
A Novel 3D Tree-Modeling Method of Incorporating Small-Scale Spatial Structure Parameters in a Heterogeneous Forest Environment
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
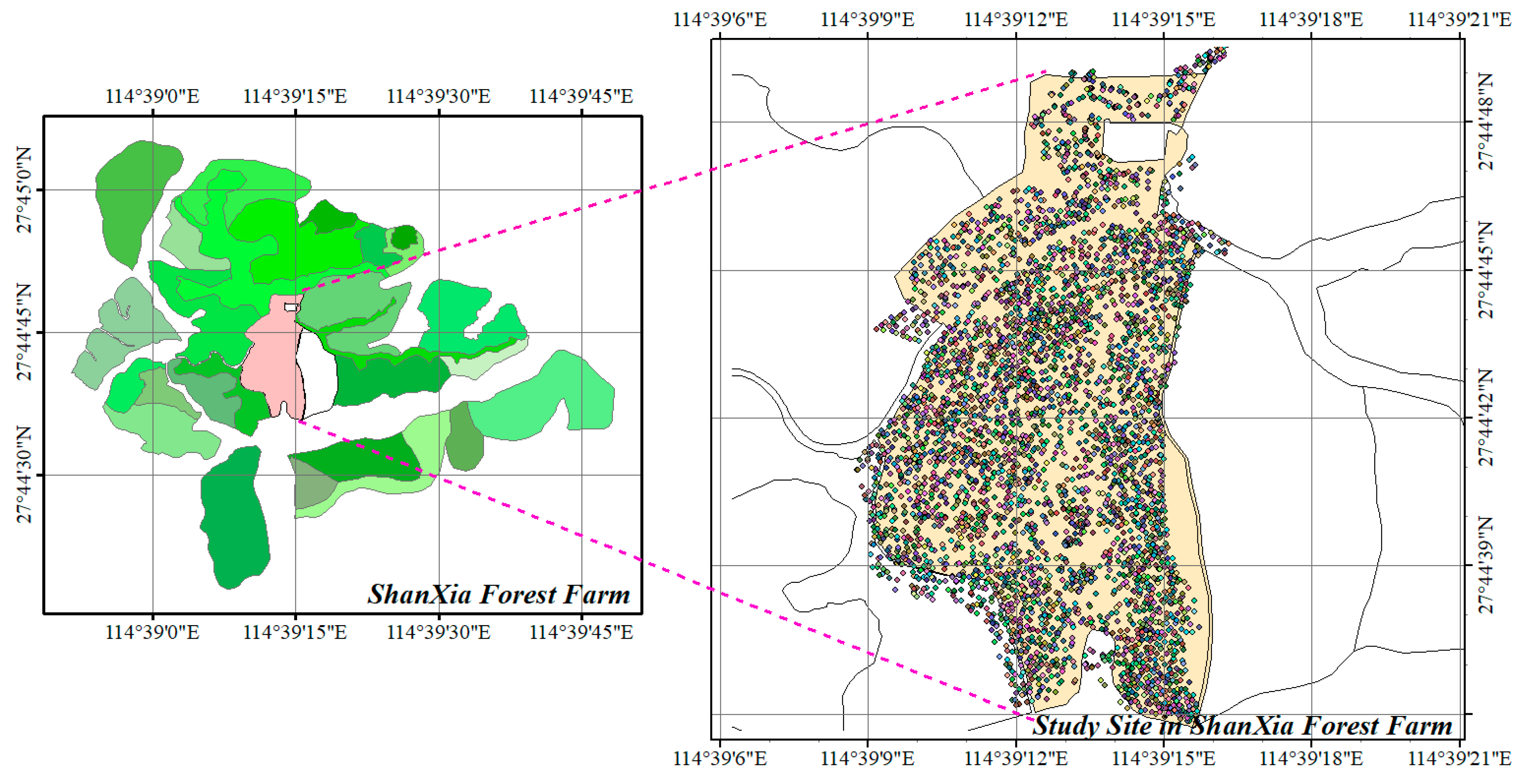
2.1. Study Areas
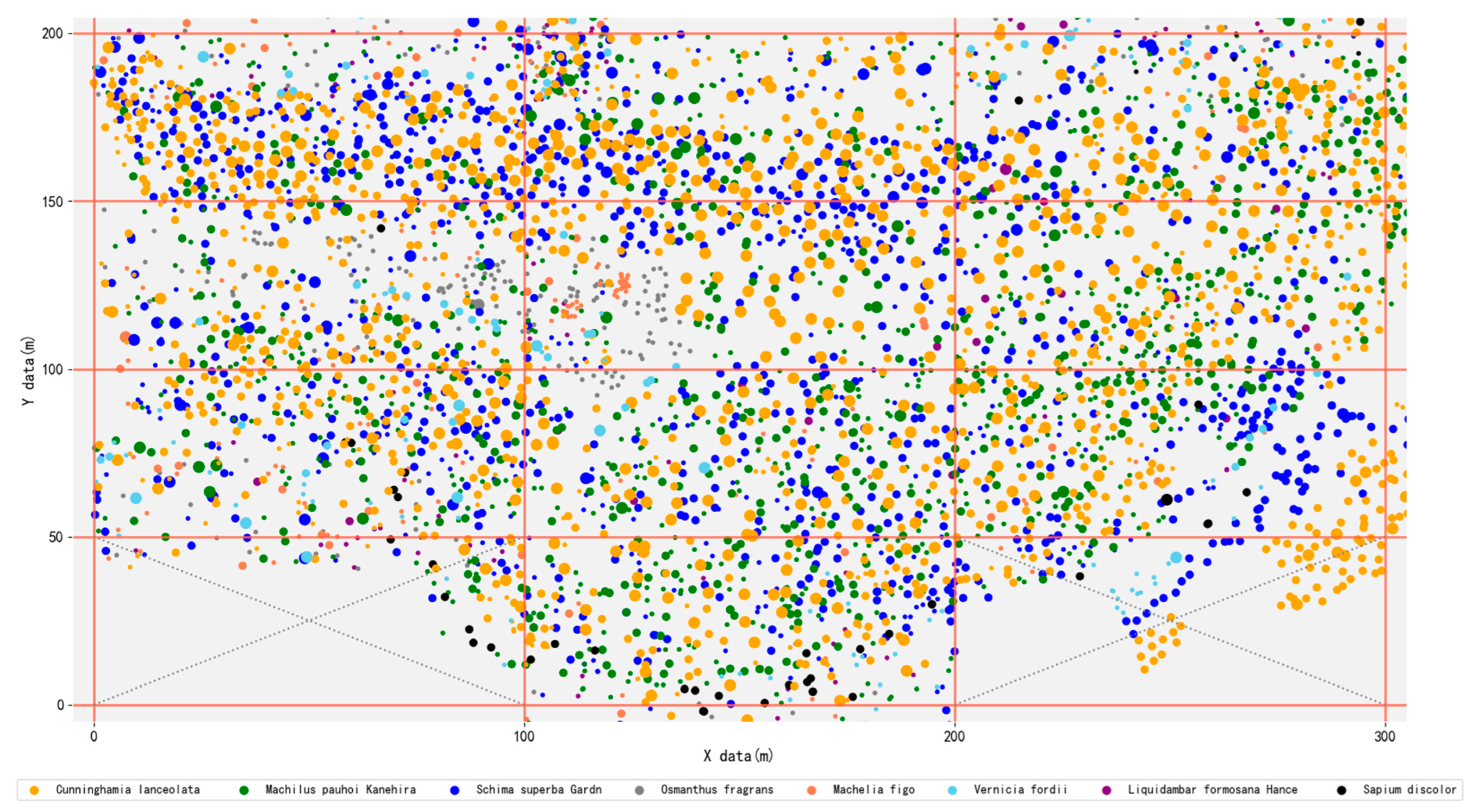
2.2. Data Collection
2.3. Data Analysis
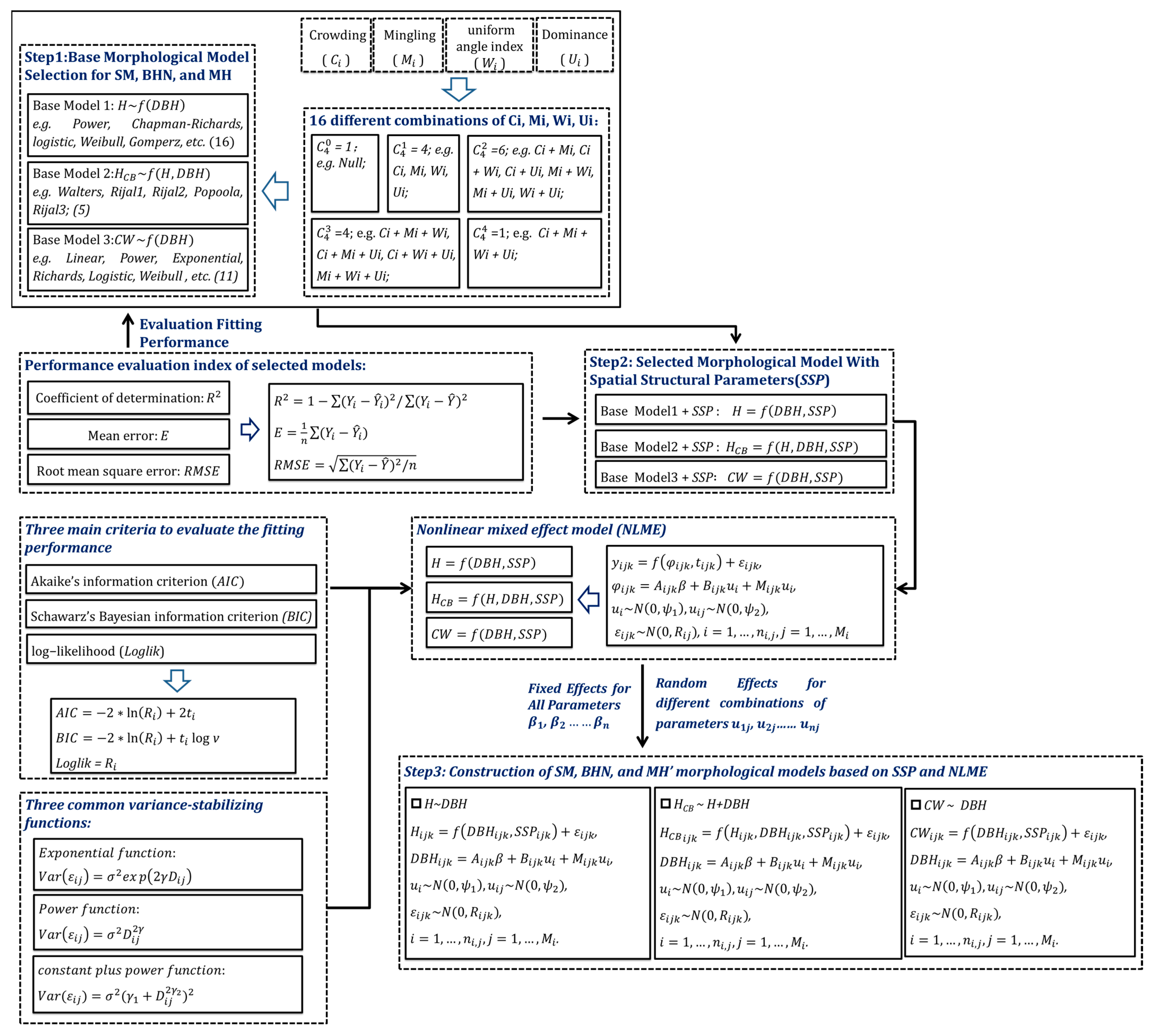
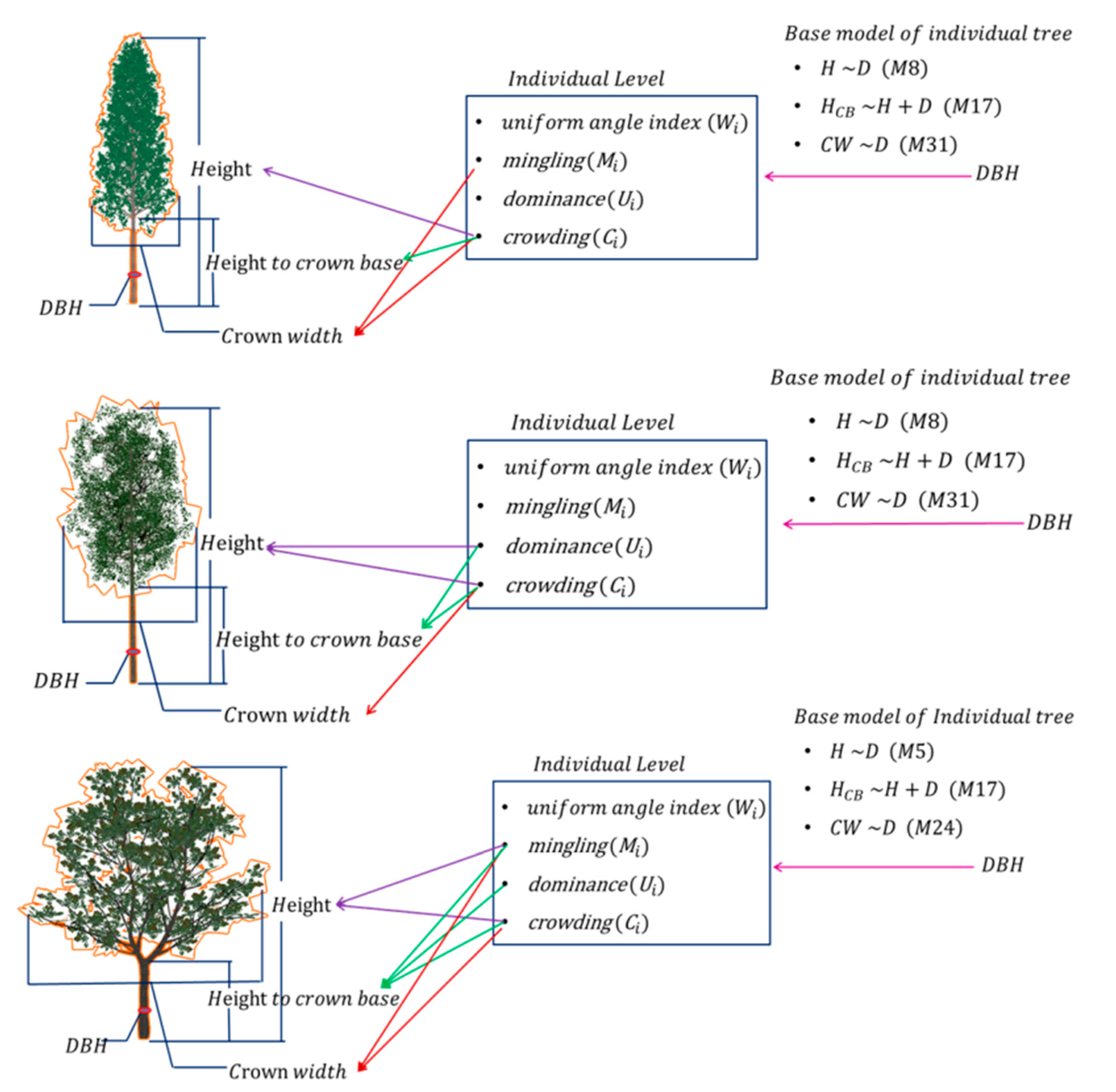
2.3.1. Identification of Dominant Populations for 3D Tree Modeling by the Important Value Index
2.3.2. Nonlinear Mixed-Effect Morphological Model of Three Predominant Trees
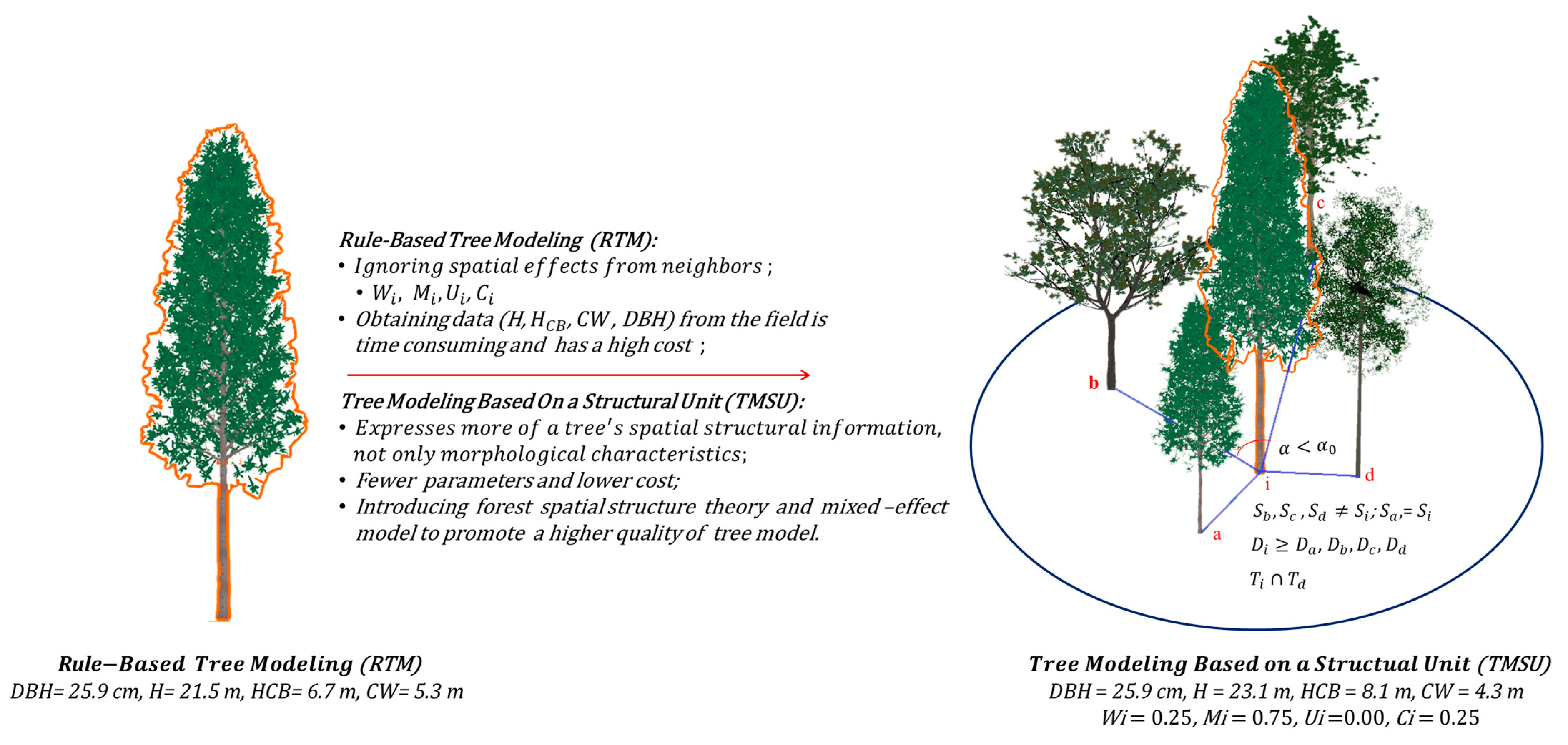
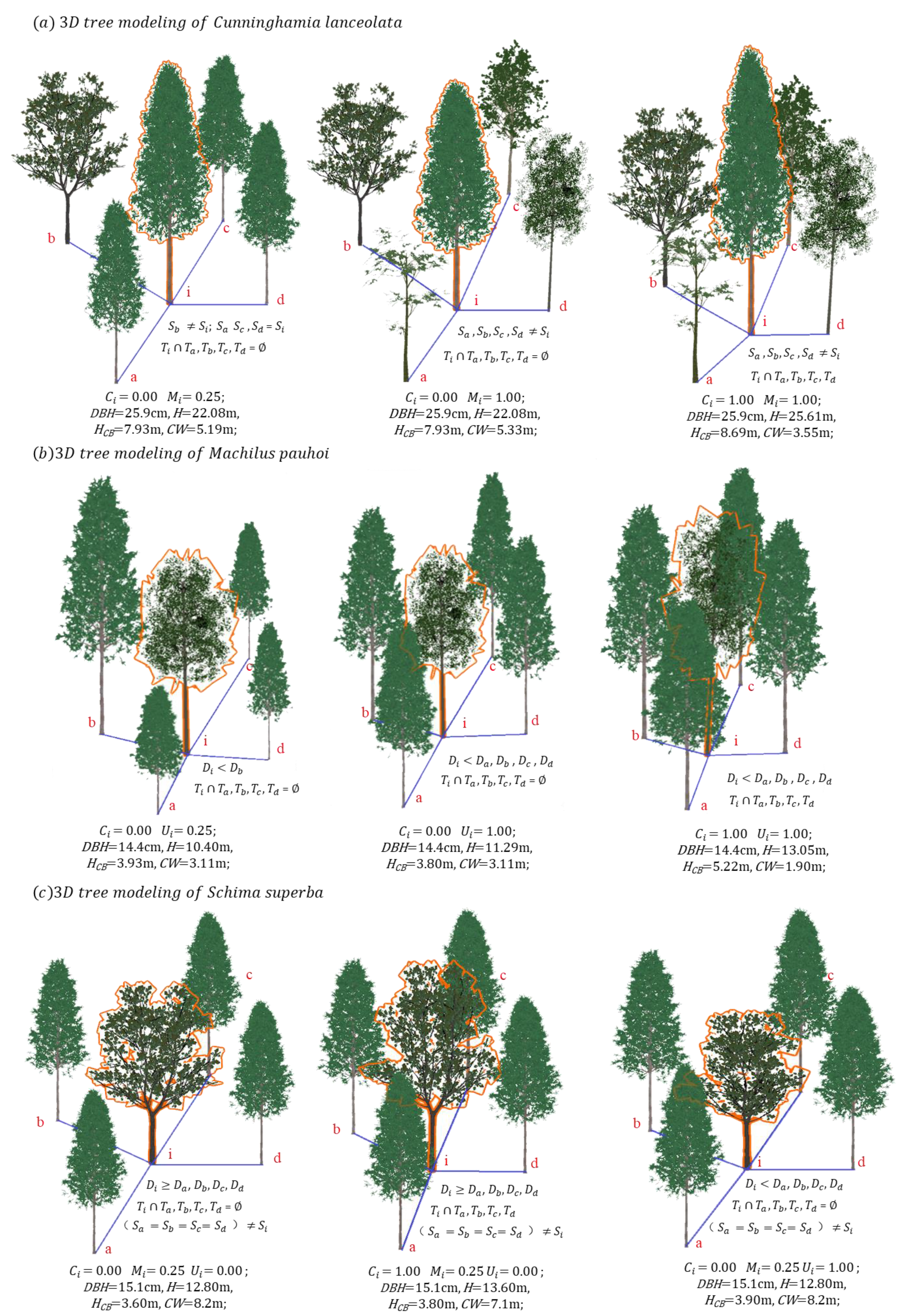
2.3.3. 3D Tree Modeling Based on a Structural Unit
3. Results
3.1. Three-Dimensional Tree Morphological Model Development at the Individual Tree Level
3.2. Analysis of 3D Tree Modeling Based on a Structural Unit
4. Discussion
4.1. The Development of Morphological Characteristic Models Involving Forest Spatial Structure in TMSU
4.2. Three-Dimensional Tree Modeling in a Highly Heterogeneous Forest Environment
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Paulus, S.; Dupuis, J.; Mahlein, A.K.; Kuhlmann, H. Surface feature based classification of plant organs from 3D laserscanned point clouds for plant phenotyping. BMC Bioinform. 2013, 14, 238. [Google Scholar] [CrossRef] [Green Version]
- Thapa, S.; Zhu, F.; Walia, H.; Yu, H.; Ge, Y. A novel LiDAR-based instrument for high-throughput, 3D measurement of morphological traits in maize and sorghum. Sensors 2018, 18, 1187. [Google Scholar] [CrossRef] [Green Version]
- Indirabai, I.; Nair, M.V.H.; Jaishanker, R.N.; Nidamanuri, R.R. Terrestrial laser scanner based 3D reconstruction of trees and retrieval of leaf area index in a forest environment. Ecol. Inform. 2019, 53, 100986. [Google Scholar] [CrossRef]
- Sun, R.; Jia, J.; Jaeger, M. Intelligent tree modeling based on L-system. In Proceedings of the 2009 IEEE 10th International Conference on Computer-Aided Industrial Design & Conceptual Design, Wenzhou, China, 26–29 November 2009; pp. 1096–1100. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, D.; Zhou, K.; Jia, J. Rule and reuse based lightweight modeling and real time web3D rendering of forest scenes. In Proceedings of the 23rd International ACM Conference on 3D Web Technology, Poznań, Poland, 20–22 June 2018; pp. 1–8. [Google Scholar] [CrossRef]
- Tang, L.; Peng, X.; Chen, C.; Huang, H.; Lin, D. Three-dimensional Forest growth simulation in virtual geographic environments. Earth Sci. Inform. 2019, 12, 31–41. [Google Scholar] [CrossRef]
- Kim, D.; Kang, W.H.; Hwang, I.; Kim, J.; Kim, J.H.; Park, K.S.; Son, J.E. Use of structurally-accurate 3D plant models for estimating light interception and photosynthesis of sweet pepper (Capsicum annuum) plants. Comput. Electron. Agric. 2020, 177, 105689. [Google Scholar] [CrossRef]
- Zhang, X.; Li, H.; Dai, M.; Ma, W.; Quan, L. Data-driven synthetic modeling of trees. IEEE Trans. Vis. Comput. Graph. 2014, 20, 1214–1226. [Google Scholar] [CrossRef]
- Seidel, D.; Hoffmann, N.; Ehbrecht, M.; Juchheim, J.; Ammer, C. How neighborhood affects tree diameter increment–new insights from terrestrial laser scanning and some methodical considerations. For. Ecol. Manag. 2015, 336, 119–128. [Google Scholar] [CrossRef]
- Bayer, D.; Seifert, S.; Pretzsch, H. Structural crown properties of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica [L.]) in mixed versus pure stands revealed by terrestrial laser scanning. Trees 2013, 27, 1035–1047. [Google Scholar] [CrossRef]
- Seidel, D.; Leuschner, C.; Müller, A.; Krause, B. Crown plasticity in mixed forests—Quantifying asymmetry as a measure of competition using terrestrial laser scanning. For. Ecol. Manag. 2011, 261, 2123–2132. [Google Scholar] [CrossRef]
- Forrester, D.I.; Benneter, A.; Bouriaud, O.; Bauhus, J. Diversity and competition influence tree allometric relationships–developing functions for mixed-species forests. J. Ecol. 2017, 105, 761–774. [Google Scholar] [CrossRef] [Green Version]
- Vincent, G.; Harja, D. Exploring ecological significance of tree crown plasticity through three-dimensional modelling. Ann. Bot. 2008, 101, 1221–1231. [Google Scholar] [CrossRef] [Green Version]
- Purves, D.W.; Lichstein, J.W.; Pacala, S.W. Crown plasticity and competition for canopy space: A new spatially implicit model parameterized for 250 North American tree species. PLoS ONE 2007, 2, e870. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Jucker, T.; Bouriaud, O.; Coomes, D.A. Crown plasticity enables trees to optimize canopy packing in mixed-species forests. Funct. Ecol. 2015, 29, 1078–1086. [Google Scholar] [CrossRef] [Green Version]
- Lines, E.R.; Zavala, M.A.; Purves, D.W.; Coomes, D.A. Predictable changes in aboveground allometry of trees along gradients of temperature, aridity and competition. Glob. Ecol. Biogeogr. 2012, 21, 1017–1028. [Google Scholar] [CrossRef]
- Grueters, U.; Seltmann, T.; Schmidt, H.; Horn, H.; Pranchai, A.; Vovides, A.G.; Peters, R.; Vogt, J.; Dahdouh-Guebas, F.; Berger, U. The mangrove forest dynamics model mesoFON. Ecol. Model. 2014, 291, 28–41. [Google Scholar] [CrossRef]
- Pommerening, A.; Gaulton, R.; Magdon, P.; Myllymäki, M. CanopyShotNoise–An individual-based tree canopy modelling framework for projecting remote-sensing data and ecological sensitivity analysis. Int. J. Remote Sens. 2021, 42, 6837–6865. [Google Scholar] [CrossRef]
- Fischer, F.J.; Labriere, N.; Vincent, G.; Hérault, B.; Alonso, A.; Memiaghe, H.; Bissiengou, P.; Kenfack, D.; Saatchi, S.; Chaveet, J. A simulation method to infer tree allometry and forest structure from airborne laser scanning and forest inventories. Remote Sens. Environ. 2020, 251, 112056. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, Q.; Luo, P.; Ye, Q.; Sharma, R.P.; Duan, G.; Zhang, H.; Fu, L. Nonlinear mixed-effects height to crown base model based on both airborne LiDAR and field datasets for Picea crassifolia Kom trees in northwest China. For. Ecol. Manag. 2020, 474, 118323. [Google Scholar] [CrossRef]
- Ma, A.; Miao, Z.; Xie, L.; Dong, L.; Li, F. Crown width prediction for Larix olgensis plantations in Northeast China based on nonlinear mixed-effects model and quantile regression. Trees 2022, 36, 1761–1776. [Google Scholar] [CrossRef]
- Sharma, R.P.; Štefančík, I.; Vacek, Z.; Vacek, S. Generalized nonlinear mixed-effects individual tree diameter increment models for beech forests in Slovakia. Forests 2019, 10, 451. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.P.; Vacek, Z.; Vacek, S.; Kučera, M. A nonlinear mixed-effects height-to-diameter ratio model for several tree species based on Czech national forest inventory data. Forests 2019, 10, 70. [Google Scholar] [CrossRef] [Green Version]
- Bronisz, K.; Mehtätalo, L. Mixed-effects generalized height–diameter model for young silver birch stands on post-agricultural lands. For. Ecol. Manag. 2020, 460, 117901. [Google Scholar] [CrossRef]
- Ciceu, A.; Garcia-Duro, J.; Seceleanu, I.; Badea, O. A generalized nonlinear mixed-effects height–diameter model for Norway spruce in mixed-uneven aged stands. For. Ecol. Manag. 2020, 477, 118507. [Google Scholar] [CrossRef]
- Fu, L.; Zhang, H.; Sharma, R.P.; Pang, L.; Wang, G. A generalized nonlinear mixed-effects height to crown base model for Mongolian oak in northeast China. For. Ecol. Manag. 2017, 384, 34–43. [Google Scholar] [CrossRef]
- Fu, L.; Sharma, R.P.; Hao, K.; Tang, S. A generalized interregional nonlinear mixed-effects crown width model for Prince Rupprecht larch in northern China. For. Ecol. Manag. 2017, 389, 364–373. [Google Scholar] [CrossRef]
- Zhang, S.; Sun, J.; Duan, A.; Zhang, J. Variable-exponent taper equation based on multilevel nonlinear mixed effect for Chinese fir in China. Forests 2021, 12, 126. [Google Scholar] [CrossRef]
- Bragg, D.C. A local basal area adjustment for crown width prediction. North. J. Appl. For. 2001, 18, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Marshall, D.D.; Johnson, G.P.; Hann, D.W. Crown profile equations for stand-grown western hemlock trees in northwestern Oregon. Can. J. For. Res. 2003, 33, 2059–2066. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Hui, G.; Zhao, Z.; Hu, Y.; Ye, S. Spatial structural characteristics of three hardwood species in Korean pine broad-leaved forest—Validating the bivariate distribution of structural parameters from the point of tree population. For. Ecol. Manag. 2014, 314, 17–25. [Google Scholar] [CrossRef]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; Wiley: New York, NY, USA, 1974. [Google Scholar]
- Mehtätalo, L.; de-Miguel, S.; Gregoire, T.G. Modeling height-diameter curves for prediction. Can. J. For. Res. 2015, 45, 826–837. [Google Scholar] [CrossRef] [Green Version]
- Leduc, D.; Goelz, J. A height–diameter curve for longleaf pine plantations in the Gulf Coastal Plain. South. J. Appl. For. 2009, 33, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Von Gadow, K.; Hui, G.Y. Characterizing forest spatial structure and diversity. In Sustainable Forestry in Temperate Regions; Björk, L., Ed.; SUFOR, University of Lund: Lund, Sweden, 2002; pp. 20–30. [Google Scholar]
- Hui, G.; Zhang, G.; Zhao, Z.; Yang, A. Methods of forest structure research: A review. Curr. For. Rep. 2019, 5, 142–154. [Google Scholar] [CrossRef]
- Gadow, K.; Zhang, G.Q.; Durrheim, G.; Drew, D.; Seydack, A. Diversity and production in an Afromontane Forest. For. Ecosyst. 2016, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhang, H.; Li, Y.; Yang, T.; Shen, K. Three-Dimensional Visualization Simulation of Chinese Fir Stand Growth Based on Unity3D. In Proceedings of the 2018 3rd International Conference on Mechanical, Control and Computer Engineering (ICMCCE), Huhhot, China, 14–16 September 2018; pp. 530–534. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.Q.; Ju, H.B.; Tang, X.; Hu, B. 3-D tree model and its visualization in Guozigou forest farm. For. Res. Beijing 2016, 29, 74–79. [Google Scholar] [CrossRef]
- Lu, K.N.; Zhang, H.Q.; Liu, M.; Ouyang, G. Design and implementation of individual tree growth visualization system of Cunninghamia lanceolata. For. Res. Beijing 2012, 25, 207–211. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, H.; Chen, Y.F.; Liu, M. Study on visual simulation technology of Cunninghamia lanceolata morphological characters. For. Res. Beijing 2010, 23, 59–64. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, M. Tree growth simulation method based on improved IFS algorithm. In Proceedings of the 2009 International Conference on Computational Intelligence and Software Engineering, Wuhan, China, 11–13 December 2009; pp. 1–5. [Google Scholar] [CrossRef]
- Fu, L.; Sun, H.; Sharma, R.P.; Lei, Y.; Zhang, H.; Tang, S. Nonlinear mixed-effects crown width models for individual trees of Chinese fir (Cunninghamia lanceolata) in south-central China. For. Ecol. Manag. 2013, 302, 210–220. [Google Scholar] [CrossRef]
- Knutti, R. Should we believe model predictions of future climate change? Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2008, 366, 4647–4664. [Google Scholar] [CrossRef]
- Leng, W.; He, H.S.; Bu, R.; Dai, L.; Hu, Y.; Wang, X. Predicting the distributions of suitable habitat for three larch species under climate warming in Northeastern China. For. Ecol. Manag. 2008, 254, 420–428. [Google Scholar] [CrossRef]
- Lei, X.; Yu, L.; Hong, L. Climate-sensitive integrated stand growth model (CS-ISGM) of Changbai larch (Larix olgensis) plantations. For. Ecol. Manag. 2016, 376, 265–275. [Google Scholar] [CrossRef]
- Tasoulas, E.; Varras, G.; Tsirogiannis, I.; Myriounis, C. Development of a GIS application for urban forestry management planning. Procedia Technol. 2013, 8, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Clark, P.J.; Evans, F.C. Distance to nearest neighbor as a measure of spatial relationships in populations. Ecology 1954, 35, 445–453. [Google Scholar] [CrossRef]
- Morisita, M. Estimation of population density by spacing method. Mem. Fac. Sci. Kyushu Univ. Ser. E Biol. 1954, 1, 187–197. [Google Scholar] [CrossRef]
- Ripley, B.D. Modelling spatial patterns. J. R. Stat. Soc. Ser. B Methodol. 1977, 39, 172–192. [Google Scholar] [CrossRef]
- Upton, G.; Fingleton, B. Spatial Data Analysis by Example. Volume 1: Point Pattern and Quantitative Data; John Wiley & Sons Ltd.: Chichester, UK, 1985. [Google Scholar]
- Wiegand, T.; AMoloney, K. Rings, circles, and nullal. Rule and reuse based lightweight mecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Lexerød, N.L.; Eid, T. An evaluation of different diameter diversity indices based on criteria related to forest management planning. For. Ecol. Manag. 2006, 222, 17–28. [Google Scholar] [CrossRef]
- Getzin, S.; Dean, C.; He, F.; Trofymow, J.A.; Wiegand, K.; Wiegand, T. Spatial patterns and competition of tree species in a Douglastems, chronosequence on Vancouver Island. Ecography 2006, 29, 671–682. [Google Scholar] [CrossRef]
- Corral-Rivas, J.J.; Wehenkel, C.; Castellanos-Bocaz, H.A.; Vargas-Larreta, B.; Diéguez-Aranda, U. A permutation test of spatial randomness: Application to nearest neighbour indices in forest stands. J. For. Res. 2010, 15, 218–225. [Google Scholar] [CrossRef]
- Gadow, K.; Füldner, K. Zur bestandesbeschreibung in der forsteinrichtung. Forst Und Holz 1993, 48, 602–606. [Google Scholar]
- Gadow, V.K.; Hui, G.Y. Modelling Forest Development; Springer Science & Business Media: Göttingen, Germany, 1999. [Google Scholar]
- Aguirre, O.; Hui, G.; von Gadow, K.; Jiménez, J. An analysis of spatial forest structure using neighbourhood-based variables. For. Ecol. Manag. 2003, 183, 137–145. [Google Scholar] [CrossRef]
- Hui, G.; Wang, Y.; Zhang, G.; Zhao, Z.; Bai, C.; Liu, W. A novel approach for assessing the neighborhood competition in two different aged forests. For. Ecol. Manag. 2018, 422, 49–58. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Density | IV (%) | Stand Characteristics | ||||||
|---|---|---|---|---|---|---|---|---|---|
| (Trees ha−1) | Upper Layer | Median Layer | Lower Layer | All | DBH | Height | Basal Area | Dominant Height (m) | |
| (cm) | (m) | (m2) | |||||||
| Cunninghamia lanceolata (Lamb.) Hook. | 273.0 | 14.57 ± 7.14 | 15.13 ± 5.19 | 3.28 ± 1.86 | 17.18 ± 3.74 | 24.86 ± 8.91 | 16.50 ± 5.40 | 80.83 | 26.8 |
| Machilus pauhoi Kanehira | 252.6 | 2.53 ± 1.68 | 10.38 ± 2.73 | 7.04 ± 3.13 | 11.13 ± 3.44 | 13.82 ± 8.25 | 10.63 ± 3.83 | 25.72 | 23.5 |
| Schima superba Gardn. et Champ. | 259.0 | 1.21 ± 1.25 | 12.78 ± 4.21 | 4.32 ± 1.88 | 12.39 ± 4.39 | 15.32 ± 7.02 | 11.85 ± 3.55 | 27.67 | 23.4 |
| Osmanthus fragrans (Thunb.) Lour. | 62.0 | 0.00 ± 0.00 | 0.00 ± 0.00 | 1.82 ± 3.56 | 1.65 ± 2.94 | 10.37 ± 3.31 | 5.74 ± 1.12 | 2.88 | 8.2 |
| Machelia figo (Lour.) Spreng. | 48.0 | 0.07 ± 0.21 | 0.81 ± 0.90 | 1.12 ± 1.35 | 1.59 ± 1.60 | 13.69 ± 8.03 | 9.30 ± 3.76 | 4.36 | 18.8 |
| Vernicia fordii (Hemsl.) Airy Shaw | 35.6 | 0.21 ± 0.55 | 1.34 ± 1.23 | 0.41 ± 0.74 | 1.52 ± 1.65 | 22.60 ± 9.47 | 14.21 ± 3.40 | 7.54 | 21.1 |
| Liquidambar formosana Hance | 27.0 | 0.03 ± 0.09 | 0.63 ± 0.90 | 0.74 ± 0.86 | 1.24 ± 1.43 | 16.26 ± 7.63 | 12.19 ± 3.45 | 3.01 | 18.2 |
| Sapium discolor (Champ. ex Benth.) Muell. Arg. | 21.8 | 0.00 ± 0.00 | 0.91 ± 1.76 | 0.13 ± 0.40 | 0.74 ± 1.21 | 22.22 ± 7.83 | 13.33 ± 1.92 | 4.21 | 15.4 |
| Model | Formula | Model Form | Model | Formula | Model Form |
|---|---|---|---|---|---|
| M1 | H~1.3 + DBH2/(a∗DBH + b)2 | Naslund | M17 | HCB~H/(1 + exp(a + b∗DBH)) | Walters |
| M2 | H~1.3 + a∗DBH/(1 + DBH)b | Curtis | M18 | HCB~H/(1 + exp(a + b∗DBH))1/2 | Rijal 1 |
| M3 | H~1.3 + a∗exp(−b/DBH) | Schumacher | M19 | HCB~H/(1 + exp(a + b∗DBH))1/6 | Rijal 2 |
| M4 | H~1.3 + a∗(1 − exp(−b∗DBH)) | Meyer | M20 | HCB~H/(a + exp(−b∗DBH)) | Popoola |
| M5 | H~1.3 + a∗DBHb | Power | M21 | HCB~H/(1 − a∗exp(−b∗DBH)) | Rijal 3 |
| M6 | H~1.3 + a∗DBH/(b + DBH) | Michaelis-Menten | M22 | CW~a + b∗DBH | Linear |
| M7 | H~1.3 + exp(a − b/(DBH + 1)) | Wykoff | M23 | CW~a + b∗DBH + c∗DBH2 | Quadratic |
| M8 | H~1.3 + a∗((1 − exp(-b∗DBH))) | Chapman–Richards | M24 | CW~a∗DBHb | Power |
| M9 | H~1.3 + DBH2/(a∗DBH2 + b∗DBH + c) | Prodan | M25 | CW~a∗(1−exp(−b∗DBH)) | Monomolecular |
| M10 | H~1.3 + a/(1 + b∗exp(−c∗DBH)) | logistic | M26 | CW~(DBH/(a + b∗DBH))2 | Hossfeld I |
| M11 | H~1.3 + a∗(1-exp(−b∗DBHc)) | Weibull | M27 | CW~a∗(b)DBH | Compound |
| M12 | H~1.3 + a∗exp(−b∗exp(−c∗DBH)) | Gomperz | M28 | CW~exp(a + b∗DBH) | Growth |
| M13 | H~1.3 + a∗DBH(b∗DBH−c) | Sibbesen | M29 | CW~a∗exp(b∗DBH) | Exponential |
| M14 | H~1.3 + a∗exp(−b∗DBH−c) | Korf | M30 | CW~a∗(1 − exp(−b∗DBH)) | Richards |
| M15 | H~1.3 + a∗exp(−b/(DBH + c)) | Ratkowsky | M31 | CW~a/(1 + b∗exp(−c∗DBH)) | Logistic |
| M16 | H~1.3 + a/(1 + 1/(b∗DBHc)) | Hossfeld IV | M32 | CW~a∗(1 − exp(−b∗DBH)) | Weibull |
| Species | Model | Evaluation Indices | Residual Variance Function | |||
|---|---|---|---|---|---|---|
| Null | Exponential Function | Power Function | Constant Plus Power Function | |||
| SM | Equation (23) | AIC | 4336.224 | 4337.334 | 4337.458 | 4339.477 |
| Loglik | −2158.11 | −2157.667 | −2157.729 | −2157.739 | ||
| p Value | p > 0.05 | p > 0.05 | p > 0.05 | |||
| Equation (24) | AIC | 3438.982 | 3421.818 | 3421.136 | 3423.136 | |
| Loglik | −1712.491 | −1702.909 | −1702.568 | −1702.568 | ||
| p Value | p < 0.0001 | p < 0.0001 | p < 0.0001 | |||
| Equation (25) | AIC | 1935.082 | 1935.461 | 1934.881 | 1936.881 | |
| Loglik | −960.5411 | −959.7304 | −959.441 | −959.4404 | ||
| p Value | p > 0.05 | p > 0.05 | p > 0.05 | |||
| BHN | Equation (26) | AIC | 3825.354 | 3827.209 | 3825.14 | 3827.14 |
| Loglik | −1906.677 | −1906.604 | −1905.57 | −1905.57 | ||
| p Value | p > 0.05 | p > 0.05 | p > 0.05 | |||
| Equation (27) | AIC | 3322.544 | 3249.631 | 3231.132 | 3235.042 | |
| Loglik | −1906.677 | −1616.815 | −1607.57 | −1608.521 | ||
| p Value | p < 0.0001 | p < 0.0001 | p < 0.0001 | |||
| Equation (28) | AIC | 3160.599 | 2365.384 | 2449.84 | 2342.501 | |
| Loglik | −1574.299 | −1175.692 | −1217.92 | −1163.251 | ||
| p Value | p < 0.0001 | p < 0.0001 | p < 0.0001 | p < 0.0001 | ||
| MH | Equation (29) | AIC | 4316.121 | 4313.26 | 4310.291 | 4312.291 |
| Loglik | −2150.061 | −2147.63 | −2146.145 | −2146.145 | ||
| p Value | p < 0.05 | p < 0.05 | p < 0.05 | |||
| Equation (30) | AIC | 3798.104 | 3701.172 | 3699.936 | 3701.532 | |
| Loglik | −1888.052 | −1838.586 | −1837.968 | −1837.766 | ||
| p Value | p < 0.0001 | p < 0.0001 | p < 0.0001 | |||
| Equation (31) | AIC | 2462.686 | 2418.486 | 2427.477 | 2416.827 | |
| Loglik | −1223.343 | −1200.243 | −1204.739 | −1198.413 | ||
| p Value | p < 0.0001 | p < 0.0001 | p < 0.0001 | |||
| Species | Formula | Model | Fixed Parameters | Evaluation Indexes | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 𝛽1 | 𝛽2 | 𝛽3 | 𝛽4 | 𝛽5 | AIC | Loglik | p Value | |||
| SM | H-D | Equation (5) | 33.4665 | 0.0266 | —— | —— | —— | 4686.538 | −2340.27 | a |
| Equation (14) | 32.5791 | 0.0252 | −0.0037 | —— | —— | 4350.543 | −2168.27 | b *** | ||
| Equation (23) | 36.7543 | 0.0236 | −0.0045 | —— | —— | 4336.224 | −2158.11 | c *** | ||
| HCB-H | Equation (6) | 0.4501 | −0.0012 | —— | —— | —— | 3488.336 | −1741.17 | a | |
| Equation (15) | 0.4230 | 0.0061 | −0.0087 | —— | —— | 3441.651 | −1716.83 | b *** | ||
| Equation (24) | 0.3896 | 0.0073 | −0.0096 | —— | —— | 3421.136 | −1702.57 | c ** | ||
| CW-D | Equation (7) | 5.9591 | 2.4952 | 0.0538 | —— | —— | 2224.718 | −1108.36 | a | |
| Equation (16) | 5.5826 | 2.3971 | 0.0461 | −0.0367 | 0.0153 | 2030.871 | −1009.44 | b *** | ||
| Equation (25) | 5.5212 | 2.4130 | 0.0520 | −0.0310 | 0.0162 | 1934.881 | −959.441 | c *** | ||
| BHN | H-D | Equation (8) | 19.3411 | 0.0540 | —— | —— | —— | 3970.13 | −1982.07 | a |
| Equation (17) | 19.6529 | 0.0399 | −0.0123 | −0.0072 | —— | 3935.849 | −1962.93 | b *** | ||
| Equation (26) | 20.0990 | 0.0399 | −0.0133 | −0.0078 | —— | 3825.14 | −1905.57 | c *** | ||
| HCB-H | Equation (9) | 0.2933 | 0.0043 | —— | —— | —— | 3404.81 | −1699.41 | a | |
| Equation (18) | 0.0384 | −0.0310 | 0.0202 | —— | —— | 3335.732 | −1663.87 | b *** | ||
| Equation (27) | 0.0408 | −0.0308 | 0.0133 | —— | —— | 3231.132 | −1607.57 | c *** | ||
| CW-D | Equation (10) | 8.3246 | 4.1547 | 0.0885 | —— | —— | 3218.003 | −1605 | a | |
| Equation (19) | 8.4723 | 3.7601 | 0.0501 | −0.0386 | —— | 3188.058 | −1589.03 | b *** | ||
| Equation (28) | 6.7421 | 3.2953 | 0.0721 | −0.0455 | —— | 2342.501 | −1163.25 | c *** | ||
| MH | H-D | Equation (11) | 2.5166 | 0.5378 | —— | —— | —— | 4577.814 | −2285.91 | a |
| Equation (20) | 2.6616 | 0.5135 | −0.3857 | 0.3370 | —— | 4526.287 | −2258.14 | b *** | ||
| Equation (29) | 2.3741 | 0.5686 | −0.2014 | 0.0622 | —— | 4310.291 | −2146.15 | c *** | ||
| HCB-H | Equation (12) | 0.8568 | −0.0036 | —— | —— | —— | 3980.823 | −1987.41 | a | |
| Equation (21) | 0.5636 | 0.0253 | 0.0185 | −0.0103 | —— | 3919.156 | −1954.58 | b *** | ||
| Equation (30) | 0.5479 | 0.0277 | 0.0132 | −0.0026 | —— | 3699.936 | −1837.97 | c *** | ||
| CW-D | Equation (13) | 0.8478 | 0.5819 | —— | —— | —— | 2749.965 | −1371.98 | a | |
| Equation (22) | 0.5918 | 0.5606 | 0.3178 | 0.1041 | —— | 2554.176 | −1272.09 | b *** | ||
| Equation (31) | 0.6514 | 0.5591 | 0.2820 | 0.0519 | —— | 2416.827 | −1198.41 | c *** | ||
| Species | Formula | Model | Variance Components | Fit Statistics | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | E | RMSE | |||||||||
| SM | H-D | Equation (5) | —— | —— | —— | —— | —— | —— | 0.7155 | −0.0201 | 2.7644 |
| Equation (14) | —— | —— | —— | —— | —— | —— | 0.7369 | −0.0199 | 2.6584 | ||
| Equation (23) | 4.99 × 100 | 7.51 × 10−5 | 2.90 × 10−5 | −9.86 × 10−1 | 8.83 × 10−1 | −9.47 × 10−1 | 0.8203 | −0.0190 | 2.1971 | ||
| HCB-H | Equation (6) | —— | —— | —— | —— | —— | —— | 0.6121 | 0.0635 | 1.4820 | |
| Equation (15) | —— | —— | —— | —— | —— | —— | 0.6313 | 0.0739 | 1.4450 | ||
| Equation (24) | 3.78 × 10−6 | —— | —— | —— | —— | —— | 0.6406 | 0.0670 | 1.4265 | ||
| CW-D | Equation (7) | —— | —— | —— | —— | —— | —— | 0.4276 | −0.0004 | 0.7671 | |
| Equation (16) | —— | —— | —— | —— | —— | —— | 0.5341 | −0.0004 | 0.6921 | ||
| Equation (25) | 1.24 × 10−1 | 2.95 × 10−2 | —— | −1.81 × 10−1 | —— | —— | 0.5965 | −0.0013 | 0.6441 | ||
| BHN | H-D | Equation (8) | —— | —— | —— | —— | —— | —— | 0.6752 | 0.0250 | 2.1741 |
| Equation (17) | —— | —— | —— | —— | —— | —— | 0.6887 | 0.0201 | 2.1285 | ||
| Equation (26) | 3.17 × 10−6 | 9.95 × 10−5 | —— | −8.83 × 10−1 | —— | —— | 0.7376 | 0.0205 | 1.9542 | ||
| HCB-H | Equation (9) | —— | —— | —— | —— | —— | —— | 0.4095 | 0.0482 | 1.5897 | |
| Equation (18) | —— | —— | —— | —— | —— | —— | 0.4542 | 0.0383 | 1.5284 | ||
| Equation (27) | 2.49 × 10−5 | 1.61 × 10−4 | —— | −4.30 × 10−1 | —— | —— | 0.4860 | 0.0467 | 1.4832 | ||
| CW-D | Equation (10) | —— | —— | —— | —— | —— | —— | 0.4594 | −0.0012 | 1.4319 | |
| Equation (19) | —— | —— | —— | —— | —— | —— | 0.4782 | −0.0015 | 1.4068 | ||
| Equation (28) | 3.59 × 100 | 3.41 × 100 | 3.47 × 10−13 | 1.00 × 100 | −3.20 × 10−2 | −3.20 × 10−2 | 0.5271 | 0.0032 | 1.3393 | ||
| MH | H-D | Equation (11) | —— | —— | —— | —— | —— | —— | 0.5161 | 0.0008 | 2.4594 |
| Equation (20) | —— | —— | —— | —— | —— | —— | 0.5426 | −0.0021 | 2.3912 | ||
| Equation (29) | 7.43 × 10−2 | —— | —— | —— | —— | —— | 0.6473 | −0.0029 | 2.0999 | ||
| HCB-H | Equation (12) | —— | —— | —— | —— | —— | —— | 0.3018 | −0.0148 | 1.8170 | |
| Equation (21) | —— | —— | —— | —— | —— | —— | 0.3468 | −0.0177 | 1.7575 | ||
| Equation (30) | 3.77 × 10−2 | —— | —— | —— | —— | —— | 0.4282 | 0.0078 | 1.6443 | ||
| CW-D | Equation (13) | —— | —— | —— | —— | —— | —— | 0.5322 | 0.0020 | 0.9734 | |
| Equation (22) | —— | —— | —— | —— | —— | —— | 0.6180 | 0.0054 | 0.8796 | ||
| Equation (31) | 1.10 × 10−2 | 2.76 × 10−3 | —— | −9.95 × 10−1 | —— | —— | 0.6658 | 0.0077 | 0.8228 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhang, H.; Zhang, H.; Yang, T.; Zhang, J.; Liu, Y. A Novel 3D Tree-Modeling Method of Incorporating Small-Scale Spatial Structure Parameters in a Heterogeneous Forest Environment. Forests 2023, 14, 639. https://doi.org/10.3390/f14030639
Wang L, Zhang H, Zhang H, Yang T, Zhang J, Liu Y. A Novel 3D Tree-Modeling Method of Incorporating Small-Scale Spatial Structure Parameters in a Heterogeneous Forest Environment. Forests. 2023; 14(3):639. https://doi.org/10.3390/f14030639
Chicago/Turabian StyleWang, Linlong, Huaiqing Zhang, Huacong Zhang, Tingdong Yang, Jing Zhang, and Yang Liu. 2023. "A Novel 3D Tree-Modeling Method of Incorporating Small-Scale Spatial Structure Parameters in a Heterogeneous Forest Environment" Forests 14, no. 3: 639. https://doi.org/10.3390/f14030639