
Distinct Responses of European Beech (Fagus sylvatica L.) to Drought Intensity and Length—A Review of the Impacts of the 2003 and 2018–2019 Drought Events in Central Europe

Abstract
:1. Introduction
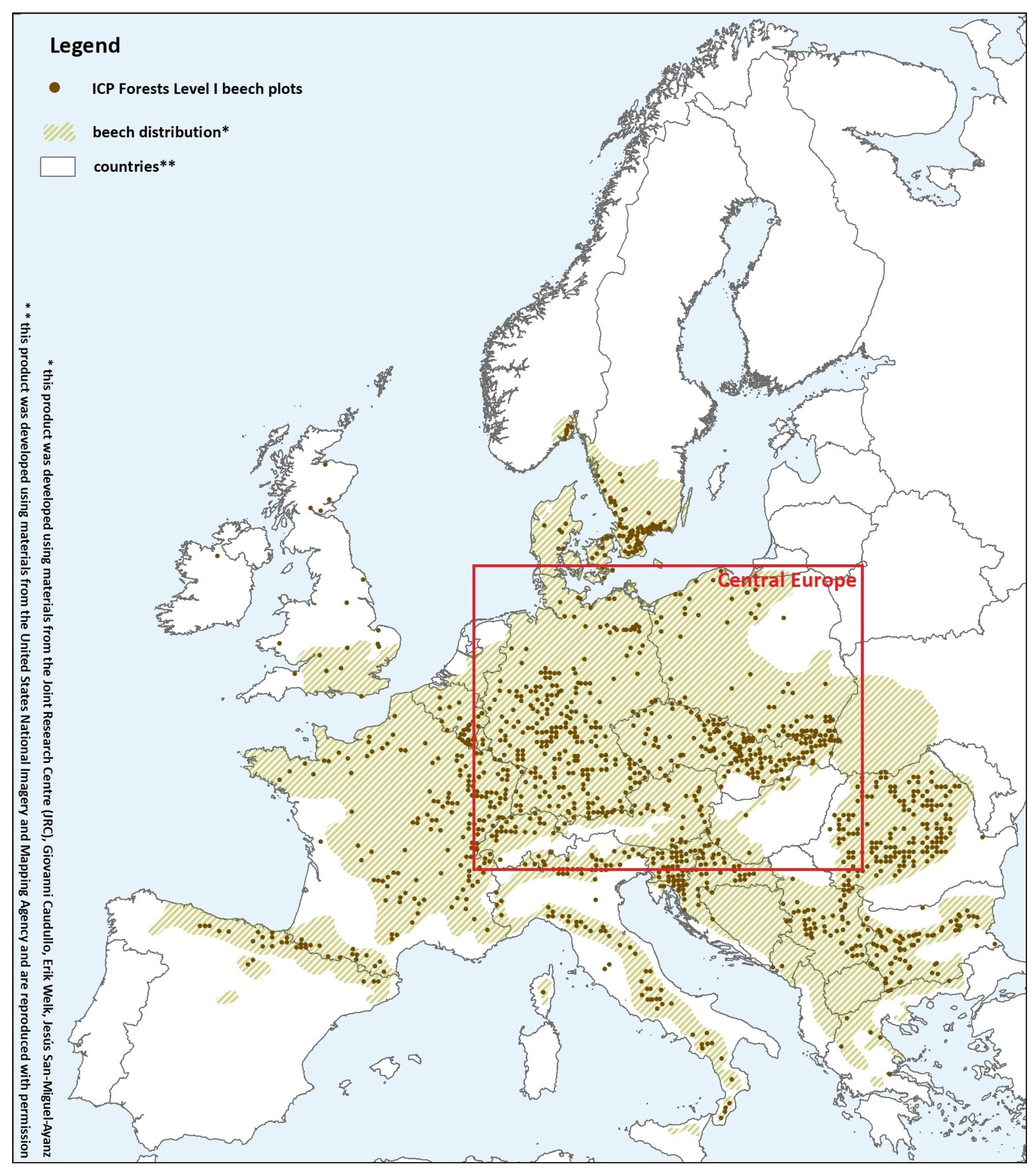
2. Climatic and Edaphic Conditions in Beech Dominated Areas
3. Single Year vs. Consecutive Drought Periods
4. Stress Response under Extreme Events
4.1. Negative Impacts of the 2003 Drought
4.2. Negative Impact of the Consecutive Droughts in 2018 and 2019
5. Unravelling Legacy Effects of Defoliation in the Wake of the 2018 Drought
5.1. Linking Defoliation and Growth
5.2. Beech Rooting Depth, Allometry, and Legacy Effects
6. Future Management Decisions for Beech
6.1. Site Conditions and Drought Response of Beech
6.2. Mixed versus Pure Stands of Beech
6.3. Stand Thinning to Mitigate Negative Legacy Effects in Beech
7. Outlook and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to Quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Ann. Rev. Ecol. Evol. Sys. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- Meyer, B.F.; Buras, A.; Rammig, A.; Zang, C.S. Higher Susceptibility of Beech to Drought in Comparison to Oak. Dendrochronologia 2020, 64, 125780. [Google Scholar] [CrossRef]
- Muffler, L.; Weigel, R.; Hacket-Pain, A.J.; Klisz, M.; van der Maaten, E.; Wilmking, M.; Kreyling, J.; van der Maaten-Theunissen, M. Lowest Drought Sensitivity and Decreasing Growth Synchrony towards the Dry Distribution Margin of European Beech. J. Biogeogr. 2020, 47, 1910–1921. [Google Scholar] [CrossRef]
- Scharnweber, T.; Manthey, M.; Criegee, C.; Bauwe, A.; Schröder, C.; Wilmking, M. Drought Matters—Declining Precipitation Influences Growth of Fagus sylvatica L. and Quercus robur L. in North-Eastern Germany. For. Ecol. Manag. 2011, 262, 947–961. [Google Scholar] [CrossRef]
- Bundesministerium für Ernährung und Landwirtschaft—BMEL. Ergebnisse der Waldzustandserhebung 2021; Referat 515—Nachhaltige Waldbewirtschaftung; BMEL: Bonn, Germany, 2022; pp. 1–76. [Google Scholar]
- West, E.; Morley, P.J.; Jump, A.S.; Donoghue, D.N.M. Satellite data track spatial and temporal declines in European beech forest canopy characteristics associated with intense drought events in the Rhön Biosphere Reserve, central Germany. Plant Biol. 2022, 24, 1120–1131. [Google Scholar] [CrossRef]
- Burton, P.J.; Jentsch, A.; Walker, L.R. The Ecology of Disturbance Interactions. Bioscience 2020, 70, 854–870. [Google Scholar] [CrossRef]
- Kannenberg, S.A.; Schwalm, C.R.; Anderegg, W.R.L. Ghosts of the Past: How Drought Legacy Effects Shape Forest Functioning and Carbon Cycling. Ecol. Lett. 2020, 23, 891–901. [Google Scholar] [CrossRef] [Green Version]
- Arend, M.; Link, R.M.; Zahnd, C.; Hoch, G.; Schuldt, B.; Kahmen, A. Lack of Hydraulic Recovery as a Cause of Post-Drought Foliage Reduction and Canopy Decline in European Beech. New Phytol. 2022, 234, 1195–1205. [Google Scholar] [CrossRef]
- Wang, W.; Peng, C.; Kneeshaw, D.D.; Larocque, G.R.; Luo, Z. Drought-Induced Tree Mortality: Ecological Consequences, Causes, and Modeling. Environ. Rev. 2012, 20, 109–121. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of Plant Survival and Mortality during Drought: Why Do Some Plants Survive While Others Succumb to Drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Giesecke, T.; Hickler, T.; Kunkel, T.; Sykes, M.T.; Bradshaw, R.H.W. Towards an Understanding of the Holocene Distribution of Fagus sylvatica L. J. Biogeogr. 2007, 34, 118–131. [Google Scholar] [CrossRef]
- Geßler, A.; Keitel, C.; Kreuzwieser, J.; Matyssek, R.; Seiler, W.; Rennenberg, H. Potential Risks for European Beech (Fagus sylvatica L.) in a Changing Climate. Trees 2007, 21, 1–11. [Google Scholar] [CrossRef]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Forests: Vegetation Ecology of Central Europe; Revised and Extended Version of the 6th German Edition; Springer International: Cham, Switzerland, 2017; pp. 1–972. [Google Scholar]
- Buras, A.; Rammig, A.; Zang, C.S. Quantifying Impacts of the 2018 Drought on European Ecosystems in Comparison to 2003. Biogeosciences 2020, 17, 1655–1672. [Google Scholar] [CrossRef] [Green Version]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.E.; Hauck, M.; Hajek, P.; et al. A First Assessment of the Impact of the Extreme 2018 Summer Drought on Central European Forests. Basic Appl. Ecol. 2020, 45, 86–103. [Google Scholar] [CrossRef]
- Bigler, C.; Vitasse, Y. Premature leaf discoloration of European deciduous trees is caused by drought and heat in late spring and cold spells in early fall. Agric. For. Meteorol. 2020, 307, 108492. [Google Scholar] [CrossRef]
- Rohner, B.; Kumar, S.; Liechti, K.; Gessler, A.; Ferretti, M. Tree Vitality Indicators Revealed a Rapid Response of Beech Forests to the 2018 Drought. Ecol. Indic. 2021, 120, 106903. [Google Scholar] [CrossRef]
- Scharnweber, T.; Smiljanic, M.; Cruz-García, R.; Manthey, M.; Wilmking, M. Tree Growth at the End of the 21st Century—The Extreme Years 2018/19 as Template for Future Growth Conditions. Environ. Res. Lett. 2020, 15, 074022. [Google Scholar] [CrossRef] [Green Version]
- Michel, A.; Prescher, A.K.; Schwärzel, K. (Eds.) Forest Condition in Europe: The 2020 Assessment; ICP Forests Technical Report under the UNECE Convention on Long-Range Transboundary Air Pollution (Air Convention); Thünen Institute: Eberswalde, Germany, 2020; pp. 1–98. [Google Scholar]
- Walthert, L.; Ganthaler, A.; Mayr, S.; Saurer, M.; Waldner, P.; Walser, M.; Zweifel, R.; von Arx, G. From the Comfort Zone to Crown Dieback: Sequence of Physiological Stress Thresholds in Mature European Beech Trees across Progressive Drought. Sci. Total Environ. 2021, 753, 141792. [Google Scholar] [CrossRef]
- Wohlgemuth, T.; Kistler, M.; Aymon, C.; Hagedorn, F.; Gessler, A.; Gossner, M.M.; Queloz, V.; Vögtli, I.; Wasem, U.; Vitasse, Y.; et al. Früher Laubfall der Buche während der Sommertrockenheit 2018: Resistenz oder Schwächesymptom? Schw. Zeit. For. 2020, 171, 257–269. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European Tree Species to Drought Stress in Mixed versus Pure Forests: Evidence of Stress Release by Inter-Specific Facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of Drought Tolerance in Major European Temperate Forest Trees: Climatic Drivers and Levels of Variability. Glob. Change Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef] [PubMed]
- Hammond, W.M.; Williams, A.P.; Abatzoglou, J.T.; Adams, H.D.; Klein, T.; López, R.; Sáenz-Romero, C.; Hartmann, H.; Breshears, D.D.; Allen, C.D. Global field observations of tree die-off reveal hotter-drought fingerprint for Earth’s forests. Nat. Commun. 2022, 13, 1761. [Google Scholar] [CrossRef] [PubMed]
- Senf, C.; Seidl, R. Persistent Impacts of the 2018 Drought on Forest Disturbance Regimes in Europe. Biogeosciences 2021, 18, 5223–5230. [Google Scholar] [CrossRef]
- Muñoz-Sabater, J. ERA5-Land Monthly Averaged Data from 1981 to Present; Copernicus Climate Change Service (C3S) Climate Data Store (CDS), 2019. Available online: https://cds.climate.copernicus.eu/cdsapp#!/dataset/10.24381/cds.68d2bb30?tab=overview (accessed on 12 March 2022).
- Deutscher Wetterdienst—DWD. Wetter und Klima—Temperature and Precipitation. Available online: https://www.dwd.de/EN/climate_environment/climatemonitoring/europe/europe_node.html (accessed on 1 July 2022).
- Ramonet, M.; Ciais, P.; Apadula, F.; Bartyzel, J.; Bastos, A.; Bergamaschi, P.; Blanc, P.E.; Brunner, D.; Caracciolo Di Torchiarolo, L.; Calzolari, F.; et al. The Fingerprint of the Summer 2018 Drought in Europe on Ground-Based Atmospheric CO2 Measurements. Philos. Trans. R. Soc. B 2020, 375, 20190513. [Google Scholar] [CrossRef]
- Rakovec, O.; Samaniego, L.; Hari, V.; Markonis, Y.; Moravec, V.; Thober, S.; Hanel, M.; Kumar, R. The 2018–2020 Multi-Year Drought Sets a New Benchmark in Europe. Earth’s Future 2022, 10, EF002394. [Google Scholar] [CrossRef]
- Shah, J.; Hari, V.; Rakovec, O.; Markonis, Y.; Samaniego, L.; Mishra, V.; Hanel, M.; Hinz, C.; Kumar, R. Increasing Footprint of Climate Warming on Flash Droughts Occurrence in Europe. Environ. Res. Lett. 2022, 17, 064017. [Google Scholar] [CrossRef]
- Großmann, M. Extremjahr 2018 hinterlässt Spuren im Nationalpark Hainich. Trockenheit, hohe Temperaturen und permanente Sonneneinstrahlung bringen selbst Rotbuchen an ihre Grenzen. Press Release—Nationalpark Hainich, 28 June 2019. Available online: https://www.nationalpark-hainich.de/de/aktuelles/aktuelles-presse/einzelansicht/extremjahr-2018-hinterlaesst-spuren-im-nationalpark-hainich.html (accessed on 25 July 2021).
- Corcobado, T.; Cech, T.L.; Brandstetter, M.; Daxer, A.; Hüttler, C.; Kudláček, T.; Jung, M.H.; Jung, T. Decline of European Beech in Austria: Involvement of Phytophthora Spp. and Contributing Biotic and Abiotic Factors. Forests 2020, 11, 895. [Google Scholar] [CrossRef]
- Langer, G.J.; Bußkamp, J. Fungi Associated with Woody Tissues of European Beech and Their Impact on Tree Health. Front. Microbiol. 2021, 12, 2476. [Google Scholar] [CrossRef]
- Purahong, W.; Tanunchai, B.; Wahdan, S.F.M.; Buscot, F.; Schulze, E.D. Molecular Screening of Microorganisms Associated with Discolored Wood in Dead European Beech Trees Suffered from Extreme Drought Event Using Next Generation Sequencing. Plants 2021, 10, 2092. [Google Scholar] [CrossRef]
- Thornthwaite, C.W. An approach towards a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Beguería, S.; Vicente-Serrano, S.M. SPEI: Calculation of the Standardised Precipitation-Evapotranspiration Index. R Package Version 1.7, 2019. Available online: https://CRAN.R-project.org/package=SPEI (accessed on 11 May 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 11 May 2022).
- Eurostat. Geographical Information and Maps—GISCO. Shapefile-Countries 2020, 1:1 Million. Available online: https://ec.europa.eu/eurostat/web/gisco/geodata/reference-data/administrative-units-statistical-units/countries (accessed on 16 February 2022).
- Pierce, D. ncdf4: Interface to Unidata netCDF (Version 4 or Earlier) Format Data Files. R Package Version 1.19; 2021. Available online: https://CRAN.R-project.org/package=ncdf4 (accessed on 11 February 2022).
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 3.5-2; 2021. Available online: https://CRAN.R-project.org/package=raster (accessed on 11 February 2022).
- Rebetez, M.; Mayer, H.; Dupont, O.; Schindler, D.; Gartner, K.; Kropp, J.P.; Menzel, A. Heat and Drought 2003 in Europe: A Climate Synthesis. Ann. For. Sci. 2006, 63, 569–577. [Google Scholar] [CrossRef] [Green Version]
- Bastos, A.; Fu, Z.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Pongratz, J.; Weber, U.; Reichstein, M.; Anthoni, P.; Arneth, A.; et al. Impacts of Extreme Summers on European Ecosystems: A Comparative Analysis of 2003, 2010 and 2018. Philos. Trans. R. Soc. B 2020, 375, 20190507. [Google Scholar] [CrossRef]
- Schnabel, F.; Purrucker, S.; Schmitt, L.; Engelmann, R.A.; Kahl, A.; Richter, R.; Seele-Dilbat, C.; Skiadaresis, G.; Wirth, C. Cumulative Growth and Stress Responses to the 2018–2019 Drought in a European Floodplain Forest. Glob. Change Biol. 2022, 28, 1870–1883. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant Responses to Rising Vapor Pressure Deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a Potent Driver of Regional Forest Drought Stress and Tree Mortality. Nat. Clim. Change 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Bartelink, H.H. Allometric Relationships for Biomass and Leaf Area of Beech (Fagus sylvatica L). Ann. Sci. For. 1997, 54, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Thomas, F.M.; Bögelein, R.; Werner, W. Interaction between Douglas Fir and European Beech—Investigations in Pure and Mixed Stands. Forstarchiv 2015, 86, 83–91. [Google Scholar]
- Pretzsch, H.; Uhl, E.; Biber, P.; Schütze, G.; Coates, K.D. Change of Allometry between Coarse Root and Shoot of Lodgepole Pine (Pinus contorta DOUGL. Ex. LOUD) along a Stress Gradient in the Sub-Boreal Forest Zone of British Columbia. Scand. J. For. Res. 2012, 27, 532–544. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef] [PubMed]
- Peuke, A.D.; Schraml, C.; Hartung, W.; Rennenberg, H. Identification of drought-sensitive beech ecotypes by physiological parameters. New Phytol. 2002, 154, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Szejner, P.; Belmecheri, S.; Ehleringer, J.R.; Monson, R.K. Recent Increases in Drought Frequency Cause Observed Multi-Year Drought Legacies in the Tree Rings of Semi-Arid Forests. Oecologia 2020, 192, 241–259. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of Tree Mortality under Drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef]
- Stojnić, S.; Suchocka, M.; Benito-Garzón, M.; Torres-Ruiz, J.M.; Cochard, H.; Bolte, A.; Cocozza, C.; Cvjetković, B.; De Luis, M.; Martinez-Vilalta, J.; et al. Variation in Xylem Vulnerability to Embolism in European Beech from Geographically Marginal Populations. Tree Physiol. 2018, 38, 173–185. [Google Scholar] [CrossRef]
- Leuschner, C. Drought Response of European Beech (Fagus sylvatica L.)—A Review. Perspect. Plant Ecol. Evol. Syst. 2020, 47, 125576. [Google Scholar] [CrossRef]
- Salmon, Y.; Dietrich, L.; Sevanto, S.; Hölttä, T.; Dannoura, M.; Epron, D. Drought impacts on tree phloem: From cell-level responses to ecological significance. Tree Physiol. 2019, 39, 173–191. [Google Scholar] [CrossRef] [Green Version]
- Hesse, B.D.; Goisser, M.; Hartmann, H.; Grams, T.E.E. Repeated summer drought delays sugar export from the leaf and impairs phloem transport in mature beech. Tree Physiol. 2019, 39, 192–200. [Google Scholar] [CrossRef]
- Camarero, J.J. The drought–dieback–death conundrum in trees and forests. Plant Ecol. Div. 2021, 14, 1–12. [Google Scholar] [CrossRef]
- Tallieu, C.; Badeau, V.; Allard, D.; Nageleisen, L.M.; Bréda, N. Year-to-Year Crown Condition Poorly Contributes to Ring Width Variations of Beech Trees in French ICP Level I Network. For. Ecol. Manag. 2020, 465, 118071. [Google Scholar] [CrossRef]
- Gérard, B.; Bréda, N. Radial Distribution of Carbohydrate Reserves in the Trunk of Declining European Beech Trees (Fagus sylvatica L.). Ann. For. Sci. 2014, 71, 675–682. [Google Scholar] [CrossRef] [Green Version]
- Weithmann, G.; Schuldt, B.; Link, R.M.; Heil, D.; Hoeber, S.; John, H.; Müller-Haubold, H.; Schüller, L.M.; Schumann, K.; Leuschner, C. Leaf Trait Modification in European Beech Trees in Response to Climatic and Edaphic Drought. Plant Biol. 2022, 24, 1272–1286. [Google Scholar] [CrossRef] [PubMed]
- Beck, W. Impact of drought and heat on tree and stand vitality—Results of the study commissioned by the Federal Ministry of Food, Agriculture and Consumer Protection. In TRACE, Scientific Technical Report 11/07; Maaten-Theunissen, M., Spiecker, H., Gärtner, H., Helle, G., Heinrich, I., Eds.; GFZ Potsdam: Potsdam, Germany, 2011; Volume 9, pp. 20–27. [Google Scholar]
- Beck, W.; Heußner, K.U. Increased sensitivity in ring width series of common beech after 1990—Climatic impact or normal patterns due to ageing. In TRACE, Scientific Technical Report 12/03; Gärtner, H., Philippe, R., Montés, P., Bertel, O., Heinrich, I., Helle, G., Eds.; GFZ Potsdam: Potsdam, Germany, 2012; Volume 10, pp. 13–19. [Google Scholar]
- Anderegg, W.R.L.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive Drought Legacies in Forest Ecosystems and Their Implications for Carbon Cycle Models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [Green Version]
- Tomasella, M. Hydraulics of European Beech and Norway Spruce under Experimental Drought. Ph.D. Thesis, Technical University of Munich, Munich, Germany, 24 January 2018. [Google Scholar]
- Petrík, P.; Zavadilová, I.; Šigut, L.; Kowalska, N.; Petek-Petrik, A.; Szatniewska, J.; Jocher, G.; Pavelka, M. Impact of Environmental Conditions and Seasonality on Ecosystem Transpiration and Evapotranspiration Partitioning (T/ET Ratio) of Pure European Beech Forest. Water 2022, 14, 3015. [Google Scholar] [CrossRef]
- Kahmen, A.; Basler, D.; Hoch, G.; Link, R.M.; Schuldt, B.; Zahnd, C.; Arend, M. Root water uptake depth determines the hydraulic vulnerability of temperate European tree species during the extreme 2018 drought. Plant Biol. 2022, 24, 1224–1239. [Google Scholar] [CrossRef] [PubMed]
- Bolte, A.; Czajkowski, T.; Kompa, T. The North-Eastern Distribution Range of European Beech—A Review. Forestry 2007, 80, 413–429. [Google Scholar] [CrossRef] [Green Version]
- Von Wühlish, G.; Muhs, H.J. Current state of European beech (Fagus sylvatica L.) Forests in Germany. In COST Action E 52—Genetic Resources of Beech in Europe—Current State; Frýdl, J., Novotný, P., Fennessy, J., von Wühlisch, G., Eds.; Johann Heinrich von Thünen-Institut: Braunschweig, Germany, 2011; pp. 113–121. [Google Scholar]
- Lorenz, M.; Becher, G.; Mues, V.; Fischer, R.; Ulrich, E.; Dobbertin, M.; Stofer, S. Forest Condition in Europe, 2004 Technical Report; Federal Research Centre for Forestry and Forest Products (BFH): Hamburg, Germany, 2004; pp. 1–96. [Google Scholar]
- Lorenz, M.; Becher, G.; Mues, V.; Fischer, R.; Becker, R.; Calatayud, V.; Dise, N.; Krause, G.H.M.; Sanz, M.; Ulrich, E. Forest Condition in Europe, 2005 Technical Report; Federal Research Centre for Forestry and Forest Products (BFH): Hamburg, Germany, 2005; pp. 1–96. [Google Scholar]
- European Soil Bureau Network. Soil Atlas of Europe; Office for Official Publications of the European Communities, European Commission: Luxembourg, 2005; pp. 1–128. [Google Scholar]
- Krupková, L.; Havránková, K.; Krejza, J.; Sedlák, P.; Marek, M.V. Impact of Water Scarcity on Spruce and Beech Forests. J. For. Res. 2019, 30, 899–909. [Google Scholar] [CrossRef]
- Gessler, A.; Bächli, L.; Freund, E.R.; Treydte, K.; Schaub, M.; Haeni, M.; Weiler, M.; Seeger, S.; Marshall, J.; Hug, C.; et al. Drought Reduces Water Uptake in Beech from the Drying Topsoil, but No Compensatory Uptake Occurs from Deeper Soil Layers. New Phytol. 2022, 233, 194–206. [Google Scholar] [CrossRef]
- Meier, I.C.; Knutzen, F.; Eder, L.M.; Müller-Haubold, H.; Goebel, M.O.; Bachmann, J.; Hertel, D.; Leuschner, C. The Deep Root System of Fagus Sylvatica on Sandy Soil: Structure and Variation Across a Precipitation Gradient. Ecosystems 2018, 21, 280–296. [Google Scholar] [CrossRef]
- Stojanovic, M.; Drumond, A.; Nieto, R.; Gimeno, L. Anomalies in Moisture Supply during the 2003 Drought Event in Europe: A Lagrangian Analysis. Water 2018, 10, 467. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.L.; Broquet, G.; Gerbig, C.; Koch, T.; Lang, M.; Monteil, G.; Munassar, S.; Nickless, A.; Scholze, M.; Ramonet, M.; et al. Changes in Net Ecosystem Exchange over Europe during the 2018 Drought Based on Atmospheric Observations. Philos. Trans. R. Soc. B 2020, 375, 20190512. [Google Scholar] [CrossRef]
- Copernicus Climate Change Service—C3S. Dry and Warm Spring and Summer. Available online: https://climate.copernicus.eu/dry-and-warm-spring-and-summer (accessed on 25 July 2021).
- Laaha, G.; Gauster, T.; Tallaksen, L.M.; Vidal, J.P.; Stahl, K.; Prudhomme, C.; Heudorfer, B.; Vlnas, R.; Ionita, M.; Van Lanen, H.A.J.; et al. The European 2015 Drought from a Hydrological Perspective. Hydrol. Earth Syst. Sci. 2017, 21, 3001–3024. [Google Scholar] [CrossRef] [Green Version]
- Hanel, M.; Rakovec, O.; Markonis, Y.; Máca, P.; Samaniego, L.; Kyselý, J.; Kumar, R. Revisiting the Recent European Droughts from a Long-Term Perspective. Sci. Rep. 2018, 8, 9499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brück-Dyckhoff, C.; Petercord, R.; Schopf, R. Vitality Loss of European Beech (Fagus sylvatica L.) and Infestation by the European Beech Splendour Beetle (Agrilus viridis L., Buprestidae, coleoptera). For. Ecol. Manag. 2019, 432, 150–156. [Google Scholar] [CrossRef]
- Jung, T. Beech Decline in Central Europe Driven by the Interaction between Phytophthora Infections and Climatic Extremes. For. Pathol. 2009, 39, 73–94. [Google Scholar] [CrossRef]
- Kahle, H.P. Impact of the drought in 2003 on intra- and inter-annual stem radial growth of beech and spruce along an altitudinal gradient in the Black Forest, Germany. In Proceedings of the Dendrosymposium: TRACE—Tree Rings in Archaeology, Climatology and Ecology, Fribourg, Switzerland, 21–23 April 2005. [Google Scholar]
- Leuzinger, S.; Zotz, G.; Asshoff, R.; Körner, C. Responses of Deciduous Forest Trees to Severe Drought in Central Europe. Tree Physiol. 2005, 25, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Mette, T.; Dolos, K.; Meinardus, C.; Bräuning, A.; Reineking, B.; Blaschke, M.; Pretzsch, H.; Beierkuhnlein, C.; Gohlke, A.; Wellstein, C. Climatic Turning Point for Beech and Oak under Climate Change in Central Europe. Ecosphere 2013, 4, 1–19. [Google Scholar] [CrossRef]
- Dobbertin, M. Tree Growth as Indicator of Tree Vitality and of Tree Reaction to Environmental Stress: A Review. Eur. J. For. Res. 2005, 124, 319–333. [Google Scholar] [CrossRef]
- Landesanstalt für Wald und Forstwirtschaft Bayern—LWF. Waldzustandsbericht; Bayerisches Staatsministerium für Landwirtschaft und Forsten: Munich, Germany, 2004. [Google Scholar]
- Chakraborty, T.; Saha, S.; Reif, A. Decrease in Available Soil Water Storage Capacity Reduces Vitality of Young Understorey European Beeches (Fagus sylvatica l.)—A Case Study from the Black Forest, Germany. Plants 2013, 2, 676–698. [Google Scholar] [CrossRef] [PubMed]
- Rukh, S.; Poschenrieder, W.; Heym, M.; Pretzsch, H. Drought Resistance of Norway Spruce (Picea abies [L.] Karst) and European Beech (Fagus sylvatica [L.]) in Mixed vs. Monospecific Stands and on Dry vs. Wet Sites. From Evidence at the Tree Level to Relevance at the Stand Level. Forests 2020, 11, 639. [Google Scholar] [CrossRef]
- Walentowski, H.; Falk, W.; Mette, T.; Kunz, J.; Bräuning, A.; Meinardus, C.; Zang, C.; Sutcliffe, L.M.E.; Leuschner, C. Assessing Future Suitability of Tree Species under Climate Change by Multiple Methods: A Case Study in Southern Germany. Ann. For. Res. 2017, 60, 101–126. [Google Scholar] [CrossRef] [Green Version]
- Kohler, M.; Köckemann, B.; Peichl, M.; Schmitt, J.; Reif, A. Impacts of the drought 2003 on the crown condition of suppressed and intermediate beech trees (Fagus sylvatica L.) at the ecotone between beech and downy oak forest in the nature reserve Innerberg, Südbaden. Allg. For. Jagd. 2006, 177, 86–91. [Google Scholar]
- Lakatos, F.; Molnár, M. Mass Mortality of Beech (Fagus sylvatica L.) in South-West Hungary. Acta Silv. Lignaria Hung. 2009, 5, 75–82. [Google Scholar]
- Orlikowski, L.B.; Oszako, T.; Szkuta, G. First Record on Phytophthora Spp. Associated with the Decline of European Beech Stand in South-West Poland. Phytopathol. Pol. 2006, 42, 37–46. [Google Scholar]
- Mihál, I.; Bučinová, K.; Pavlíková, J. Mycoflora of beech forests in the Kremnické vrchy Mts (Central Slovakia). Folia Oecol. 2009, 36, 23–31. [Google Scholar]
- Nussbaumer, A.; Meusburger, K.; Schmitt, M.; Waldner, P.; Gehrig, R.; Haeni, M.; Rigling, A.; Brunner, I.; Thimonier, A. Extreme Summer Heat and Drought Lead to Early Fruit Abortion in European Beech. Sci. Rep. 2020, 10, 5334. [Google Scholar] [CrossRef] [Green Version]
- Weithmann, G.; Link, R.M.; Banzragch, B.E.; Würzberg, L.; Leuschner, C.; Schuldt, B. Soil Water Availability and Branch Age Explain Variability in Xylem Safety of European Beech in Central Europe. Oecologia 2022, 198, 629–644. [Google Scholar] [CrossRef]
- Hacke, U.; Sauter, J.J. Vulnerability of Xylem to Embolism in Relation to Leaf Water Potential and Stomatal Conductance in Fagus sylvatica f. Purpurea and Populus balsamifera. J. Exp. Bot. 1995, 46, 1177–1183. [Google Scholar] [CrossRef] [Green Version]
- Herbette, S.; Wortemann, R.; Awad, H.; Huc, R.; Cochard, H.; Barigah, T.S. Insights into Xylem Vulnerability to Cavitation in Fagus sylvatica L.: Phenotypic and Environmental Sources of Variability. Tree Physiol. 2010, 30, 1448–1455. [Google Scholar] [CrossRef] [PubMed]
- Aranda, I.; Cano, F.J.; Gascó, A.; Cochard, H.; Nardini, A.; Mancha, J.A.; López, R.; Sánchez-Gómez, D. Variation in Photosynthetic Performance and Hydraulic Architecture across European Beech (Fagus sylvatica L.) Populations Supports the Case for Local Adaptation to Water Stress. Tree Physiol. 2015, 35, 34–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuldt, B.; Knutzen, F.; Delzon, S.; Jansen, S.; Müller-Haubold, H.; Burlett, R.; Clough, Y.; Leuschner, C. How Adaptable Is the Hydraulic System of European Beech in the Face of Climate Change-Related Precipitation Reduction? New Phytol. 2016, 210, 443–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer, C.; Grams, T.E.E.; Rötzer, T.; Feldermann, A.; Pretzsch, H. Drought stress reaction of growth and δ13C in tree rings of European beech and Norway spruce in monospecific versus mixed stands along a precipitation gradient. Forests 2017, 8, 177. [Google Scholar] [CrossRef] [Green Version]
- Obladen, N.; Dechering, P.; Skiadaresis, G.; Tegel, W.; Keßler, J.; Höllerl, S.; Kaps, S.; Hertel, M.; Dulamsuren, C.; Seifert, T.; et al. Tree Mortality of European Beech and Norway Spruce Induced by 2018-2019 Hot Droughts in Central Germany. Agric. For. Meteorol. 2021, 307, 108482. [Google Scholar] [CrossRef]
- Debel, A.; Meier, W.J.H.; Bräuning, A. Climate Signals for Growth Variations of F. sylvatica, P. abies, and P. sylvestris in Southeast Germany over the Past 50 Years. Forests 2021, 12, 1433. [Google Scholar] [CrossRef]
- Michel, A.; Prescher, A.K.; Schwärzel, K. (Eds.) Forest Condition in Europe: 2019 Technical Report of ICP Forests; Report under the UNECE Convention on Long-range Transboundary Air Pollution (Air Convention); BFW—Austrian Research Centre for Forests: Vienna, Austria, 2019; pp. 1–108. [Google Scholar]
- Berliner Forsten—BF. Waldzustandsbericht 2021 des Landes Berlin; Senatsverwaltung Umwelt, Verkehr und Klimaschutz (SENUVK), Berliner Forsten: Berlin, Germany, 2021; pp. 1–27. [Google Scholar]
- Landeskompetenzzentrum Forst Eberswalde—LFE. Waldzustandsbericht 2021 des Landes Brandenburg; LFE des Landesbetrieb Forst Brandenburg—Ministerium für Landwirtschaft, Umwelt und Klimaschutz des Landes Brandenburg (MLUK): Eberswalde, Germany, 2021; pp. 1–48. [Google Scholar]
- Forstliches Forschungs- und Kompetenzzentrum des Thüringen Forst—FFK. Waldzustandsbericht 2021, Forstliches Umweltmonitoring in Thüringen; Thüringer Ministerium für Infrastruktur und Landwirtschaft (TMIL), Referat Presse: Erfurt, Germany, 2021; pp. 1–29. [Google Scholar]
- Landesforst Mecklenburg-Vorpommern—LM. Waldzustandsbericht 2019, Ergebnisse der Waldzustandserhebung; Ministerium für Landwirtschaft und Umwelt Mecklenburg-Vorpommern: Schwerin, Germany, 2019; pp. 1–47. [Google Scholar]
- Gillner, S.; Rüger, N.; Roloff, A.; Berger, U. Low Relative Growth Rates Predict Future Mortality of Common Beech (Fagus sylvatica L.). For. Ecol. Manag. 2013, 302, 372–378. [Google Scholar] [CrossRef]
- Braun, S.; Hopf, S.E.; Tresch, S.; Remund, J.; Schindler, C. 37 Years of Forest Monitoring in Switzerland: Drought Effects on Fagus sylvatica. Front. For. Glob. Change 2021, 4, 159. [Google Scholar] [CrossRef]
- Brun, P.; Psomas, A.; Ginzler, C.; Thuiller, W.; Zappa, M.; Zimmermann, N.E. Large-Scale Early-Wilting Response of Central European Forests to the 2018 Extreme Drought. Glob. Change Biol. 2020, 26, 7021–7035. [Google Scholar] [CrossRef]
- Chakraborty, T.; Saha, S.; Matzarakis, A.; Reif, A. Influence of Multiple Biotic and Abiotic Factors on the Crown Die-Back of European Beech Trees at Their Drought Limit. Flora 2017, 229, 58–70. [Google Scholar] [CrossRef]
- Piedallu, C.; Gégout, J.C.; Perez, V.; Lebourgeois, F. Soil Water Balance Performs Better than Climatic Water Variables in Tree Species Distribution Modelling. Glob. Ecol. Biogeogr. 2013, 22, 470–482. [Google Scholar] [CrossRef]
- Trnka, M.; Vizina, A.; Hanel, M.; Balek, J.; Fischer, M.; Hlavinka, P.; Semerádová, D.; Štěpánek, P.; Zahradníček, P.; Skalák, P.; et al. Increasing Available Water Capacity as a Factor for Increasing Drought Resilience or Potential Conflict over Water Resources under Present and Future Climate Conditions. Agric. Water Manag. 2022, 264, 107460. [Google Scholar] [CrossRef]
- Luo, W.; Elser, J.J.; Lü, X.T.; Wang, Z.; Bai, E.; Yan, C.; Wang, C.; Li, M.H.; Zimmermann, N.E.; Han, X.; et al. Plant Nutrients Do Not Covary with Soil Nutrients under Changing Climatic Conditions. Glob. Biogeochem. Cycles 2015, 29, 1298–1308. [Google Scholar] [CrossRef] [Green Version]
- Simpson, A.H.; Richardson, S.J.; Laughlin, D.C. Soil–Climate Interactions Explain Variation in Foliar, Stem, Root and Reproductive Traits across Temperate Forests. Glob. Ecol. Biogeogr. 2016, 25, 964–978. [Google Scholar] [CrossRef]
- Toïgo, M.; Nicolas, M.; Jonard, M.; Croisé, L.; Nageleisen, L.M.; Jactel, H. Temporal Trends in Tree Defoliation and Response to Multiple Biotic and Abiotic Stresses. For. Ecol. Manag. 2020, 477, 118476. [Google Scholar] [CrossRef]
- Grace, J.; Berninger, F.; Nagy, L. Impacts of Climate Change on the Tree Line. Ann. Bot. 2002, 90, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Michelot, A.; Simard, S.; Rathgeber, C.; Dufrêne, E.; Damesin, C. Comparing the Intra-Annual Wood Formation of Three European Species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as Related to Leaf Phenology and Non-Structural Carbohydrate Dynamics. Tree Physiol. 2012, 32, 1033–1045. [Google Scholar] [CrossRef] [Green Version]
- Gavinet, J.; Ourcival, J.M.; Limousin, J.M. Rainfall Exclusion and Thinning Can Alter the Relationships between Forest Functioning and Drought. New Phytol. 2019, 223, 1267–1279. [Google Scholar] [CrossRef]
- McDowell, N.G.; Sapes, G.; Pivovaroff, A.; Adams, H.D.; Allen, C.D.; Anderegg, W.R.L.; Arend, M.; Breshears, D.D.; Brodribb, T.; Choat, B.; et al. Mechanisms of Woody-Plant Mortality under Rising Drought, CO2 and Vapour Pressure Deficit. Nat. Rev. Earth Environ. 2022, 3, 294–308. [Google Scholar] [CrossRef]
- Seidling, W.; Ziche, D.; Beck, W. Climate responses and interrelations of stem increment and crown transparency in Norway spruce, Scots pine and common beech. For. Ecol. Manag. 2012, 284, 196–204. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Cantero, A.; Sánchez-Salguero, R.; Sánchez-Miranda, A.; Granda, E.; Serra-Maluquer, X.; Ibáñez, R. Forest Growth Responses to Drought at Short- and Long-Term Scales in Spain: Squeezing the Stress Memory from Tree Rings. Front. Ecol. Evol. 2018, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Latte, N.; Lebourgeois, F.; Claessens, H. Increased Tree-Growth Synchronization of Beech (Fagus Sylvatica L.) in Response to Climate Change in Northwestern Europe. Dendrochronologia 2015, 33, 69–77. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of Tree Resilience: Effects of Successive Low-Growth Episodes in Old Ponderosa Pine Forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Gessler, A.; Bottero, A.; Marshall, J.; Arend, M. The Way Back: Recovery of Trees from Drought and Its Implication for Acclimation. New Phytol. 2020, 228, 1704–1709. [Google Scholar] [CrossRef] [PubMed]
- DeSoto, L.; Cailleret, M.; Sterck, F.; Jansen, S.; Kramer, K.; Robert, E.M.R.; Aakala, T.; Amoroso, M.M.; Bigler, C.; Camarero, J.J.; et al. Low Growth Resilience to Drought Is Related to Future Mortality Risk in Trees. Nat. Commun. 2020, 11, 545. [Google Scholar] [CrossRef] [Green Version]
- Peltier, D.M.P.; Fell, M.; Ogle, K. Legacy Effects of Drought in the Southwestern United States: A Multi-Species Synthesis. Ecol. Monogr. 2016, 86, 312–326. [Google Scholar] [CrossRef]
- Ogle, K.; Barber, J.J.; Barron-Gafford, G.A.; Bentley, L.P.; Young, J.M.; Huxman, T.E.; Loik, M.E.; Tissue, D.T. Quantifying Ecological Memory in Plant and Ecosystem Processes. Ecol. Lett. 2015, 18, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Bose, A.K.; Scherrer, D.; Camarero, J.J.; Ziche, D.; Babst, F.; Bigler, C.; Bolte, A.; Dorado-Liñán, I.; Etzold, S.; Fonti, P.; et al. Climate Sensitivity and Drought Seasonality Determine Post-Drought Growth Recovery of Quercus petraea and Quercus robur in Europe. Sci. Total Environ. 2021, 784, 147222. [Google Scholar] [CrossRef]
- Huang, M.; Wang, X.; Keenan, T.F.; Piao, S. Drought Timing Influences the Legacy of Tree Growth Recovery. Glob. Change Biol. 2018, 24, 3546–3559. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.J.; O’Grady, A.P.; Pinkard, E.A.; Brodribb, T.J.; Arndt, S.K.; Blackman, C.J.; Duursma, R.A.; Fensham, R.J.; Hilbert, D.W.; Nitschke, C.R.; et al. An Ecoclimatic Framework for Evaluating the Resilience of Vegetation to Water Deficit. Glob. Change Biol. 2016, 22, 1677–1689. [Google Scholar] [CrossRef]
- Ferretti, M.; Bacaro, G.; Brunialti, G.; Calderisi, M.; Croisé, L.; Frati, L.; Nicolas, M. Tree Canopy Defoliation Can Reveal Growth Decline in Mid-Latitude Temperate Forests. Ecol. Indic. 2021, 127, 107749. [Google Scholar] [CrossRef]
- Johnson, D.M.; Sherrard, M.E.; Domec, J.C.; Jackson, R.B. Role of Aquaporin Activity in Regulating Deep and Shallow Root Hydraulic Conductance during Extreme Drought. Trees 2014, 28, 1323–1331. [Google Scholar] [CrossRef]
- Brinkmann, N.; Eugster, W.; Buchmann, N.; Kahmen, A. Species-Specific Differences in Water Uptake Depth of Mature Temperate Trees Vary with Water Availability in the Soil. Plant Biol. 2019, 21, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeger, S.; Weiler, M. Temporal Dynamics of Tree Xylem Water Isotopes: In Situ Monitoring and Modeling. Biogeosciences 2021, 18, 4603–4627. [Google Scholar] [CrossRef]
- Lüttschwager, D.; Jochheim, H. Drought Primarily Reduces Canopy Transpiration of Exposed Beech Trees and Decreases the Share of Water Uptake from Deeper Soil Layers. Forests 2020, 11, 537. [Google Scholar] [CrossRef]
- Kannenberg, S.A.; Maxwell, J.T.; Pederson, N.; D’Orangeville, L.; Ficklin, D.L.; Phillips, R.P. Drought Legacies Are Dependent on Water Table Depth, Wood Anatomy and Drought Timing across the Eastern US. Ecol. Lett. 2019, 22, 119–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhellemont, M.; Sousa-Silva, R.; Maes, S.L.; Van den Bulcke, J.; Hertzog, L.; De Groote, S.R.E.; Van Acker, J.; Bonte, D.; Martel, A.; Lens, L.; et al. Distinct Growth Responses to Drought for Oak and Beech in Temperate Mixed Forests. Sci. Total Environ. 2019, 650, 3017–3026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, P.; Bugmann, H.; Pluess, A.R.; Walthert, L.; Rigling, A. Drought Response and Changing Mean Sensitivity of European Beech Close to the Dry Distribution Limit. Trees 2013, 27, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Arnič, D.; Gričar, J.; Jevšenak, J.; Božič, G.; von Arx, G.; Prislan, P. Different Wood Anatomical and Growth Responses in European Beech (Fagus sylvatica L.) at Three Forest Sites in Slovenia. Front. Plant Sci. 2021, 12, 1551. [Google Scholar] [CrossRef]
- Kasper, J.; Leuschner, C.; Walentowski, H.; Petritan, A.M.; Weigel, R. Winners and Losers of Climate Warming: Declining Growth in Fagus and Tilia vs. Stable Growth in Three Quercus Species in the Natural Beech–Oak Forest Ecotone (Western Romania). For. Ecol. Manag. 2022, 506, 119892. [Google Scholar] [CrossRef]
- Kolář, T.; Čermák, P.; Trnka, M.; Žid, T.; Rybníček, M. Temporal Changes in the Climate Sensitivity of Norway Spruce and European Beech along an Elevation Gradient in Central Europe. Agric. For. Meteorol. 2017, 239, 24–33. [Google Scholar] [CrossRef]
- Gennaretti, F.; Ogée, J.; Sainte-Marie, J.; Cuntz, M. Mining Ecophysiological Responses of European Beech Ecosystems to Drought. Agric. For. Meteorol. 2020, 280, 107780. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; Čufar, K.; de Luis, M.; Merela, M.; Rossi, S. Growing Season and Radial Growth Predicted for Fagus Sylvatica under Climate Change. Clim. Change 2019, 153, 181–197. [Google Scholar] [CrossRef]
- Giagli, K.; Gričar, J.; Vavrčík, H.; Menšík, L.; Gryc, V. The Effects of Drought on Wood Formation in Fagus sylvatica during two Contrasting Years. IAWA J. 2016, 37, 332–348. [Google Scholar] [CrossRef]
- Lebourgeois, F.; Bréda, N.; Ulrich, E.; Granier, A. Climate-Tree-Growth Relationships of European Beech (Fagus sylvatica L.) in the French Permanent Plot Network (RENECOFOR). Trees 2005, 19, 385–401. [Google Scholar] [CrossRef]
- Chakraborty, T.; Reif, A.; Matzarakis, A.; Saha, S. How Does Radial Growth of Water-Stressed Populations of European Beech (Fagus Sylvatica L.) Trees Vary under Multiple Drought Events? Forests 2021, 12, 129. [Google Scholar] [CrossRef]
- Hlásny, T.; Barcza, Z.; Fabrika, M.; Balázs, B.; Churkina, G.; Pajtík, J.; Sedmák, R.; Turcani, M. Climate change impacts on growth and carbon balance of forests in Central Europe. Clim. Res. 2011, 47, 219–236. [Google Scholar] [CrossRef] [Green Version]
- Dulamsuren, C.; Hauck, M.; Kopp, G.; Ruff, M.; Leuschner, C. European Beech Responds to Climate Change with Growth Decline at Lower, and Growth Increase at Higher Elevations in the Center of Its Distribution Range (SW Germany). Trees 2017, 31, 673–686. [Google Scholar] [CrossRef]
- Buras, A.; Menzel, A. Projecting Tree Species Composition Changes of European Forests for 2061–2090 under RCP 4.5 and RCP 8.5 Scenarios. Front. Plant Sci. 2019, 9, 1986. [Google Scholar] [CrossRef] [Green Version]
- Saltré, F.; Duputié, A.; Gaucherel, C.; Chuine, I. How Climate, Migration Ability and Habitat Fragmentation Affect the Projected Future Distribution of European Beech. Glob. Change Biol. 2015, 21, 897–910. [Google Scholar] [CrossRef]
- Jonard, F.; André, F.; Ponette, Q.; Vincke, C.; Jonard, M. Sap Flux Density and Stomatal Conductance of European Beech and Common Oak Trees in Pure and Mixed Stands during the Summer Drought of 2003. J. Hydrol. 2011, 409, 371–381. [Google Scholar] [CrossRef]
- Schwarz, J.A.; Bauhus, J. Benefits of Mixtures on Growth Performance of Silver Fir (Abies alba) and European Beech (Fagus sylvatica) Increase with Tree Size Without Reducing Drought Tolerance. Front. For. Glob. Change 2019, 2, 79. [Google Scholar] [CrossRef] [Green Version]
- Metz, J.; Annighöfer, P.; Schall, P.; Zimmermann, J.; Kahl, T.; Schulze, E.D.; Ammer, C. Site-Adapted Admixed Tree Species Reduce Drought Susceptibility of Mature European Beech. Glob. Change Biol. 2016, 22, 903–920. [Google Scholar] [CrossRef] [PubMed]
- Scharnweber, T.; Manthey, M.; Wilmking, M. Differential Radial Growth Patterns between Beech (Fagus sylvatica L.) and Oak (Quercus robur L.) on Periodically Waterlogged Soils. Tree Physiol. 2013, 33, 425–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossiord, C. Having the Right Neighbors: How Tree Species Diversity Modulates Drought Impacts on Forests. New Phytol. 2020, 228, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Hafner, B.D.; Tomasella, M.; Häberle, K.H.; Goebel, M.; Matyssek, R.; Grams, T.E.E. Hydraulic Redistribution under Moderate Drought among English Oak, European Beech and Norway Spruce Determined by Deuterium Isotope Labeling in a Split-Root Experiment. Tree Physiol. 2017, 37, 950–960. [Google Scholar] [CrossRef] [Green Version]
- Thurm, E.A.; Uhl, E.; Pretzsch, H. Mixture Reduces Climate Sensitivity of Douglas-Fir Stem Growth. For. Ecol. Manag. 2016, 376, 205–220. [Google Scholar] [CrossRef]
- Lévesque, M.; Walthert, L.; Weber, P. Soil Nutrients Influence Growth Response of Temperate Tree Species to Drought. J. Ecol. 2016, 104, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Ammer, C. Unraveling the importance of inter- and intraspecific competition for the adaptation of forests to climate change. In Progress in Botany, 1st ed.; Cánovas, F.M., Lüttge, U., Matyssek, R., Eds.; Springer: Cham, Switzerland, 2017; Volume 78, pp. 345–367. [Google Scholar]
- Williams, M.I.; Dumroese, R.K. Preparing for Climate Change: Forestry and Assisted Migration. J. For. 2013, 111, 287–297. [Google Scholar] [CrossRef]
- Spathelf, P.; Stanturf, J.; Kleine, M.; Jandl, R.; Chiatante, D.; Bolte, A. Adaptive Measures: Integrating Adaptive Forest Management and Forest Landscape Restoration. Ann. For. Sci. 2018, 75, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Dounavi, A.; Netzer, F.; Celepirovic, N.; Ivanković, M.; Burger, J.; Figueroa, A.G.; Schön, S.; Simon, J.; Cremer, E.; Fussi, B.; et al. Genetic and Physiological Differences of European Beech Provenances (F. sylvatica L.) Exposed to Drought Stress. For. Ecol. Manag. 2016, 361, 226–236. [Google Scholar] [CrossRef]
- Bolte, A.; Czajkowski, T.; Cocozza, C.; Tognetti, R.; De Miguel, M.; Pšidová, E.; Ditmarová, L.; Dinca, L.; Delzon, S.; Cochard, H.; et al. Desiccation and Mortality Dynamics in Seedlings of Different European Beech (Fagus sylvatica L.) Populations under Extreme Drought Conditions. Front. Plant Sci. 2016, 7, 751. [Google Scholar] [CrossRef] [Green Version]
- Sohn, J.A.; Saha, S.; Bauhus, J. Potential of Forest Thinning to Mitigate Drought Stress: A Meta-Analysis. For. Ecol. Manag. 2016, 380, 261–273. [Google Scholar] [CrossRef]
- Van der Maaten, E. Thinning Prolongs Growth Duration of European Beech (Fagus sylvatica L.) across a Valley in Southwestern Germany. For. Ecol. Manag. 2013, 306, 135–141. [Google Scholar] [CrossRef]
- Diaconu, D.; Kahle, H.P.; Spiecker, H. Thinning Increases Drought Tolerance of European Beech: A Case Study on Two Forested Slopes on Opposite Sides of a Valley. Eur. J. For. Res. 2017, 136, 319–328. [Google Scholar] [CrossRef]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate Forest Trees and Stands under Severe Drought: A Review of Ecophysiological Responses, Adaptation Processes and Long-Term Consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef] [Green Version]
- Breda, N.; Granier, A.; Aussenac, G. Effects of Thinning on Soil and Tree Water Relations, Transpiration and Growth in an Oak Forest (Quercus petraea (Matt.) Liebl.). Tree Physiol. 1995, 15, 295–306. [Google Scholar] [CrossRef]
- Meyer, P.; Spînu, A.P.; Mölder, A.; Bauhus, J. Management Alters Drought-Induced Mortality Patterns in European Beech (Fagus sylvatica L.) Forests. Plant Biol. 2022, 24, 1157–1170. [Google Scholar] [CrossRef]
- Bolte, A.; Höhl, M.; Hennig, P.; Schad, T.; Kroiher, F.; Seintsch, B.; Englert, H.; Rosenkranz, L. Zukunftsaufgabe Waldanpassung. AFZ Wald 2021, 76, 12–16. [Google Scholar]
- Intergovernmental Panel on Climate Change—IPCC. Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2022; pp. 1–151. [Google Scholar]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.T.; et al. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Ammer, C.; Fichtner, A.; Fischer, A.; Gossner, M.M.; Meyer, P.; Seidl, R.; Thomas, F.M.; Annighöfer, P.; Kreyling, J.; Ohse, B.; et al. Key Ecological Research Questions for Central European Forests. Basic Appl. Ecol. 2018, 32, 3–25. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
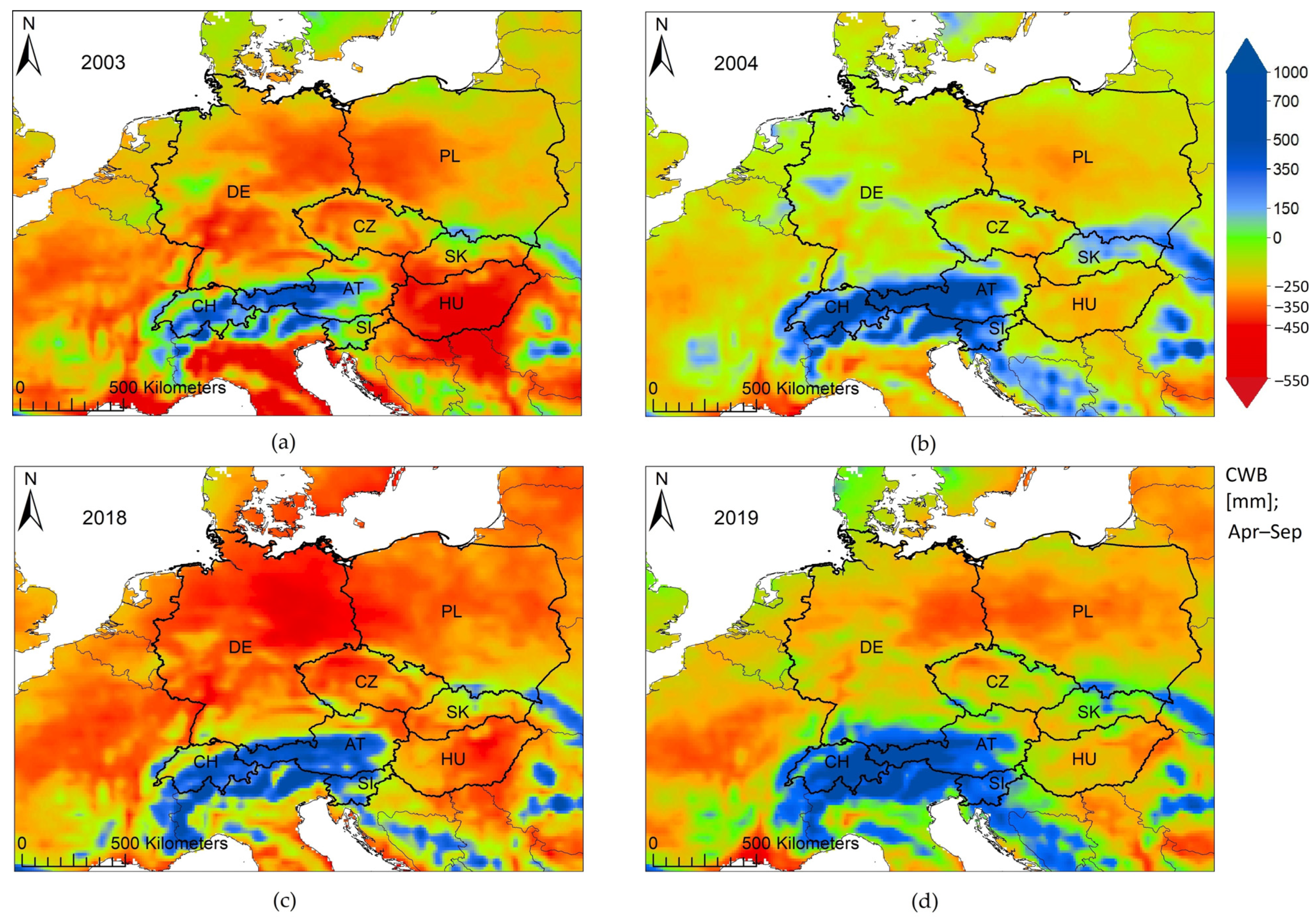
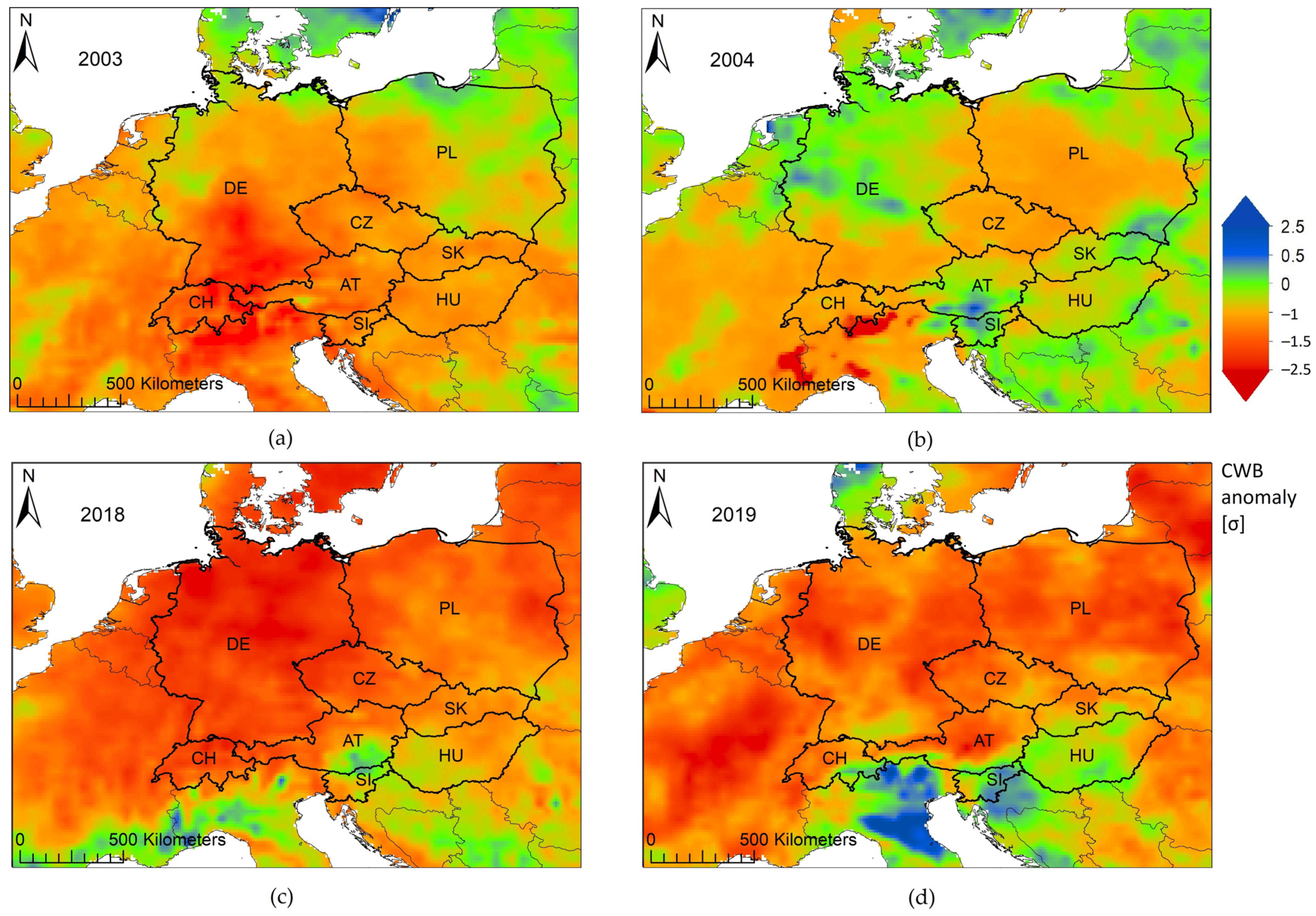
| Year | Unusual Weather Patterns | Temperature Anomalies (1961–1990) a | Precipitation Anomalies % (1961–1990) b | CWB c absolute sum (mm) | Standardized CWB Anomalies (1991–2020) d | References |
|---|---|---|---|---|---|---|
| 2003 | Heat: starting in May, peaking in June and August; Lower than average precipitation from February until June and then again in August | Mean temperature during June and August at least +2 to +4 °C above the long-term average over large areas across Europe; Min temperatures: at least +2 °C of the long-term average in June and August over large CE areas Max temperatures: warmest months June and August, anomalies beyond +4 ° C across almost all CE countries | −50% to −75% of the long-term monthly mean values in June and August across Germany, Poland, Austria, Slovakia, Czechia and Hungary; September precipitation decreasing to −75% in southern Germany and Switzerland | −225 mm; 49% of the grid points across CE below −250 mm | −1.5 σ (standard deviation) | [28,29,44,45,80] |
| 2004 | Relatively favorable climatic conditions compared to 2003 with on average milder temperatures and higher precipitation months | Mean temperatures remaining 3 ° C below the long-term average (1981–2010) in May, June and July and 0 up to +4 °C above the long-term average in April, August and October | Precipitation in March between −20% and −60% in Germany, Austria Switzerland, parts of Poland in April; from −20% to +150% in May, July and August, with the exception of Czechia and Poland: −40% to −80% in August and September, respectively | −38 mm | +0.1 σ | [28,29] |
| 2018 | Impact on a larger area than in 2003; Heat: began in April (mean temperature +4 to +6 °C of long-term average); Lower than average precipitation in February, April, June, July, August, and October; Germany being constantly under stress of below-average precipitation | Mean temperature: +4 to +6 °C in April and May over large CE areas, even above +6 °C in April across large parts of Czechia, small parts of Germany, Poland and Austria, varying between +2 and up to +6 °C from June to September; maximum temperature anomalies +4 σ over larger CE than the long-term average in July (1981-2018; Buras et al. [16]) | In February down to −80% to −100%, in April −40% to −80% lower than long-term average and impacting a large CE region, with June/July anomalies down to -60% to -80% in Austria, Germany and Switzerland; August again −40% to −80% in Germany, Poland and Czechia; also below average in October again in parts of Germany and Hungary; November, also dry down to −80% across almost all of Germany and Poland (smaller regions down to −100%), larger parts of Czechia and parts of Austria, Switzerland and Slovakia | −253 mm; 71% of the grid points across CE below −250 mm | −1.9 σ | [16,17,28,29,30,45] |
| 2019 | Secondary drought after 2018; As a post-drought year more unfavorable than 2004, with a lower climatic water balance during the vegetation period | +2 to +4 °C higher mean temperatures than the long-term average in February, April, July, August, October; June being the warmest with +3 up to +6 °C above average, reaching up to +7 °C across large areas in Poland | Down to −60% to −80% in Germany, Switzerland., Austria, Hungary and Slovakia in February; April ranging from −40% to −80% in Germany, Czechia, parts of Slovakia and Austria; and up to −100 % on large parts of Poland; June and July from −60% to −80% over a large CE region; August down to −60% in eastern Germany and central and western Poland | −132 mm | −0.8 σ | [28,29] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rukh, S.; Sanders, T.G.M.; Krüger, I.; Schad, T.; Bolte, A. Distinct Responses of European Beech (Fagus sylvatica L.) to Drought Intensity and Length—A Review of the Impacts of the 2003 and 2018–2019 Drought Events in Central Europe. Forests 2023, 14, 248. https://doi.org/10.3390/f14020248
Rukh S, Sanders TGM, Krüger I, Schad T, Bolte A. Distinct Responses of European Beech (Fagus sylvatica L.) to Drought Intensity and Length—A Review of the Impacts of the 2003 and 2018–2019 Drought Events in Central Europe. Forests. 2023; 14(2):248. https://doi.org/10.3390/f14020248
Chicago/Turabian StyleRukh, Shah, Tanja G. M. Sanders, Inken Krüger, Tobias Schad, and Andreas Bolte. 2023. "Distinct Responses of European Beech (Fagus sylvatica L.) to Drought Intensity and Length—A Review of the Impacts of the 2003 and 2018–2019 Drought Events in Central Europe" Forests 14, no. 2: 248. https://doi.org/10.3390/f14020248