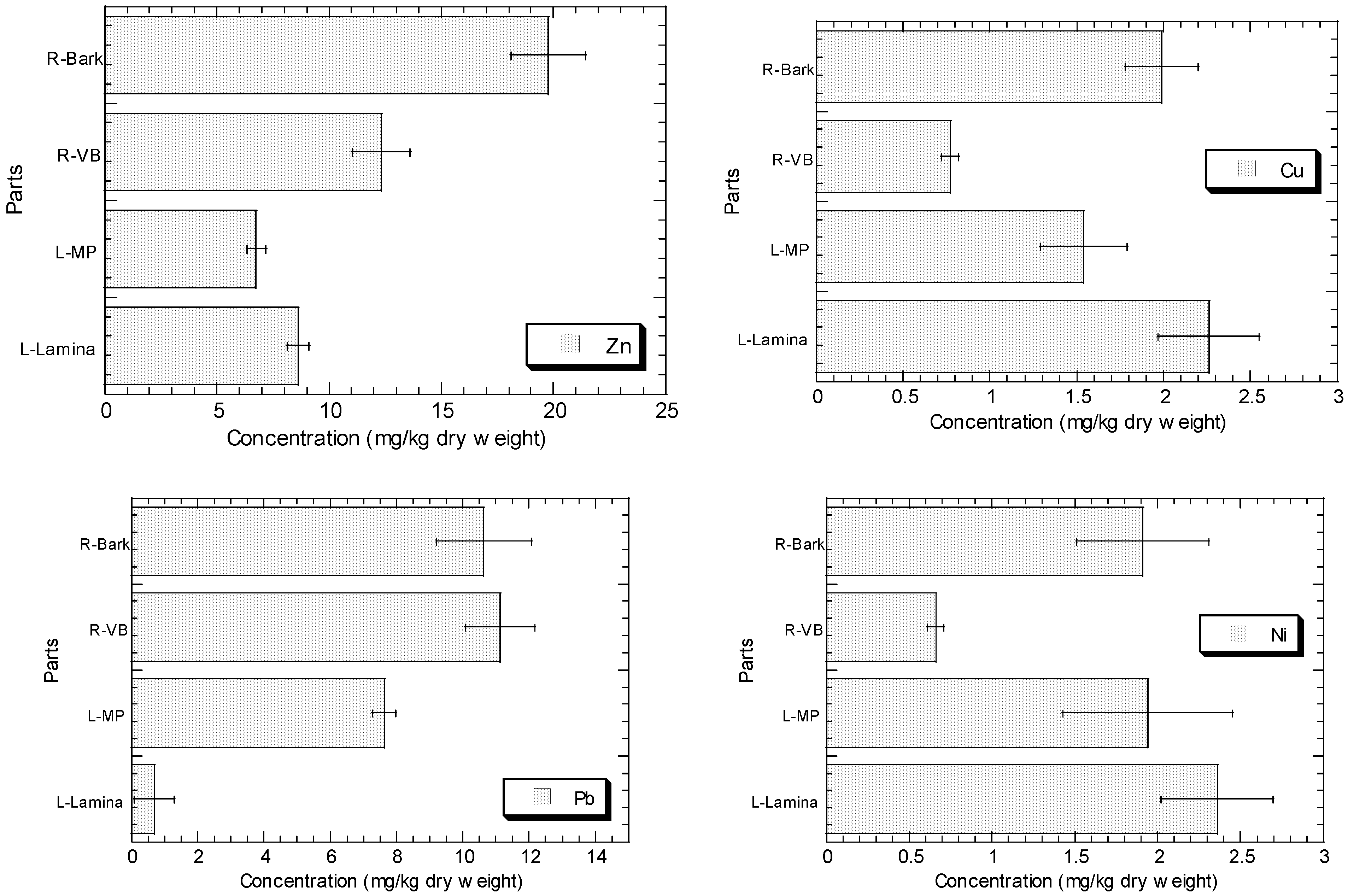
4.1. The Levels of Essential Fe, Cu and Zn in Lamina Are Significantly (p < 0.05) Higher Than in Midrib plus Petiole
The higher number of significant and positive correlations between the essential metal lamina–geochemical fractions than those between Fe MP–geochemical fractions could support the higher levels of Fe, Cu, and Zn in the lamina than MP. This phenomenon was also indicated by the differences in influential variables selected for the accumulation of Fe, Cu, and Zn between lamina and MP.
This demonstrated that the lamina had better potential to accumulate the necessary essential Fe, Cu, and Zn than the MP. The higher levels of essential metals in the lamina could be because these metals are needed for the metabolism during the photosynthetic activities of the lamina, and these metals are required to produce carbohydrates for the plant.
The lamina is in charge of the plant’s photosynthesis. Managing the leaf lamina function by the leaf lamina area is crucial for maintaining the viability of plant life, depending on the ecology. It contains chloroplast, which gives leaves their green colour, so a leaf is referred to as the kitchen of the plant. The petiole, which connects the leaf’s flat surface to the plant, is referred to as the lamina. The main portion of the leaf, or lamina, has a network of stomata that allow carbon dioxide and oxygen to be taken in and exhaled. These stomata, which are connected by primary and secondary veins, open and close in response to the plant’s needs for processing food. The lamina receives its structure and shape from the midrib. Chlorophyll is found in the lamina, which serves as a trap for the energy from sunshine. The mixture of water and carbon dioxide then uses the energy trapped to produce glucose. The plant’s food, glucose, is created and transported to different sections of the plant by the phloem, or vascular tissue, which is also found in the lamina. The primary purpose of green leaves is a process known as photosynthesis [
82].
Essential micronutrients, Cu, Fe, and Zn, and their uptake and allocation to plant organs, such as photosynthetic leaf tissue, are high and active [
51]. The highest value of the essential metals compared to non-essential metals of
R. apiculata is closely related to the role of this element in growth and plant cell maintenance. Due to the significant roles that vital metals play in the organelle mechanisms of the leaf structure, it is possible that the greater amounts of Fe, Cu, and Zn in the lamina are the result. The Fe substance is required to provide them with the chlorophyll that gives them their distinctive green colour and absorbs light energy. Cu and Zn are required to construct a protective leaf epidermis at the lamina levels.
Fe is a necessary transition metal, with a redox-active character, under biological conditions [
83] since it is a crucial micronutrient [
84]. Redox enzymes use several Fe compounds as cofactors. They involve metabolic activities such as DNA synthesis, respiration, and photosynthesis [
84]. Despite being common in most well-aerated soils, Fe has a low biological activity because, at neutral pH levels, it largely forms very insoluble ferric compounds. Iron plays a crucial role in numerous physiological and metabolic processes in plants. It is needed for various biological processes, as it is a component of numerous essential enzymes, including cytochromes of the electron transport chain. Fe is required to maintain chloroplast structure and function in plants, and it is involved in synthesising chlorophyll. However, little is known about how Fe status is regulated in leaves.
Cu is a vital cofactor for many metalloproteins, and it is involved in some physiological processes. However, elevated bioaccumulation of Cu in the lamina could stunt the growth and compromises crucial cellular functions, such as photosynthetic electron transport [
85]. Different mechanisms have developed in plants to properly manage Cu homeostasis as a function of the environmental Cu level because Cu is both a vital cofactor and a harmful element, including a complex network of metal trafficking routes. Plastocyanin (PC) is an abundant and crucial Cu protein needed for photosynthesis in higher plants, according to Shahbaz et al. [
86]. Due to a lack of PC, severe Cu shortage may result in a flaw in photosynthetic electron transport.
Besides playing a crucial part in the growth, Zn also plays a role in plants’ ability to protect themselves, with a focus on superoxide dismutases [
87,
88,
89]. The amount of available Zn and the type of plant impact Zn distribution and transport in plants. Zn concentrations are often higher in developing tissue than in mature tissue when plants have low to adequate Zn supplies [
88]. Accumulation has been seen in plants’ root cortices and leaves that can tolerate harmful Zn levels. Zn was accumulated in the cell walls or stored in vacuoles in the lamina [
89].
4.2. The Levels of Non-Essential Pb and Ni in Midrib plus Petiole Is Significantly (p < 0.05) Higher Than in Lamina
The higher number of significant and positive correlations between Ni MP–geochemical fractions are higher than those between Ni lamina–geochemical fractions, supporting that the non-essential metal in MP is higher than in lamina.
The non-essentiality of Pb and Ni to plant growth, with low solubility and their passive uptake, caused their translocation from roots to other plant organs to be generally low [
51]. The lower mean values of Pb and Ni also indicated that these non-essential elements are not significantly involved in the major process of
R. apiculata. This phenomenon was also indicated by the differences in influential variables selected for accumulating Pb and Ni between lamina and MP.
Although Pb is not necessary for plants, it is easily absorbed and accumulates in many plant tissues [
90]. Particle size, cation exchange capability, pH, root exudation, and other physico-chemical factors affect how much Pb plants can absorb. Mechanisms of Pb detoxification include binding to glutathione and amino acids, sequestering Pb in the vacuole, phytochelatin production, and other processes. Pb tolerance is linked to a plant’s ability to confine Pb to its cell walls, producing osmolytes and activating its antioxidant defence system. Using phytoremediation and rhizofiltration technologies to clean up Pb-contaminated soils appears to have many potential metabolisms, while growth and photosynthetic activity of the lamina were found to be harmed by the high intake of Pb. However, excessive Pb bioaccumulation in plant leaves might result in a 42% reduction in root growth [
91].
Although generally considered a non-essential metal, Ni is a component of urease and hydrogenase. It has been acknowledged as a necessary element for plants and was the most recent element to be added to the list of essential nutrients for plants [
92]. According to van der Ent et al. [
93], particular metal sequestering and transport processes cause a high level of Ni accumulation in hyperaccumulator plants. Understanding these processes is essential for understanding how these plants regulate the metabolism of transition elements. The chemical speciation of Ni
2+ is unmistakably linked to citrate and does not significantly vary between species, plant tissues, or transport fluids.
4.3. The Lamina Has a Better Potential as a Phytoremediator of Cu, Zn and Fe, while Midrib plus Petiole Has a Better Potential as a Phytoremediator of Pb and Ni
The overall R values between the essential metal (Fe, Cu, and Zn) lamina–BCF pairwises, which are higher than those between MP–BCF pairwises, could support that lamina is a better phytoremediator of Cu, Zn, and Fe than MP. The relatively similar numbers of influential variables selected for the accumulation of Zn lamina (8 variables) compared to Zn MP (9 variables), as well as similar numbers of influential variables for the accumulation of Cu lamina (9 variables) and Cu MP (9 variables), could be used to understand why lamina could be a good phytoremediator for Zn and Cu for lamina.
The overall R values between Ni MP–BCF pairwises are higher than those between Ni lamina–BCF pairwises, supporting that MP has a better potential as a phytoremediator of Ni. The clear higher numbers of influential variables selected for the accumulation of Pb MP (10 variables) compared to Pb lamina (4 variables), as well as for the accumulation of Ni MP (10 variables) compared to Ni lamina (5 variables), indicated one point. The more variables with higher R values for Pb and Ni in MP supported the correlation pairwises mentioned previously. Hence, this could be a proxy of MP, which is a better phytoremediator for Pb and Ni than lamina.
According to Kamaruzzaman et al. [
52], BCF values showed that
R. apiculata contains substantially less lead than silt, at just 0.96 times the levels. It is believed that the mechanism behind the increased accumulation of metals in plant tissues is the translocation of air absorbed by lenticels in pneumatophores to subterranean roots. In the anaerobic soil environment, this leads to the production of oxidised rhizospheres, a decline in complexing sulphides, a reduction in the stability of Fe plaques, and an increase in the concentration of exchangeable trace metals [
94].
MacFarlane and Burchett [
95] reported that
A. marina revealed that Pb showed little absorption and minimal movement, while Cu and Zn had the largest accumulation. Sazon and Veronica [
18] reported that Zn (0.7) in
Avicennia sp. had the highest BCF values, whereas metals with high mean levels were in sediments (4244 mg/kg Ni and 137,049 mg/kg Fe) had lower BCF values. They concluded that
R. apiculata could be useful in the remediation of Ni-Fe-laden sediments, particularly for phytostabilization, based on the limited translocation of Ni and Fe in these plants.
From the present study, all values of BCF
OO, BCF
RES, and BCF
SUM were found below 1.0, indicating a low PTM uptake. The BCF
SUM value is conventionally used, which is based on total metal concentration. However, the present BCF values were higher than 1.0 for BCF
EFLE and BCF
AR, indicating the substantial update of PTM at the EFLE and AR fractions of the mangrove habitat sediments. Even though BCF calculates species’ potential for phytoremediation [
1], the effectiveness that was based on the total metal concentration of the sediments is debatable. This debate is somewhat masked by the many reported studies concluding that hyperaccumulators are interpreted by BCF > 1, which are also tolerant to high concentrations of PTMs [
96].
Usman et al. [
3] claimed that the BCF, which measures the ratio of metal concentration in tissue compared to that in sediment, can be used to quantify how well plants and aquatic creatures can absorb contaminants from sediments. Qiu et al. [
1] reported that the leaf BCF values for Cu, Ni, and Zn are, respectively, 1.25–9.08, 0–1.02, and 0.20–4.71. They found that most of these BCF values were deemed excessive (>1), indicating that
A. marina may be a highly effective plant for bioaccumulating PTMs.
Low bioavailability of PTMs in sediments and/or preventing metal uptake by mangroves can account for the lowest BCF values found in highly metal-contaminated sediments. It was speculated that, by complexing with organic matter and/or precipitating sulphides under decreasing conditions, mangrove sediments could immobilise PTMs in inaccessible forms [
94].
Due to greater metal concentrations in the leaves (284–496 mg/kg) and BCF values >1, Usman and Mohamed [
97] concluded that
A. marina might be categorised as a possible accumulator for Cu (1.25–9.08). However, they cannot be regarded as prospective hyperaccumulators since they lack the necessary quantities of Cu in the above-ground tissues, which should be >1000 mg/kg [
69]. In
R. apiculata from Setiu Mangrove Forest, Terengganu, Malaysia, Kamaruzzaman et al. [
52] reported Cu concentrations of 2.73 and 9.42 mg/kg dry weight in the leaves and sediments, respectively, and Zn concentrations of 1.43 and 11.7 mg/kg dry weight in the leaves and sediments, respectively. According to Khan et al. [
54], essential Fe and Cu contents in tissue samples from
R. apiculata were higher at the 80-year-old site than at the 15-year-old location. Due to the 80-year-old compartment’s proximity to an urban area, this may be a side effect.
Interestingly, mangroves thriving in less contaminated environments had the highest BCF values [
98]. In contrast, plants from the most contaminated area had the highest translocation factors, showing that
Avicennia schaueriana could adapt to unfavourable circumstances [
97]. Nath et al. [
25] concluded that
A. marina could be used as phytostabilizers in this heavily altered estuary, shielding the aquatic ecosystem from direct or indirect sources of PTM contamination. Usman et al. [
3] also investigated the capacity of mangroves (
A. marina) from Farasan Island, the coast of the Red Sea, to store and translocate PTMs inside their numerous compartments. Based on BCF values, their most often obtained values were found to be too high (>1), indicating that
A. marina can be regarded as a highly effective plant for the bioaccumulation of PTMs.
4.4. Lamina Is a Potential Biomonitoring Agent of Potentially Toxic Metals
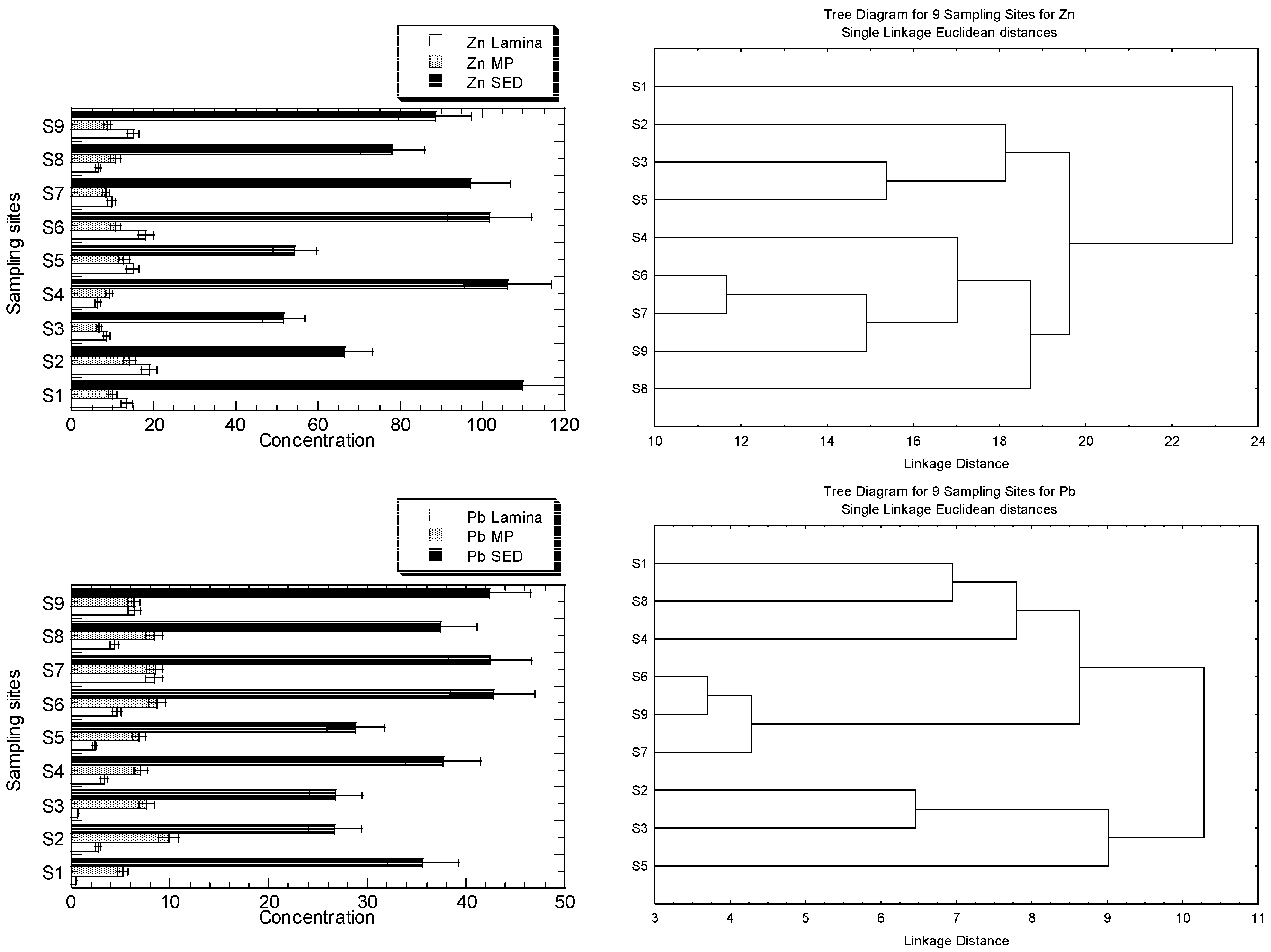
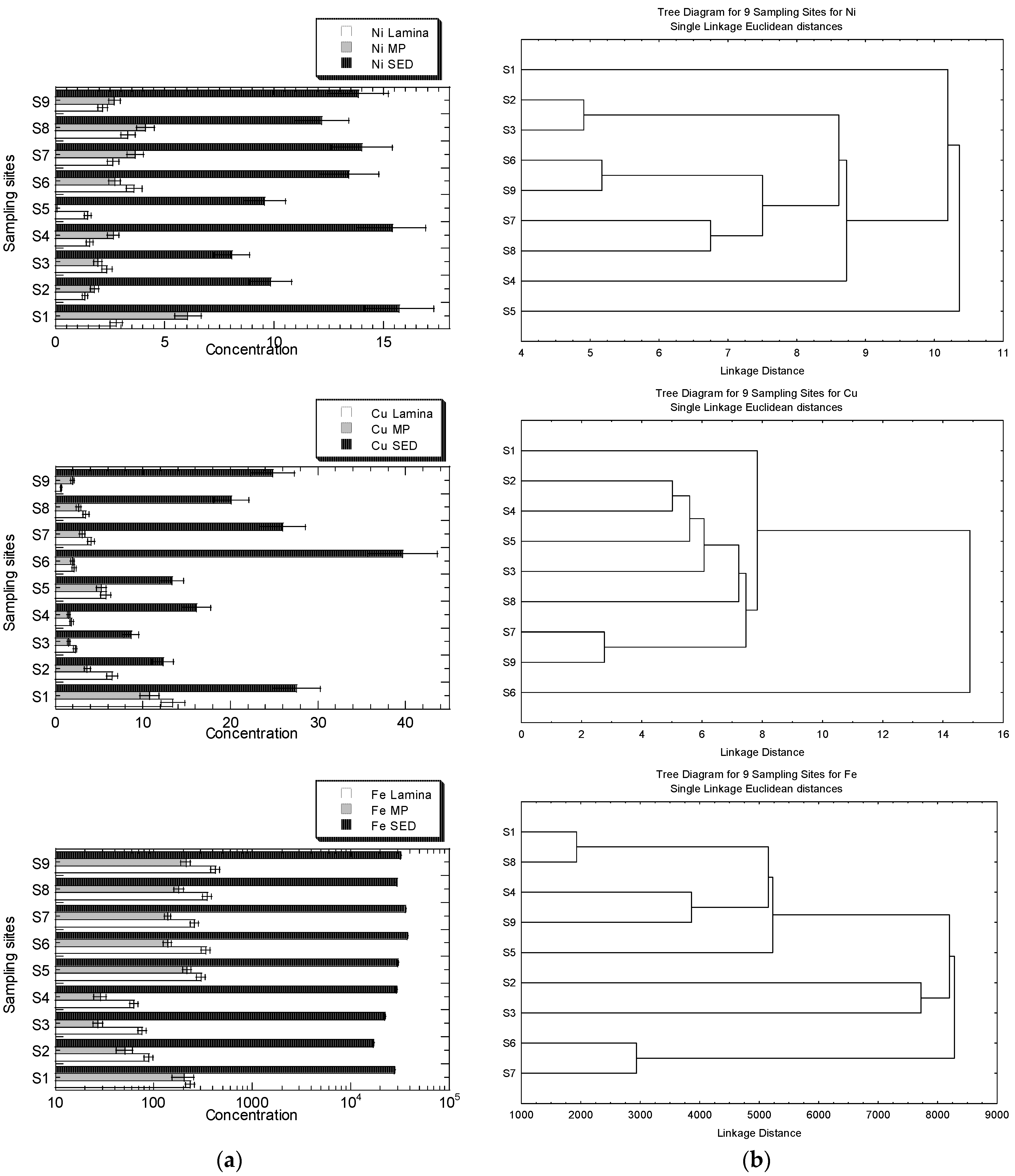
The MP generally exhibits negative connections with all geochemical fractions, but the Zn lamina exhibits positive relationships with all geochemical fractions. Except for EFLE, all geochemical fractions and total summation fractions indicate positive associations between Fe levels in the two leaf portions (lamina and MP), with lamina exhibiting higher R values for SUM, AR, OO, and RES. The Pb lamina exhibits favourable correlations with each geochemical fraction (except for AR). The MP, however, exhibits adverse associations with each geochemical fraction (except for EFLE, AR and OO). Except for AR, all geochemical fractions had poor associations with the Cu levels in the lamina and MP of leaves. Except for EFLE, the Ni concentrations in both the lamina and the MP of leaves demonstrate positive associations with all geochemical fractions and total summation fractions, with the MP displaying higher R values for SUM, OO, and RES.
Some authors have reported the positive correlation of metals between mangrove leaves and their habitat sediments. Except for Cu and Cd, which don’t correlate with other metal concentrations, Aljahdali and Alhassan [
19] found a substantial link between the PTM concentrations in sediment and
A. marina leaves. Bakshi et al. [
24] found that the substantial association between the metal concentration in
A. officinalis leaves collected from Sundarban mangrove wetland and the sediment metals suggested that there has been extensive bioaccumulation. According to Usman et al. [
3], the highest levels of Cu in the leaves of
A. marina are correlated with the highest Cu levels in the nearby sediments.
According to Kamaruzzaman et al. [
52], Pb and Cu are observed to have greater concentrations in the sediments in Setiu mangrove habitats. Their concentrations are on the rise, especially close to the estuary, which is consistent with their predominately lithogenous origin. Furthermore, compared to the coarse-grained sediments of the inner portion of the rivers, the fine-grained sediments in the mangrove and the surrounding area are characterised by higher quantities of these trace elements. The contents of Pb and Cu are positively correlated with grain size, as documented by several scientists [
99,
100], indicating the role of the fine fraction in their assimilation into the sediments. Lower Pb and Cu ratios and larger mean particle sizes indicate sediments located further from the estuary. Cu content was higher than Pb, according to Yunasfi et al. [
39], who used
R. apiculata samples from Belawan (North Sumatra).
Ganeshkumar et al. [
49] suggested that
R. mucronata could be used as a biomonitor for monitoring Pb pollution in the Muthupet mangrove ecosystem, based on the high accumulation and translocation rate of Pb. Based on the values of the biota–sediment accumulation factors (BSAF) of metals in mangrove tissues, Qiu and Qiu [
48] concluded that
Rhizophora stylosa from Dongzhai Harbor and Sanya Bay of Hainan Island can be used as a biomonitor of Cu and Zn pollution with temporal monitoring. Being a good biomonitor of metal pollutants is attributed to the mangrove’s characteristics, including tolerance to metal pollution in the mangrove ecosystem. For instance, Shams et al. [
42] confirmed that the antioxidative defence system plays a critical role in
R. mucronata to tolerate the multiple PTMs stress. Nualla-ong et al. [
41] also suggested that the concentration of glutathione and phytochelatin synthase play an important role in PTM tolerance in
R. mucronata seedlings. Based on the study of the efficiency of
R. apiculata mangrove plants in Zn metal removal made from Zn artificial solution using a laboratory scale reactor, Razif and Farhan [
40] reported the positive efficiency of Zn removal of
R. apiculata.
4.5. Low Metal Uptake in the Mangrove Leaves
According to data from
Avicennia marina, gathered from eight stations at the Rabigh lagoon (Saudi Arabia), Aljahdali and Alhassan [
19] found that there was a positive connection between antioxidants and Pb, which may be due to
A. marina’s capacity to exclude or detoxify this metal through its mechanism of exclusion or detoxification. Therefore, low levels of PTM bioaccumulation in the mangrove leaves are expected.
According to Arumugam et al. [
16], samples of
A. marina have higher concentrations of non-essential metals such as Cd and Pb, in which the metal accumulation was influenced by the seasons, with the summer and the post-monsoon being the most polluted. When compared to non-essential metals, they found that essential elements such as Cu and Zn were more abundant in the sediment of the rhizosphere of
A. marina. As a result, the plant was using Cu and Zn for their active metabolism quickly. This phenomenon could explain the low bioaccumulation of Cu and Zn in the leaves of
R. apiculata from the present study.
Using both species-level analyses and a phylogenetic approach, MacFarlane et al. [
22] conducted a comparative analysis analysing patterns of accumulation and partitioning of Cu, Pb, and Zn in mangroves from accessible field-based studies to date.
Avicennia mangroves have several adaptive mechanisms for overcoming the difficulty of saline and extremely anoxic conditions. Although they are different species of mangrove, metal accumulation and partitioning for Cu, Pb, and Zn were discovered to be comparable across
Avicennia genera and vast geographical ecosystems. With root BCF values of 1, metals were collected in roots to concentrations similar to those of nearby sediments. For all metals, the root BCFs were consistent during the exposure range. The metal levels in leaves were either half or less than those in roots. Cu and Zn, two important elements, saw a decline in leaf BCF when environmental concentrations rose. Regardless of ambient quantities, the non-essential metal Pb was not found in leaf tissue. As a result, mangroves, as a whole, often act as species that exclude non-essential metals and regulate vital metals.
Greger [
101] hypothesised that a high degree of soil salinity, which leads to the development of metal-chloride complexes, may be to blame for plants’ inadequate absorption of heavy metals. The PTMs are less readily absorbed and accumulated by plants due to these complexes. Kamaruzzaman et al. [
52] reported, in contrast to the current study, that the concentration of Cu was highest in sediments, followed by roots, barks, and leaves. Roots had larger quantities of Fe and Cu than shoots, indicating effective absorption and accumulation that may be related to improved oxidation in the rhizosphere [
102].
Collected from the Alinsaog River (Philippines), Sazon and Veronica [
18] reported the concentrations (mg/kg dry weight) of Zn, Ni, and Fe in leaves of
R. apiculata, and the sediments surrounding it were 13.9, 95.1, 351.2, and 137,049 mg/kg dry weight, respectively. According to Ariyanto et al. [
21], the mean concentrations of Pb in the leaves and habitat sediments of
R. apiculata mangrove were 0.00035 mg/kg dry weight and sediment (9.06·0.05 mg/kg), respectively, in Asahan East Coast, Asahan Regency, North Sumatra.
Additionally, Usman et al. [
3] demonstrated that the metal contents of
A. marina’s leaf, branch, and root varied. The leaves of
A. marina were the component of the plant that accumulated the greatest Cd and Cu in the majority of the research sites. However, there is no discernible pattern among the examined sites for Zn and Ni. According to Usman et al. [
3], the metal levels in the mangrove tissues were high compared to mangrove concentrations worldwide for most of the studied metals [
1,
94,
95,
103]. Qiu et al. [
1] reported that nine species of mangroves from Hainan Island, China have average amounts of Cu, Zn, Cd, and Pb of 2.8, 8.7, 0.03, and 1.1 mg/kg dry weight, respectively. Defew et al. [
4] measured leaf concentrations of Cu and Zn at 3.73 and 35.8 mg/kg dry weight, respectively, in a study on the assessment of metal contamination in mangrove sediments and leaves from Punta Mala Bay, Pacific Panama.
Based on
R. apiculata collected from Balok mangrove forest, Pahang, Kamaruzzaman et al. [
53] reported that the mean Pb concentrations (mg/kg dry weight) in the leaf and root of
R. apiculata and their habitat surface sediment were 4.30, 22.5, and 31.2, respectively. The average Cu concentrations recorded were 2.93, 4.81, and 15.5, respectively. Based on
R. apiculata from the Setiu mangrove forest (Terengganu), Kamaruzzaman et al. [
52] also reported the mean concentrations (mg/kg dry weight) of Cu in the mangrove’s leaf, root, and their habitat sediment as: 2.73, 5.21, and 9.42, respectively. Meanwhile, the mean concentrations (mg/kg dry weight) of Pb in leaf, root, and sediment were 1.43, 2.05, and 11.66, respectively. Abdullah et al. [
51] reported ratios of 48.5:9.23 between roots and leaves for Fe, 3.60:1.14 between roots and leaves for Zn, and 0.22:0.08 between roots and leaves for Pb in
R. apiculata collected from Kuala Sepetang. Based on
R. stylosa collected from Belawan Stream Estuary (Medan, Northern Sumatra), Yunasfi et al. [
39] reported that the levels of Cu and Pb were higher in the roots and stems than in the leaves of the mangrove. Based on
R. mucronata in the Kelantan Delta, Baruddin et al. [
47] found that the roots accumulated higher levels of Pb than the leaves.
The above three findings confirmed that the levels of Cu, Pb, and Zn were lowest in leaves of
R. apiculata, followed by roots and habitat sediments. This also indicated that metal accumulation generally occurs at the root levels of
R. apiculata, with restricted transport to leafy or aerial portions of the mangrove plants. Based on
Rhizophora spp. collected from Tanjung Piai, Halim et al. [
40] concluded that the
Rhizophora did not meet the criteria for hyperaccumulators due to low metal accumulation, necessitating further research to identify species suited for phytoextraction of Cd, Pb, and Zn. Pahalawattaarachchi et al. [
44] concluded that the trend of the phytostabilization capacity of
R. mucronata in the Alibag mangrove ecosystem is confined to only the highly abundant metals in nature, and the phytoremediation capacity of
R. mucronata varies from metal to metal.
These indicated that plants actively avoid the uptake of PTMs. Qiu and Qiu [
48] proposed that total organic carbon in the mangrove sediment could be one of the important factors for regulating metals in mangrove tissues. Hence, the present findings indicated that the leaves of
R. apiculata played important roles as biofilters and natural pollution treatment centres because the root system could manage to trap the PTMs into the sediments, as well as particulates, which are transported by the current into the oceans from the estuaries [
52,
53,
99,
100].
The low level of metal uptake by
R. apiculata from the present study suggested that
A. apiculata from Sepang and Lukut may not be stressed due to the unpolluted condition of the study sites. According to Kabata–Pendias and Pendias [
104], the gene plants’ general heavy metal concentration depends on the amounts of metals that were measured in the plant tissues. At Yanbu Red Sea, Saudi Arabia, Alharbi et al. [
20] evaluated the geographical distribution and concentrations (mg/kg dry weight) of Cu, Ni, Pb, and Zn in the mangrove
Avicennia marina) leaves (16.1–56.3, 18.1–40.2, 2.3–9.90, and 36.8–84.9, respectively) and their habitat sediments (27.3–242, 17.2–217, 11.5–111, and 48.8–512, respectively). These results showed that the mangrove leaves accumulated far lower levels of PTMs than their habitat sediments.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}