


Different Environmental and Phylogenetic Controls over the Altitudinal Variation in Leaf N and P Resorption Traits between Woody and Herbaceous Plants
 ,
,
Abstract
:1. Instruction
2. Methods
2.1. Study Site
2.2. Field Sampling and Chemical Analyses
2.3. Calculations of Leaf Nutrient Resorption
2.4. Species Phylogeny
2.5. Climatic Data
2.6. Data Analysis
3. Results
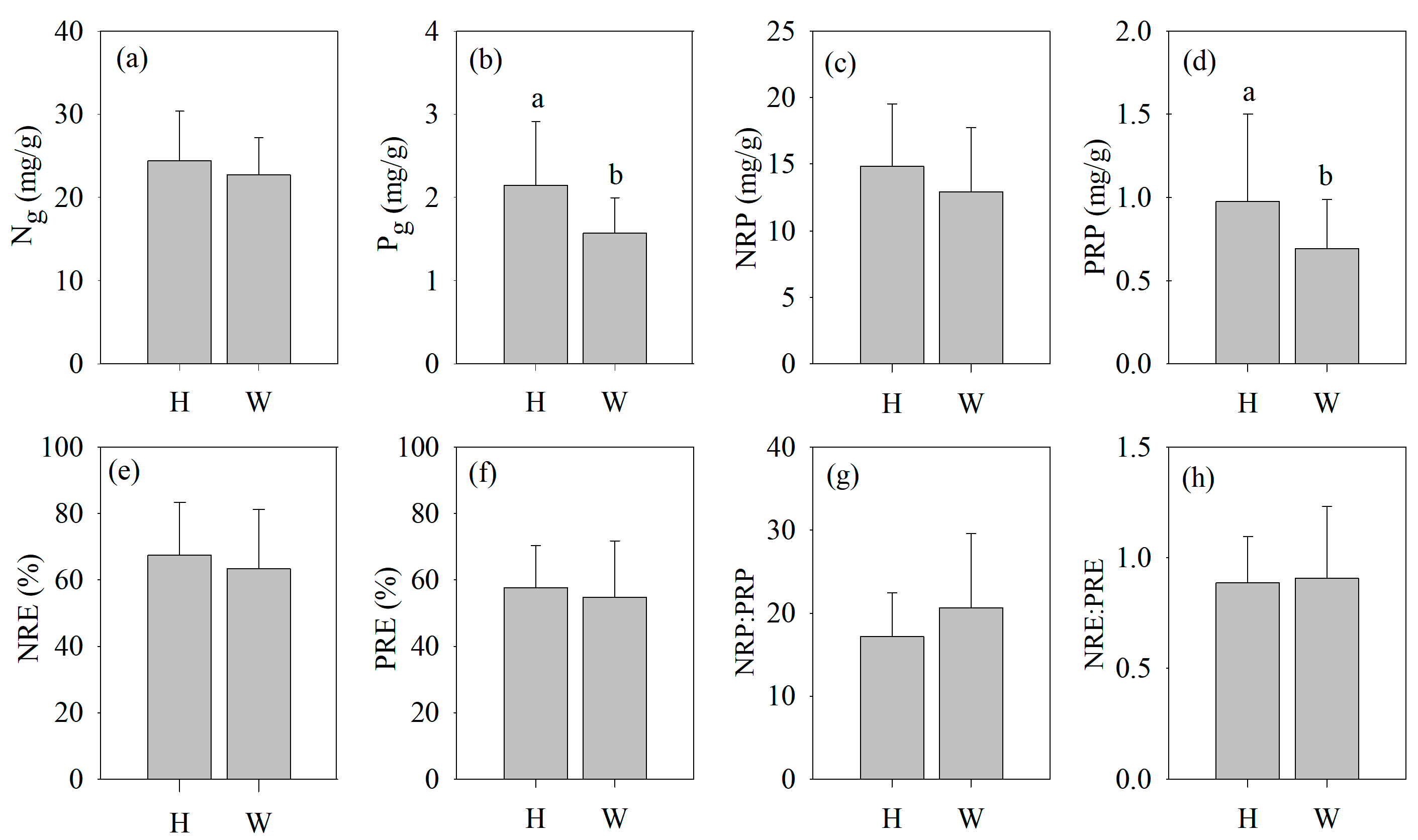
3.1. Variation in Leaf N and P Nutrient Traits and Resorption Traits
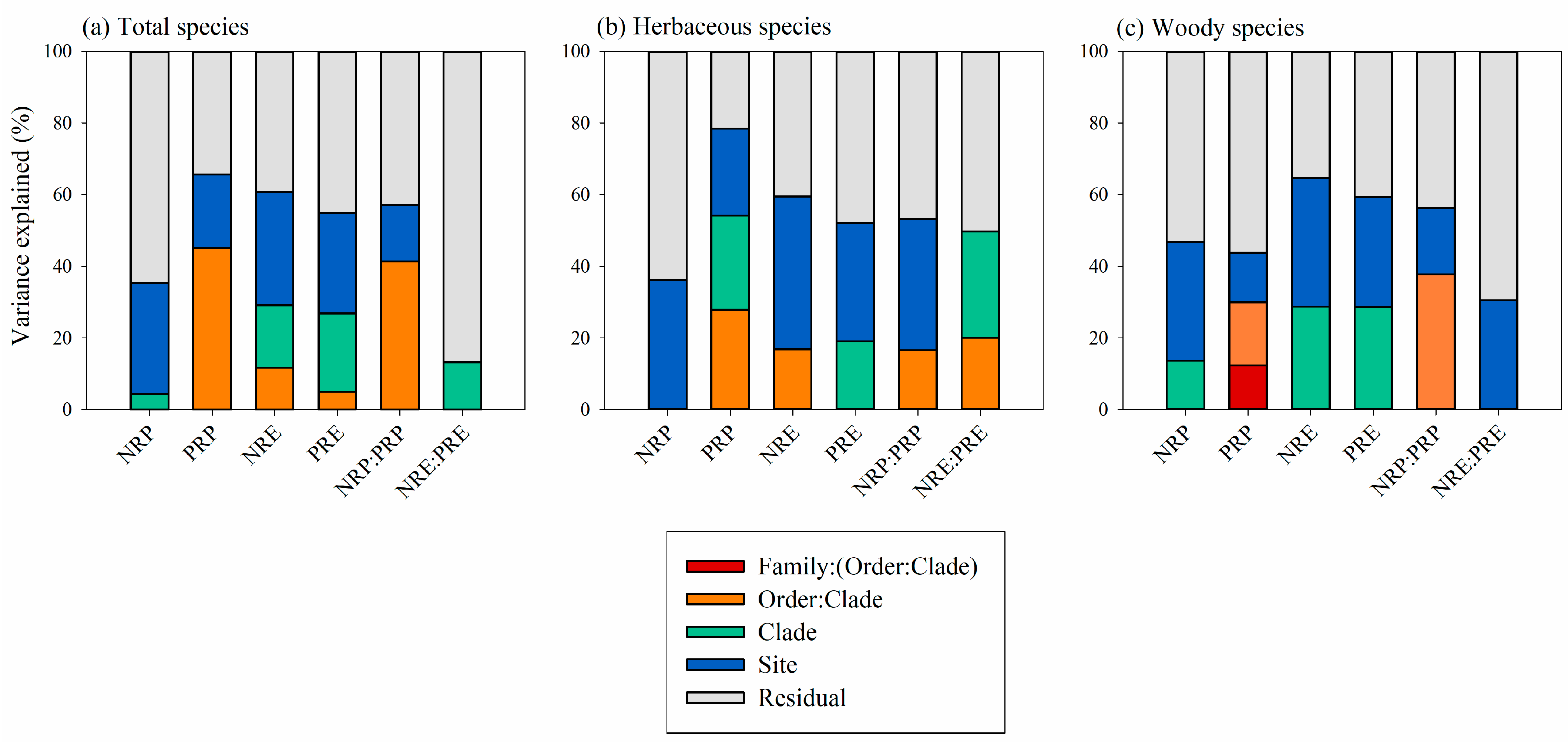
3.2. Phylogenetic Effects on the Variation in Leaf Resorption Traits
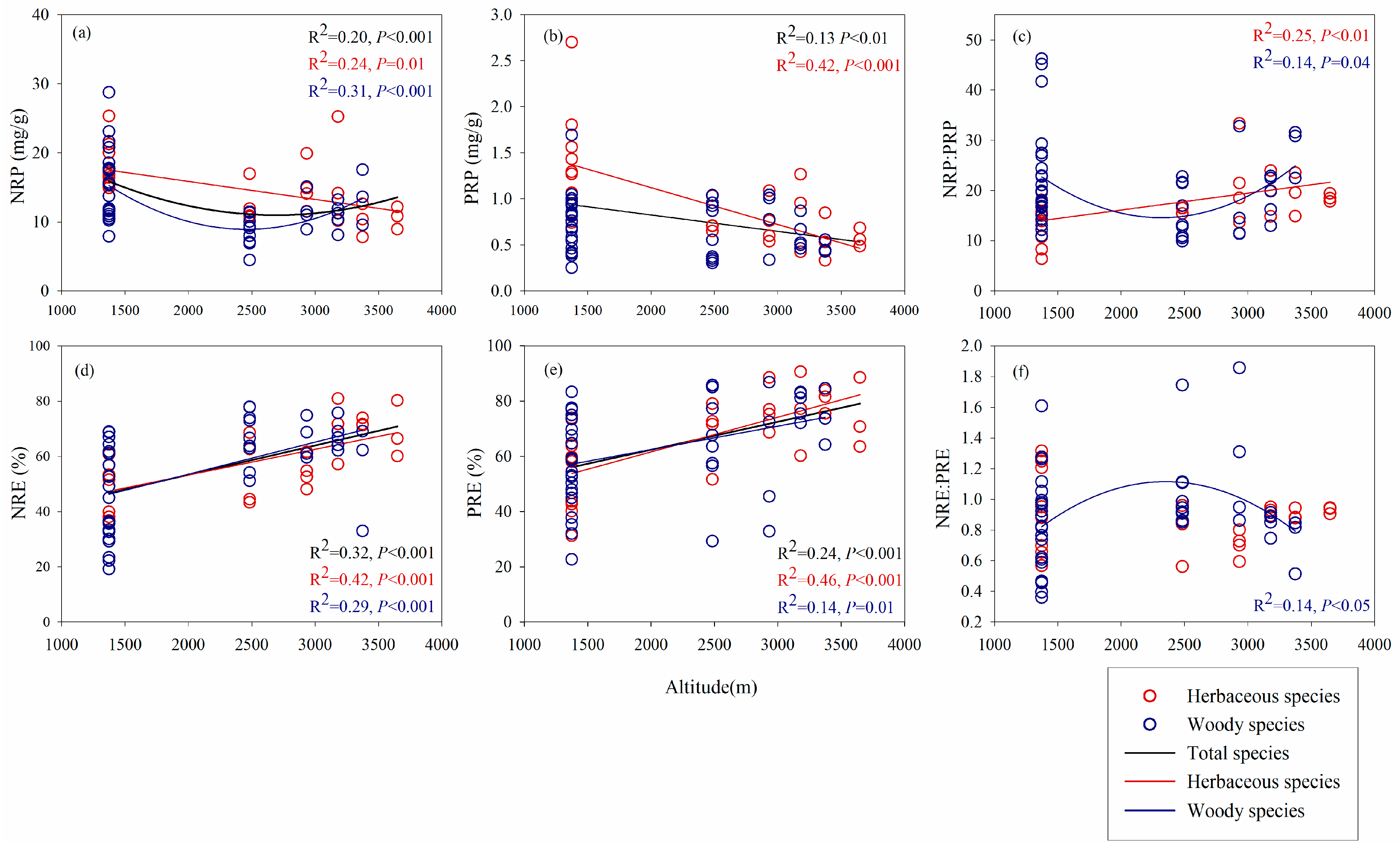
3.3. Altitudinal Patterns of Leaf N and P Resorption Traits
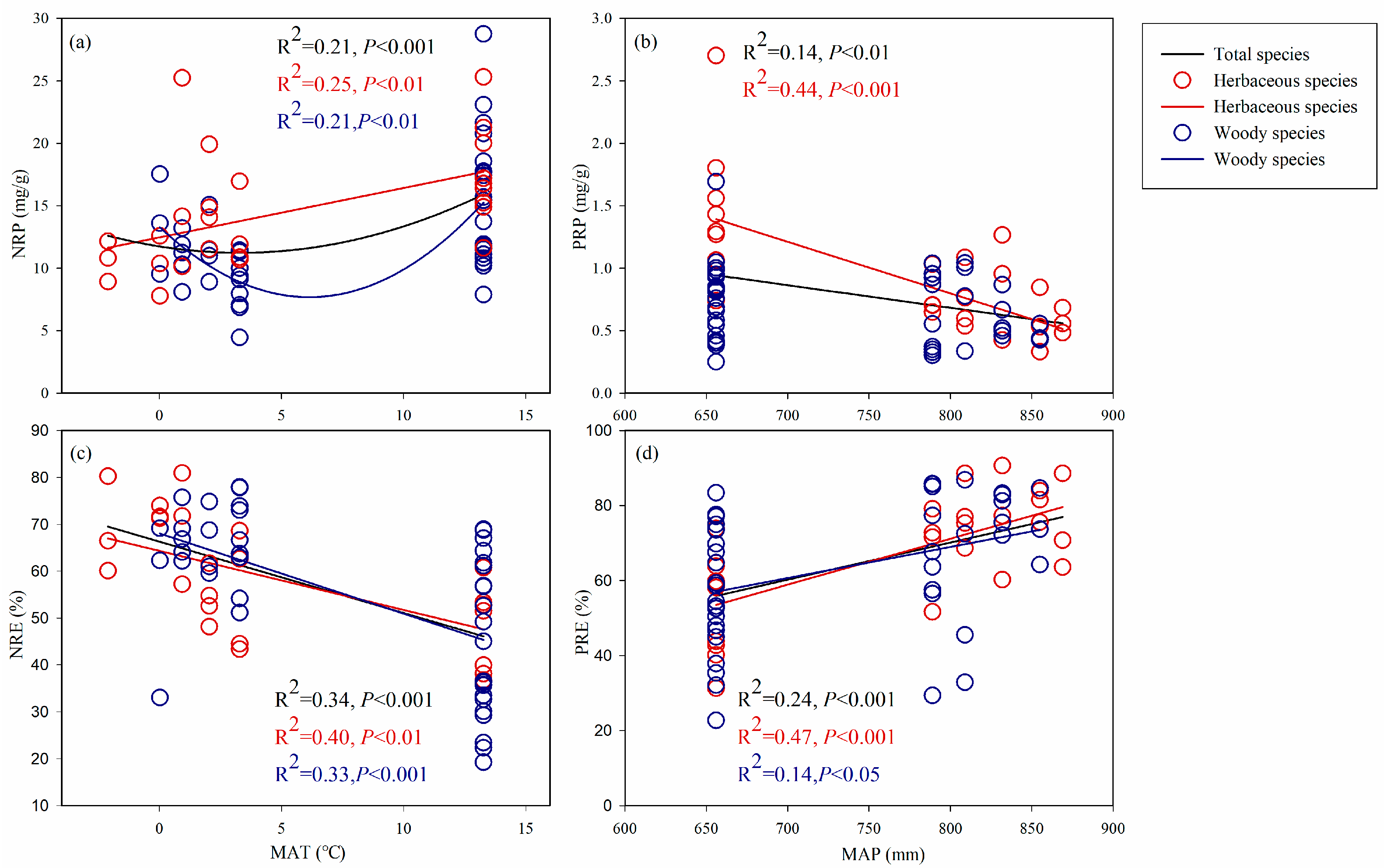
3.4. Environmental Effects on the Variation in Leaf Resorption Traits
4. Discussion
4.1. Phylogenetic Influence on Leaf Resorption Traits
4.2. Environmental Influence on the Variation in Leaf Resorption Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| NuRP | nutrient resorption proficiency |
| NuRE | nutrient resorption efficiency |
| NRP | nitrogen resorption proficiency |
| PRP | phosphorus resorption proficiency |
| NRE | nitrogen resorption efficiency |
| PRE | phosphorus resorption efficiency |
References
- Brant, A.N.; Chen, H.Y.H. Patterns and mechanisms of nutrient resorption in plants. Crit. Rev. Plant Sci. 2015, 34, 471–486. [Google Scholar] [CrossRef]
- Estiarte, M.; Penuelas, J. Alteration of the phenology of leaf senescence and fall in winter deciduous species by climate change: Effects on nutrient proficiency. Glob. Chang. Biol. 2015, 21, 1005–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.Y.; Chen, H.Y.H. Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Glob. Ecol. Biogeogr. 2009, 18, 11–18. [Google Scholar] [CrossRef]
- Yuan, Z.Y.Y.; Chen, H.Y.H. Global trends in senesced-leaf nitrogen and phosphorus. Glob. Ecol. Biogeogr. 2009, 18, 532–542. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Negative effects of fertilization on plant nutrient resorption. Ecology 2015, 96, 373–380. [Google Scholar] [CrossRef]
- Killingbeck, K.T. Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiency. Ecology 1996, 77, 1716–1727. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar]
- Du, E.Z.; Terrer, C.; Pellegrini, A.F.A.; Ahlstrom, A.; van Lissa, C.J.; Zhao, X.; Xia, N.; Wu, X.H.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Tang, L.Y.; Han, W.X.; Chen, Y.H.; Fang, J.Y. Resorption proficiency and efficiency of leaf nutrients in woody plants in eastern China. J. Plant Ecol. 2013, 6, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Vergutz, L.; Manzoni, S.; Porporato, A.; Novais, R.F.; Jackson, R.B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.; Zhu, J.J.; Yang, K. Leaf nitrogen and phosphorus resorption of woody species in response to climatic conditions and soil nutrients: A meta-analysis. J. For. Res. 2018, 29, 905–913. [Google Scholar] [CrossRef]
- Sun, X.; Kang, H.; Chen, H.Y.H.; Bjorn, B.; Samuel, B.F.; Liu, C. Biogeographic patterns of nutrient resorption from Quercus variabilis Blume leaves across China. Plant Biol. 2016, 18, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.S.; Shi, P.L.; Wu, J.S.; Xiong, D.P.; Zong, N.; Zhang, X.Z. Foliar nutrient resorption patterns of four functional plants along a precipitation gradient on the Tibetan Changtang Plateau. Ecol. Evol. 2017, 7, 7201–7212. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.P.; Zhu, Y.F.; Zhang, S.H.; Feng, Y.Z.; Zhang, W.; Han, X.H. Global scaling the leaf nitrogen and phosphorus resorption of woody species: Revisiting some commonly held views. Sci. Total Environ. 2021, 788, 147807. [Google Scholar] [CrossRef]
- Hao, Z.; Kuang, Y.W.; Kang, M. Untangling the influence of phylogeny, soil and climate on leaf element concentrations in a biodiversity hotspot. Funct. Ecol. 2015, 29, 165–176. [Google Scholar] [CrossRef]
- Zhao, N.; Yu, G.R.; He, N.P.; Wang, Q.F.; Guo, D.L.; Zhang, X.Y.; Wang, R.L.; Xu, Z.W.; Jiao, C.C.; Li, N.N.; et al. Coordinated pattern of multi-element variability in leaves and roots across Chinese forest biomes. Glob. Ecol. Biogeogr. 2016, 25, 359–367. [Google Scholar] [CrossRef]
- Lang, S.I.; Aerts, R.; van Logtestijn, R.S.P.; Schweikert, W.; Klahn, T.; Quested, H.M.; van Hal, J.R.; Cornelissen, J.H.C. Mapping nutrient resorption efficiencies of subarctic cryptogams and seed plants onto the Tree of Life. Ecol. Evol. 2014, 4, 2217–2227. [Google Scholar] [CrossRef]
- Zhang, J.H.; Tang, Z.Y.; Luo, Y.K.; Chi, X.L.; Chen, Y.H.; Fang, J.Y.; Shen, H.H. Resorption efficiency of leaf nutrients in woody plants on Mt. Dongling of Beijing, North China. J. Plant Ecol. 2015, 8, 530–538. [Google Scholar] [CrossRef] [Green Version]
- Kobe, R.K.; Lepczyk, C.A.; Iyer, M. Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology 2005, 86, 2780–2792. [Google Scholar] [CrossRef] [Green Version]
- Han, W.X.; Tang, L.Y.; Chen, Y.H.; Fang, J.Y. Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants. PLoS ONE 2013, 8, e83366. [Google Scholar] [CrossRef]
- Prieto, I.; Querejeta, J.I. Simulated climate change decreases nutrient resorption from senescing leaves. Glob. Chang. Biol. 2020, 26, 1795–1807. [Google Scholar] [CrossRef] [PubMed]
- He, M.S.; Yan, Z.B.; Cui, X.Q.; Gong, Y.M.; Li, K.H.; Han, W.X. Scaling the leaf nutrient resorption efficiency: Nitrogen vs phosphorus in global plants. Sci. Total Environ. 2020, 729, 138920. [Google Scholar] [CrossRef] [PubMed]
- Freschet, G.T.; Cornelissen, J.H.; van Logtestijn, R.S.; Aerts, R. Substantial nutrient resorption from leaves, stems and roots in a subarctic flora: What is the link with other resource economics traits? New Phytol. 2010, 186, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Wang, R.; Chen, H.; Liu, X.; Wang, Z.; Wen, J.; Zhang, S. Plant phylogeny and growth form as drivers of the altitudinal variation in woody leaf vein traits. Front. Plant Sci. 2020, 10, 1735. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Fang, J. Temperature variation along the northern and southern slopes of Mt. Taibai, China. Agric. For. Meteorol. 2006, 139, 200–207. [Google Scholar] [CrossRef]
- Du, B.M.; Ji, H.W.; Peng, C.; Liu, X.J.; Liu, C.J. Altitudinal patterns of leaf stoichiometry and nutrient resorption in Quercus variabilis in the Baotianman Mountains, China. Plant Soil 2017, 413, 193–202. [Google Scholar] [CrossRef]
- Gerdol, R.; Iacumin, P.; Brancaleoni, L. Differential effects of soil chemistry on the foliar resorption of nitrogen and phosphorus across altitudinal gradients. Funct. Ecol. 2019, 33, 1351–1361. [Google Scholar] [CrossRef]
- Chase, M.W.; Byng, J.; Christenhusz, M.; Fay, M.; Judd, W.; Mabberley, D.; Sennikov, A.; Soltis, D.; Soltis, P.S.; Stevens, P.F.J. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar]
- Zanne, A.E.; Tank, D.C.; Cornwell, W.K.; Eastman, J.M.; Smith, S.A.; FitzJohn, R.G.; McGlinn, D.J.; O’Meara, B.C.; Moles, A.T.; Reich, P.B. Three keys to the radiation of angiosperms into freezing environments. Nature 2014, 506, 89–92. [Google Scholar] [CrossRef]
- Konietschke, F.; Placzek, M.; Schaarschmidt, F.; Hothorn, L.A. nparcomp: An R software package for nonparametric multiple comparisons and simultaneous confidence intervals. J. Stat. Softw. 2015, 64, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Blomberg, S.P.; Garland, T.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar] [PubMed]
- Watanabe, T.; Broadley, M.R.; Jansen, S.; White, P.J.; Takada, J.; Satake, K.; Takamatsu, T.; Tuah, S.J.; Osaki, M. Evolutionary control of leaf element composition in plants. New Phytol. 2007, 174, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.C.D. Extension of Nakagawa & Schielzeth’s R-GLMM(2) to random slopes models. Methods Ecol. Evol. 2014, 5, 944–946. [Google Scholar] [PubMed] [Green Version]
- Wang, X.C.; Wang, Q.; Chen, Y.; Zhao, R.; Zhang, J.H.; Quan, X.K.; Liu, F.; Wang, C.K. Coloration and phenology manifest nutrient variability in senesced leaves of 46 temperate deciduous woody species. J. Plant Ecol. 2022, 15, 700–710. [Google Scholar] [CrossRef]
- Hoch, W.A.; Singsaas, E.L.; McCown, B.H. Resorption protection. Anthocyanins facilitate nutrient recovery in autumn by shielding leaves from potentially damaging light levels. Plant Physiol. 2003, 133, 1296–1305. [Google Scholar] [CrossRef] [Green Version]
- Hörtensteiner, S. Chlorophyll degradation during senescence. Annu. Rev. Plant Biol. 2006, 57, 55–77. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Press Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Veneklaas, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.R.; Shane, M.W.; White, P.J.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef]
- Li, C.H.; Yu, H.L.; Xu, Y.X.; Zhu, W.W.; Wang, P.; Huang, J.Y. Close linkages between leaf functional traits and soil and leaf C:N:P stoichiometry under altered precipitation in a desert steppe in northwestern China. Plant Ecol. 2022, 223, 407–421. [Google Scholar] [CrossRef]
- Nearing, M.A. The mechanics of soil detachment by raindrops and runoff. Eurasian Soil Sci. 1997, 30, 552–556. [Google Scholar]
- Zhang, R.R.; Li, M.; Yuan, X.; Pan, Z.C. Influence of rainfall intensity and slope on suspended solids and phosphorus losses in runoff. Environ. Sci. Pollut. R 2019, 26, 33963–33975. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niinemets, U.; Tamm, U. Species differences in timing of leaf fall and foliage chemistry modify nutrient resorption efficiency in deciduous temperate forest stands. Tree Physiol. 2005, 25, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Zurdo, P.; Escudero, A.; Mediavilla, S. N resorption efficiency and proficiency in response to winter cold in three evergreen species. Plant Soil 2015, 394, 87–98. [Google Scholar] [CrossRef]
- Lü, X.T.; Reed, S.; Yu, Q.; He, N.P.; Wang, Z.W.; Han, X.G. Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semiarid grassland. Glob. Chang. Biol. 2013, 19, 2775–2784. [Google Scholar] [CrossRef]
- Ren, Y.; Liu, M.; Tian, L.; Tian, X.; Li, Z. Biodiversity, Conservation and Management of Taibaishan Nature Reserve; China Forestry Publishing House: Beijing, China, 2006; pp. 2–26. (In Chinese) [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
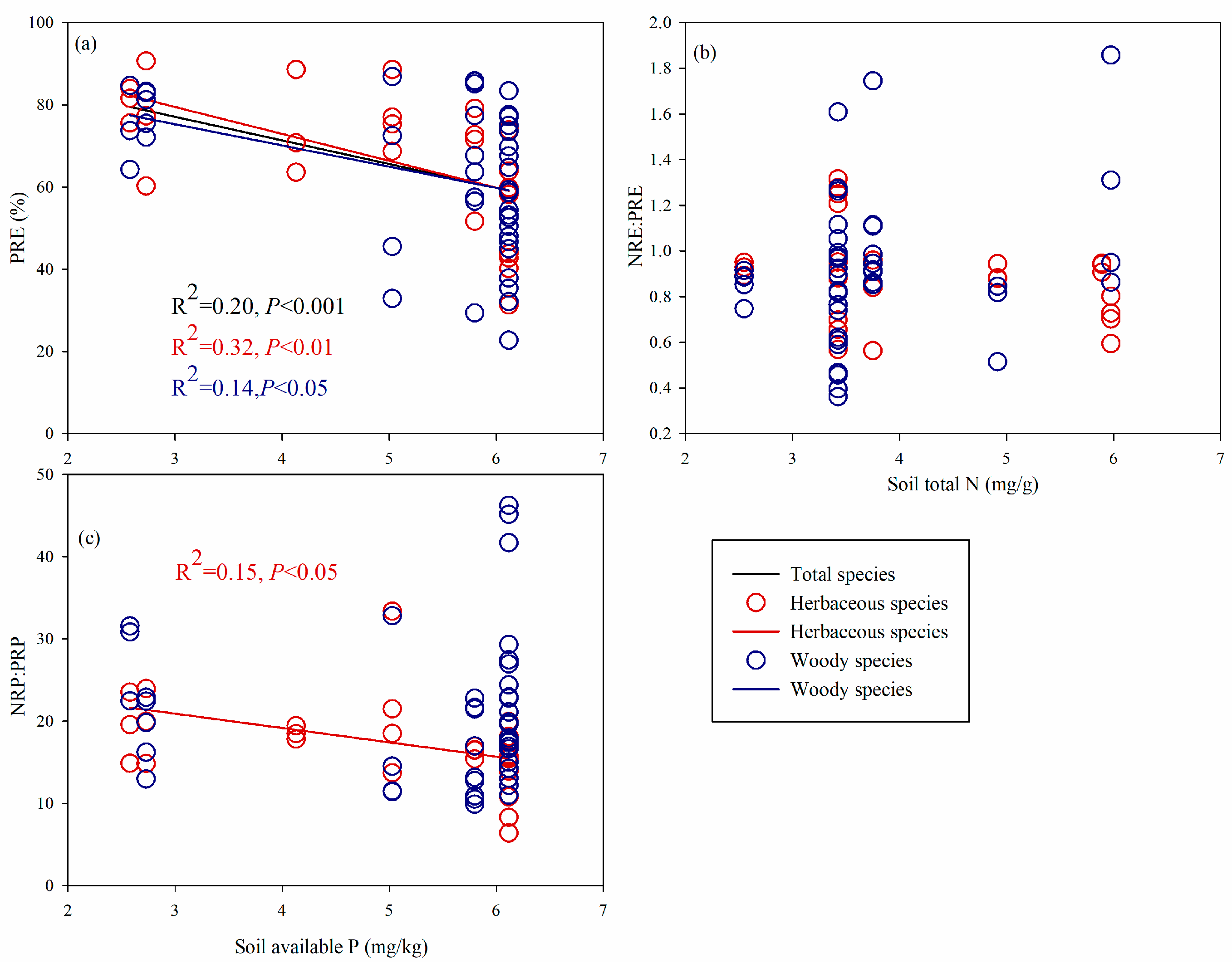{kind=link}
| n | Ng | Pg | NRE | PRE | NRP | PRP | NRP:PRP | NRE:PRE | |
|---|---|---|---|---|---|---|---|---|---|
| Total species | 53 | ||||||||
| Blomberg’s K | 0.60 | 0.43 | 0.31 | 0.34 | 0.44 | 0.35 | 0.49 | 0.45 | |
| Blomberg’s P | 0.03 | 0.25 | 0.44 | 0.69 | 0.40 | 0.54 | 0.04 | 0.07 | |
| herbaceous species | 16 | ||||||||
| Blomberg’s K | 0.85 | 0.96 | 0.70 | 0.32 | 0.69 | 0.28 | 0.35 | 0.36 | |
| Blomberg’s P | 0.04 | 0.02 | 0.10 | 0.97 | 0.20 | 0.96 | 0.92 | 0.92 | |
| woody species | 37 | ||||||||
| Blomberg’s K | 0.43 | 0.32 | 0.33 | 0.39 | 0.37 | 0.48 | 0.57 | 0.53 | |
| Blomberg’s P | 0.35 | 0.81 | 0.82 | 0.54 | 0.68 | 0.19 | 0.08 | 0.15 |
| NRP | PRP | NRE | PRE | NRP:PRP | NRE:PRE | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | SS% | p | Estimate | SS% | p | Estimate | SS% | p | Estimate | SS% | p | Estimate | SS% | p | Estimate | SS% | p | |
| Intercept | 12.67 | <0.001 | 4.12 | <0.001 | 64.39 | <0.001 | −30.89 | 0.28 | 26.29 | <0.001 | 1.12 | <0.001 | ||||||
| fixed effect | ||||||||||||||||||
| MAT | 0.37 | 12.38 | 0.08 | −1.27 | 24.75 | <0.01 | ||||||||||||
| MAP | −0.004 | 10.25 | <0.001 | 0.13 | 18.54 | <0.01 | ||||||||||||
| SAP | −1.81 | 1.67 | 0.17 | |||||||||||||||
| STN | −0.06 | 0.66 | 0.3 | |||||||||||||||
| PGF | −2.56 | 4.40 | 0.14 | −2.82 | 10.23 | <0.001 | 3.67 | 0.01 | 0.47 | 33.55 | 0.14 | 0.35 | −3.48 | 3.51 | 0.67 | −0.57 | 0.10 | 0.06 |
| MAT:PGF | −0.01 | 18.20 | 0.97 | −0.45 | 25.27 | 0.43 | ||||||||||||
| MAP:PGF | 0.01 | 29.13 | <0.01 | 0.05 | 19.43 | 0.34 | ||||||||||||
| SAP:PGF | 1.37 | 6.68 | 0.37 | |||||||||||||||
| STN:PGF | 0.15 | 7.15 | 0.04 | |||||||||||||||
| ramdom effect | ||||||||||||||||||
| site | 4.91 | 0.14 | 0 | 1 | 0 | 0.39 | 0 | 0.39 | 2.94 | 0.21 | 0 | 0.38 | ||||||
| Total model | ||||||||||||||||||
| R2m | 0.22 | 0.37 | 0.34 | 0.24 | 0.07 | 0.07 | ||||||||||||
| R2c | 0.28 | 0.37 | 0.34 | 0.24 | 0.1 | 0.07 | ||||||||||||
| p | <0.01 | <0.001 | <0.001 | <0.01 | 0.3 | 0.4 | ||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Chen, S.; Wang, X.; Liu, X.; Wang, X.; Zhu, R.; Mo, W.; Wang, R.; Zhang, S. Different Environmental and Phylogenetic Controls over the Altitudinal Variation in Leaf N and P Resorption Traits between Woody and Herbaceous Plants. Forests 2023, 14, 5. https://doi.org/10.3390/f14010005
Chen H, Chen S, Wang X, Liu X, Wang X, Zhu R, Mo W, Wang R, Zhang S. Different Environmental and Phylogenetic Controls over the Altitudinal Variation in Leaf N and P Resorption Traits between Woody and Herbaceous Plants. Forests. 2023; 14(1):5. https://doi.org/10.3390/f14010005
Chicago/Turabian StyleChen, Haoxuan, Shuang Chen, Xiaochun Wang, Xinrui Liu, Xue Wang, Rong Zhu, Weiyi Mo, Ruili Wang, and Shuoxin Zhang. 2023. "Different Environmental and Phylogenetic Controls over the Altitudinal Variation in Leaf N and P Resorption Traits between Woody and Herbaceous Plants" Forests 14, no. 1: 5. https://doi.org/10.3390/f14010005