
Nematodes Consume Four Species of a Common, Wood-Decay Fungus


Abstract
:1. Introduction
2. Materials and Methods
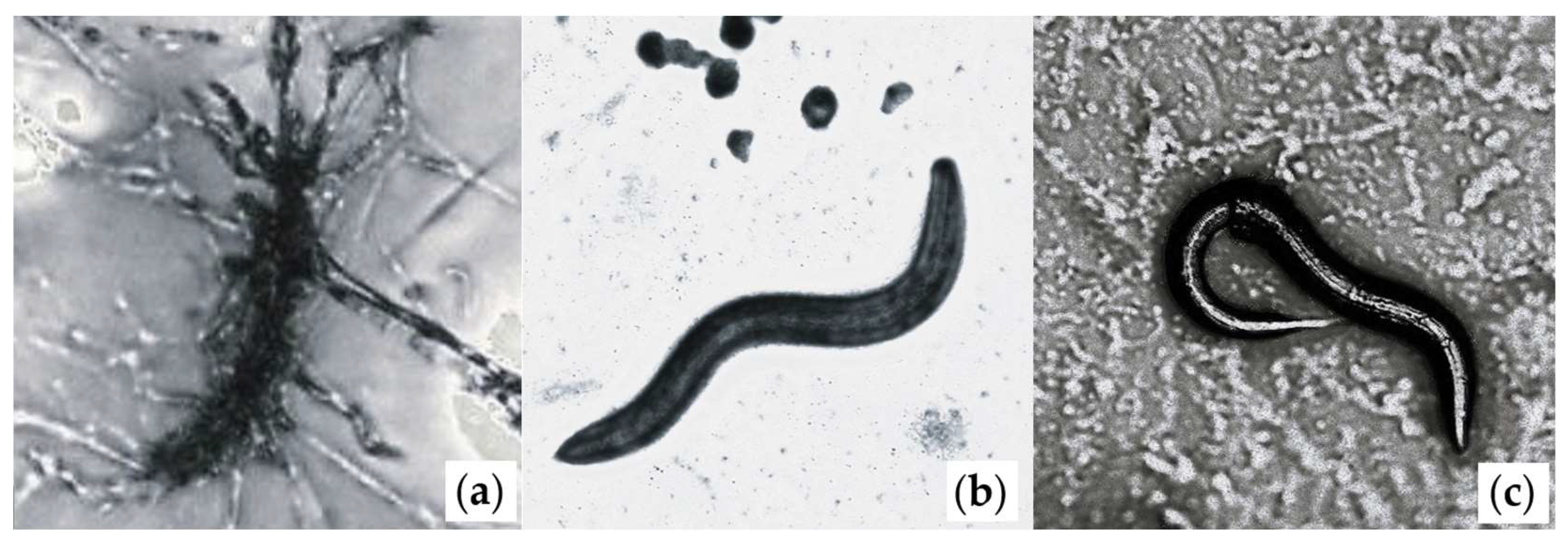
2.1. Fungal Cultures
2.2. Nematode Cultures
2.3. Fomitopsis–Nematode Interaction Experiments
2.4. Data Collection
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| F. mounceae | F. ochracea | F. pinicola | F. schrenkii | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CAN | OR | MI | WA | SE.1 | SE.2 | AZ | CO.2 | F | N | - | |
| 4. | N | N | N | N | N | N | N | - | 0 | 7 | 1 |
| 5. | - | N | N | F | N | N | N | N | 1 | 6 | 1 |
| 6. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 7. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 11. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 12. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 13. | N | N | N | N | N | - | N | N | 0 | 7 | 1 |
| 17. | N | N | N | N | N | N | N | 0 | 7 | 0 | |
| 18. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 21. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 23. | N | N | N | N | N | - | - | N | 0 | 8 | 0 |
| 24. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| F | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | ||
| N | 11 | 12 | 12 | 11 | 12 | 11 | 11 | 11 | 91 | ||
| - | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 3 | ||
| F. mounceae | F. ochracea | F. pinicola | F. schrenkii | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| NH | WI | AK | NH | EE | RU | CO.1 | SD | F | N | - | |
| 1. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 3. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 4. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 5. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 6. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 7. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 10. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 11. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 12. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 13. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 14. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 15. | N | N | N | - | N | N | N | N | 0 | 7 | 1 |
| 16. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 20. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 21. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 23. | N | N | N | N | N | N | - | N | 0 | 7 | 1 |
| 24. | N | N | N | N | N | N | N | 0 | 7 | 0 | |
| F | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| N | 16 | 17 | 17 | 16 | 17 | 17 | 16 | 17 | 133 | ||
| - | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 2 | ||
| F. mounceae | F. ochracea | F. pinicola | F. schrenkii | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| NH | WI | AK | NH | EE | RU | CO.1 | SD | F | N | - | |
| 1. | N | N | N | N | N | N | 0 | 6 | 0 | ||
| 4. | N | N | N | N | N | N | N | 0 | 7 | 0 | |
| 5. | N | N | N | N | N | N | N | N | 0 | 8 | 0 |
| 7. | N | N | N | N | N | N | N | 0 | 7 | 0 | |
| 10. | N | N | N | N | N | N | - | 0 | 6 | 1 | |
| 12. | N | N | N | F | N | N | N | N | 1 | 7 | 0 |
| 13. | N | N | N | N | N | N | - | 0 | 6 | 1 | |
| 14. | N | N | N | N | - | N | - | N | 0 | 6 | 2 |
| 15. | N | N | N | N | N | N | N | 0 | 7 | 0 | |
| 20. | N | N | N | N | N | N | N | 0 | 7 | 0 | |
| 21. | N | N | N | F | N | N | N | N | 1 | 7 | 0 |
| 23. | N | N | N | F | N | N | F | 2 | 5 | 0 | |
| 24. | N | N | N | N | N | N | F | 1 | 6 | 0 | |
| F | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 2 | 5 | ||
| N | 12 | 13 | 12 | 10 | 12 | 5 | 12 | 9 | 85 | ||
| - | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 2 | 4 | ||
References
- van den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berbee, M.L.; Strullu-Derrien, C.; Delaux, P.-M.; Strother, P.K.; Kenrick, P.; Selosse, M.-A.; Taylor, J.W. Genomic and fossil windows into the secret lives of the most ancient fungi. Nat. Rev. Microbiol. 2020, 18, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Barron, G.L.; Thorn, R.G. Destruction of nematodes by species of Pleurotus. Can. J. Bot. 1987, 65, 774–778. [Google Scholar] [CrossRef]
- Marlin, M.; Wolf, A.; Alomran, M.; Carta, L.; Newcombe, G. Nematophagous Pleurotus species consume some nematode species but are themselves consumed by others. Forests 2019, 10, 404. [Google Scholar] [CrossRef] [Green Version]
- Balaes, T.; Tănase, C. Basidiomycetes as potential biocontrol agents against nematodes. Rom. Biotechnol. Lett. 2016, 21, 11185–11193. [Google Scholar]
- Blaxter, M.; Koutsovoulos, G. The evolution of parasitism in Nematoda. Parasitology 2015, 142, S26–S39. [Google Scholar] [CrossRef] [Green Version]
- Catalogue of Life. Available online: https://www.catalogueoflife.org/data/taxon/F (accessed on 10 November 2021).
- van Ooij, C. Hungry fungus eats nematode. Nat. Rev. Microbiol. 2011, 9, 766–767. [Google Scholar] [CrossRef]
- Thorn, R.G.; Barron, G.L. Carnivorous mushrooms. Science 1984, 224, 76–78. [Google Scholar] [CrossRef] [Green Version]
- Högberg, N.; Holdenrieder, O.; Stenlid, J. Population structure of the wood decay fungus Fomitopsis pinicola. Heredity 1999, 83, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Balaes, T.; Boroş, L.; Şesan, T.; Tănase, C. Basidiomycete-based method for biocontrol of phytopathogenic nematodes. J. Plant Dev. 2015, 22, 83–94. [Google Scholar]
- de Freitas Soares, F.E.; Sufiate, B.L.; de Queiroz, J.H. Nematophagous fungi: Far beyond the endoparasite, predator and ovicidal groups. Agric. Nat. Resour. 2018, 52, 1–8. [Google Scholar] [CrossRef]
- Haight, J.-E.; Laursen, G.A.; Glaeser, J.A.; Taylor, D.L. Phylogeny of Fomitopsis pinicola: A species complex. Mycologia 2016, 108, 925–938. [Google Scholar] [CrossRef] [Green Version]
- Haight, J.-E.; Nakasone, K.K.; Laursen, G.A.; Redhead, S.A.; Taylor, D.L.; Glaeser, J.A. Fomitopsis mounceae and F. schrenkii—Two new species from North America in the F. pinicola complex. Mycologia 2019, 111, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Lee, Y.Y.; Chang, Y.C.; Pon, W.L.; Lee, S.P.; Wali, N.; Nakazawa, T.; Honda, Y.; Shie, J.J.; Hsueh, Y.P. A carnivorous mushroom paralyzes and kills nematodes via a volatile ketone. Sci. Adv. 2023, 9, 4809. [Google Scholar] [CrossRef]
- Barron, G.L. Microcolonies of bacteria as a nutrient source for lignicolous and other fungi. Can. J. Bot. 1988, 66, 2505–2510. [Google Scholar] [CrossRef]
- Carta, L.K. Bacterial-feeding nematode growth and preference for biocontrol isolates of the bacterium Burkholderia cepacia. J. Nematol. 2000, 32, 362–369. [Google Scholar]
- Maynard, D.S.; Covey, K.R.; Crowther, T.W.; Sokol, N.W.; Morrison, E.W.; Frey, S.D.; Van Diepen, L.T.A.; Bradford, M.A. Species associations overwhelm abiotic conditions to dictate the structure and function of wood-decay fungal communities. Ecology 2018, 99, 801–811. [Google Scholar] [CrossRef]
- Barron, G.L. Predatory fungi, wood decay, and the carbon cycle. Biodiversity 2003, 4, 3–9. [Google Scholar] [CrossRef]
- Boddy, L.; Jones, T.H. Interactions between basidiomycota and invertebrates. In British Mycological Society Symposia Series; Boddy, L., Frankland, J.C., van West, P., Eds.; Academic Press: New York, NY, USA, 2008; Volume 28, pp. 155–179. [Google Scholar] [CrossRef]
- Crowther, T.W.; Boddy, L.; Jones, T.H. Outcomes of fungal interactions are determined by soil invertebrate grazers. Ecol. Lett. 2011, 14, 1134–1142. [Google Scholar] [CrossRef]
- Bradford, M.A.; Warren II, R.J.; Baldrian, P.; Crowther, T.W.; Maynard, D.S.; Oldfield, E.E.; Wieder, W.R.; Wood, S.A.; King, J.R. Climate fails to predict wood decomposition at regional scales. Nat. Clim. Chang. 2014, 4, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Moll, J.; Roy, F.; Bässler, C.; Heilmann-Clausen, J.; Hofrichter, M.; Kellner, H.; Krabel, D.; Schmidt, J.H.; Buscot, F.; Hoppe, B. First evidence that nematode communities in deadwood are related to tree species identity and to co-occurring fungi and prokaryotes. Microorganisms 2021, 9, 1454. [Google Scholar] [CrossRef] [PubMed]
- Hiscox, J.; O’leary, J.; Boddy, L. Fungus wars: Basidiomycete battles in wood decay. Stud. Mycol. 2018, 89, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Ishizaki, T.; Nomura, N.; Watanabe, K. Screening of mushrooms for nematophagous activity against the pinewood nematode, Bursaphelenchus xylophilus. Nematol. Res. 2015, 45, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Han, M.L.; Xu, T.M.; Wang, Y.; Wu, D.M.; Cui, B.K. Taxonomy and phylogeny of the Fomitopsis pinicola complex with descriptions of six new species from east Asia. Front. Microbiol. 2021, 12, 644979. [Google Scholar] [CrossRef]
- Reale, J. The Impact of a Fungus-Feeding Nematode (Aphelenchoides sp.) on Decomposition of Trembling Aspen Wood by Various Wood-Decay Fungi. Bachelor’s Thesis, Lakehead University, Thunder Bay, ON, Canada, 2018. [Google Scholar]
- Consiglio, G.; Setti, L.; Thorn, R.G. New species of Hohenbuehelia, with comments on the Hohenbuehelia atrocoerulea–Nematoctonus robustus species complex. Pers.-Mol. Phylogeny Evol. Fungi 2018, 41, 202–212. [Google Scholar] [CrossRef]
- De la Cruz, R.G.; Knudsen, G.R.; Carta, L.K.; Newcombe, G. Either low inoculum or a multi-trophic interaction can reduce the ability of Sclerotinia sclerotiorum to kill an invasive plant. Rhizosphere 2018, 5, 76–80. [Google Scholar] [CrossRef]
- Baynes, M.A.; Russell, D.M.; Newcombe, G.; Carta, L.K.; Rossman, A.Y.; Ismaiel, A. A mutualistic interaction between a fungivorous nematode and a fungus within the endophytic community of Bromus tectorum. Fungal Ecol. 2012, 5, 610–623. [Google Scholar] [CrossRef]
- Jaffee, B.A. Wood, nematodes, and the nematode-trapping fungus Arthrobotrys oligospora. Soil Biol. Biochem. 2004, 36, 1171–1178. [Google Scholar] [CrossRef]

| Unique I.D. | Genus | Species | Country | State | Collection Number | Experiment |
|---|---|---|---|---|---|---|
| F.m.CAN | Fomitopsis | mounceae | Canada | Alberta Province | JEH-82 | I,II |
| F.m.NH | Fomitopsis | mounceae | USA | New Hampshire | FP-125086-T | I,III,IV |
| F.m.OR | Fomitopsis | mounceae | USA | Oregon | CS-1 | I,II |
| F.m.WI | Fomitopsis | mounceae | USA | Wisconsin | JEH-147 | I,III,IV |
| F.o.AK | Fomitopsis | ochracea | USA | Alaska | LT-16 | I,III,IV |
| F.o.MI | Fomitopsis | ochracea | USA | Michigan | HHB-3331-Sp | I,II |
| F.o.NH | Fomitopsis | ochracea | USA | New Hampshire | FP-125083-T | I,III,IV |
| F.o.WA | Fomitopsis | ochracea | USA | Washington | HHB-14787-T | I,II |
| F.p.EE | Fomitopsis | pinicola | Estonia | Ida-Viru | DR-EST-11 | I,III,IV |
| F.p.RU | Fomitopsis | pinicola | Russia | Moscow | TS-Fp-24 | I,III,IV |
| F.p.SE.1 | Fomitopsis | pinicola | Sweden | Östergötland | AT-Fp-1 | I,II |
| F.p.SE.2 | Fomitopsis | pinicola | Sweden | - | FCUG-2034 | I,II |
| F.s.AZ | Fomitopsis | schrenkii | USA | Arizona | RLG-10752-Sp | I,II |
| F.s.CO.1 | Fomitopsis | schrenkii | USA | Colorado | FP-105881-R | I,III,IV |
| F.s.CO.2 | Fomitopsis | schrenkii | USA | Colorado | JW-F.p.1 | I,II |
| F.s.SD | Fomitopsis | schrenkii | USA | South Dakota | JEH-150 | I,III,IV |
| Unique I.D. | Family | Genus | Species | Collection Number | Experiment |
|---|---|---|---|---|---|
| 1. | Cephalobidae | Acrobeloides | amurensis | PS1146 | I,III,IV |
| 2. | Cephalobidae | Acrobeloides | apiculatus | LKC60 | I |
| 3. | Cephalobidae | Acrobeloides | varius (nanus) | LKC27 | III |
| 4. | Cephalobidae | Acrobeloides | varius (nanus) | LKC52 | I,II,III,IV |
| 5. | Cephalobidae | Zeldia | punctata | PS1192 | II,III,IV |
| 6. | Cephalobidae | Zeldia | sp. | PS1194 | II,III |
| 7. | Diplogasteridae | Pristionchus | aerivorus | LKC54 | I,II,III,IV |
| 8. | Diplogasteridae | Pristionchus | entomophagus | KR2984 | I |
| 9. | Panagrolamidae | Panagrellus | redivivus | PS1163 | I |
| 10. | Panagrolamidae | Panagrolaimus | detritophagus | LKC56 | III,IV |
| 11. | Panagrolamidae | Panagrolaimus | detritophagus | PS1162 | II,III |
| 12. | Panagrolamidae | Panagrolaimus | hygrophilus | PS1732 | II,III,IV |
| 13. | Panagrolamidae | Panagrolaimus | rigidus | LKC53 | II,III,IV |
| 14. | Panagrolamidae | Panagrolaimus | sp. | LKC39 | III,IV |
| 15. | Panagrolamidae | Panagrolaimus | sp. | LKC40 | III,IV |
| 16. | Panagrolamidae | Panagrolaimus | sp. | LKC46 | III |
| 17. | Rhabditidae | Caenorhabditis | elegans | N2 | I,II |
| 18. | Rhabditidae | Diploscapter | lycostoma | PS2017 | II |
| 19. | Rhabditidae | Mesorhabditis | inarimensis | LKC51 | I |
| 20. | Rhabditidae | Metarhabditis | rainai | LKC20 | III,IV |
| 21. | Rhabditidae | Oscheius | dolichura | LKC50 | I,II,III,IV |
| 22. | Rhabditidae | Oscheius | myriophila | DF5020 | I |
| 23. | Rhabditidae | Oscheius | tipulae | LKC57 | I,II,III,IV |
| 24. | Rhabditidae | Poikilolaimus | oxycercus | LKC64 | I,II,III,IV |
| F. mounceae | F. ochracea | F. pinicola | F. schrenkii | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CAN | NH | OR | WI | AK | MI | NH | WA | EE | SE.1 | SE.2 | RU | AZ | CO.1 | CO.2 | SD | F | N | - | |
| 1. | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | N | 0 | 16 | 0 |
| 2. | N | N | N | N | N | N | N | N | N | N | F | N | N | F | N | N | 2 | 14 | 0 |
| 4. | N | N | N | N | N | N | N | N | N | N | - | N | F | - | N | N | 1 | 13 | 2 |
| 7. | - | N | N | N | N | N | N | N | N | N | N | - | N | N | N | N | 0 | 14 | 2 |
| 8. | N | N | N | N | N | N | N | N | N | N | F | N | N | F | N | F | 3 | 13 | 0 |
| 9. | N | N | N | N | N | N | N | N | N | N | F | N | N | F | - | - | 2 | 12 | 2 |
| 17. | F | N | N | - | N | N | - | N | - | N | F | - | F | F | N | F | 5 | 7 | 4 |
| 19. | F | N | N | N | N | N | N | N | N | N | F | - | F | F | N | F | 5 | 1 0 | 1 |
| 21. | N | N | N | N | N | N | N | N | N | N | - | - | N | F | N | N | 1 | 13 | 2 |
| 22. | N | N | N | N | F | N | - | N | N | N | F | N | N | F | - | F | 4 | 10 | 2 |
| 23. | N | N | N | N | N | N | N | N | N | N | N | N | N | - | N | - | 0 | 14 | 2 |
| 24. | - | N | N | N | N | N | N | N | N | N | F | N | N | F | N | - | 2 | 12 | 2 |
| F | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 3 | 8 | 0 | 4 | 25 | ||
| N | 8 | 12 | 12 | 11 | 11 | 12 | 10 | 12 | 11 | 12 | 3 | 8 | 9 | 2 | 10 | 5 | 148 | ||
| - | 2 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 1 | 0 | 2 | 4 | 0 | 2 | 2 | 3 | 19 | ||
| F. mounceae | F. ochracea | F. pinicola | F. schrenkii | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CAN | NH | OR | WI | AK | MI | NH | WA | EE | SE.1 | SE.2 | RU | AZ | CO.1 | CO.2 | SD | F | N | - | |
| 1. | N,N (I,III) | N,N,N (I,III,IV) | N,N,N (I,III,IV) | N,N,N (I,III,IV) | N,N,N (I,III,IV) | N,N (I,III) | N,N,N (I,III,IV) | N,N,N (I,III,IV) | 0 | 22 | 0 | ||||||||
| 4. | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | -,N (I,II) | N,N (I,III) | F,N (I,II) | -,N,N (I,III,IV) | N,- (I,II) | N,N,N (I,III,IV) | 1 | 35 | 3 |
| 5. | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | 0 | 16 | 0 | ||||||||
| 7. | -,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | N,N (I,II) | -,N (I,III) | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | 0 | 37 | 2 |
| 10. | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,- (III,IV) | 0 | 13 | 1 | |||||||||
| 12. | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | 0 | 16 | 0 | ||||||||
| 13. | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,- (III,IV) | 0 | 13 | 1 | |||||||||
| 14. | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,- (III,IV) | N,N (III,IV) | N,- (III,IV) | N,N (III,IV) | 0 | 14 | 2 | ||||||||
| 15. | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | -,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | 0 | 13 | 1 | |||||||||
| 17. | F,N (I,II) | N,N (I,II) | N,N (I,II) | N,N (I,II) | N,N (I,II) | F,N (I,II) | N,N (I,II) | 2 | 12 | 0 | |||||||||
| 20. | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | N,N (III,IV) | 0 | 14 | 0 | |||||||||
| 21. | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N,N (I,III,IV) | N,N (I,II) | N,N,F (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | -,N (I,II) | -,N,N (I,III,IV) | N,N (I,II) | F,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | 2 | 36 | 2 |
| 23. | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N,N (I,III,IV) | N,N (I,II) | N,N,F (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | N,N (I,II) | N,N (I,III) | N,N (I,II) | -,-,N (I,III,IV) | N,N (I,II) | -,N,F (I,III,IV) | 2 | 34 | 3 |
| 24. | -,N (I,II) | N,N (I,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | N,N,N (I,III,IV) | N,N (I,II) | F,N (I,II) | N,N (I,III) | N,N (I,II) | F,N,N (I,III,IV) | N,N (I,II) | -,N,F (I,III,IV) | 3 | 33 | 2 |
| F | 1 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 0 | 1 | 2 | 0 | 2 | 10 | ||
| N | 9 | 30 | 12 | 32 | 30 | 12 | 29 | 12 | 31 | 12 | 8 | 19 | 9 | 26 | 11 | 26 | 308 | ||
| - | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 2 | 2 | 0 | 4 | 1 | 4 | 17 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferson-Mitchell, A.; Carta, L.; Haight, J.-E.; Newcombe, G. Nematodes Consume Four Species of a Common, Wood-Decay Fungus. Forests 2023, 14, 634. https://doi.org/10.3390/f14030634
Ferson-Mitchell A, Carta L, Haight J-E, Newcombe G. Nematodes Consume Four Species of a Common, Wood-Decay Fungus. Forests. 2023; 14(3):634. https://doi.org/10.3390/f14030634
Chicago/Turabian StyleFerson-Mitchell, Abigail, Lynn Carta, John-Erich Haight, and George Newcombe. 2023. "Nematodes Consume Four Species of a Common, Wood-Decay Fungus" Forests 14, no. 3: 634. https://doi.org/10.3390/f14030634