
Lower Sensitivity of Soil Carbon and Nitrogen to Regional Temperature Change in Karst Forests Than in Non-Karst Forests
 and
and
Abstract
:1. Introduction
2. Methods and Materials
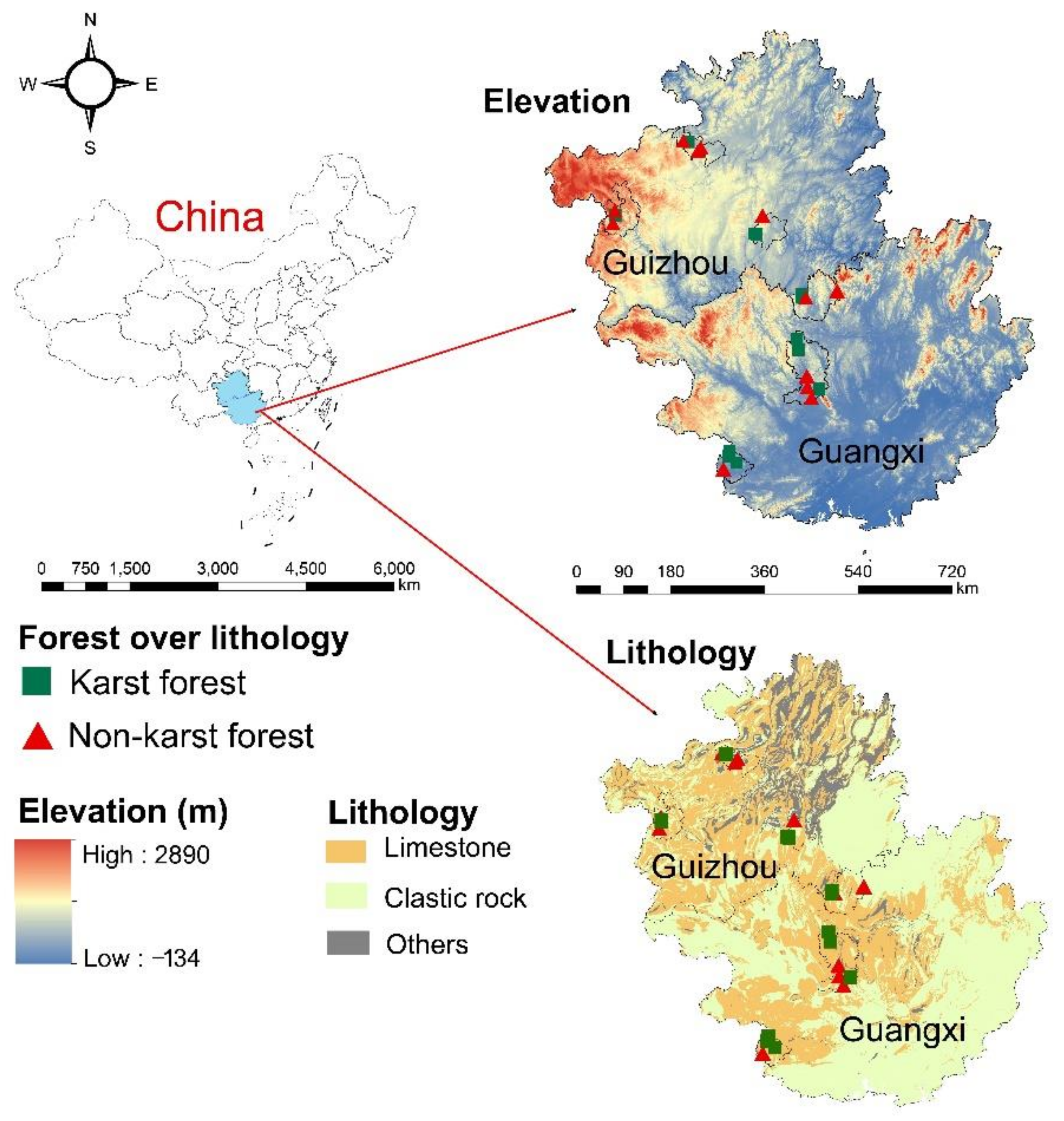
2.1. Study Area
2.2. Experimental Design and Sample Collection
2.3. Sample Processing and Analysis
2.4. Statistical Analysis
3. Results
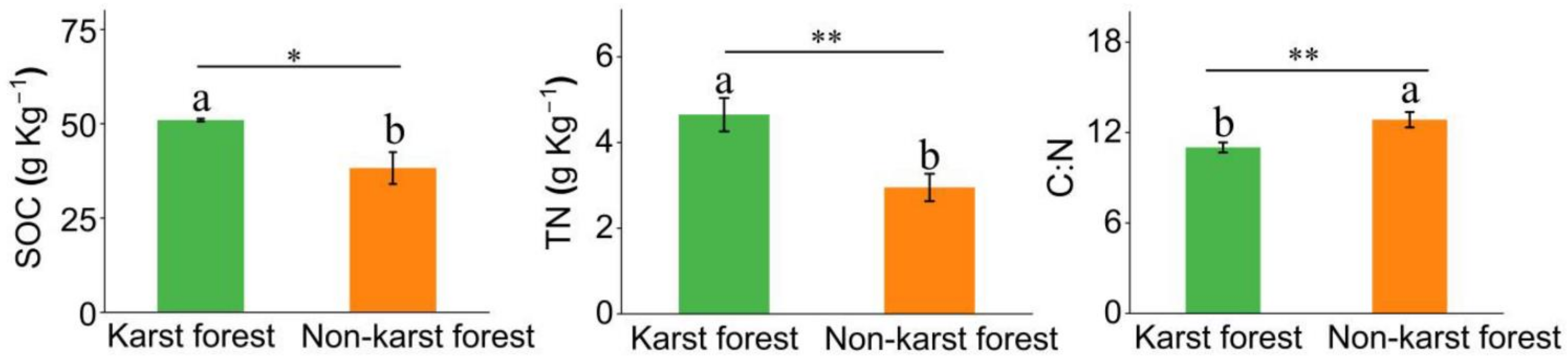
3.1. Differences in Soil Properties between Karst and Non-Karst Forests
3.2. Contents of SOC and TN in Karst and Non-Karst Forests
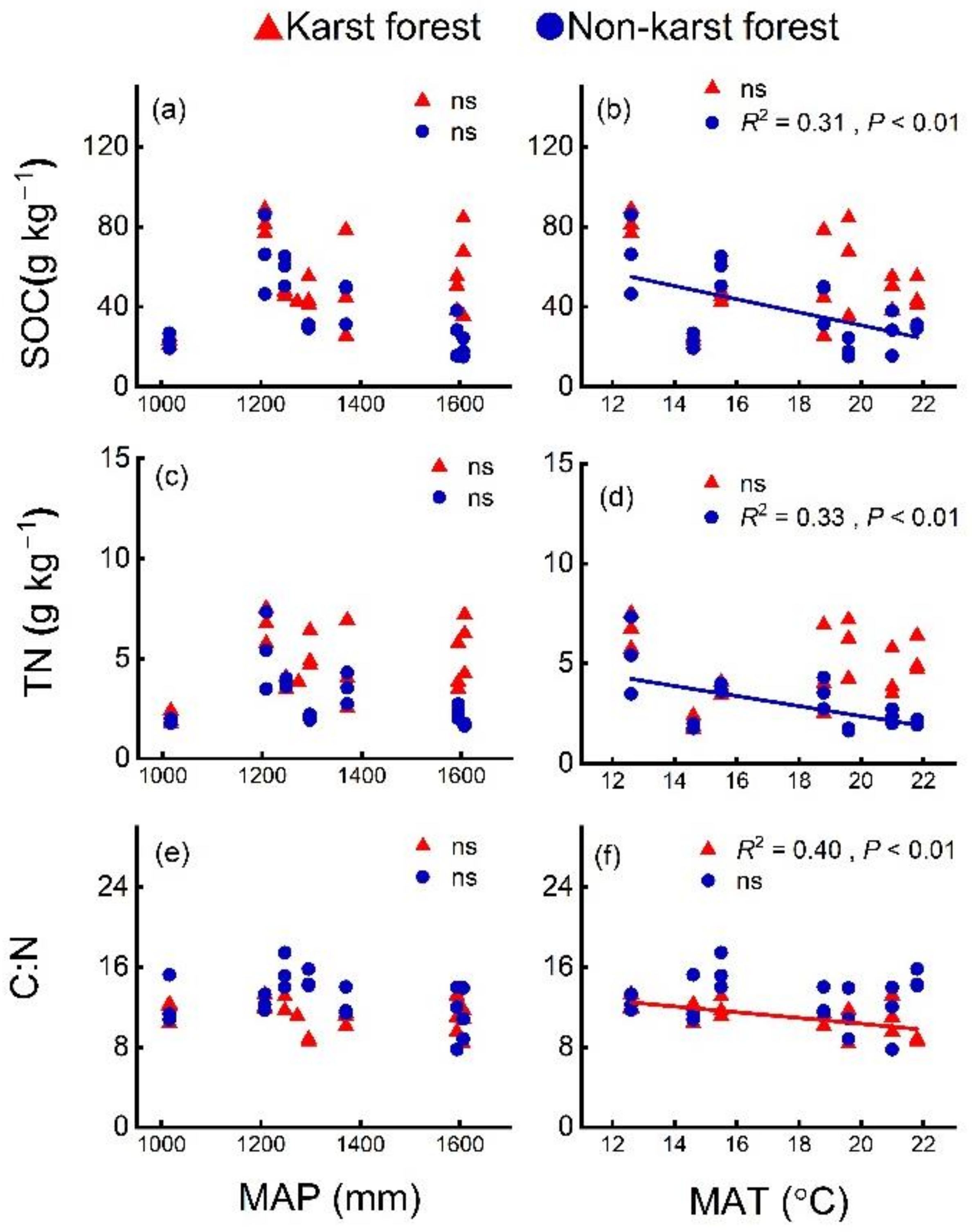
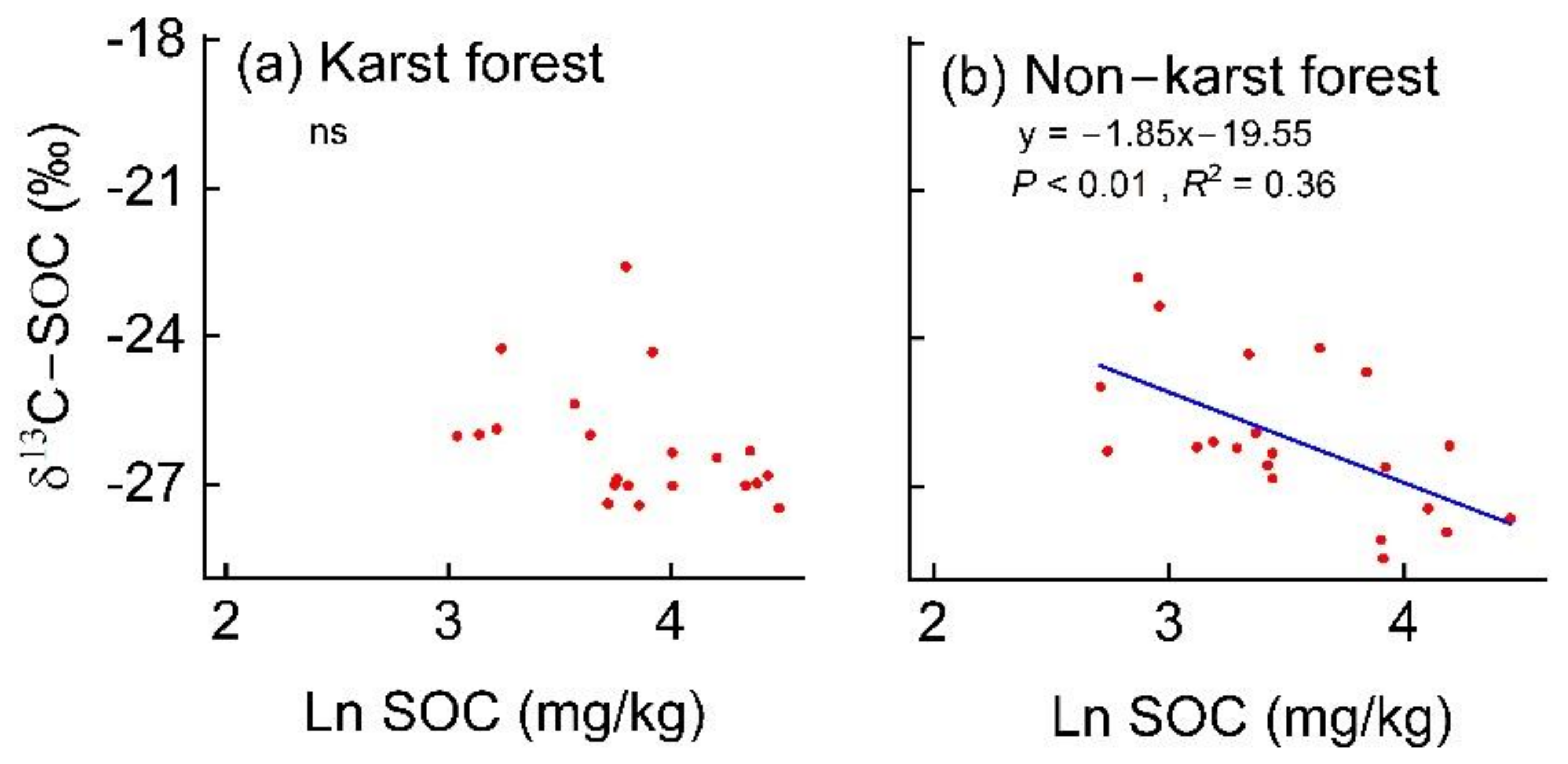
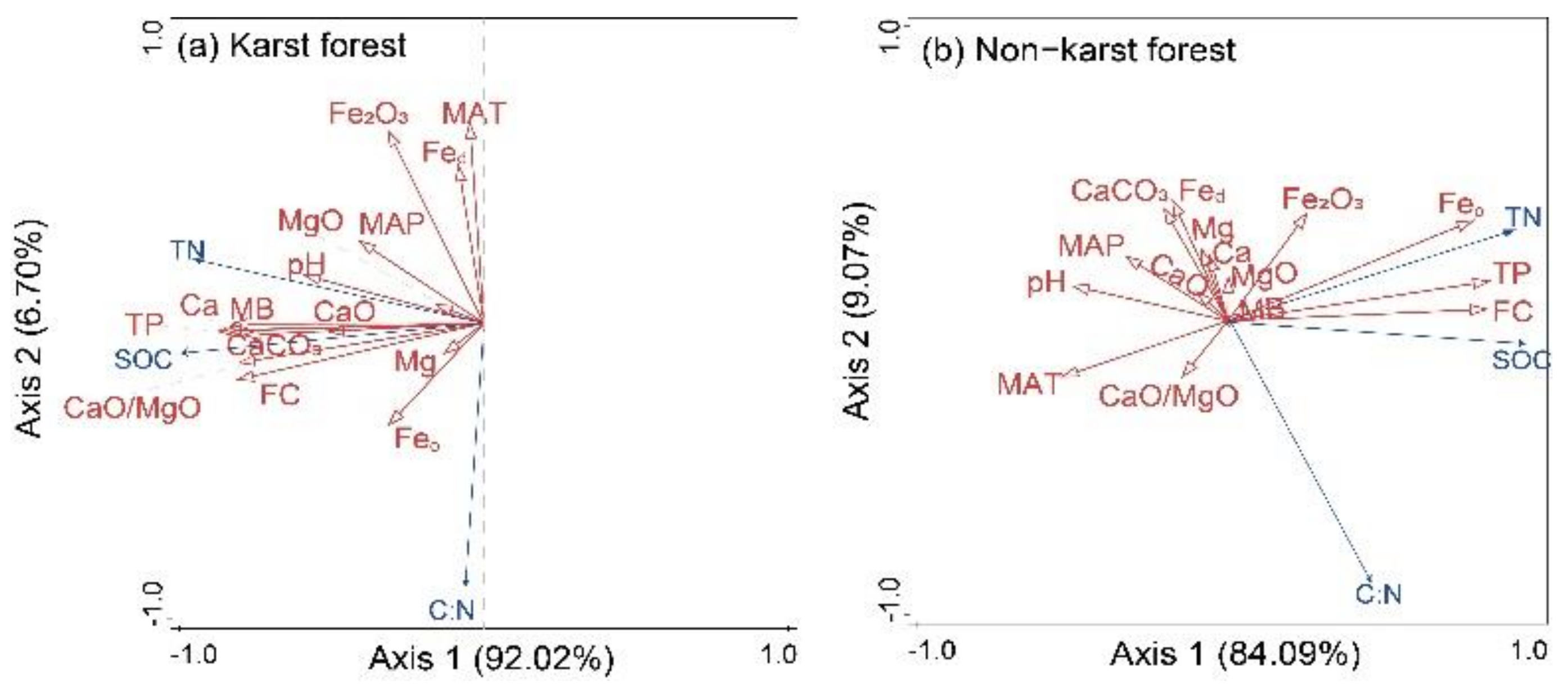
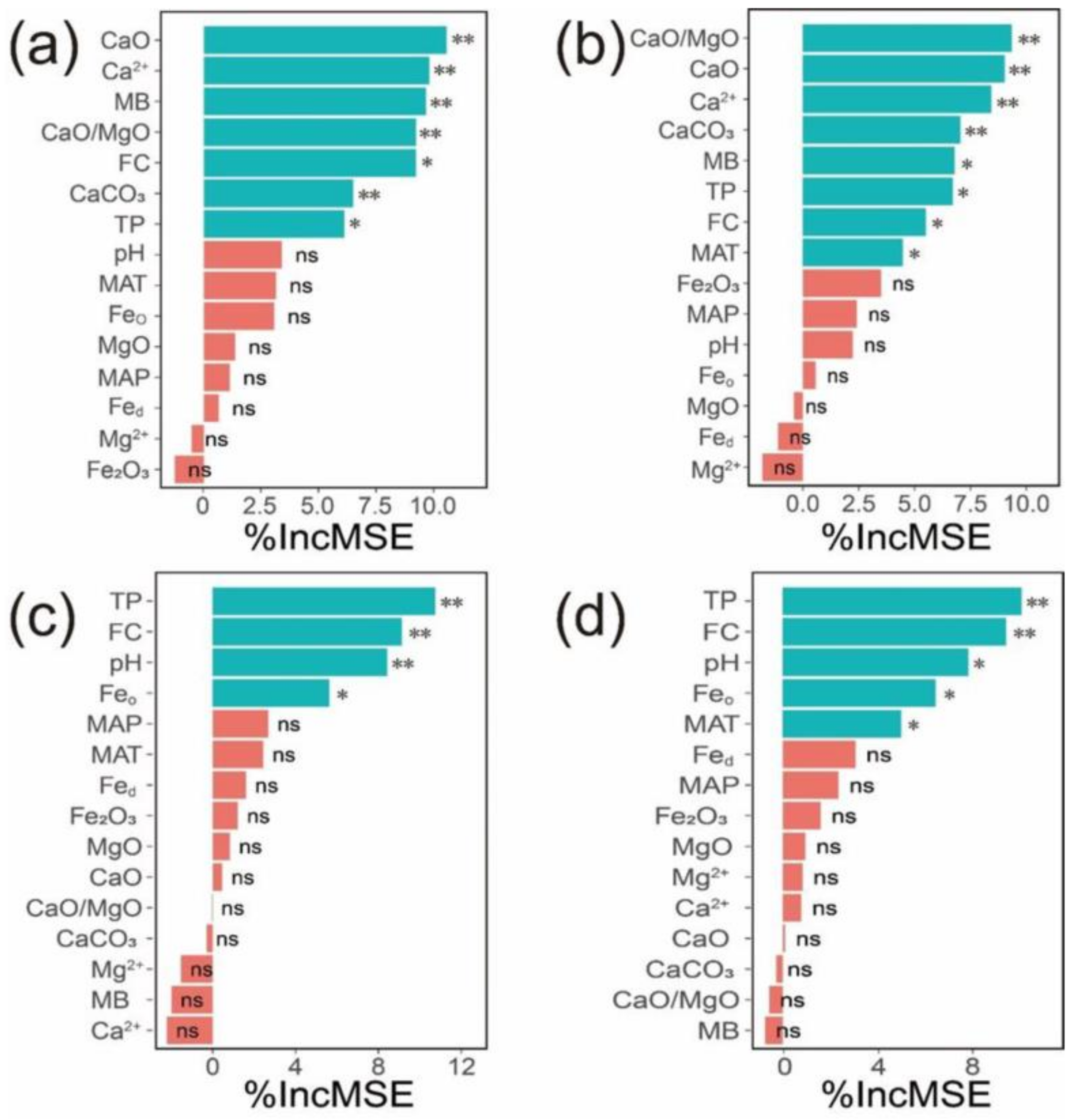
3.3. Soil Properties Correlated with SOC and TN Contents
4. Discussion
4.1. Differences in Soil Carbon and Nitrogen between Karst and Non-Karst Forests
4.2. Lower Temperature Sensitivity of SOC and TN in Karst Forests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Eswaran, H.; Van Den Berg, E.; Reich, P. Organic Carbon in Soils of the World. Soil Sci. Soc. Am. J. 1993, 57, 192–194. [Google Scholar] [CrossRef]
- Lal, R. Soil Carbon Sequestration Impacts on Global Climate Change and Food Security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature Sensitivity of Soil Carbon Decomposition and Feedbacks to Climate Change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef]
- Das, A.; Babu, S.; Datta, M.; Kumar, S.; Singh, R.; Avasthe, R.; Rathore, S.S.; Yadav, S.K.; Singh, V.K. Restoring Soil Carbon in Marginal Land of Indian Himalayas: Impact of Crop Intensification and Conservation Tillage. J. Environ. Manag. 2022, 318, 115603. [Google Scholar] [CrossRef]
- Ninan, K.N.; Inoue, M. Valuing Forest Ecosystem Services: What We Know and What We Don’t. Ecol. Econ. 2013, 93, 137–149. [Google Scholar] [CrossRef]
- Parhizkar, M.; Shabanpour, M.; Lucas-Borja, M.E.; Zema, D.A.; Li, S.; Tanaka, N.; Cerdà, A. Effects of Length and Application Rate of Rice Straw Mulch on Surface Runoff and Soil Loss under Laboratory Simulated Rainfall. Int. J. Sediment Res. 2021, 36, 468–478. [Google Scholar] [CrossRef]
- Bastin, J.-F.; Finegold, Y.; Garcia, C.; Mollicone, D.; Rezende, M.; Routh, D.; Zohner, C.M.; Crowther, T.W. The Global Tree Restoration Potential. Science 2019, 365, 76–79. [Google Scholar] [CrossRef]
- Chhabra, A.; Palria, S.; Dadhwal, V.K. Soil Organic Carbon Pool in Indian Forests. For. Ecol. Manag. 2003, 173, 187–199. [Google Scholar] [CrossRef]
- Galka, B.; Labaz, B.; Bogacz, A.; Bojko, O.; Kabala, C. Conversion of Norway Spruce Forests Will Reduce Organic Carbon Pools in the Mountain Soils of SW Poland. Geoderma 2014, 213, 287–295. [Google Scholar] [CrossRef]
- Fujii, K.; Sukartiningsih; Hayakawa, C.; Inagaki, Y.; Kosaki, T. Effects of Land Use Change on Turnover and Storage of Soil Organic Matter in a Tropical Forest. Plant Soil 2020, 446, 425–439. [Google Scholar] [CrossRef]
- Guan, J.-H.; Deng, L.; Zhang, J.-G.; He, Q.-Y.; Shi, W.-Y.; Li, G.; Du, S. Soil Organic Carbon Density and Its Driving Factors in Forest Ecosystems across a Northwestern Province in China. Geoderma 2019, 352, 1–12. [Google Scholar] [CrossRef]
- Albaladejo, J.; Ortiz, R.; Garcia-Franco, N.; Ruiz-Navarro, A.; Almagro, M.; García-Pintado, J.; Martínez-Mena, M. Land Use and Climate Change Impacts on Soil Organic Carbon Stocks in Semi-Arid Spain. J. Soils Sediments 2012, 13, 265–277. [Google Scholar] [CrossRef]
- Feng, W.; Liang, J.; Hale, L.E.; Jung, C.G.; Chen, J.; Zhou, J.; Xu, M.; Yuan, M.; Wu, L.; Bracho, R.; et al. Enhanced Decomposition of Stable Soil Organic Carbon and Microbial Catabolic Potentials by Long-Term Field Warming. Glob. Change Biol. 2017, 23, 4765–4776. [Google Scholar] [CrossRef]
- Hu, P.; Liu, S.-J.; Ye, Y.; Zhang, W.; Wang, K.-L.; Su, Y.-R. Effects of Environmental Factors on Soil Organic Carbon under Natural or Managed Vegetation Restoration. Land Degrad. Dev. 2018, 29, 387–397. [Google Scholar] [CrossRef]
- Li, D.; Niu, S.; Luo, Y. Global Patterns of the Dynamics of Soil Carbon and Nitrogen Stocks Following Afforestation: A Meta-Analysis. New Phytol. 2012, 195, 172–181. [Google Scholar] [CrossRef]
- Lützow, M.V.; Kögel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of Organic Matter in Temperate Soils: Mechanisms and Their Relevance under Different Soil Conditions-a Review: Mechanisms for Organic Matter Stabilization in Soils. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Luo, Y.; Su, B.; Currie, W.S.; Dukes, J.S.; Finzi, A.; Hartwig, U.; Hungate, B.; Mc MURTRIE, R.E.; Oren, R.; Parton, W.J.; et al. Progressive Nitrogen Limitation of Ecosystem Responses to Rising Atmospheric Carbon Dioxide. BioScience 2004, 54, 731. [Google Scholar] [CrossRef]
- Six, J.; Conant, R.T.; Paul, E.A.; Paustian, K. Stabilization Mechanisms of Soil Organic Matter: Implications for C-Saturation of Soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Deng, L.; Wang, K.-B.; Chen, M.-L.; Shangguan, Z.-P.; Sweeney, S. Soil Organic Carbon Storage Capacity Positively Related to Forest Succession on the Loess Plateau, China. CATENA 2013, 110, 1–7. [Google Scholar] [CrossRef]
- Callesen, I.; Liski, J.; Raulund-Rasmussen, K.; Olsson, M.T.; Tau-Strand, L.; Vesterdal, L.; Westman, C.J. Soil Carbon Stores in Nordic Well-Drained Forest Soils—Relationships with Climate and Texture Class. Glob. Change Biol. 2003, 9, 358–370. [Google Scholar] [CrossRef]
- Raich, J.W.; Russell, A.E.; Kitayama, K.; Parton, W.J.; Vitousek, P.M. TEMPERATURE INFLUENCES CARBON ACCUMULATION IN MOIST TROPICAL FORESTS. Ecology 2006, 87, 76–87. [Google Scholar] [CrossRef]
- Tashi, S.; Singh, B.; Keitel, C.; Adams, M. Soil Carbon and Nitrogen Stocks in Forests along an Altitudinal Gradient in the Eastern Himalayas and a Meta-Analysis of Global Data. Glob. Change Biol. 2016, 22, 2255–2268. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, E.; Matos, M.; Ríos, S.; Santiago, C.; Marín-Spiotta, E. Clay and Climate Are Poor Predictors of Regional-Scale Soil Carbon Storage in the US Caribbean. Geoderma 2019, 354, 113841. [Google Scholar] [CrossRef]
- Adopted, I. Climate Change 2014 Synthesis Report; IPCC: Geneva, Szwitzerland, 2014. [Google Scholar]
- Maaroufi, N.I.; De Long, J.R. Global Change Impacts on Forest Soils: Linkage Between Soil Biota and Carbon-Nitrogen-Phosphorus Stoichiometry. Front. For. Glob. Change 2020, 3, 16. [Google Scholar] [CrossRef]
- Menyailo, O.V. Forest Soil Carbon and Climate Changes. Forests 2022, 13, 398. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, C.; Chen, H.; Yue, Y.; Zhang, W.; Zhang, M.; Qi, X.; Fu, Z. Karst Landscapes of China: Patterns, Ecosystem Processes and Services. Landscape Ecol. 2019, 34, 2743–2763. [Google Scholar] [CrossRef]
- Jiang, Z.; Lian, Y.; Qin, X. Rocky Desertification in Southwest China: Impacts, Causes, and Restoration. Earth Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Wang, M.; Chen, H.; Zhang, W.; Wang, K. Soil Nutrients and Stoichiometric Ratios as Affected by Land Use and Lithology at County Scale in a Karst Area, Southwest China. Sci. Total Environ. 2018, 619, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Zhang, W.; Xiao, L.; Yang, R.; Xiao, D.; Zhao, J.; Wang, W.; Chen, H.; Wang, K. Moss-Dominated Biological Soil Crusts Modulate Soil Nitrogen Following Vegetation Restoration in a Subtropical Karst Region. Geoderma 2019, 352, 70–79. [Google Scholar] [CrossRef]
- Wen, L.; Li, D.; Yang, L.; Luo, P.; Chen, H.; Xiao, K.; Song, T.; Zhang, W.; He, X.; Chen, H.; et al. Rapid Recuperation of Soil Nitrogen Following Agricultural Abandonment in a Karst Area, Southwest China. Biogeochemistry 2016, 129, 341–354. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, W.; Wang, K.; Hou, Y. Soil Organic Carbon and Total Nitrogen as Affected by Land Use Types in Karst and Non-Karst Areas of Northwest Guangxi, China. J. Sci. Food Agric. 2012, 92, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, J.; Dürr, H.H.; Moosdorf, N.; Meybeck, M.; Kempe, S. The Geochemical Composition of the Terrestrial Surface (without Soils) and Comparison with the Upper Continental Crust. Int. J. Earth Sci. 2012, 101, 365–376. [Google Scholar] [CrossRef]
- D’Amico, M.; Gorra, R.; Freppaz, M. Small-Scale Variability of Soil Properties and Soil–Vegetation Relationships in Patterned Ground on Different Lithologies (NW Italian Alps). CATENA 2015, 135, 47–58. [Google Scholar] [CrossRef]
- Reith, F.; Brugger, J.; Zammit, C.M.; Gregg, A.L.; Goldfarb, K.C.; Andersen, G.L.; DeSantis, T.Z.; Piceno, Y.M.; Brodie, E.L.; Lu, Z.; et al. Influence of Geogenic Factors on Microbial Communities in Metallogenic Australian Soils. ISME J 2012, 6, 2107–2118. [Google Scholar] [CrossRef]
- Liu, M.; Xu, X.; Sun, A.Y.; Wang, K.; Liu, W.; Zhang, X. Is Southwestern China Experiencing More Frequent Precipitation Extremes? Environ. Res. Lett. 2014, 9, 064002. [Google Scholar] [CrossRef]
- Ahmed, Y.A.-R.; Pichler, V.; Homolák, M.; Gömöryová, E.; Nagy, D.; Pichlerová, M.; Gregor, J. High Organic Carbon Stock in a Karstic Soil of the Middle-European Forest Province Persists after Centuries-Long Agroforestry Management. Eur. J. For. Res. 2012, 131, 1669–1680. [Google Scholar] [CrossRef]
- Wen, L.; Li, D.; Chen, H.; Wang, K. Dynamics of Soil Organic Carbon in Density Fractions during Post-Agricultural Succession over Two Lithology Types, Southwest China. J. Environ. Manag. 2017, 201, 199–206. [Google Scholar] [CrossRef]
- Fu, H.; Jian, X.; Zhang, W.; Shang, F. A Comparative Study of Methods for Determining Carbonate Content in Marine and Terrestrial Sediments. Mar. Pet. Geol. 2020, 116, 104337. [Google Scholar] [CrossRef]
- Willers, C.; Jansen van Rensburg, P.J.; Claassens, S. Microbial Signature Lipid Biomarker Analysis–an Approach That Is Still Preferred, Even amid Various Method Modifications. J. Appl. Microbiol. 2015, 118, 1251–1263. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M. Impacts of Carbon and Flooding on Soil Microbial Communities: Phospholipid Fatty Acid Profiles and Substrate Utilization Patterns. Microb. Ecol. 1998, 35, 265–278. [Google Scholar] [CrossRef]
- Li, D.; Liu, J.; Chen, H.; Zheng, L.; Wang, K. Soil Microbial Community Responses to Forage Grass Cultivation in Degraded Karst Soils, Southwest China. Land Degrad. Dev. 2018, 29, 4262–4270. [Google Scholar] [CrossRef]
- Chung, H.; Zak, D.R.; Reich, P.B.; Ellsworth, D.S. Plant Species Richness, Elevated CO2, and Atmospheric Nitrogen Deposition Alter Soil Microbial Community Composition and Function. Glob. Chang. Biol. 2007, 13, 980–989. [Google Scholar] [CrossRef]
- Hoffmann, U.; Hoffmann, T.; Jurasinski, G.; Glatzel, S.; Kuhn, N.J. Assessing the Spatial Variability of Soil Organic Carbon Stocks in an Alpine Setting (Grindelwald, Swiss Alps). Geoderma 2014, 232, 270–283. [Google Scholar] [CrossRef]
- Viscarra Rossel, R.A.; Webster, R.; Bui, E.N.; Baldock, J.A. Baseline Map of Organic Carbon in Australian Soil to Support National Carbon Accounting and Monitoring under Climate Change. Glob. Change Biol. 2014, 20, 2953–2970. [Google Scholar] [CrossRef]
- Doetterl, S.; Stevens, A.; Six, J.; Merckx, R.; Oost, K.; Casanova, M.; Casanova-Katny, A.; Muñoz, C.; Boudin, M.; Zagal, E.; et al. Soil Carbon Storage Controlled by Interactions between Geochemistry and Climate. Nat. Geosci. 2015, 8, 780–783. [Google Scholar] [CrossRef]
- Kosmas, C.; Gerontidis, S.; Marathianou, M. The Effect of Land Use Change on Soils and Vegetation over Various Lithological Formations on Lesvos (Greece). CATENA 2000, 40, 51–68. [Google Scholar] [CrossRef]
- Oyonarte, C.; Aranda, V.; Durante, P. Soil Surface Properties in Mediterranean Mountain Ecosystems: Effects of Environmental Factors and Implications of Management. For. Ecol. Manag. 2008, 254, 156–165. [Google Scholar] [CrossRef]
- Li, D.; Wang, Z.; Sun, X.; Zhang, Q.; Wang, K. Tree Species Effects on Asymbiotic N2 Fixation in Subtropical Karst and Non-Karst Forests. Soil Biol. Biochem. 2018, 117, 185–190. [Google Scholar] [CrossRef]
- Kaiser, M.; Ellerbrock, R.H.; Wulf, M.; Dultz, S.; Hierath, C.; Sommer, M. The Influence of Mineral Characteristics on Organic Matter Content, Composition, and Stability of Topsoils under Long-Term Arable and Forest Land Use. J. Geophys. Res. Biogeosci. 2012, 117. [Google Scholar] [CrossRef]
- Chorover, J.; Amistadi, M.K. Reaction of Forest Floor Organic Matter at Goethite, Birnessite and Smectite Surfaces. Geochim. Cosmochim. Acta 2001, 65, 95–109. [Google Scholar] [CrossRef]
- Hu, P.; Zhang, W.; Xu, L.; Xiao, J.; Luo, Y.; Wang, K.-L. Lithologic Control of Microbial-Derived Carbon in Forest Soils. Soil Biol. Biochem. 2022, 167, 108600. [Google Scholar] [CrossRef]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The Importance of Anabolism in Microbial Control over Soil Carbon Storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef]
- Bailey, V.L.; Smith, J.L.; Bolton, H. Fungal-to-Bacterial Ratios in Soils Investigated for Enhanced C Sequestration. Soil Biol. Biochem. 2002, 34, 997–1007. [Google Scholar] [CrossRef]
- Xiao, D.; He, X.; Zhang, W.; Hu, P.; Sun, M.; Wang, K.-L. Comparison of Bacterial and Fungal Diversity and Network Connectivity in Karst and Non-Karst Forests in Southwest China. Sci. Total Environ. 2022, 822, 153179. [Google Scholar] [CrossRef]
- Pietri, J.C.A.; Brookes, P.C. Substrate Inputs and PH as Factors Controlling Microbial Biomass, Activity and Community Structure in an Arable Soil. Soil Biol. Biochem. 2009, 41, 1396–1405. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil Bacterial and Fungal Communities across a PH Gradient in an Arable Soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Golchin, A.; Clarke, P.; Oades, J.M. The Heterogeneous Nature of Microbial Products as Shown by Solid-State 13C CPIMAS NMR Spectroscopy. Biogeochemistry 1996, 34, 71–97. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.; Xiao, K.; Wang, K. Soil Microbial Processes and Resource Limitation in Karst and Non-Karst Forests. Funct. Ecol. 2018, 32, 1400–1409. [Google Scholar]
- LeBauer, D.S.; Treseder, K.K. NITROGEN LIMITATION OF NET PRIMARY PRODUCTIVITY IN TERRESTRIAL ECOSYSTEMS IS GLOBALLY DISTRIBUTED. Ecology 2008, 89, 371–379. [Google Scholar]
- Li, D.; Wen, L.; Yang, L.; Luo, P.; Xiao, K.; Chen, H.; Zhang, W.; He, X.; Chen, H.; Wang, K. Dynamics of Soil Organic Carbon and Nitrogen Following Agricultural Abandonment in a Karst Region. J. Geophys. Res. Biogeosci. 2017, 122, 230–242. [Google Scholar] [CrossRef]
- Hu, Y.; Xiang, D.; Veresoglou, S.D.; Chen, F.; Chen, Y.; Hao, Z.; Zhang, X.; Chen, B. Soil Organic Carbon and Soil Structure Are Driving Microbial Abundance and Community Composition across the Arid and Semi-Arid Grasslands in Northern China. Soil Biol. Biochem. 2014, 77, 51–57. [Google Scholar] [CrossRef]
- Chen, D.; Mi, J.; Chu, P.; Cheng, J.; Zhang, L.; Pan, Q.; Bai, Y. Patterns and Drivers of Soil Microbial Communities along a Precipitation Gradient on the Mongolian Plateau. Landsc. Ecol. 2014, 30, 1669–1682. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Ding, J.; Peng, Y.; Li, F.; Yang, G.-B.; Liu, L.; Qin, S.-Q.; Fang, K.; Yang, Y. Patterns and Drivers of Soil Microbial Communities in Tibetan Alpine and Global Terrestrial Ecosystems. J. Biogeogr. 2016, 43, 2027–2039. [Google Scholar] [CrossRef]
- Zhao, Z.; Wei, X.; Wang, X.; Ma, T.; Huang, L.; Gao, H.; Fan, J.; Li, X.; Jia, X. Concentration and Mineralization of Organic Carbon in Forest Soils along a Climatic Gradient. For. Ecol. Manag. 2019, 432, 246–255. [Google Scholar] [CrossRef]
- Gutiérrez-Girón, A.; Díaz-Pinés, E.; Rubio, A.; Gavilán, R.G. Both Altitude and Vegetation Affect Temperature Sensitivity of Soil Organic Matter Decomposition in Mediterranean High Mountain Soils. Geoderma 2015, 237, 1–8. [Google Scholar] [CrossRef]
- Eclesia, R.P.; Jobbagy, E.G.; Jackson, R.B.; Rizzotto, M.; Piñeiro, G. Stabilization of New Carbon Inputs Rather than Old Carbon Decomposition Determines Soil Organic Carbon Shifts Following Woody or Herbaceous Vegetation Transitions. Plant Soil 2016, 409, 99–116. [Google Scholar] [CrossRef]
- Rowley, M.C.; Grand, S.; Verrecchia, É.P. Calcium-Mediated Stabilisation of Soil Organic Carbon. Biogeochemistry 2018, 137, 27–49. [Google Scholar] [CrossRef]
- Zhang, W.; Parker, K.M.; Luo, Y.; Wan, S.; Wallace, L.L.; Hu, S. Soil Microbial Responses to Experimental Warming and Clipping in a Tallgrass Prairie. Glob. Change Biol. 2005, 11, 266–277. [Google Scholar] [CrossRef]
- Garten, C.T., Jr. Relationships among Forest Soil C Isotopic Composition, Partitioning, and Turnover Times. Can. J. For. Res. 2006, 36, 2157–2167. [Google Scholar] [CrossRef]
- Guillaume, T.; Damris, M.; Kuzyakov, Y. Losses of Soil Carbon by Converting Tropical Forest to Plantations: Erosion and Decomposition Estimated by Δ13C. Glob. Change Biol. 2015, 21, 3548–3560. [Google Scholar] [CrossRef]
- Powers, J.S.; Schlesinger, W.H. Geographic and Vertical Patterns of Stable Carbon Isotopes in Tropical Rain Forest Soils of Costa Rica. Geoderma 2002, 109, 141–160. [Google Scholar] [CrossRef]
- Holloway, J.M.; Dahlgren, R.A. Nitrogen in Rock: Occurrences and Biogeochemical Implications. Glob. Biogeochem. Cycles 2002, 16, 65-1–65-17. [Google Scholar] [CrossRef]
- Houlton, B.Z.; Morford, S.L.; Dahlgren, R.A. Convergent Evidence for Widespread Rock Nitrogen Sources in Earth’s Surface Environment. Science 2018, 360, 58–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Property | Karst Forest | Non-Karst Forest |
|---|---|---|
| Field capacity/% | 48.27 (2.46) | 49.71 (3.44) |
| Total porosity/% | 62.67 (1.46) | 61.98 (2.27) |
| pH | 5.89 (a) | 4.22 (b) |
| Ca/nmol·kg−1 | 35.65 (3.05) a | 6.75 (2.18) b |
| Mg/nmol·kg−1 | 9.39 (1.78) a | 2.20 (0.41) b |
| Fed/nmol·kg−1 | 3.11 (0.33) | 6.82 (1.12) |
| Feo/nmol·kg−1 | 55.42 (4.71) | 45.82 (4.05) b |
| CaCO3/% | 2.00 (0.21) a | 0.92 (0.15) b |
| CaO/% | 1.87 (0.51) a | 0.36 (0.08) b |
| MgO/% | 1.74 (0.23) a | 0.80 (0.12) b |
| Fe2O3/% | 9.43 (0.61) a | 7.10 (0.82) b |
| CaO/MgO | 1.11 (0.17) a | 0.52 (0.12) b |
| MB/nmol·kg−1 | 95.96 (8.91) a | 38.40 (4.02) b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yang, R.; Hu, P.; Xiao, D.; Wang, Z.; Zhang, W.; Wang, K. Lower Sensitivity of Soil Carbon and Nitrogen to Regional Temperature Change in Karst Forests Than in Non-Karst Forests. Forests 2023, 14, 355. https://doi.org/10.3390/f14020355
Li Y, Yang R, Hu P, Xiao D, Wang Z, Zhang W, Wang K. Lower Sensitivity of Soil Carbon and Nitrogen to Regional Temperature Change in Karst Forests Than in Non-Karst Forests. Forests. 2023; 14(2):355. https://doi.org/10.3390/f14020355
Chicago/Turabian StyleLi, Yunfan, Rong Yang, Peilei Hu, Dan Xiao, Zhongcheng Wang, Wei Zhang, and Kelin Wang. 2023. "Lower Sensitivity of Soil Carbon and Nitrogen to Regional Temperature Change in Karst Forests Than in Non-Karst Forests" Forests 14, no. 2: 355. https://doi.org/10.3390/f14020355