

Predicting Distribution and Range Dynamics of Three Threatened Cypripedium Species under Climate Change Scenario in Western Himalaya
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
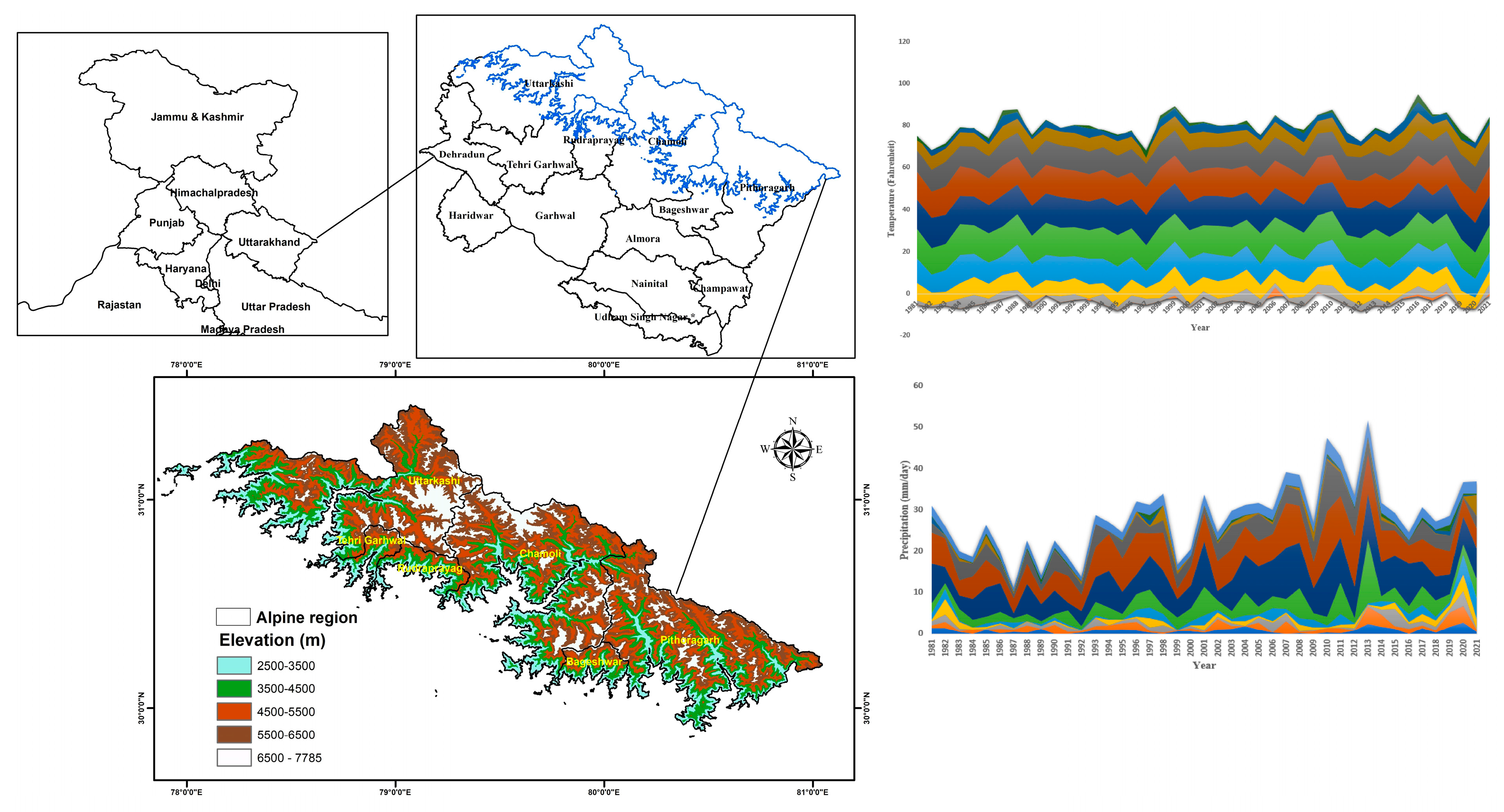
2.1. Study Area
2.2. Study Species
2.3. Data Collection
2.4. Predictor Variables
2.5. Predictive Modeling
3. Results
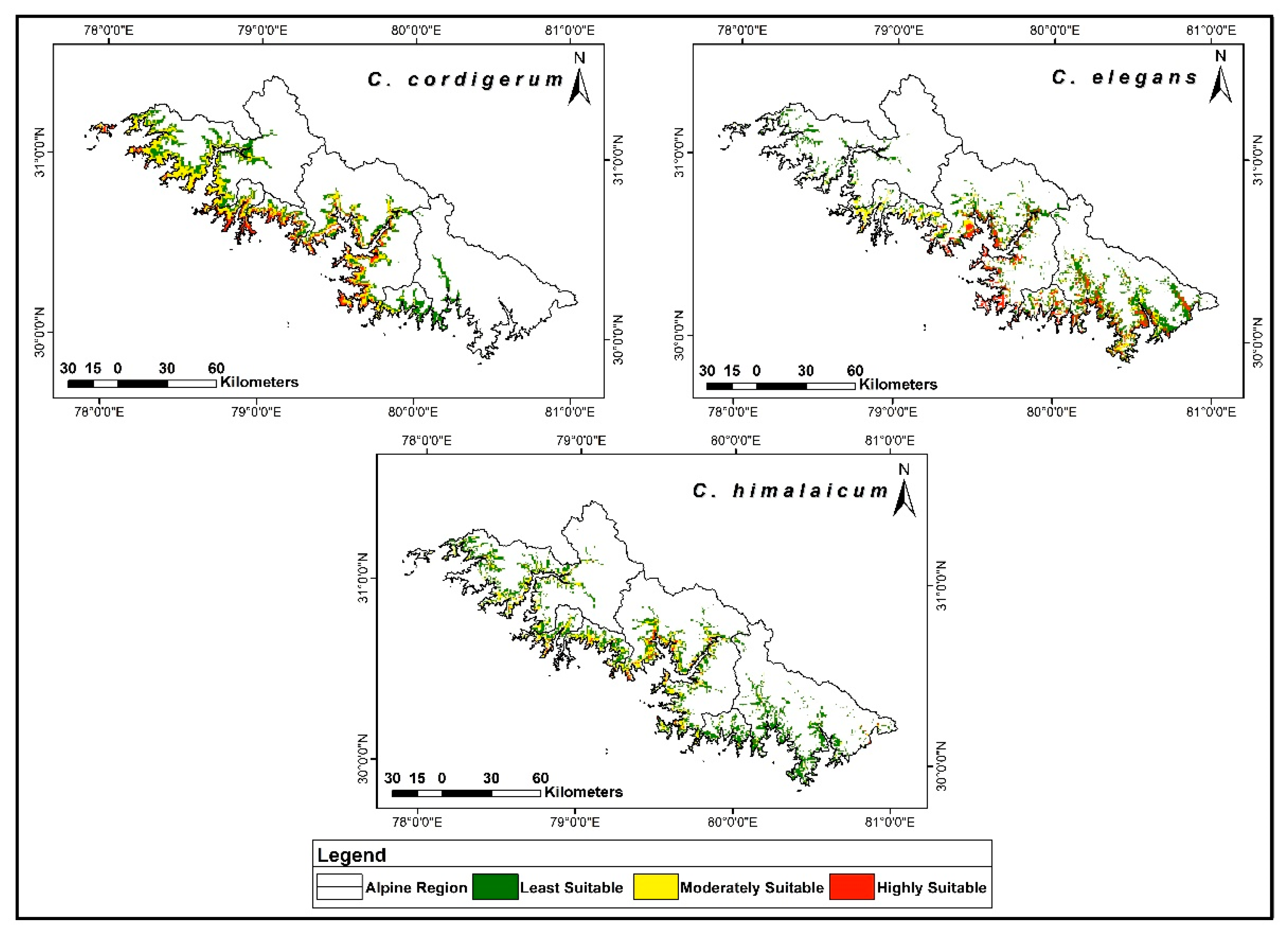
3.1. Habitat Preferences of the Species
3.2. Potential Distribution and Habitat Preferences
3.3. Habitat Suitability under Current Climatic Conditions
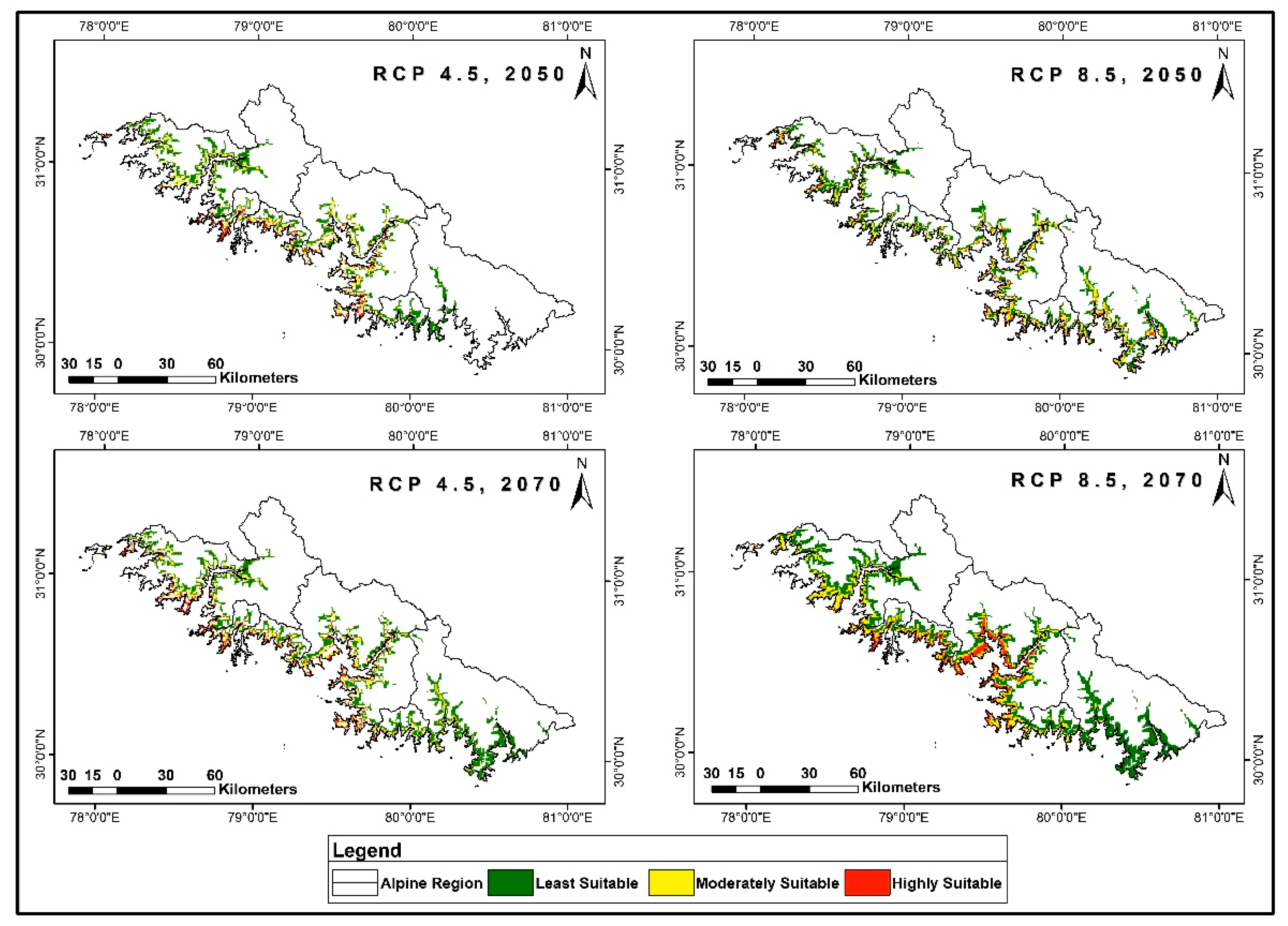
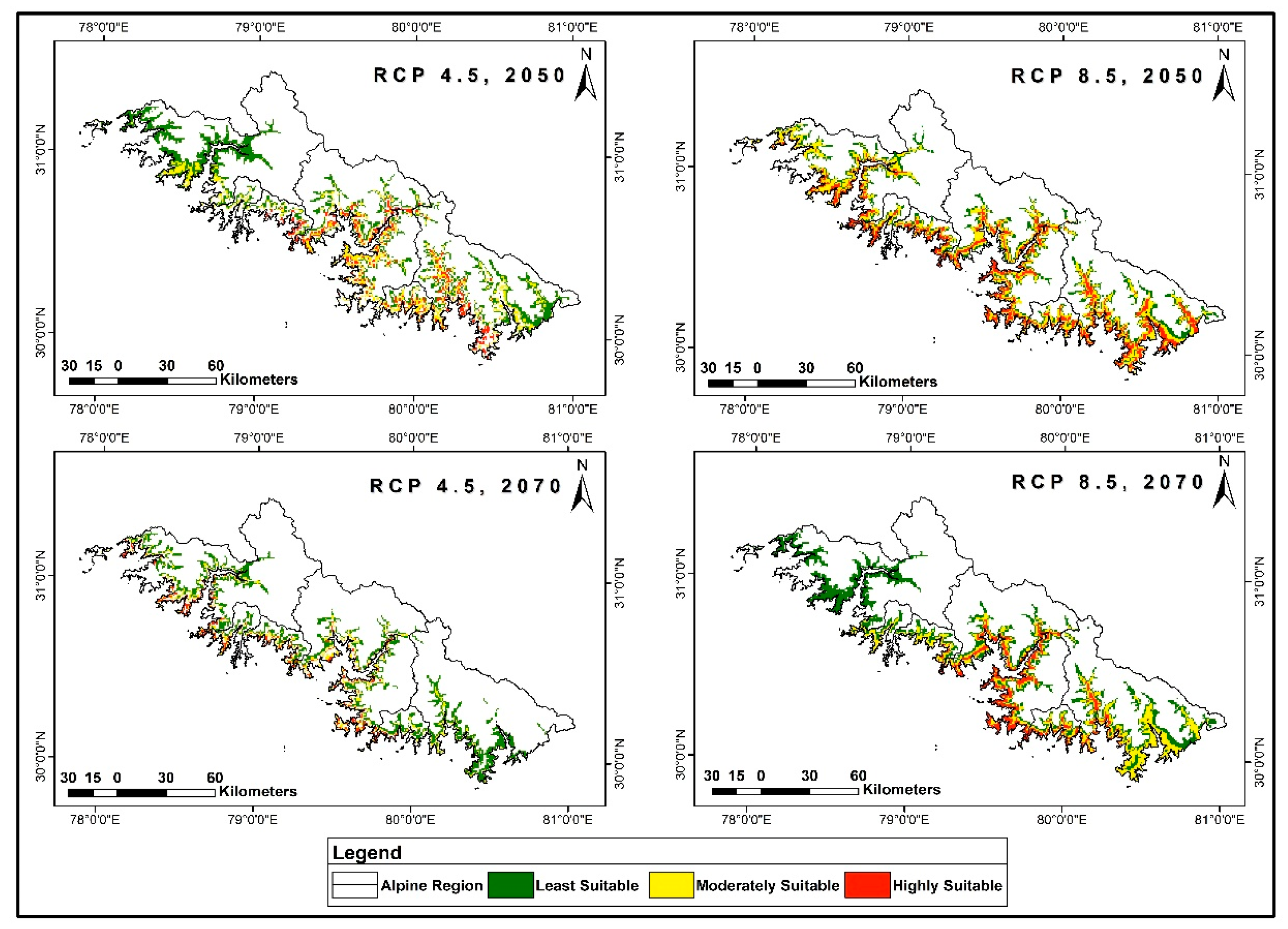
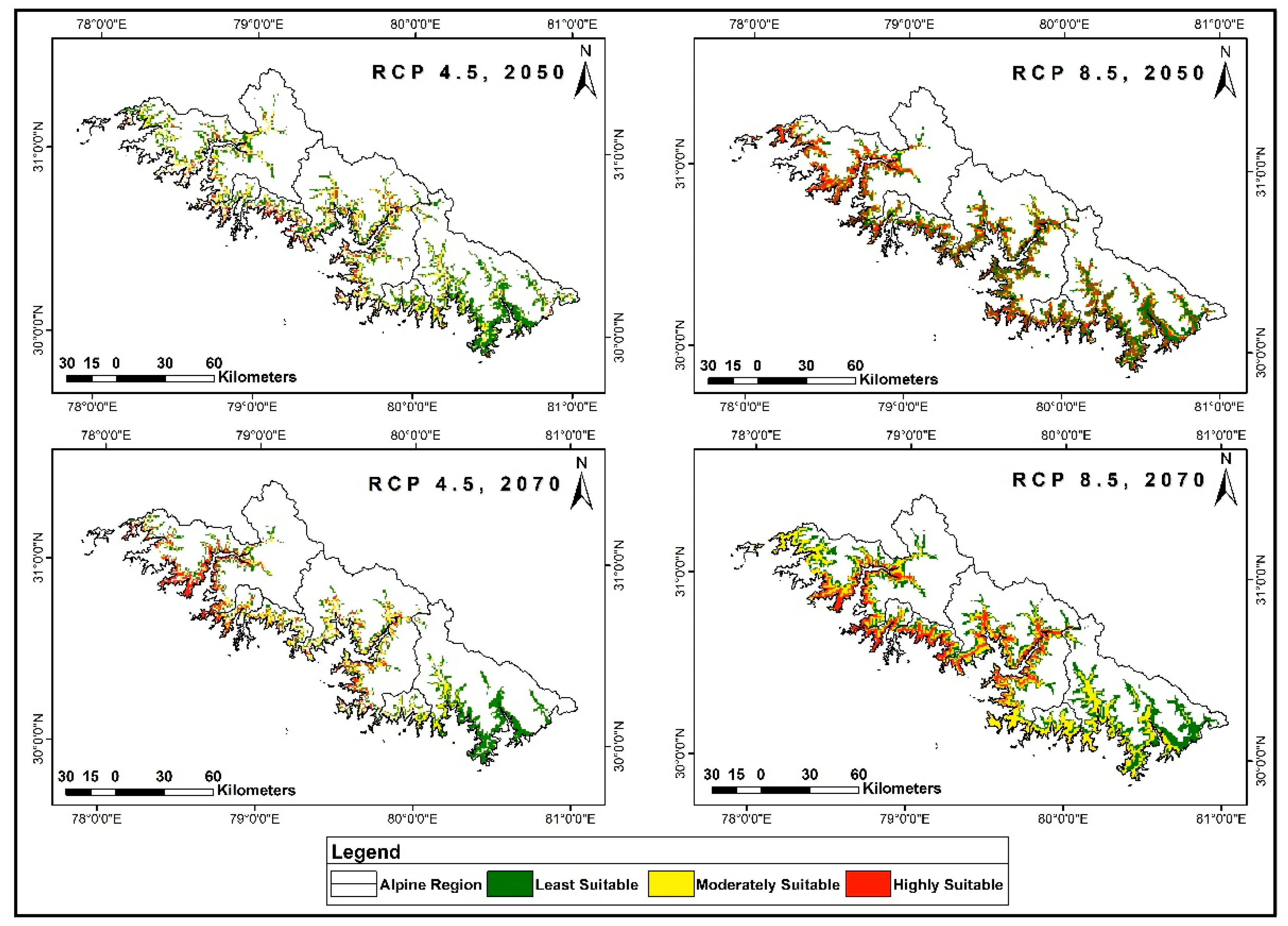
3.4. Distribution Prediction in Future Climate Scenarios
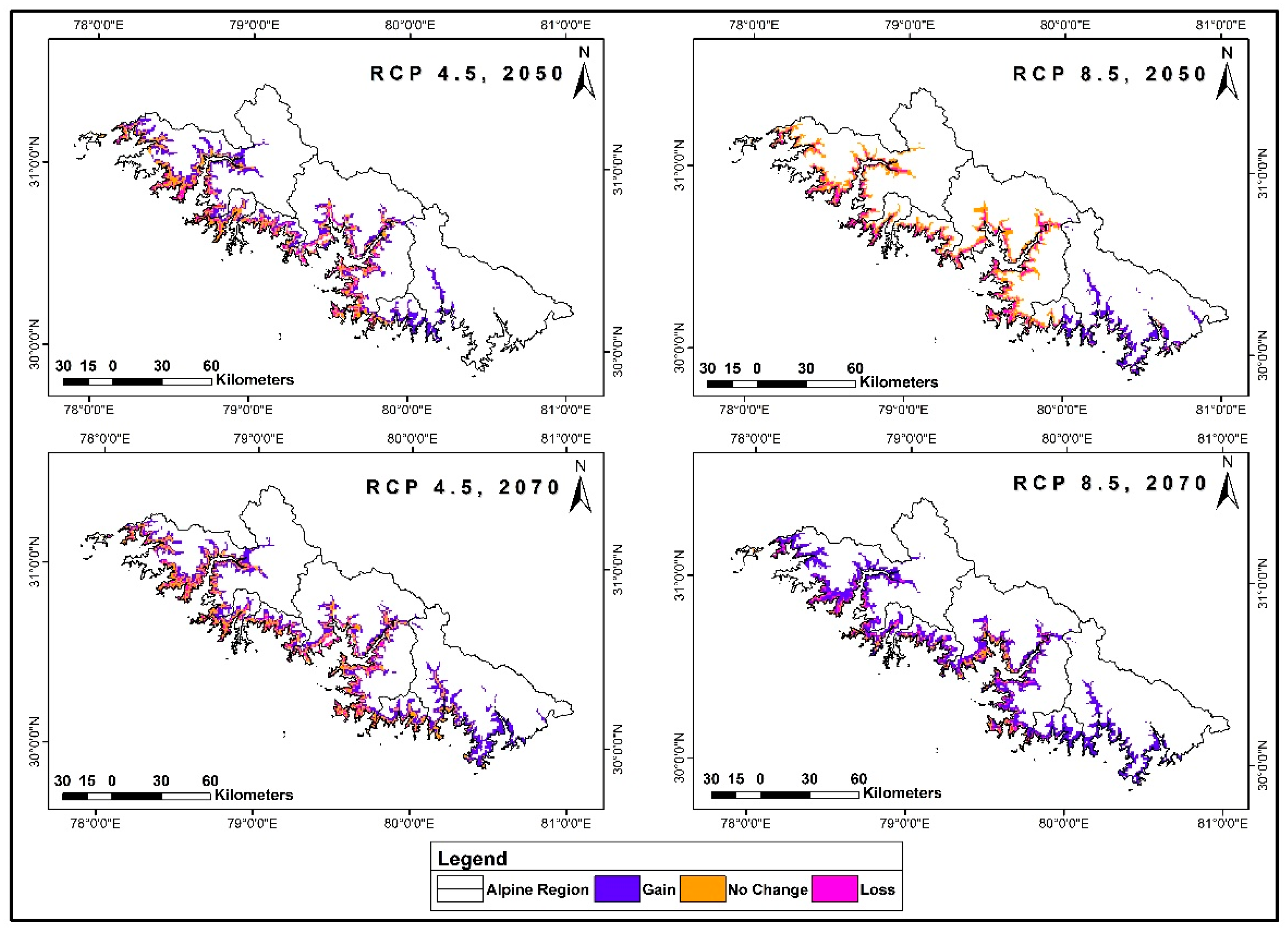
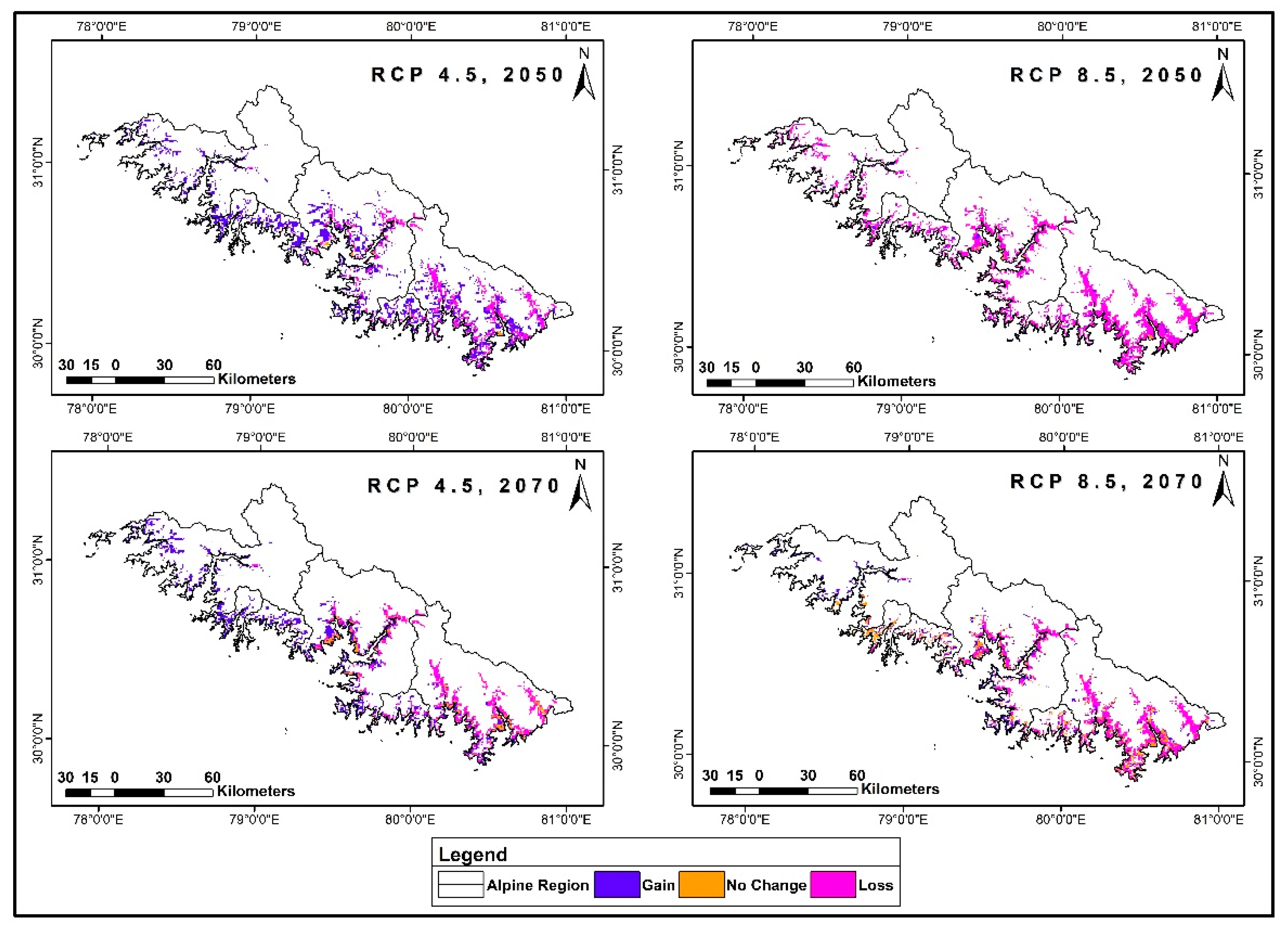
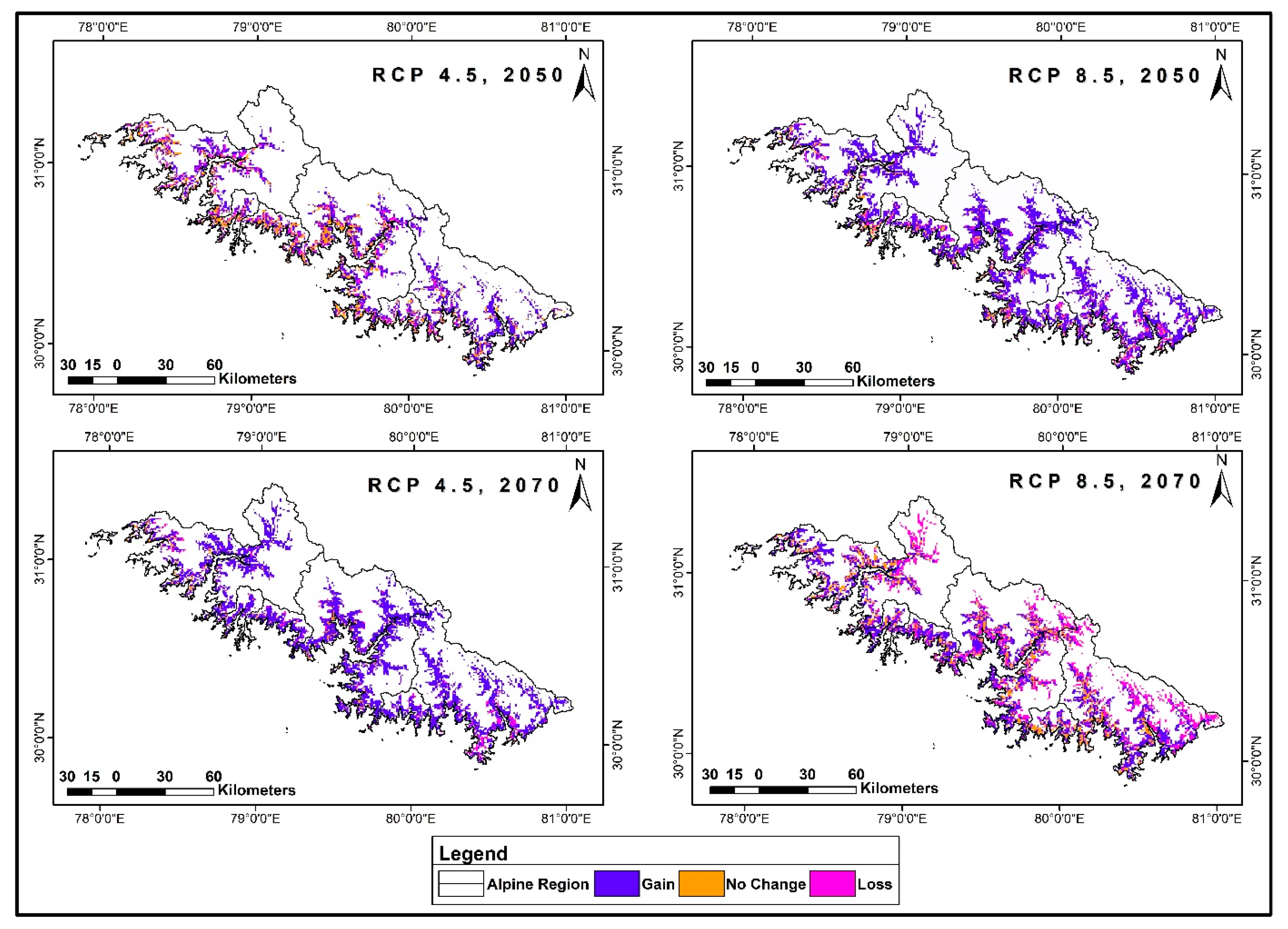
3.5. Range Dynamics under Future Climatic Scenarios
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N. Threatened biotas: “hot spots” in tropical forests. Environmentalist 1988, 8, 187–208. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.K.; Rana, H.K.; Ranjitkar, S.; Ghimire, S.K.; Gurmachhan, C.M.; O’Neill, A.R.; Sun, H. Climate-change threats to distribution, habitats, sustainability and conservation of highly traded medicinal and aromatic plants in Nepal. Ecol. Indic. 2020, 115, 106435. [Google Scholar] [CrossRef]
- Brandt, J.S.; Haynes, M.A.; Kuemmerle, T.; Waller, D.M.; Radeloff, V.C. Regime shift on the roof of the world: Alpine meadows converting to shrublands in the southern Himalayas. Biol. Conserv. 2013, 158, 116–127. [Google Scholar] [CrossRef]
- Singh, L.; Tariq, M.; Sekar, K.C.; Bhatt, I.D.; Nandi, S.K. Ecological niche modelling: An important tool for predicting suitable habitat and conservation of the Himalayan medicinal herbs. ENVIS Bull. Himal. Ecol. 2017, 25, 154–155. [Google Scholar]
- Thakur, S.; Negi, V.S.; Dhyani, R.; Satish, K.V.; Bhatt, I.D. Vulnerability assessments of mountain forest ecosystems: A global synthesis. Trees For. People 2021, 6, 100156. [Google Scholar] [CrossRef]
- Negi, V.S.; Pathak, R.; Rawal, R.S.; Bhatt, I.D.; Sharma, S. Long-term ecological monitoring on forest ecosystems in Indian Himalayan Region: Criteria and indicator approach. Ecol. Indic. 2019, 102, 374–381. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Cribb, P. The Genus Cypripedium; Timber Press Inc.: Portland, OR, USA, 1997. [Google Scholar]
- Jalal, J.S.; Jayanthi, J. An annonated checklist of the the orchid of western Himalaya, India. Lankesteriana 2015, 15, 7–50. [Google Scholar] [CrossRef] [Green Version]
- Jalal, J.S.; Kumar, P.; Rawat, G.S.; Pangtey, Y.P.S. Orchidaceae, Uttarakhand, Western Himalaya, India. Check List 2008, 4, 304–320. [Google Scholar] [CrossRef]
- Diez, J.M.; Pulliam, H.R. Hierarchical analysis of species distributions and abundance across environmental gradients. Ecol. Soc. Am. 2007, 88, 3144–3152. [Google Scholar] [CrossRef]
- Rai, I.D.; Adhikari, B.S.; Rawat, G.S. A unique patch of timberline ecotone with three species of Lady’s slipper orchids in Garhwal Himalaya, India. J. Threat Taxa 2010, 2, 766–769. [Google Scholar] [CrossRef] [Green Version]
- Kull, T. Cypripedium calceolus L. J. Ecol. 1999, 87, 913–924. [Google Scholar] [CrossRef]
- Warren, J.; McLaughlin, M.; Bardsley, J.; Eich, J.; Esche, C.A.; Kropkowski, L.; Risch, S. The Strengths and Challenges of Implementing EBP in Healthcare Systems. Worldviews Evid. Based Nurs. 2016, 13, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Polak, T.; Saltz, D. Reintroduction as an Ecosystem Restoration Technique. Conserv. Biol. 2011, 25, 424–425. [Google Scholar] [CrossRef] [PubMed]
- Suyal, R.; Bhatt, D.; Rawal, R.S.; Tewari, L.M. Status of two threatened astavarga herbs, Polygonatumcirrhifolium and Malaxismuscifera, in West Himalaya: Conservation implications. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 695–704. [Google Scholar] [CrossRef]
- Hooker, J.D. A Sketch of the Flora of British India; Eyre and Spottiswoode: London, UK, 1907; Volume 1, pp. 157–212. [Google Scholar]
- Bachman, S.; Moat, J.; Hill, A.W.; de la Torre, J.; Scott, B. Supporting Red List threat assessments with GeoCAT: Geospatial conservation assessment tool. ZooKeys 2011, 150, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Fick, S.E.; Hijmans, R.J. WorldClim2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modelling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberon, R.G.; Pearson, R.P.; Anderson, E.; Martinez-Meyer, M.; Araujo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Thuiller, W.; Araujo, M.B.; Lavorel, S. Do we need land-cover data to model species distributions in Europe? J. Biogeogr. 2004, 31, 353–361. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudık, M.; Ferrier, S.; Guisan, A.; Hijmans, R.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis‐Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Araujo, M.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Li, G.Q.; Du, S. Simulating the potential distribution of Elaeagnus angustifolia L. based on climatic constraints in China. Ecol. Eng. 2018, 113, 27–34. [Google Scholar] [CrossRef]
- Santiz, E.C.; Lorenzo, C.; Carrillo-Reyes, A.; Navarrete, D.A.; Islebe, G. Effect of climate change on the distribution of a critically threatened species. Therya 2016, 7, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; De Siqeira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Wei, Y.; Wang, X. Maxent modeling for predicting the potential distribution of Sanghuang, an important group of medicinal fungi in China. Fungal Ecol. 2015, 17, 140–145. [Google Scholar] [CrossRef]
- Li, R.; Xu, M.; Wong, M.H.G.; Qiu, S.; Sheng, Q.; Li, X.; Song, Z. Climate change-induced decline in bamboo habitats and species diversity: Implications for giant panda conservation. Divers. Distrib. 2015, 21, 379–391. [Google Scholar] [CrossRef]
- Lenoir, J.; Ge’gout, J.C.; Marquet, P.; De Ruffray, P.; Brisse, H.J.S. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from south western China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Casajus, N.; Lek, S.; Grenouillet, G. Uncertainty in ensemble forecasting of species distribution. Glob. Chang. Biol. 2010, 16, 1145–1157. [Google Scholar] [CrossRef]
- Chitale, V.S.; Behera, M.D.; Roy, P.S. Future of endemic flora of biodiversity hotspots in India. PLoS ONE 2014, 9, e115264. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.; Bhatt, I.D.; Negi, V.S.; Nandi, S.K.; Rawal, R.S.; Bisht, A.K. Population status, threats, and conservation options of the orchid Dactylorhiza hatagirea in Indian Western Himalaya. Reg. Environ. Chang. 2021, 21, 40. [Google Scholar] [CrossRef]
- Chen, I.C.; Hill, J.K.; Ohlemuller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Riofrı´o-Dillon, G.; De Ruffray, P.; Vidal, C.; Pierrat, J.C.; Gégout, J.C. Changes in plant community composition lag behind climate warming in lowland forests. Nature 2011, 479, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Kutiel, P.; Lavee, H. Effect of slope aspect on soil and vegetation properties along an aridity transect. Isr. J. Plant Sci. 1999, 47, 169. [Google Scholar] [CrossRef]
- Boisvert-Marsh, L.; Perie, C.; de Blois, S. Shifting with climate? Evidence for recent changes in tree species distribution at high latitudes. Ecosphere 2014, 5, 1–33. [Google Scholar] [CrossRef]
- Yu, F.; Wang, T.; Groen, T.A.; Skidmore, A.K.; Yang, X.; Ma, K.; Wu, Z. Climate and land use changes will degrade the distribution of Rhododendrons in China. Sci. Total Environ. 2019, 659, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.K.; Rawat, G.S. Database of Himalayan plants based on published floras during a century. Data 2017, 2, 36. [Google Scholar] [CrossRef] [Green Version]
- Sobrino, E.; Gonzalez, A.; Sanz-Elorza, M.; Dana, E.; Sanchez-Mata, D.; Gavilan, R. The expansion of thermophilic plants in Iberian peninsula as a sign of climate change. In Fingerprints of Climate Change. Adaptive Behaviour and Shifting Species Range; Walther, G.R., Burga, C.A., Edwards, P.J., Eds.; Kulwer Publishers: Dordrecht, The Netherlands, 2001; pp. 163–184. [Google Scholar]
- Hassan, T.; Hamid, M.; Wani, S.A.; Malik, A.H.; Waza, S.A.; Khuroo, A.A. Substantial shifts in flowering phenology of Sternbergiavernalis in the Himalaya: Supplementing decadal field records with historical and experimental evidences. Sci. Total Environ. 2021, 795, 148811. [Google Scholar] [CrossRef] [PubMed]
- Bhutiyani, M.R.; Kale, V.S.; Pawar, N.J. Climate change and the precipitation variations in the northwestern Himalaya: 1866–2006. Int. J. Climatol. 2010, 30, 535–548. [Google Scholar] [CrossRef]
- Livensperger, C.; Steltzer, H.; Darrouzet-Nardi, A.; Sullivan, P.F.; Wallenstein, M.; Weintraub, M.N. Earlier snowmelt and warming lead to earlier but not necessarily more plant growth. Ann. Bot. 2016, 8, plw021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manish, K.; Telwala, Y.; Nautiyal, D.C.; Pandit, M.K. Modelling the impacts of future climate change on plant communities in the Himalaya: A case study from Eastern Himalaya, India. Model Earth Syst. Environ. 2016, 2, 92. [Google Scholar] [CrossRef] [Green Version]
- Chuine, I. Why does phenology drive species distribution? Philos. Trans. R. Soc. B 2010, 365, 3149–3160. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.K.; Jenamani, R.K.; Kalsi, S.R.; Panda, S.K. Some evidences of climate change in twentieth-century India. Clim. Chang. 2007, 85, 299–321. [Google Scholar] [CrossRef]
- Keller, F.; Goyette, S.; Beniston, M. Sensitivity analysis of snow cover to climate change scenarios and their impact on plant habitats in alpine terrain. Clim. Chang. 2005, 72, 299–319. [Google Scholar] [CrossRef]
- Hossain, M.M. Therapeutic orchids: Traditional uses and recent advances—An overview. Fitoterapia 2011, 82, 102–140. [Google Scholar] [CrossRef]
- Deb, C.R.; Imchen, T. Orchids of horticultural importance from Nagaland, India. Pleione 2011, 5, 44–48. [Google Scholar]
- Suyal, R.; Joshi, P.; Bahukhandi, A.; Bhandari, S. Diversity and Distribution Pattern of Orchids Along an Altitudinal Gradient: Pindari Valley, West Himalaya. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2022, 92, 817–824. [Google Scholar] [CrossRef]
- Jalal, J.S.; Rawat, G.S.; Pankaj, K. Status, distribution and habitats of orchids in Uttarakhand. J. Orchid. Soc. India 2010, 24, 35–41. [Google Scholar]
- Suyal, R.; Rawal, R.S.; Jalal, J.S. Noteworthy additions to the orchids of Kumaun Himalaya, India. Indian For. 2018, 144, 778–780. [Google Scholar]
- Tsiftsis, S.; Djordjević, V. Modelling sexually deceptive orchid species distributions under future climates: The importance of plant–pollinator interactions. Sci. Rep. 2020, 10, 10623. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | C. himalaicum | C. cordigerum | C. elegans |
|---|---|---|---|
| Bio-2 | 44.9 | 2.5 | 1.4 |
| Aspect | 25.2 | 14.5 | 8.4 |
| Bio-12 | 11.4 | 3.4 | -- |
| Bio-11 | 9.3 | 16.6 | 6.2 |
| Altitude | 7.7 | 1.3 | 22.1 |
| Bio-3 | -- | 4.9 | 48.4 |
| Slope | 1.1 | 4.1 | -- |
| Vegetation type | -- | 2.4 | -- |
| Bio-4 | -- | 0.4 | 7.9 |
| Soil type | 0.5 | 30.1 | -- |
| Year 2050 | Year 2070 | |||||||
|---|---|---|---|---|---|---|---|---|
| RCP 2.6 | RCP 8.6 | RCP 2.6 | RCP 8.6 | |||||
| AUC value | 0.84 | 0.75 | 0.92 | 0.87 | ||||
| TSS value | 0.75 | 0.81 | 0.88 | 0.74 | ||||
| Percentage of contribution | Bio-1 | Bio-19 | Bio-12 | Bio-6 | Bio-19 | Bio-1 | Bio-13 | Bio-6 |
| Value (%) | 34 | 27 | 41 | 25 | 37 | 24 | 31 | 30 |
| Area in km2 (10th percentile training presence threshold rule) | 190 | 124 | 140 | 107 | ||||
| Percentage of Area | 2.6 | 1.5 | 1.7 | 1.3 | ||||
| Highest probability of species occurrence | 0.74 | 0.65 | 0.71 | 0.81 | ||||
| 2050 | Year 2070 | |||||||
|---|---|---|---|---|---|---|---|---|
| RCP 2.6 | RCP 8.6 | RCP 2.6 | RCP 8.6 | |||||
| AUC value | 0.71 | 0.71 | 0.81 | 0.91 | ||||
| TSS value | 0.75 | 0.81 | 0.79 | 0.69 | ||||
| Percentage of contribution | Altitude | Bio-19 | Bio-1 | Bio-12 | Bio-12 | Bio-6 | Bio-13 | Bio-19 |
| Value (%) | 40 | 21 | 35 | 27 | 23 | 31 | 34 | 29 |
| Area in km2 ((10th percentile training presence threshold rule) | 97 | 55 | 86 | 51 | ||||
| Percentage of Area | 1.2 | 0.7 | 1.09 | 0.6 | ||||
| Highest probability of species occurrence | 0.65 | 0.71 | 0.68 | 0.7 | ||||
| 2050 | Year 2070 | |||||||
|---|---|---|---|---|---|---|---|---|
| RCP 2.6 | RCP 8.6 | RCP 2.6 | RCP 8.6 | |||||
| AUC value | 0.78 | 0.80 | 0.89 | 0.70 | ||||
| TSS value | 0.80 | 0.82 | 0.87 | 0.90 | ||||
| Percentage of contribution | Bio-12 | Bio-19 | Bio-13 | Slope | Bio-19 | Bio-1 | Bio-12 | Bio-19 |
| value | 37 | 25 | 34 | 26 | 37 | 24 | 37 | 32 |
| Area in km2 ((10th percentile training presence threshold rule) | 102 | 189 | 199 | 154 | ||||
| Percentage of Area | 1.3 | 2.4 | 2.5 | 1.9 | ||||
| Highest probability of species occurrence | 0.61 | 0.62 | 0.65 | 0.59 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chandra, N.; Singh, G.; Rai, I.D.; Mishra, A.P.; Kazmi, M.Y.; Pandey, A.; Jalal, J.S.; Costache, R.; Almohamad, H.; Al-Mutiry, M.; et al. Predicting Distribution and Range Dynamics of Three Threatened Cypripedium Species under Climate Change Scenario in Western Himalaya. Forests 2023, 14, 633. https://doi.org/10.3390/f14030633
Chandra N, Singh G, Rai ID, Mishra AP, Kazmi MY, Pandey A, Jalal JS, Costache R, Almohamad H, Al-Mutiry M, et al. Predicting Distribution and Range Dynamics of Three Threatened Cypripedium Species under Climate Change Scenario in Western Himalaya. Forests. 2023; 14(3):633. https://doi.org/10.3390/f14030633
Chicago/Turabian StyleChandra, Naveen, Gajendra Singh, Ishwari Datt Rai, Arun Pratap Mishra, Mohd. Yahya Kazmi, Arvind Pandey, Jeewan Singh Jalal, Romulus Costache, Hussein Almohamad, Motrih Al-Mutiry, and et al. 2023. "Predicting Distribution and Range Dynamics of Three Threatened Cypripedium Species under Climate Change Scenario in Western Himalaya" Forests 14, no. 3: 633. https://doi.org/10.3390/f14030633