi. β-lactone interferes with the biosynthesis of ergosterol by down-regulating the expression of HMG-CoA synthase [73]. The significance of bassidioferrin and β-lactone to the fungal virulence is unknown.
A few enzymes in the ergosterol biosynthesis pathway have been reported to be essential for fungal viability [74,75,76,77], vegetative differentiation, and virulence [78]. Ortholog clusters related to ergosterol biosynthesis and sterol transport were present in all the Ganoderma species/strains analyzed. The PRYs are sterol-binding and -export proteins belonging to the CAP (cysteine-rich secretory protein, antigen 5, and pathogenesis-related 1) protein family [79]. Fungal pathogens sequester sterol from the host tissue through PRYs, which facilitate tissue penetration [79]. The PRY2 proteins are secreted glycoproteins while the PRY3 proteins are known to attach to glycosylphosphatidylinositol at the fungal cell wall [80,81]. PRY2 can export acetylated cholesterol [82] and was found to be hypersensitive to plant eugenol oil [81]. Although pathogen-specific PRYs may display distinct ligand-binding properties and/or protein stabilities [79], it is difficult to predict the pathogenicity of these putative PRYs solely based on the number of conserved cysteine residues involved in intramolecular disulfide bridges without evidence from laboratory experiments. Nevertheless, one of the PRY2 orthologs from G. boninense strain G3 was found to be highly similar to a sequence from G. boninense PER71, which was found to be expressed during oil palm root infection [20].
The infection mechanisms shared by the Ganoderma species/strains with other fungal pathogens can be inferred from the presence of annotated pathogenesis-related protein orthologs in their proteomes. The fungal infection process extends from germination or multiplication of an infective apparatus at the infection site to the establishment of a biotrophic or necrotrophic relationship with its host [83]. The proteins involved may include the global transcriptional regulator sge1, which participates in conidiogenesis and regulates the expression of fungal effectors during infection [56], and the C2H2 finger domain transcription factor dvrA, which may participate in the interactions between pathogen and host and regulate virulence negatively by changing the damaging capacity of a pathogen to host cells [58]. In addition, proteases (such as extracellular metalloproteinase, metallocarboxypeptidase A-like protein, aspartic-type endopeptidase, leucine aminopeptidase, and tripeptidyl-peptidases proteases) may facilitate the degradation of host proteins by these Ganoderma species/strains [61], while calcineurin subunit B (a calcium-dependent, calmodulin stimulated protein phosphatase) could play a central role in virulence and antifungal drug action [59] in Ganoderma species/strains. The presence of a glycerophosphodiester transporter in Ganoderma species/strains may mediate the uptake of glycerophosphoinositol as a source of inositol and phosphate [84,85] and expand the ability of fungi to utilize glycerophosphoinositol, hence enabling them to occupy a variety of environments within their host organisms [84]. Meanwhile, the LysM domain-containing protein ARB_00327 might protect the fungal hyphae from plant chitinases [86] and suppress host defenses by sequestering chitin oligosaccharides that trigger host immunity [60].
The current study predicted and compared putative fungal effectors from different Ganoderma species/strains including species that are pathogenic and non-pathogenic to oil palms. The approach herein is different from the approach employed by Khairi et al. [37], who predicted effector proteins from the pan genomes of G. boninense. Few predicted effectors were conserved in all seven species, and many putative effectors are unique to only one Ganoderma species/strain. These putative effectors are possibly responsible for host specificity and specific host–pathogen interactions. About 41% of the predicted effectors from G. boninense strain G3 were found to be expressed in G. boninense PER71 during oil palm infection [20]. These groups of unique fungal effectors may have more potential to be associated with basal stem rot disease in oil palm. Some of these putative effectors were also found to be expressed in planta by a separate study [37] using different RNAseq datasets. Among these unique effectors are putative allergen Asp F15, thaumatin-like protein 1, PRY2, protein rot1, cytochrome P450 monooxygenase FSL4, cytochrome P450 734A, thioredoxin-like protein AAED1, and hydrophobin. Asp F15 is an allergen which belongs to the cerato-platanin family. The cerato-platanin protein could also be a phytotoxin which causes plant disease [87], and it was reported to induce cell necrosis and phytoalexin synthesis in plants [88]. Moreover, hydrophobin and thaumatin-like protein were reported to be involved in the development and senescence of the fruiting body, respectively [89,90]. In addition, thaumatin-like protein may have β-1,3-glucanase activities which can modify fungal cell walls [91]. Although fusarielins exhibit antibacterial, antifungal, anticancer, anti-angiogenic, and antiproliferative properties [92], the role of these polyketides in plant pathogenesis is not clear. Since most of these candidate effectors were found to be present only in oil palm roots treated with G. boninense but not in oil palm root samples treated with Trichoderma, these candidate effectors could serve as targets for the development of antifungal agents that are specific to the pathogen. The roles of these candidate effectors in oil palm infection await further elucidation.
5. Conclusions
The comparative genomics analyses of the proteomes of Ganoderma species/strains in this study revealed that: 1. G. boninense is likely a hemibiotroph based on its CAZyme content (in support of its aggressiveness as a potent pathogen to oil palm) while all the other Ganoderma species/strains being analyzed were predicted to adopt a saprophytic or a symbiotic lifestyle; 2. Ganoderma species/strains shared many CAZymes and secondary metabolite core gene clusters, in line with their ability to degrade plant cell walls and to produce high values secondary metabolites; however, there are also unique CAZyme classes and secondary metabolite core genes that are unique in individual species; 3. Protein orthologs that are related to pathogenesis including those related to mycotoxin and secondary metabolite biosynthesis, ergosterol biosynthesis and transport, as well as virulence factors were identified for further functional experiments; 4. Potential effectors were also identified for Ganoderma species/strains, especially unique effectors that were expressed in G. boninense in oil palm roots, causing basal stem rot disease. These are useful resources for future analyses of the fungal pathogenesis of Ganoderma species.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f14030653/s1, Table S1: Information of the Ganoderma species and genome assemblies.
Funding
This project was funded by Geran Putra Berimpak (Project No. UPM/7002/1/GPB/2018) from the Universiti Putra Malaysia.
Data Availability Statement
The datasets generated during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The author declares no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Baby, S.; Johnson, A.J.; Govindan, B. Secondary metabolites from Ganoderma. Phytochem. 2015, 114, 66–101. [Google Scholar] [CrossRef]
- Mohammed, C.L.; Rimbawanto, A.; Page, D.E. Management of basidiomycete root- and stem-rot diseases in oil palm, rubber and tropical hardwood plantation crops. For. Path. 2014, 44, 428–446. [Google Scholar] [CrossRef]
- Kandan, A.; Bhaskaran, R.; Samiyappan, R. Ganoderma—A basal stem rot disease of coconut palm in south Asia and Asia pacific regions. Arch. Phytopathol. Pflanzenschutz. 2010, 43, 1445–1449. [Google Scholar] [CrossRef]
- Loyd, A.L.; Linder, E.R.; Anger, N.A.; Richter, B.S.; Blanchette, R.A.; Smith, J.A. Pathogenicity of Ganoderma species on landscape trees in the Southeastern United States. Plant Dis. 2018, 102, 1944–1949. [Google Scholar] [CrossRef] [Green Version]
- Bishop, K.S.; Kao, C.H.; Xu, Y.; Glucina, M.P.; Paterson, R.R.; Ferguson, L.R. From 2000 years of Ganoderma lucidum to recent developments in nutraceuticals. Phytochemistry 2015, 114, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, I.C.; Heleno, S.A.; Reis, F.S.; Stojkovic, D.; Queiroz, M.J.; Vasconcelos, M.H.; Sokovic, M. Chemical features of Ganoderma polysaccharides with antioxidant, antitumor and antimicrobial activities. Phytochemistry 2015, 114, 38–55. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Liu, J.; Xia, J.; Wang, C.; Li, X.; Deng, Y.; Bao, N.; Zhang, Z.; Qiu, M. Lanostane triterpenoids from Ganoderma hainanense J. D. Zhao. Phytochemistry 2015, 114, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiang, Y.; Zhang, M.; Zhang, L. Ganoderma sinense polysaccharide: An adjunctive drug used for cancer treatment. Prog. Mol. Biol. Transl. Sci. 2019, 163, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Tian, D.; Liu, Y.; Li, H.; Zhu, J.; Li, M.; Xin, M.; Xia, J. Review of the molecular mechanisms of Ganoderma lucidum triterpenoids: Ganoderic acids A, C2, D, F, DM, X and Y. Eur. J. Med. Chem. 2019, 174, 130–141. [Google Scholar] [CrossRef]
- Pattanayak, S.; Das, S.; Biswal, G. Ganoderma: The wild mushroom with wonderful health benefits. J. Pharmacogn. Phytochem. 2020, 9, 313–316. [Google Scholar] [CrossRef]
- Jiang, N.; Hu, S.; Peng, B.; Li, Z.; Yuan, X.; Xiao, S.; Fu, Y. Genome of Ganoderma species provides Insights Into the evolution, conifers substrate utilization, and terpene synthesis for Ganoderma tsugae. Front. Microbiol. 2021, 12, 724451. [Google Scholar] [CrossRef]
- Tian, Y.Z.; Wang, Z.F.; Liu, Y.D.; Zhang, G.Z.; Li, G. The whole-genome sequencing and analysis of a Ganoderma lucidum strain provide insights into the genetic basis of its high triterpene content. Genomics 2021, 113, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.F.; Lebreton, A.; Xing, J.H.; Fang, Y.X.; Si, J.; Morin, E.; Miyauchi, S.; Drula, E.; Ahrendt, S.; Cobaugh, K.; et al. Phylogenomics and Comparative Genomics Highlight Specific Genetic Features in Ganoderma Species. J. Fungi 2022, 8, 311. [Google Scholar] [CrossRef] [PubMed]
- Kües, U.; Nelson, D.R.; Liu, C.; Yu, G.J.; Zhang, J.; Li, J.; Wang, X.C.; Sun, H. Genome analysis of medicinal Ganoderma spp. with plant-pathogenic and saprotrophic life-styles. Phytochemistry 2015, 114, 18–37. [Google Scholar] [CrossRef]
- Chen, S.; Xu, J.; Liu, C.; Zhu, Y.; Nelson, D.R.; Zhou, S.; Li, C.; Wang, L.; Guo, X.; Sun, Y.; et al. Genome sequence of the model medicinal mushroom Ganoderma lucidum. Nat. Commun. 2012, 3, 913. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.J.; Wang, M.; Huang, J.; Yin, Y.L.; Chen, Y.J.; Jiang, S.; Jin, Y.X.; Lan, X.Q.; Wong, B.H.; Liang, Y.; et al. Deep insight into the Ganoderma lucidum by comprehensive analysis of its transcriptome. PLoS ONE 2012, 7, e44031. [Google Scholar] [CrossRef]
- Huang, Y.H.; Wu, H.Y.; Wu, K.M.; Liu, T.T.; Liou, R.F.; Tsai, S.F.; Shiao, M.S.; Ho, L.T.; Tzean, S.S.; Yang, U.C. Generation and analysis of the expressed sequence tags from the mycelium of Ganoderma lucidum. PLoS ONE 2013, 8, e61127. [Google Scholar] [CrossRef] [Green Version]
- Ramzi, A.B.; Che Me, M.L.; Ruslan, U.S.; Baharum, S.N.; Nor Muhammad, N.A. Insight into plant cell wall degradation and pathogenesis of Ganoderma boninense via comparative genome analysis. PeerJ 2019, 7, e8065. [Google Scholar] [CrossRef] [Green Version]
- Ho, P.Y.; Namasivayam, P.; Sundram, S.; Ho, C.L. Expression of genes encoding manganese peroxidase and laccase of Ganoderma boninense in response to nitrogen sources, hydrogen peroxide and phytohormones. Genes 2020, 11, 1263. [Google Scholar] [CrossRef]
- Ho, C.L.; Tan, Y.C.; Yeoh, K.A.; Ghazali, A.K.; Yee, W.Y.; Hoh, C.C. De novo transcriptome analyses of host-fungal interactions in oil palm (Elaeis guineensis Jacq.). BMC Genom. 2016, 17, 66. [Google Scholar] [CrossRef] [Green Version]
- Ko, E.M.; Leem, Y.E.; Choi, H.T. Purification and characterization of laccase isozymes from the white-rot basidiomycete Ganoderma lucidum. Appl. Microbiol. Biotechnol. 2001, 57, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Teerapatsakul, C.; Abe, N.; Bucke, C.; Chitradon, L. Novel laccases of Ganoderma sp. KU-Alk4, regulated by different glucose concentration in alkaline media. World J. Microbiol. Biotechnol. 2007, 23, 1559–1567. [Google Scholar] [CrossRef]
- Kumar, A.; Kant, K.; Kumar., P.; Ramchiary., N. Laccase isozymes from Ganoderma lucidum MDU-7: Isolation, characterization, catalytic properties and differential role during oxidative stress. J. Mol. Catal. Enzymat. 2015, 113, 68–75. [Google Scholar] [CrossRef]
- Torres-Farradá, G.; Manzano León, A.M.; Rineau, F.; Ledo Alonso, L.L.; Sánchez-López, M.I.; Thijs, S.; Colpaert, J.; Ramos-Leal, M.; Guerra, G.; Vangronsveld, J. Diversity of ligninolytic enzymes and their genes in strains of the genus Ganoderma: Applicable for biodegradation of xenobiotic compounds? Front. Microbiol. 2017, 8, 898. [Google Scholar] [CrossRef] [Green Version]
- Si, J.; Wu, Y.; Ma, H.F.; Cao, Y.J.; Sun, Y.F.; Cui, B.K. Selection of a pH- and temperature-stable laccase from Ganoderma australe and its application for bioremediation of textile dyes. J. Environ. Manag. 2021, 299, 113619. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Guo, M.Y.; Gao, Y.H.; Bai, X.H.; Zhou, X.W. Expression and characteristics of manganese peroxidase from Ganoderma lucidum in Pichia pastoris and its application in the degradation of four dyes and phenol. BMC Biotechnol. 2017, 17, 19. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, R.; Ma, L.; Fan, F.; Gong, Y.; Wan, X.; Jiang, M.; Zhang, X.; Yang, Y. Decolorization of different dyes by a newly isolated white-rot fungi strain Ganoderma sp. En3 and cloning and functional analysis of its laccase gene. J. Hazard. Mater. 2011, 192, 855–873. [Google Scholar] [CrossRef]
- You, L.F.; Liu, Z.M.; Lin, J.F.; Guo, L.Q.; Huang, X.L.; Yang, H.X. Molecular cloning of a laccase gene from Ganoderma lucidum and heterologous expression in Pichia pastoris. J. Basic Microbiol. 2014, 54, S134–S141. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Deng, W.; Shen, M.; Yan, G.; Zhao, W.; Yang, Y. A laccase Gl-LAC-4 purified from white-rot fungus Ganoderma lucidum had a strong ability to degrade and detoxify the alkylphenol pollutants 4-n-octylphenol and 2-phenylphenol. J. Hazard. Mater. 2021, 408, 124775. [Google Scholar] [CrossRef] [PubMed]
- Teh, C.Y.; Pang, C.; Tor, X.Y.; Ho, P.; Lim, Y.; Namasivayam, P.; Ho, C.L. Molecular cloning and functional analysis of a necrosis and ethylene inducing protein (NEP) from Ganoderma boninense. Physiol. Mol. Plant Pathol. 2019, 106, 42–48. [Google Scholar] [CrossRef]
- Lim, F.-H.; Fakhrana, I.N.; Rasid, O.A.; Idris, A.S.; Ho, C.-L.; Shaharuddin, N.A.; Parveez, G.K.A. Molecular cloning and expression analysis of Ganoderma boninense cyclophilins at different growth and infection stages. Physiol. Mol. Plant Pathol. 2017, 99, 31–40. [Google Scholar] [CrossRef]
- Sulaiman, S.; Yusoff, N.; Tan, J.S.; Lee, Y.P. Deciphering the pan-genome of Ganoderma sp. to depict potential genomic components that contribute to Ganoderma boninense pathogenicity. Malays. Appl. Biol. 2018, 47, 71–80. [Google Scholar]
- Zhu, Y.; Xu, J.; Sun, C.; Zhou, S.; Xu, H.; Nelson, D.R.; Qian, J.; Song, J.; Luo, H.; Xiang, L.; et al. Chromosome-level genome map provides insights into diverse defense mechanisms in the medicinal fungus Ganoderma sinense. Sci. Rep. 2015, 5, 11087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Huang, L.; Hu, H.; Cai, M.; Liang, X.; Li, X.; Zhang, Z.; Xie, Y.; Xiao, C.; Chen, S.; et al. Whole-genome assembly of Ganoderma leucocontextum (Ganodermataceae, Fungi) discovered from the Tibetan Plateau of China. G3 2021, 11, jkab337. [Google Scholar] [CrossRef] [PubMed]
- Binder, M.; Justo, A.; Riley, R.; Salamov, A.; Lopez-Giraldez, F.; Sjökvist, E.; Copeland, A.; Foster, B.; Sun, H.; Larsson, E.; et al. Phylogenetic and phylogenomic overview of the Polyporales. Mycologia 2013, 105, 1350–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Gong, J.; Dai, W.; Kang, X.; Huang, Z.; Zhang, H.M.; Liu, W.; Liu, L.; Ma, J.; Xia, Z.; et al. The genome of Ganoderma lucidum provides insights into triterpenes biosynthesis and wood degradation. PLoS ONE 2012, 7, e36146. [Google Scholar] [CrossRef] [Green Version]
- Khairi, M.H.F.; Nor Muhammad, N.A.; Bunawan, H.; Abdul Murad, A.M.; Ramzi, A.B. Unveiling the core effector proteins of oil palm pathogen Ganoderma boninense via pan-secretome analysis. J. Fungi 2022, 8, 793. [Google Scholar] [CrossRef]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness. Methods Mol. Biol. 2019, 1962, 227–245. [Google Scholar] [CrossRef]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Eddy, S.R. A new generation of homology search tools based on probabilistic inference. Genome Inform. 2009, 23, 205–211. [Google Scholar]
- Yin, Y.; Mao, X.; Yang, J.; Chen, X.; Mao, F.; Xu, Y. dbCAN: A web resource for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2012, 40, W445–W451. [Google Scholar] [CrossRef] [PubMed]
- Hane, J.K.; Paxman, J.; Jones, D.A.B.; Oliver, R.P.; de Wit, P. “CATAStrophy”, a genome-informed trophic classification of filamentous plant pathogens—How many different types of filamentous plant pathogens are there? Front. Microbiol. 2020, 10, 3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Stoeckert, C.J., Jr.; Roos, D.S. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef] [Green Version]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Käll, L.; Krogh, A.; Sonnhammer, E.L. A combined transmembrane topology and signal peptide prediction method. J. Mol. Biol. 2004, 338, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Singh, K.B.; Taylor, J.M. Improved prediction of fungal effector proteins from secretomes with EffectorP 2.0. Mol. Plant Path. 2018, 19, 2094–2110. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Wu, S.; Su, C.; Peng, J.; Shih, Y.; Chen, L. Ganoderma multipileum, the correct name for ‘G. lucidum’ in tropical Asia. Bot. Stud. 2009, 50, 451–458. [Google Scholar]
- Kajikawa, M.; Hirai, N.; Hashimoto, T. A PIP-family protein is required for biosynthesis of tobacco alkaloids. Plant Mol. Biol. 2009, 69, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Bodega, Á.; Álvarez-Álvarez, R.; Liras, P.; Martín, J.F. Silencing of a second dimethylallyltryptophan synthase of Penicillium roqueforti reveals a novel clavine alkaloid gene cluster. Appl. Microbiol. Biotechnol. 2017, 101, 6111–6121. [Google Scholar] [CrossRef] [PubMed]
- Eichhorn, H.; Lessing, F.; Winterberg, B.; Schirawski, J.; Kämper, J.; Müller, P.; Kahmann, R. A ferroxidation/permeation iron uptake system is required for virulence in Ustilago maydis. Plant Cell 2006, 18, 3332–3345. [Google Scholar] [CrossRef] [Green Version]
- Winterberg, B.; Uhlmann, S.; Linne, U.; Lessing, F.; Marahiel, M.A.; Eichhorn, H.; Kahmann, R.; Schirawski, J. Elucidation of the complete ferrichrome A biosynthetic pathway in Ustilago maydis. Mol. Microbiol. 2010, 75, 1260–1271. [Google Scholar] [CrossRef]
- Yasmin, S.; Alcazar-Fuoli, L.; Gründlinger, M.; Puempel, T.; Cairns, T.; Blatzer, M.; Lopez, J.F.; Grimalt, J.O.; Bignell, E.; Haas, H. Mevalonate governs interdependency of ergosterol and siderophore biosyntheses in the fungal pathogen Aspergillus fumigatus. Proc. Natl. Acad. Sci. USA 2012, 109, E497–E504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michielse, C.B.; van Wijk, R.; Reijnen, L.; Manders, E.M.; Boas, S.; Olivain, C.; Alabouvette, C.; Rep, M. The nuclear protein Sge1 of Fusarium oxysporum is required for parasitic growth. PLoS Pathog. 2009, 5, e1000637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Gravelat, F.N.; Chiang, L.Y.; Chen, D.; Vanier, G.; Ejzykowicz, D.E.; Ibrahim, A.S.; Nierman, W.C.; Sheppard, D.C.; Filler, S.G. Aspergillus fumigatus AcuM regulates both iron acquisition and gluconeogenesis. Mol. Microbiol. 2010, 78, 1038–1054. [Google Scholar] [CrossRef] [Green Version]
- Ejzykowicz, D.E.; Solis, N.V.; Gravelat, F.N.; Chabot, J.; Li, X.; Sheppard, D.C.; Filler, S.G. Role of Aspergillus fumigatus DvrA in host cell interactions and virulence. Eukaryot Cell 2010, 9, 1432–1440. [Google Scholar] [CrossRef] [Green Version]
- Fox, D.S.; Cruz, M.C.; Sia, R.A.; Ke, H.; Cox, G.M.; Cardenas, M.E.; Heitman, J. Calcineurin regulatory subunit is essential for virulence and mediates interactions with FKBP12-FK506 in Cryptococcus neoformans. Mol. Microbiol. 2001, 39, 835–849. [Google Scholar] [CrossRef]
- De Jonge, R.; Thomma, B.P. Fungal LysM effectors: Extinguishers of host immunity? Trends Microbiol. 2009, 17, 151–157. [Google Scholar] [CrossRef]
- Krishnan, P.; Ma, X.; McDonald, B.A.; Brunner, P.C. Widespread signatures of selection for secreted peptidases in a fungal plant pathogen. BMC Evol. Biol. 2018, 18, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, Y.; Surendran, A.; Paterson, R.R.M.; Ali, A.; Ahmad, K. Current strategies and perspectives in detection and control of basal stem rot of oil palm. Saudi J. Biol. 2021, 28, 2840–2849. [Google Scholar] [CrossRef] [PubMed]
- Vinjusha, N.; Arun Kumar, T.K. Revision of Ganoderma species associated with stem rot of coconut palm. Mycologia 2022, 114, 157–174. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Qiao, X.; Shao, Q.; Hassan, M.; Ma, Z. Evolution of the lignin chemical structure during the bioethanol production process and its inhibition to enzymatic hydrolysis. Energy Fuels 2020, 34, 5938–5947. [Google Scholar] [CrossRef]
- Jayasuriya, H.; Silverman, K.C.; Zink, D.L.; Jenkins, R.G.; Sanchez, M.; Pelaez, F.; Vilella, D.; Lingham, R.B.; Singh, S.B. Clavaric acid: A triterpenoid inhibitor of farnesyl-protein transferase from Clavariadelphus truncatus. J. Nat. Prod. 1998, 61, 1568–1570. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Hohn, T.M. Mycotoxins in plant pathogenesis. Mol. Plant Microbe Interact. 1997, 10, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Panaccione, D.G.; Scott-Craig, J.S.; Pocard, J.A.; Walton, J.D. A cyclic peptide synthetase gene required for pathogenicity of the fungus Cochliobolus carbonum on maize. Proc. Natl. Acad. Sci. USA 1992, 89, 6590–6594. [Google Scholar] [CrossRef] [Green Version]
- Baker, S.E.; Kroken, S.; Inderbitzin, P.; Asvarak, T.; Li, B.Y.; Shi, L.; Yoder, O.C.; Turgeon, B.G. Two polyketide synthase-encoding genes are required for biosynthesis of the polyketide virulence factor, T-toxin, by Cochliobolus heterostrophus. Mol. Plant Microbe Interact. 2006, 19, 139–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koczyk, G.; Pawłowska, J.; Muszewska, A. Terpenoid biosynthesis dominates among secondary metabolite clusters in Mucoromycotina genomes. J. Fungi 2021, 7, 285. [Google Scholar] [CrossRef] [PubMed]
- Shokrollahi, N.; Ho, C.L.; Zainudin, N.A.I.M.; Wahab, M.A.B.A.; Wong, M.Y. Identification of non-ribosomal peptide synthetase in Ganoderma boninense Pat. that was expressed during the interaction with oil palm. Sci. Rep. 2021, 11, 16330. [Google Scholar] [CrossRef]
- Kröber, A.; Scherlach, K.; Hortschansky, P.; Shelest, E.; Staib, P.; Kniemeyer, O.; Brakhage, A.A. HapX mediates iron homeostasis in the pathogenic dermatophyte Arthroderma benhamiae but is dispensable for virulence. PLoS ONE 2016, 11, e0150701. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.L.; Christenson, J.K.; Wackett, L.P. Biosynthesis and chemical diversity of β-lactone natural products. Nat. Prod. Rep. 2019, 36, 458–475. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.X.; Yan, X.; Fu, W.H.; Yi, L.Q.; Tang, B.W.; Yu, L.B.; Fang, M.J.; Wu, Z.; Qiu, Y.K. New β-lactone with tea pathogenic fungus inhibitory effect from marine-derived fungus MCCC3A00957. J. Agric. Food Chem. 2019, 67, 2877–2885. [Google Scholar] [CrossRef] [PubMed]
- Lees, N.D.; Skaggs, B.; Kirsch, D.R.; Bard, M. Cloning of the late genes in the ergosterol biosynthetic pathway of Saccharomyces cerevisiae—A review. Lipids 1995, 30, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Aaron, K.E.; Pierson, C.A.; Lees, N.D.; Bard, M. The Candida albicans ERG26 gene encoding the C-3 sterol dehydrogenase (C-4 decarboxylase) is essential for growth. FEMS Yeast Res. 2001, 1, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, M.A.; Johnson, T.A.; Lees, N.D.; Barbuch, R.; Eckstein, J.A.; Bard, M. Cloning and sequencing of the Candida albicans C-4 sterol methyl oxidase gene (ERG25) and expression of an ERG25 conditional lethal mutation in Saccharomyces cerevisiae. Lipids 2000, 35, 257–262. [Google Scholar] [CrossRef]
- Pierson, C.A.; Jia, N.; Mo, C.; Lees, N.D.; Sturm, A.M.; Eckstein, J.; Barbuct, R.; Bard, M. Isolation, characterization, and regulation of the Candida albicans ERG27 gene encoding the sterol 3-keto reductase. Med. Mycol. 2004, 42, 461–473. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Jian, Y.; Chen, Y.; Kistler, H.C.; He, P.; Ma, Z.; Yin, Y. A phosphorylated transcription factor regulates sterol biosynthesis in Fusarium graminearum. Nat. Commun. 2019, 10, 1228. [Google Scholar] [CrossRef] [Green Version]
- Schneiter, R.; Di Pietro, A. The CAP protein superfamily: Function in sterol export and fungal virulence. Biomol. Concepts 2013, 4, 519–525. [Google Scholar] [CrossRef]
- Yin, Q.Y.; de Groot, P.W.; Dekker, H.L.; de Jong, L.; Klis, F.M.; de Koster, C.G. Comprehensive proteomic analysis of Saccharomyces cerevisiae cell walls: Identification of proteins covalently attached via glycosylphosphatidylinositol remnants or mild alkali-sensitive linkages. J. Biol. Chem. 2005, 280, 20894–20901. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, V.; Schneiter, R. Pathogen-Related Yeast (PRY) proteins and members of the CAP superfamily are secreted sterol-binding proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 16882–16887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, R.; Köffel, R.; Schneiter, R. An acetylation/deacetylation cycle controls the export of sterols and steroids from S. cerevisiae. EMBO J. 2007, 26, 5109–5119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravichandra, N.G. Frontiers in Phytopathology; I.K. International Publishing House Pvt Ltd.: New Delhi, India, 2016. [Google Scholar]
- Bishop, A.C.; Sun, T.; Johnson, M.E.; Bruno, V.M.; Patton-Vogt, J. Robust utilization of phospholipase-generated metabolites, glycerophosphodiesters, by Candida albicans: Role of the CaGit1 permease. Eukaryot. Cell 2011, 10, 1618–1627. [Google Scholar] [CrossRef] [Green Version]
- Bishop, A.C.; Ganguly, S.; Solis, N.V.; Cooley, B.M.; Jensen-Seaman, M.I.; Filler, S.G.; Mitchell, A.P.; Patton-Vogt, J. Glycerophosphocholine utilization by Candida albicans: Role of the Git3 transporter in virulence. J. Biol. Chem. 2013, 288, 33939–33952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Vallet, A.; Tian, H.; Rodriguez-Moreno, L.; Valkenburg, D.J.; Saleem-Batcha, R.; Wawra, S.; Kombrink, A.; Verhage, L.; de Jonge, R.; van Esse, H.P.; et al. A secreted LysM effector protects fungal hyphae through chitin-dependent homodimer polymerization. PLoS Pathog. 2020, 16, e1008652. [Google Scholar] [CrossRef]
- Pazzagli, L.; Cappugi, G.; Manao, G.; Camici, G.; Santini, A.; Scala, A. Purification, characterization, and amino acid sequence of cerato-platanin, a new phytotoxic protein from Ceratocystis fimbriata f. sp. platani. J. Biol. Chem. 1999, 274, 24959–24964. [Google Scholar] [CrossRef] [Green Version]
- Sbrana, F.; Bongini, L.; Cappugi, G.; Fanelli, D.; Guarino, A.; Pazzagli, L.; Scala, A.; Vassalli, M.; Zoppi, C.; Tiribilli, B. Atomic force microscopy images suggest aggregation mechanism in cerato-platanin. Eur. Biophys. J. 2007, 36, 727–732. [Google Scholar] [CrossRef] [Green Version]
- Cai, F.; Gao, R.; Zhao, Z.; Ding, M.; Jiang, S.; Yagtu, C.; Zhu, H.; Zhang, J.; Ebner, T.; Mayrhofer-Reinhartshuber, M.; et al. Evolutionary compromises in fungal fitness: Hydrophobins can hinder the adverse dispersal of conidiospores and challenge their survival. ISME J. 2000, 14, 2610–2624. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Watanabe, H.; Nagai, M.; Nakade, K.; Takahashi, M.; Sato, T. Lentinula edodes tlg1 encodes a thaumatin-like protein that is involved in lentinan degradation and fruiting body senescence. Plant Physiol. 2006, 141, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Grenier, J.; Potvin, C.; Trudel, J.; Asselin, A. Some thaumatin-like proteins hydrolyse polymeric beta-1,3-glucans. Plant J. 1999, 19, 473–480. [Google Scholar] [CrossRef]
- Fujimoto, H.; Aoyama, H.; Noguchi-Yachide, T.; Hashimoto, Y.; Kobayashi, H. Fusarielin A as an anti-angiogenic and anti-proliferative agent: Basic biological characterization. Chem. Pharm. Bull. 2008, 56, 298–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Carbohydrate-active enzyme (CAZyme) content among Ganoderma species/strains. (A), Number of CAZymes of each species/strain according to CAZyme superfamily: glycoside hydrolase (GH), glycosyl transferase (GT), auxiliary activity (AA), carbohydrate esterase (CE), polysaccharide lyase activity (PL), and carbohydrate-binding modules (CBM); (B), Percentage of CAZymes of each species/strain according to CAZyme superfamily; (C), Principal-component analysis (PCA) plot of fungal lifestyle predictions as inferred by CATAStrophy. PC1, principal component 1, and PC2, principal component 2. The plot shows the CAZyme-inferred phenotypic trophisms of 85 reference species of fungi with different lifestyles (i.e., 14 biotrophs, 20 hemibiotrophs, 25 nectrotrophs, 13 saprotrophs, and 36 symbionts in a training set used by CATAStrophy) and Ganoderma species/strains. Gb_G3, G. boninense strain G3; Gb_NJ3, G. boninense strain NJ3; Gm, G. multipileum; Gl_G, G. lucidum strain G260125-1; Gl_ BCRC36111, G. lucidum strain BCRC36111; Gl_ BCRC37177, G. lucidum strain BCRC37177; Gl_XN, G. lucidum strain Xiangnong; Gs, G. sinense; Gt, G. tsugae; G_BRIUMSc, Ganoderma sp. BRIUMSc. The groupings of biotrophs, hemibiotrophs, nectrotrophs, saprotrophs, and symbionts are shown by orange, blue, green, purple, and yellow circles, respectively.
Figure 1.
Carbohydrate-active enzyme (CAZyme) content among Ganoderma species/strains. (A), Number of CAZymes of each species/strain according to CAZyme superfamily: glycoside hydrolase (GH), glycosyl transferase (GT), auxiliary activity (AA), carbohydrate esterase (CE), polysaccharide lyase activity (PL), and carbohydrate-binding modules (CBM); (B), Percentage of CAZymes of each species/strain according to CAZyme superfamily; (C), Principal-component analysis (PCA) plot of fungal lifestyle predictions as inferred by CATAStrophy. PC1, principal component 1, and PC2, principal component 2. The plot shows the CAZyme-inferred phenotypic trophisms of 85 reference species of fungi with different lifestyles (i.e., 14 biotrophs, 20 hemibiotrophs, 25 nectrotrophs, 13 saprotrophs, and 36 symbionts in a training set used by CATAStrophy) and Ganoderma species/strains. Gb_G3, G. boninense strain G3; Gb_NJ3, G. boninense strain NJ3; Gm, G. multipileum; Gl_G, G. lucidum strain G260125-1; Gl_ BCRC36111, G. lucidum strain BCRC36111; Gl_ BCRC37177, G. lucidum strain BCRC37177; Gl_XN, G. lucidum strain Xiangnong; Gs, G. sinense; Gt, G. tsugae; G_BRIUMSc, Ganoderma sp. BRIUMSc. The groupings of biotrophs, hemibiotrophs, nectrotrophs, saprotrophs, and symbionts are shown by orange, blue, green, purple, and yellow circles, respectively.


Figure 2.
Counts of putative biosynthetic core gene clusters (BGCs) of Ganoderma species/strains identified by antiSMASH. The most similar known clusters were shown on top of the BGCs they belong: B, basidioferrin; and C, clavaric acid. Gb_G3, G. boninense strain G3; Gb_NJ3, G. boninense strain NJ3; Gm, G. multipileum; Gl_G, G. lucidum strain G260125-1; Gl_ BCRC36111, G. lucidum strain BCRC36111; Gl_BCRC37177, G. lucidum strain BCRC37177; Gl_XN, G. lucidum strain Xiangnong; Gs, G. sinense; Gt, G. tsugae; G_BRIUMSc, Ganoderma sp. BRIUMSc.
Figure 2.
Counts of putative biosynthetic core gene clusters (BGCs) of Ganoderma species/strains identified by antiSMASH. The most similar known clusters were shown on top of the BGCs they belong: B, basidioferrin; and C, clavaric acid. Gb_G3, G. boninense strain G3; Gb_NJ3, G. boninense strain NJ3; Gm, G. multipileum; Gl_G, G. lucidum strain G260125-1; Gl_ BCRC36111, G. lucidum strain BCRC36111; Gl_BCRC37177, G. lucidum strain BCRC37177; Gl_XN, G. lucidum strain Xiangnong; Gs, G. sinense; Gt, G. tsugae; G_BRIUMSc, Ganoderma sp. BRIUMSc.

Figure 3.
Pairwise proteome comparisons and Venn diagrams of ortholog clusters shared by Ganoderma species/strains. (A), similarity matrix of pairwise proteome comparisons. The numbers in the heat map show the number of ortholog clusters between a pair of proteomes; (B), the number of ortholog clusters shared by five representatives from different Ganoderma species; (C), the number of ortholog clusters shared by three G. lucidum strains; (D), the number of ortholog clusters shared by G. lucidum strain BCRC37177 and G. multipileum. Gb_G3, G. boninense strain G3; Gm, G. multipileum; Gl_G, G. lucidum strain G260125-1; Gl_ BCRC37177, G. lucidum strain BCRC37177; Gl_XN, G. lucidum strain Xiangnong; Gs, G. sinense; Gt, G. tsugae.
Figure 3.
Pairwise proteome comparisons and Venn diagrams of ortholog clusters shared by Ganoderma species/strains. (A), similarity matrix of pairwise proteome comparisons. The numbers in the heat map show the number of ortholog clusters between a pair of proteomes; (B), the number of ortholog clusters shared by five representatives from different Ganoderma species; (C), the number of ortholog clusters shared by three G. lucidum strains; (D), the number of ortholog clusters shared by G. lucidum strain BCRC37177 and G. multipileum. Gb_G3, G. boninense strain G3; Gm, G. multipileum; Gl_G, G. lucidum strain G260125-1; Gl_ BCRC37177, G. lucidum strain BCRC37177; Gl_XN, G. lucidum strain Xiangnong; Gs, G. sinense; Gt, G. tsugae.

Figure 4.
Predicted effectors in Ganoderma species/strains. (A), Venn diagram of effector orthologs of shared by Ganoderma species/strains. The numbers in black show the number of ortholog clusters shared by at least two Ganoderma species/strains while the numbers in red show the number of effectors that are unique to individual Ganoderma species/strains. Gb_G3, G. boninense strain G3; Gm, G. multipileum; Gl_G, G. lucidum strain G260125-1; Gl_XN, G. lucidum strain Xiangnong; Gs, G. sinense; Gt, G. tsugae. (B–D), GO classification of effectors that are unique to Gb_G3 in biological process (BP), molecular function (MF) and cellular component (CC), respectively.
Figure 4.
Predicted effectors in Ganoderma species/strains. (A), Venn diagram of effector orthologs of shared by Ganoderma species/strains. The numbers in black show the number of ortholog clusters shared by at least two Ganoderma species/strains while the numbers in red show the number of effectors that are unique to individual Ganoderma species/strains. Gb_G3, G. boninense strain G3; Gm, G. multipileum; Gl_G, G. lucidum strain G260125-1; Gl_XN, G. lucidum strain Xiangnong; Gs, G. sinense; Gt, G. tsugae. (B–D), GO classification of effectors that are unique to Gb_G3 in biological process (BP), molecular function (MF) and cellular component (CC), respectively.

Figure 5.
Heat map displaying the differential expression of 35 unique effector genes in G. boninense strain G3. The differential expression levels (in Log2[RKPM]) were calculated from the RPKM values of Ganoderma (G)- and Trichoderma (T)-inoculated oil palm root samples relative to that of untreated oil palm root sample (derived from the RNAseq data deposited at European Nucleotide Archive (ENA) under accession number: PRJEB7252). Red shows up-regulation whereby the fold change is represented by the number and color intensity (from low to high) while black indicates no expression.
Figure 5.
Heat map displaying the differential expression of 35 unique effector genes in G. boninense strain G3. The differential expression levels (in Log2[RKPM]) were calculated from the RPKM values of Ganoderma (G)- and Trichoderma (T)-inoculated oil palm root samples relative to that of untreated oil palm root sample (derived from the RNAseq data deposited at European Nucleotide Archive (ENA) under accession number: PRJEB7252). Red shows up-regulation whereby the fold change is represented by the number and color intensity (from low to high) while black indicates no expression.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The number of predicted proteins, completeness of the predicted proteome, and percentage of genes with BlastP matches to SwissProt database (≤10−5), GO, and KO annotations.
Table 1.
The number of predicted proteins, completeness of the predicted proteome, and percentage of genes with BlastP matches to SwissProt database (≤10−5), GO, and KO annotations.
| Species | Number of Predicted Proteins in the Genome | Completeness of Proteome Predicted by BUSCO | Functional Annotations | ||||||
|---|---|---|---|---|---|---|---|---|---|
| % Complete BUSCOs (C) | % Complete and Single-Copy BUSCOs (S) | % Complete and Duplicated BUSCOs (D) | % Fragmented BUSCOs (F) | % Missing BUSCOs (M) | % Genes with BlastP Matches | % Genes with BlastP Matches That are Mapped to GO | % Genes with KO Annotation | ||
| G. boninense G3 | 20,564 | 82.6 | 66.7 | 15.9 | 5.3 | 12.1 | 47.95 | 47.93 | 28.08 |
| G. boninense NJ3 | 21,487 | 66.7 | 63.6 | 3.1 | 10.9 | 22.4 | 39.47 | 39.47 | 24.46 |
| G. lucidum G260125-1 | 11,040 | 87.7 | 86.9 | 0.8 | 3.6 | 8.7 | 57.48 | 57.45 | 34.02 |
| G. lucidum Xiangnong | 12,342 | 89.4 | 88.6 | 0.8 | 1.9 | 8.7 | 53.82 | 53.80 | 30.46 |
| G. lucidum BCRC36111 | 12,843 | 75 | 71.1 | 3.9 | 7.8 | 17.2 | 52.93 | 52.90 | 32.04 |
| G. lucidum BCRC37177 | 12,533 | 90.5 | 89.8 | 0.7 | 2.9 | 6.6 | 54.62 | 54.61 | 31.86 |
| G. multipileum | 13,358 | 90.6 | 89.8 | 0.8 | 2.9 | 6.5 | 53.18 | 53.17 | 30.87 |
| G. sinense ZZ0214-1 | 15,478 | 90.1 | 89.0 | 1.1 | 5.0 | 4.9 | 47.83 | 47.70 | 27.08 |
| G. tsugae | 15,426 | 83.5 | 79.8 | 3.7 | 5.6 | 10.9 | 48.17 | 48.15 | 28.90 |
| Ganoderma sp. BRIUMSc | 18,612 | 68.8 | 64.3 | 4.5 | 12.7 | 18.5 | 47.67 | 47.65 | 29.76 |
Table 2.
Selected ortholog groups related to pathogenesis assigned by OrthoVenn.
| GO IDs | GO Annotation | Putative Identity of Protein | Number of Ortholog Group(s) | Number of Proteins | No Proteins in Each Ganoderma Species/Strains | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gb_G3 | Gl_G | Gl_XN | Gl_BCRC37177 | Gm | Gs | Gt | |||||
| Mycotoxin metabolism | |||||||||||
| GO:0009407 | P:toxin catabolic process | Glutathione S-transferase U9 | 1 | 6 | 1 | 0 | 1 | 1 | 1 | 1 | 1 |
| Glutathione S-transferase U6 | 1 | 8 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | ||
| Glutathione S-transferase U23 | 1 | 6 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | ||
| GO:0043386 | P:mycotoxin biosynthetic process | Oxidase ustYa | 2 | 12 | 3 | 1 | 2 | 1 | 1 | 2 | 2 |
| GO:0045122 | P:aflatoxin biosynthetic process | Norsolorinic acid ketoreductase | 1 | 2 | 0 | 0 | 0 | 1 | 1 | 0 | 0 |
| O-methylsterigmatocystin oxidoreductase | 2 | 8 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | ||
| FAD-binding monooxygenase aflW | 2 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Secondary metabolism | |||||||||||
| GO:0009820 | P:alkaloid metabolic process | Tryprostatin B 6-hydroxylase | 4 | 25 | 6 | 3 | 3 | 3 | 3 | 5 | 2 |
| Probable inactive reductase easA | 2 | 16 | 4 | 2 | 2 | 2 | 2 | 2 | 2 | ||
| GO:0035835 | P:indole alkaloid biosynthetic process | Chanoclavine-I aldehyde reductase fgaOx3 | 9 | 6 | 1 | 0 | 1 | 1 | 1 | 1 | 1 |
| GO:0019748 | P:secondary metabolic process | Dehydrogenase orsE | 2 | 14 | 2 | 2 | 1 | 2 | 2 | 3 | 2 |
| Isoflavone reductase (IFR) homolog A622-like | 1 | 10 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | ||
| GO:0016114 | P:terpenoid biosynthetic process | Cytochrome P450 monooxygenase andK | 4 | 17 | 4 | 2 | 2 | 2 | 2 | 3 | 2 |
| Cytochrome P450 monooxygenase mpaDE | 6 | 28 | 3 | 3 | 4 | 4 | 4 | 5 | 5 | ||
| Cytochrome P450 monooxygenase yanC | 3 | 18 | 2 | 2 | 2 | 2 | 2 | 6 | 2 | ||
| Δ (6)-protoilludene synthase | 10 | 60 | 10 | 10 | 5 | 10 | 9 | 9 | 7 | ||
| FAD-dependent monooxygenase yanF | 3 | 6 | 0 | 0 | 0 | 0 | 2 | 0 | 4 | ||
| Methyltransferase ausD | 2 | 6 | 0 | 1 | 0 | 1 | 2 | 1 | 1 | ||
| Methyltransferase trt5 | 1 | 6 | 1 | 1 | 1 | 0 | 0 | 2 | 1 | ||
| Multifunctional cytochrome P450 | 1 | 9 | 2 | 0 | 1 | 1 | 2 | 2 | 1 | ||
| N-Methyltransferase vrtF | 2 | 4 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | ||
| O-Mevalon transferase yanI | 2 | 6 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | ||
| Pathogenesis | |||||||||||
| GO:0009405 | P:pathogenesis | Acyl-CoA ligase sidI | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Aldo-keto reductase AFTS1 | 5 | 17 | 3 | 3 | 1 | 2 | 3 | 3 | 2 | ||
| Cytochrome P450 monooxygenase BOA3 | 1 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Cytochrome P450 monooxygenase BOA4 | 3 | 16 | 2 | 3 | 2 | 3 | 2 | 2 | 2 | ||
| Cytochrome P450 monooxygenase CLM2 | 5 | 22 | 3 | 2 | 4 | 3 | 3 | 3 | 4 | ||
| Enoyl-CoA hydratase AFT3-1 | 3 | 21 | 1 | 3 | 3 | 3 | 3 | 4 | 4 | ||
| Enoyl-CoA isomerase/hydratase fer4 | 1 | 8 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| FAD-dependent monooxygenase OpS4 | 8 | 44 | 4 | 6 | 6 | 8 | 8 | 7 | 5 | ||
| Hydroxymethylglutaryl (HMG)-CoA synthase | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Longiborneol synthase CLM1 | 7 | 39 | 6 | 3 | 7 | 8 | 7 | 5 | 3 | ||
| Orsellinic acid synthase ArmB | 1 | 9 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Oxidoreductase BOA17 | 8 | 41 | 10 | 5 | 4 | 6 | 6 | 6 | 4 | ||
| Reducing polyketide synthase BOA9 | 1 | 5 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | ||
| Fe-regulated protein 8 | 2 | 8 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | ||
| Ferric/cupric reductase transmembrane | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| β-1,2-xylosyltransferase 1 | 1 | 8 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| bZIP transcription factor hapX | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| C2H2 finger domain transcription factor dvrA | 1 | 8 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Calcineurin subunit B | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Global transcription regulator sge1 | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Glycerophosphoinositol permease 1 | 1 | 10 | 2 | 1 | 1 | 1 | 1 | 2 | 2 | ||
| LysM domain-containing protein ARB_00327 | 1 | 7 | 2 | 1 | 1 | 1 | 1 | 1 | 0 | ||
| Metallocarboxypeptidase A-like protein | 1 | 10 | 1 | 2 | 1 | 1 | 2 | 2 | 1 | ||
| Probable aspartic-type endopeptidase CTSD | 1 | 8 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Leucine aminopeptidase | 2 | 19 | 3 | 3 | 2 | 2 | 2 | 3 | 4 | ||
| Tripeptidyl-peptidase SED2 | 7 | 34 | 5 | 4 | 7 | 4 | 4 | 4 | 6 | ||
| Tripeptidyl-peptidase SED3 | 2 | 15 | 2 | 3 | 2 | 2 | 2 | 2 | 2 | ||
| Tripeptidyl-peptidase SED4 | 2 | 8 | 3 | 1 | 1 | 1 | 1 | 1 | 0 | ||
| Sterol biosynthesis and transport | |||||||||||
| GO:0006696 | P:ergosterol biosynthetic process | 3-keto-steroid reductase (EC 1.1.1.270) | 4 | 18 | 3 | 2 | 1 | 1 | 5 | 3 | 3 |
| C-8 sterol isomerase (EC 5.-.-.-) | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Ergosterol biosynthetic protein 28 | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| NADH-cytochrome b5 reductase 2 (EC 1.6.2.2) | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| NADPH-cytochrome P450 reductase (EC 1.6.2.4) | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Sterol 24-C-methyltransferase erg6 (EC 2.1.1.-) | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Sterol-4-α-carboxylate 3-dehydrogenase (EC 1.1.1.170) | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| GO:0032443 | P:regulation of ergosterol biosynthetic process | Damage response protein 1 | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| GO:0015248 | F:sterol transporter activity | Oxysterol-binding protein-related protein 3 (ORP-3) | 1 | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Protein PRY2 (Pathogenesis-related protein 2) | 6 | 43 | 8 | 6 | 5 | 5 | 5 | 8 | 6 | ||
| Cell wall protein PRY3 (Pathogenesis-related protein 3) | 2 | 12 | 1 | 2 | 2 | 2 | 2 | 2 | 1 | ||
| Cell wall degradation | |||||||||||
| GO:0046274 | P: lignin catabolic process | 3-O-methyltransferase | 2 | 10 | 1 | 2 | 1 | 2 | 2 | 1 | 1 |
| 4-O-methyl-glucuronoyl methylesterase | 6 | 40 | 7 | 6 | 5 | 5 | 5 | 6 | 6 | ||
| 4-O-methyltransferase | 13 | 60 | 11 | 5 | 5 | 10 | 11 | 10 | 8 | ||
| Laccase | 24 | 103 | 15 | 14 | 14 | 16 | 16 | 16 | 12 | ||
| GO:0006979 | P:response to oxidative stress | Ligninases | 3 | 16 | 3 | 2 | 2 | 2 | 2 | 2 | 3 |
| Manganese peroxidase | 3 | 19 | 4 | 2 | 2 | 2 | 2 | 4 | 3 | ||
| Versatile peroxidase | 3 | 19 | 3 | 3 | 3 | 3 | 3 | 3 | 1 | ||
Table 3.
The number of putative effectors from Ganoderma Species.
| Number of Proteins with Signal Peptide | Number of Predicted Effector Proteins (% of Number of Proteins with Signal Peptide) | Number of Predicted Effector Proteins with Annotation | Over-Represented GO Terms of Effectors | |
|---|---|---|---|---|
| G. boninense G3 | 2327 | 175 (7.5) | 80 | GO:0005576 (extracellular region) |
| G. lucidum G260125-1 | 1407 | 102 (7.2) | 59 | GO:0010466 (negative regulation of peptidase activity) GO:0004867 (serine-type endopeptidase inhibitor activity) GO:0043086 (negative regulation of catalytic activity) |
| G. lucidum Xiangnong | 1565 | 107 (6.8) | 52 | - |
| G. lucidum BCRC37177 | 1592 | 123 (7.7) | 66 | GO:0005576 (extracellular region) GO:0005615 (extracellular space) |
| G. multipileum | 1625 | 118 (7.3) | 63 | GO:0010466 (negative regulation of peptidase activity) GO:0004867 (serine-type endopeptidase inhibitor activity) |
| G. sinense ZZ0214-1 | 1908 | 153 (8.0) | 71 | GO:0005576 (extracellular region) |
| G. tsugae | 1648 | 144 (8.7) | 85 | GO:0005576 (extracellular region) GO:0030234 (enzyme regulator activity) GO:0016798 (hydrolase activity, acting on glycosyl bonds) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ho, C.-L. Comparative Genomics Analysis of Ganoderma Orthologs Involved in Plant-Pathogenesis. Forests 2023, 14, 653. https://doi.org/10.3390/f14030653
AMA Style
Ho C-L. Comparative Genomics Analysis of Ganoderma Orthologs Involved in Plant-Pathogenesis. Forests. 2023; 14(3):653. https://doi.org/10.3390/f14030653
Chicago/Turabian StyleHo, Chai-Ling. 2023. "Comparative Genomics Analysis of Ganoderma Orthologs Involved in Plant-Pathogenesis" Forests 14, no. 3: 653. https://doi.org/10.3390/f14030653
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.