
Exploring the Potential Applications of the Noninvasive Reporter Gene RUBY in Plant Genetic Transformation
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Construction of the 35S346 bp::RUBY Expression Vector
2.3. Genetic Transformation of RUBY
2.4. Genetic Transformation of 35S346 bp::RUBY Arabidopsis
2.5. RNA Extraction, RT−PCR, and RT−qPCR Analysis
2.6. Extraction and Quantitative Determination of Pigment of RUBY Transgenic Plants and Tissues
3. Results
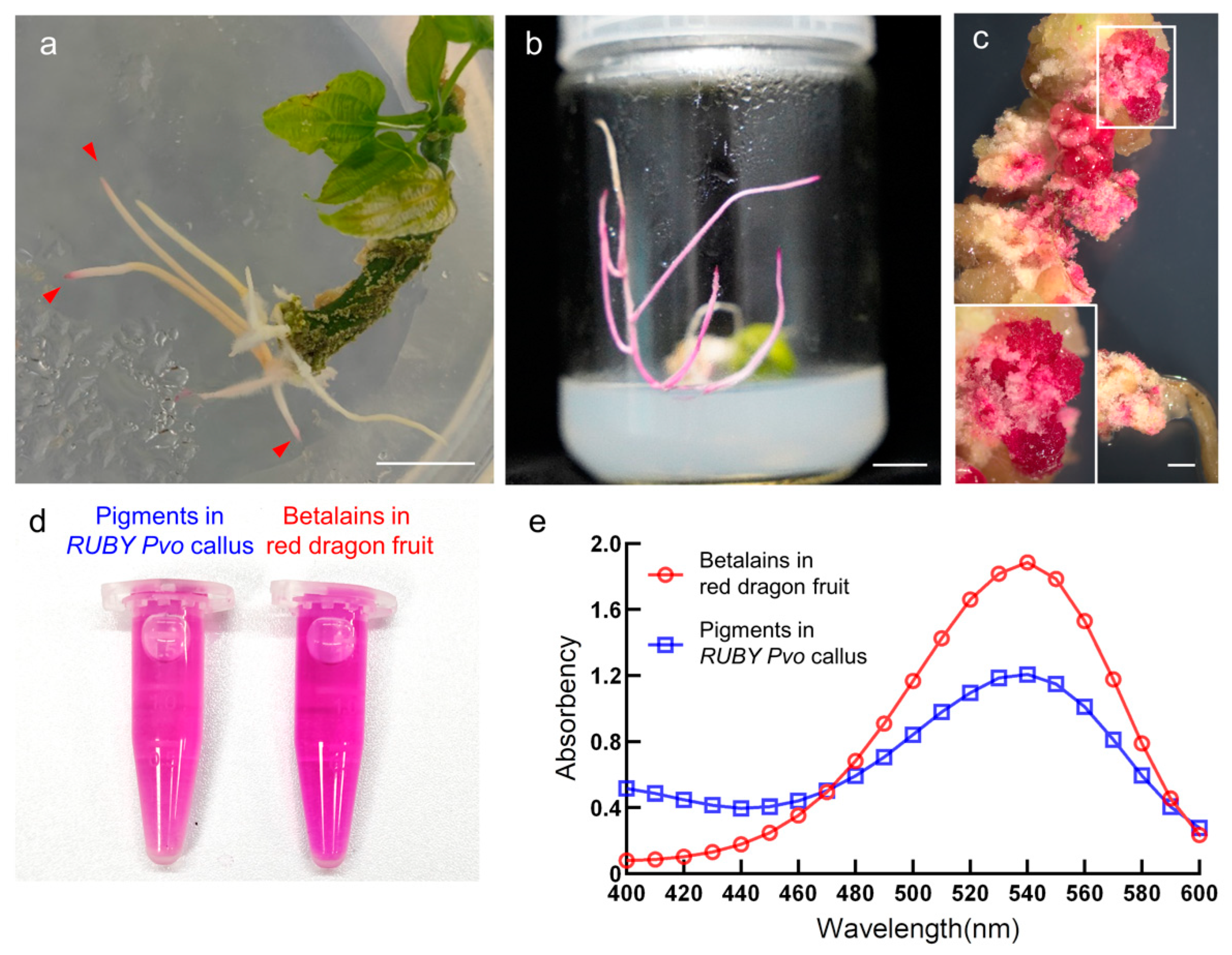
3.1. Ectopic Expression of RUBY Reconstructed the Betalain Biosynthesis Pathway in P. volubilis
3.2. Ectopic Expression of RUBY Has No Adverse Effects on the Growth and Development of Plants
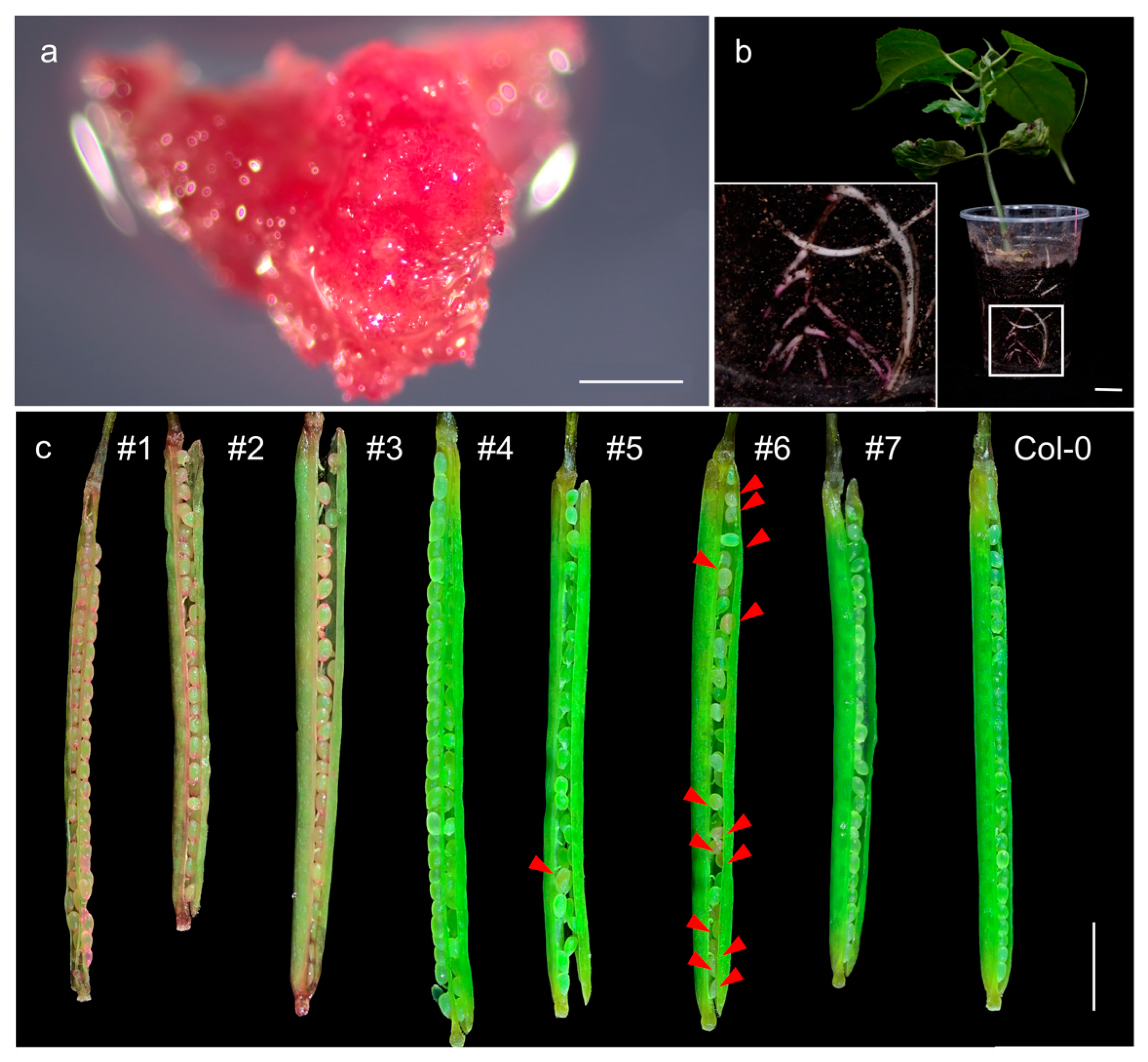
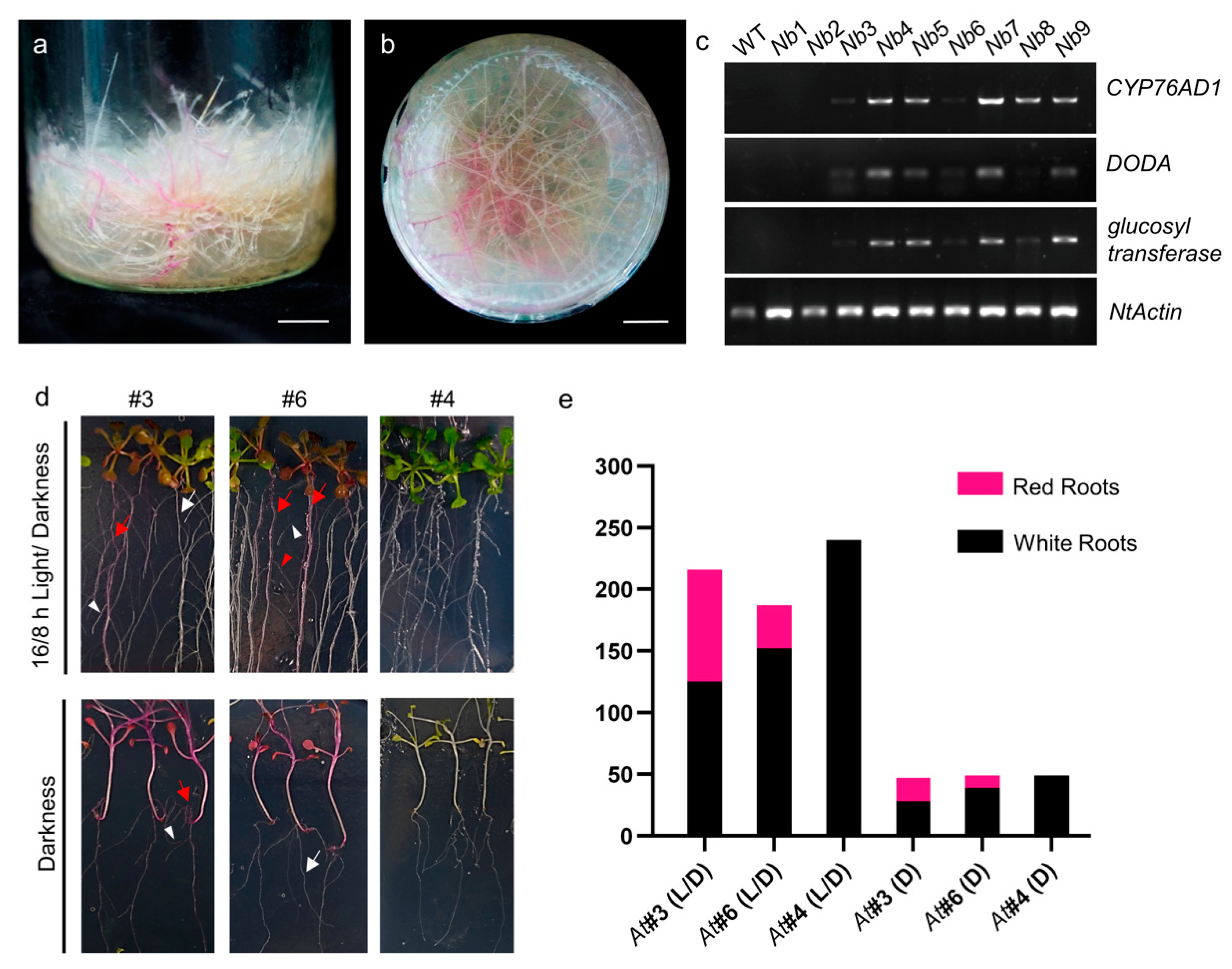
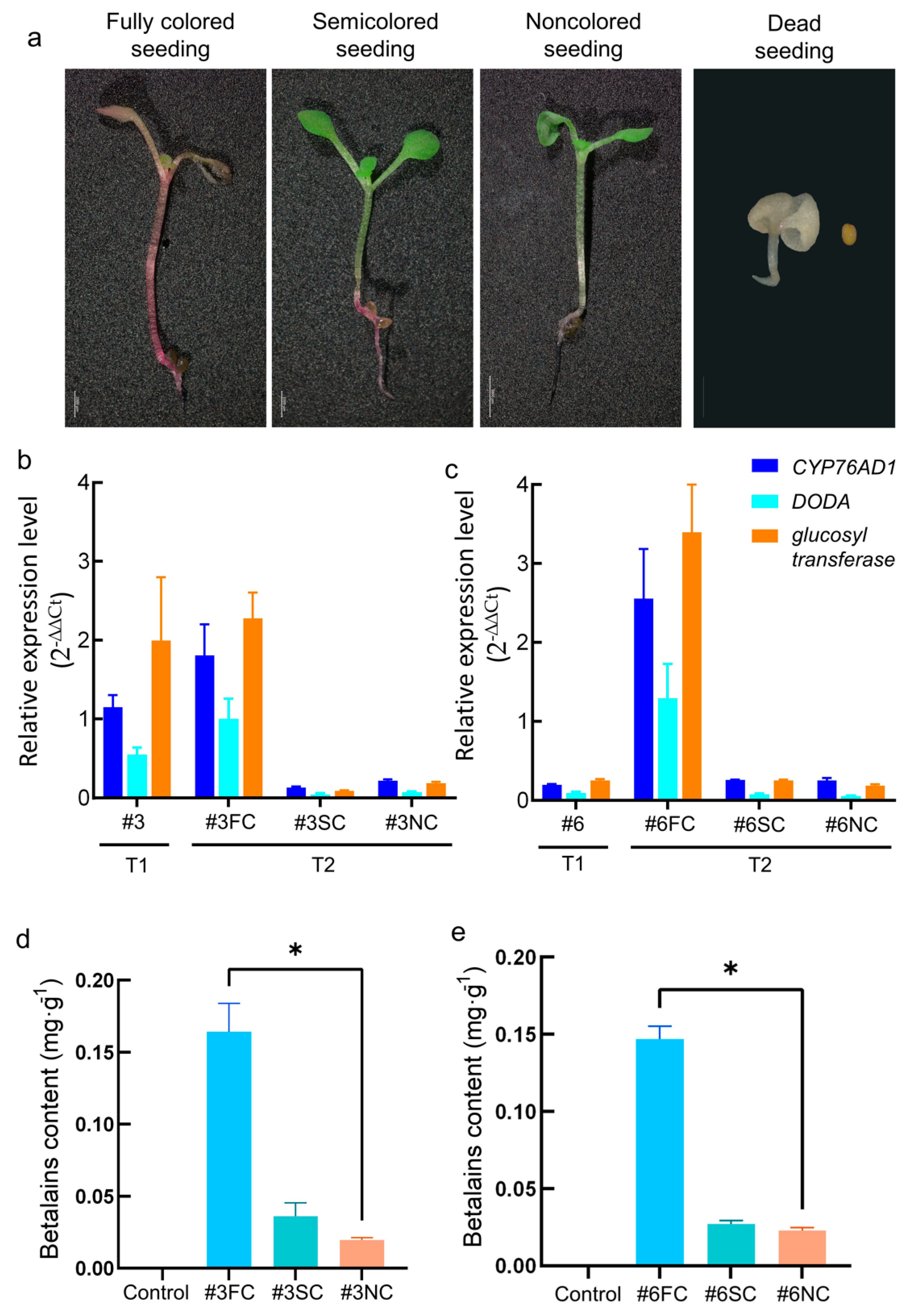
3.3. RUBY Transgenic Hairy Roots and Plants Showed Low Rates of Red Coloration
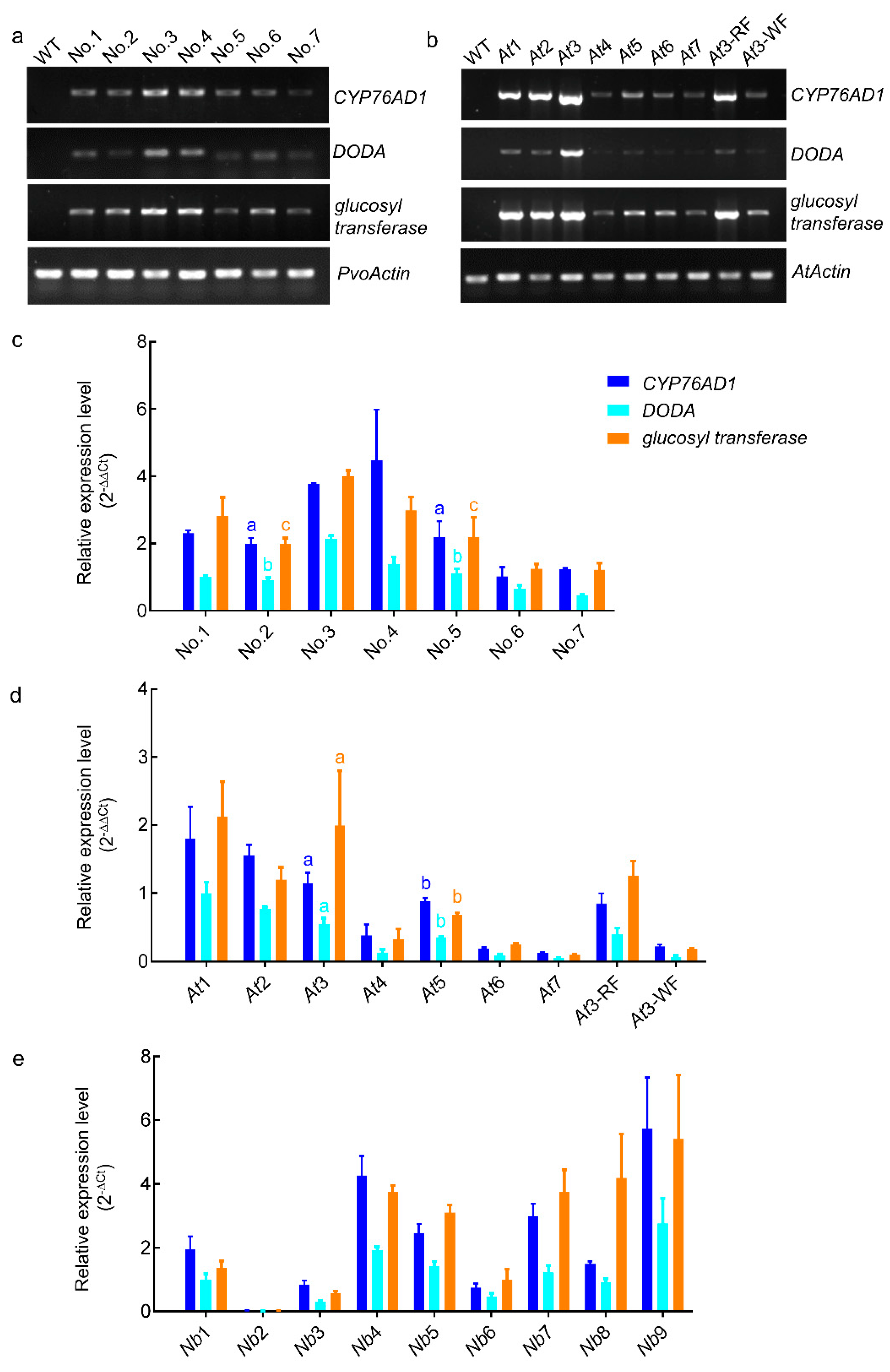
3.4. The Expression Level of RUBY Is Positively Correlated with Red Coloration
3.5. Instability of RUBY Expression in Transgenic Arabidopsis
4. Discussion
4.1. RUBY Has Broad Prospects as a Naked-Eye Visual Screening Reporter
4.2. The Risk of RUBY in Genetic Transformation
4.3. RUBY as a Reporter Needs Further Optimizations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Gandía-Herrero, F.; García-Carmona, F. Biosynthesis of betalains: Yellow and violet plant pigments. Trends Plant Sci. 2013, 18, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Kiryushkin, A.S.; Ilina, E.L.; Guseva, E.D.; Pawlowski, K.; Demchenko, K.N. Hairy CRISPR: Genome editing in plants using hairy root transformation. Plants 2021, 11, 51. [Google Scholar] [CrossRef]
- Timoneda, A.; Yunusov, T.; Quan, C.; Gavrin, A.; Brockington, S.F.; Schornack, S. MycoRed: Betalain pigments enable in vivo real-time visualisation of arbuscular mycorrhizal colonisation. PLoS Biol. 2021, 19, e3001326. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Lin, H.; Li, Q.; Ruan, Y.; Cousins, D.; Li, F.; Gao, S.; Jackson, K.; Wen, J.; Murray, J.D.; et al. Anthocyanin pigmentation as a quantitative visual marker for arbuscular mycorrhizal fungal colonization of Medicago truncatula Roots. New Phytol. 2022, 236, 1988–1998. [Google Scholar] [CrossRef]
- Polturak, G.; Breitel, D.; Grossman, N.; Sarrion-Perdigones, A.; Weithorn, E.; Pliner, M.; Orzaez, D.; Granell, A.; Rogachev, I.; Aharoni, A. Elucidation of the first committed step in betalain biosynthesis enables the heterologous engineering of betalain pigments in plants. New Phytol. 2015, 210, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, X.; Li, J.; Meng, Y.; Zhao, G.R. Improvement of betanin biosynthesis in Saccharomyces cerevisiae by metabolic engineering. Synth. Syst. Biotechnol. 2023, 8, 54–60. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhang, T.; Sun, H.; Zhan, H.; Zhao, Y. A reporter for noninvasively monitoring gene expression and plant transformation. Hortic. Res. 2020, 7, 152. [Google Scholar] [CrossRef]
- Tian, Y.S.; Fu, X.Y.; Yang, Z.Q.; Wang, B.; Gao, J.J.; Wang, M.Q.; Xu, J.; Han, H.J.; Li, Z.J.; Yao, Q.H.; et al. Metabolic engineering of rice endosperm for betanin biosynthesis. New Phytol. 2019, 225, 1915–1922. [Google Scholar] [CrossRef]
- Harris, N.N.; Javellana, J.; Davies, K.M.; Lewis, D.H.; Jameson, P.E.; Deroles, S.C.; Calcott, K.E.; Gould, K.S.; Schwinn, K.E. Betalain production is possible in anthocyanin-producing plant species given the presence of DOPA-dioxygenase and L-DOPA. BMC Plant Biol. 2012, 12, 34. [Google Scholar] [CrossRef] [Green Version]
- Polturak, G.; Grossman, N.; Vela-Corcia, D.; Dong, Y.; Nudel, A.; Pliner, M.; Levy, M.; Rogachev, I.; Aharoni, A. Engineered gray mold resistance, antioxidant capacity, and pigmentation in betalain-producing crops and ornamentals. Proc. Natl. Acad. Sci. USA 2017, 114, 9062–9067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, J.S.; Mabry, T.J.B.A. Pigment evolution in the Caryophyllales: A systematic overview. Bot. Acta 1996, 109, 360–367. [Google Scholar] [CrossRef]
- Grützner, R.; Schubert, R.; Horn, C.; Yang, C.; Vogt, T.; Marillonnet, S. Engineering betalain biosynthesis in tomato for high level betanin production in fruits. Front. Plant Sci. 2021, 12, 682443. [Google Scholar] [CrossRef]
- Alagarsamy, K.; Shamala, L.F.; Wei, S. Protocol: High-efficiency in-planta Agrobacterium-mediated transgenic hairy root induction of Camellia sinensis Var. sinensis. Plant Methods 2018, 14, 17. [Google Scholar] [CrossRef]
- Chen, K.; Hu, K.; Xi, F.; Wang, H.; Kohnen, M.V.; Gao, P.; Liao, J.; Wei, W.; Liu, X.; Zhang, H.; et al. High-efficient and transient transformation of Moso Bamboo (Phyllostachys edulis) and Ma Bamboo (Dendrocalamus latiflorus Munro). J. Plant Biol. 2023, 66, 75–86. [Google Scholar] [CrossRef]
- Cao, X.; Xie, H.; Song, M.; Lu, J.; Ma, P.; Huang, B.; Wang, M.; Tian, Y.; Chen, F.; Peng, J.; et al. Cut–dip–budding delivery system enables genetic modifications in plants without tissue culture. Innovation 2023, 4, 100345. [Google Scholar] [CrossRef]
- Mei, G.; Chen, A.; Wang, Y.; Li, S.; Wu, M.; Liu, X.; Hou, X. A simple and efficient in planta transformation method based on the active regeneration capacity of plants. bioRxiv 2023. [Google Scholar] [CrossRef]
- Ge, X.; Wang, P.; Wang, Y.; Wei, X.; Chen, Y.; Li, F. Development of an eco-friendly pink cotton germplasm by engineering betalain biosynthesis pathway. Plant Biotechnol. J. 2023, in press. [Google Scholar] [CrossRef]
- Kodahl, N. Sacha inchi (Plukenetia volubilis L.)-from lost crop of the Incas to part of the solution to global challenges? Planta 2020, 251, 80. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, F.; Kakuda, Y. Sacha inchi (Plukenetia volubilis L.): Nutritional composition, biological activity, and uses. Food Chem. 2018, 265, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Ángel, M.R.C.; German, G.A.; María, F.S.M.; Kodahl, N. Ethnobotanical knowledge in the peruvian amazon of the neglected and underutilized crop Sacha Inchi (Plukenetia volubilis L.). Econ. Bot. 2019, 73, 281–287. [Google Scholar]
- Fu, Q.; Niu, L.; Zhang, Q.; Pan, B.Z.; He, H.; Xu, Z.F. Benzyladenine treatment promotes floral feminization and fruiting in a promising oilseed crop Plukenetia volubilis. Ind. Crops Prod. 2014, 59, 295–298. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Niu, L.; Chen, M.S.; Tao, Y.B.; Wang, X.; He, H.; Pan, B.Z.; Xu, Z.F. De novo transcriptome assembly and comparative analysis between male and benzyladenine-induced female inflorescence buds of Plukenetia Volubilis. J. Plant Physiol. 2018, 221, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.D.; Pan, B.Z.; Fu, Q.; Niu, L.; Chen, M.S.; Xu, Z.F. De novo transcriptome assembly of the eight major organs of Sacha Inchi (Plukenetia Volubilis) and the identification of genes involved in α-linolenic acid metabolism. BMC Genom. 2018, 19, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kereszt, A.; Li, D.; Indrasumunar, A.; Nguyen, C.D.T.; Nontachaiyapoom, S.; Kinkema, M.; Gresshoff, P.M. Agrobacterium rhizogenes-mediated transformation of soybean to study root biology. Nat. Protoc. 2007, 2, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Yang, Q.; Dong, B.; Song, Z.; Niu, L.; Wang, L.; Cao, H.; Li, H.; Fu, Y. Development of an efficient root transgenic system for pigeon pea and its application to other important economically plants. Plant Biotechnol. J. 2019, 17, 1804–1813. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.L.; Liu, C.; Piao, C.L.; Liu, C.L. A stable Agrobacterium rhizogenes-mediated transformation of cotton (Gossypium hirsutum L.) and plant regeneration from transformed hairy root via embryogenesis. Front. Plant Sci. 2020, 11, 604255. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, D.; Fu, J.; Zhang, Z.; Qin, Y.; Hu, G.; Zhao, J. Agrobacterium rhizogenes-mediated hairy root transformation as an efficient system for gene function analysis in Litchi chinensis. Plant Methods 2021, 17, 103. [Google Scholar] [CrossRef]
- Maher, M.; Nasti, R.; Vollbrecht, M.; Starker, C.; Clark, M.; Voytas, D. Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 2020, 38, 84–89. [Google Scholar] [CrossRef]
- Odell, J.T.; Nagy, F.; Chua, N.H. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature 1985, 313, 810–812. [Google Scholar] [CrossRef]
- Holsters, M.; de Waele, D.; Depicker, A.; Messens, E.; van Montagu, M.; Schell, J. Transfection and transformation of Agrobacterium tumefaciens. Mol. Genet. Genom. 1978, 163, 181–187. [Google Scholar] [CrossRef]
- Fan, Y.L.; Zhang, X.H.; Zhong, L.J.; Wang, X.Y.; Jin, L.S.; Lyu, S.H. One-step generation of composite soybean plants with transgenic roots by Agrobacterium rhizogenes-mediated transformation. BMC Plant Biol. 2020, 20, 208. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Ding, M.; Nakano, M.; Hong, K.; Huang, R.; Becker, D.; Glazebrook, J.; Katagiri, F.; Han, X.; Tsuda, K. Letter to the editor: DNA purification-free PCR from plant tissues. Plant Cell Physiol. 2021, 62, 1503–1505. [Google Scholar] [CrossRef] [PubMed]
- Arocho, A.; Chen, B.; Ladanyi, M.; Pan, Q. Validation of the 2-DeltaDeltaCt calculation as an alternate method of data analysis for quantitative PCR of BCR-ABL P210 transcripts. Diagn. Mol. Pathol. 2006, 15, 56–61. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, M.; Corke, H. Identification and distribution of simple and acylated betacyanins in the Amaranthaceae. J. Agric. Food Chem. 2001, 49, 1971–1978. [Google Scholar] [CrossRef]
- Herbach, K.M.; Maier, C.; Stintzing, F.C.; Carle, R. Effects of processing and storage on juice colour and betacyanin stability of purple pitaya (Hylocereus polyrhizus) juice. Eur. Food Res. Technol. 2007, 224, 649–658. [Google Scholar] [CrossRef]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Gene Dev. 2006, 20, 1790–1799. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yordanov, Y.S.; Ma, C.; Strauss, S.; Busov, V.B. DR5 as a reporter system to study auxin response in Populus. Plant Cell Rep. 2013, 32, 453–463. [Google Scholar] [CrossRef]
- Choo, K.Y.; Ong, Y.Y.; Lim, R.L.H.; Tan, C.P.; Ho, C.W. Study on bioaccessibility of betacyanins from red dragon fruit (Hylocereus Polyrhizus). Food Sci. Biotechnol. 2019, 28, 1163–1169. [Google Scholar] [CrossRef]
- Piątczak, E.; Kuźma, Ł.; Skała, E.; Żebrowska, M.; Balcerczak, E.; Wysokińska, H. Iridoid and phenylethanoid glycoside production and phenotypical changes in plants regenerated from hairy roots of Rehmannia glutinosa Libosch. Plant Cell Tissue Organ Cult. 2015, 122, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Boobalan, S.; Kamalanathan, D. Tailoring enhanced production of aervine in Aerva lanata (L.) Juss. Ex Schult by Agrobacterium rhizogenes- mediated hairy root cultures. Ind. Crop. Prod. 2020, 155, 112814. [Google Scholar] [CrossRef]
- Piątczak, E.; Jeleń, A.; Makowczyńska, J.; Zielińska, S.; Kuźma, Ł.; Balcerczak, E. Establishment of hairy root cultures of Rehmannia elata N.E. Brown ex Prain and production of iridoid and phenylethanoid glycosides. Ind. Crop. Prod. 2019, 137, 308–314. [Google Scholar] [CrossRef]
- Osakabe, Y.; Sugano, S.S.; Osakabe, K. Genome engineering of woody plants: Past, present and future. J. Wood Sci. 2016, 62, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Habibi, P.; de Sa, M.F.G.; da Silva, A.L.L.; Makhzoum, A.; da Costa, J.L.; Borghetti, I.A.; Soccol, C.R. Efficient genetic transformation and regeneration system from hairy root of Origanum vulgare. Physiol. Mol. Biol. Plants 2016, 22, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Gantait, S.; Mukherjee, E. Hairy root culture technology: Applications, constraints and prospect. Appl. Microbiol. Biot. 2021, 105, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Velini, E.D.; Trindade, M.; Duke, B.S.O. Growth regulation and other secondary effects of herbicides. Weed Sci. 2010, 58, 351–354. [Google Scholar] [CrossRef]
- Wang, X.; Ryu, D.; Houtkooper, R.H.; Auwerx, J. Antibiotic use and abuse: A threat to mitochondria and chloroplasts with impact on research, health, and environment. BioEssays 2015, 37, 1045–1053. [Google Scholar] [CrossRef]
- Liang, J.; He, J. Protective role of anthocyanins in plants under low nitrogen stress. Biochem. Biophys. Res. Commun. 2018, 498, 946–953. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Mitiouchkina, T.; Mishin, A.; Somermeyer, L.; Markina, N.; Chepurnyh, T.; Guglya, E.; Karataeva, T.; Palkina, K.; Shakhova, E.; Fakhranurova, L.; et al. Plants with genetically encoded autoluminescence. Nat. Biotechnol. 2020, 38, 1000–1001. [Google Scholar] [CrossRef]
- Zhu, Q.; Yu, S.; Zeng, D.; Liu, H.; Wang, H.; Yang, Z.; Xie, X.; Shen, R.; Tan, J.; Li, H.; et al. Development of “Purple Endosperm Rice” by engineering anthocyanin biosynthesis in the endosperm with a high-efficiency transgene stacking system. Mol. Plant 2017, 10, 918–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Wu, H.; Zhu, H.; Huang, C.; Liu, C.; Chang, Y.; Kong, Z.; Zhou, Z.; Wang, G.; Lin, Y.; et al. Determining factors, regulation system, and domestication of anthocyanin biosynthesis in rice leaves. New Phytol. 2019, 223, 705–721. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhu, M.; Wu, J.; Ouyang, L.; Wang, R.; Sun, H.; Yan, L.; Wang, L.; Xu, M.; Zhan, H.; et al. Repurposing of anthocyanin biosynthesis for plant transformation and genome editing. Front. Genome Ed. 2020, 2, 607982. [Google Scholar] [CrossRef] [PubMed]
- Sunilkumar, G.; Mohr, L.A.; Lopata-Finch, E.; Emani, C.; Rathore, K.S. Developmental and tissue-specific expression of CaMV35S promoter in cotton as revealed by GFP. Plant Mol. Biol. 2002, 50, 463–474. [Google Scholar] [CrossRef]
- Amack, S.C.; Antunes, M.S. CaMV35S promoter—A plant biology and biotechnology workhorse in the era of synthetic biology. Curr. Plant Biol. 2020, 24, 100179. [Google Scholar] [CrossRef]
- Zhao, S.Z.; Sun, H.Z.; Chen, M.; Wang, B.S. Light-regulated betacyanin accumulation in Euhalophyte Suaeda Salsa Calli. Plant Cell Tissue Organ Cult. 2010, 102, 99–107. [Google Scholar] [CrossRef]
- Shin, K.S.; Murthy, H.N.; Heo, J.W.; Paek, K.Y. Induction of betalain pigmentation in hairy roots of red beet under different radiation sources. Biol. Plant. 2003, 47, 149–152. [Google Scholar] [CrossRef]
- Zheng, X.; Liu, S.; Cheng, C.; Guo, R.; Chen, Y.; Xie, L.; Mao, Y.; Lin, Y.; Zhang, Z.; Lai, Z. Cloning and expression analysis of betalain biosynthesis genes in Amaranthus Tricolor. Biotechnol. Lett. 2016, 38, 723–729. [Google Scholar] [CrossRef]
- Kiselev, K.; Aleynova, O.; Ogneva, Z.; Suprun, A.; Dubrovina, A. 35S promoter-driven transgenes are variably expressed in different organs of Arabidopsis thaliana and in response to abiotic stress. Mol. Biol. Rep. 2021, 48, 2235–2241. [Google Scholar] [CrossRef]
- Jefferson, R.A. The GUS reporter gene system. Nature 1989, 342, 837–838. [Google Scholar] [CrossRef]
- Koo, J.; Kim, Y.; Kim, J.; Yeom, M.; Lee, I.C.; Nam, H.G. A GUS/Luciferase fusion reporter for plant gene trapping and for assay of promoter activity with luciferin-dependent control of the reporter protein stability. Plant Cell Physiol. 2007, 48, 1121–1131. [Google Scholar] [CrossRef] [Green Version]
- Khidr, Y.A.; Flachowsky, H.; Haselmair-Gosch, C.; Thill, J.; Miosic, S.; Hanke, M.V.; Stich, K.; Halbwirth, H. Evaluation of a MdMYB10/GFP43 fusion gene for its suitability to act as reporter gene in promoter studies in Fragaria vesca L. ‘Rügen’. Plant Cell Tissue Organ Cult. 2017, 130, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Ramírez, E.; Lima, E.; Guzmán, A. Natural betalains supported on γ-alumina: A wide family of stable pigments. Dye. Pigment. 2015, 120, 161–168. [Google Scholar] [CrossRef]
- Khan, M.I. Stabilization of Betalains: A Review. Food Chem. 2016, 197, 1280–1285. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, Y.; Makita, Y.; Kawauchi, M.; Kageyama, A.; Kuriyama, T.; Matsui, M. Intergenic splicing-stimulated transcriptional readthrough is suppressed by nonsense-mediated mRNA decay in Arabidopsis. Commun. Biol. 2022, 5, 1390. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 35S::RUBY Col-0 T2 | Total Number of Seedlings | Surviving Seedlings (%) | Dead Seedlings (%) | The Ratio of Surviving and Dead Seedlings | Chi-Test (p-Value) | ||
|---|---|---|---|---|---|---|---|
| Fully Colored Seedlings (%) | Semicolored Seedlings (%) | Noncolored Seedlings (%) | |||||
| #1 | 420 | 63.45 ± 2.99 | 11.44 ± 0.17 | 0.22 ± 0.22 | 24.89 ± 1.56 | 3.04 ± 0.43:1 | 0.97 |
| #2 | 676 | 73.31 ± 3.90 | 6.36 ± 1.01 | 0.00 ± 0.00 | 20.34 ± 2.81 | 4.09 ± 1.08:1 | 0.30 |
| #3 | 481 | 59.10 ± 1.88 | 19.10 ± 1.84 | 0.19 ± 0.19 | 21.64 ± 0.18 | 3.62 ± 0.07:1 | 0.44 |
| #4 | 445 | 0.00 ± 0.00 | 0.00 ± 0.00 | 75.13 ± 2.02 | 24.51 ± 2.36 | 3.08 ± 0.6:1 | 0.99 |
| #5 | 521 | 0.97 ± 0.37 | 0.00 ± 0.00 | 78.25 ± 1.15 | 20.77 ± 0.10 | 3.84 ± 0.39:1 | 0.80 |
| #6 | 676 | 17.51 ± 4.17 | 33.40 ± 5.15 | 26.78 ± 2.04 | 22.31 ± 1.60 | 3.53 ± 0.56:1 | 0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Deng, S.; Huang, H.; Mo, J.; Xu, Z.-F.; Wang, Y. Exploring the Potential Applications of the Noninvasive Reporter Gene RUBY in Plant Genetic Transformation. Forests 2023, 14, 637. https://doi.org/10.3390/f14030637
Yu J, Deng S, Huang H, Mo J, Xu Z-F, Wang Y. Exploring the Potential Applications of the Noninvasive Reporter Gene RUBY in Plant Genetic Transformation. Forests. 2023; 14(3):637. https://doi.org/10.3390/f14030637
Chicago/Turabian StyleYu, Jingjing, Shiling Deng, Han Huang, Jinhui Mo, Zeng-Fu Xu, and Yi Wang. 2023. "Exploring the Potential Applications of the Noninvasive Reporter Gene RUBY in Plant Genetic Transformation" Forests 14, no. 3: 637. https://doi.org/10.3390/f14030637