

Long-Term Effects of Altered Precipitation Patterns on Alpine Vegetation Species Composition on the Qinghai-Tibet Plateau

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
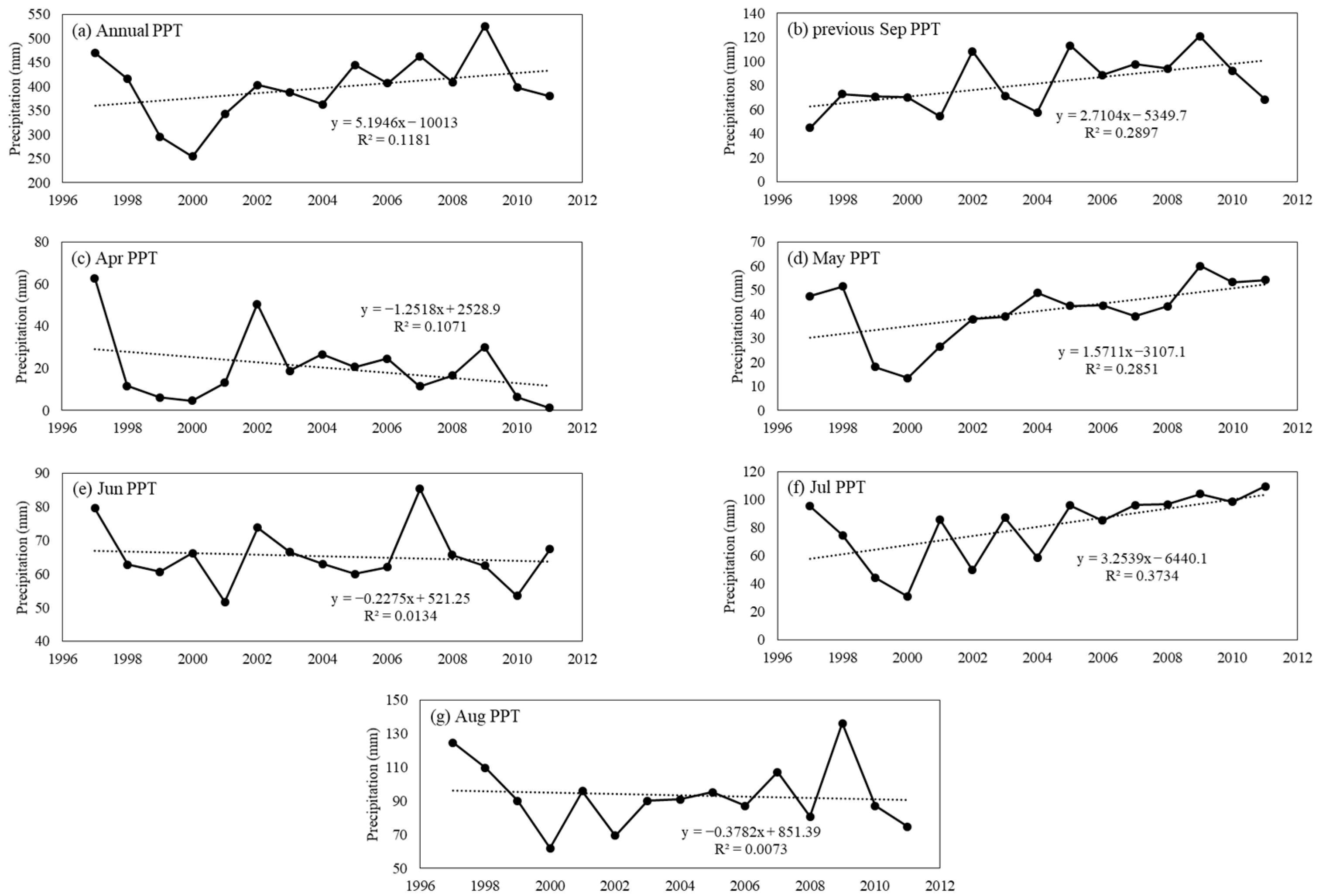
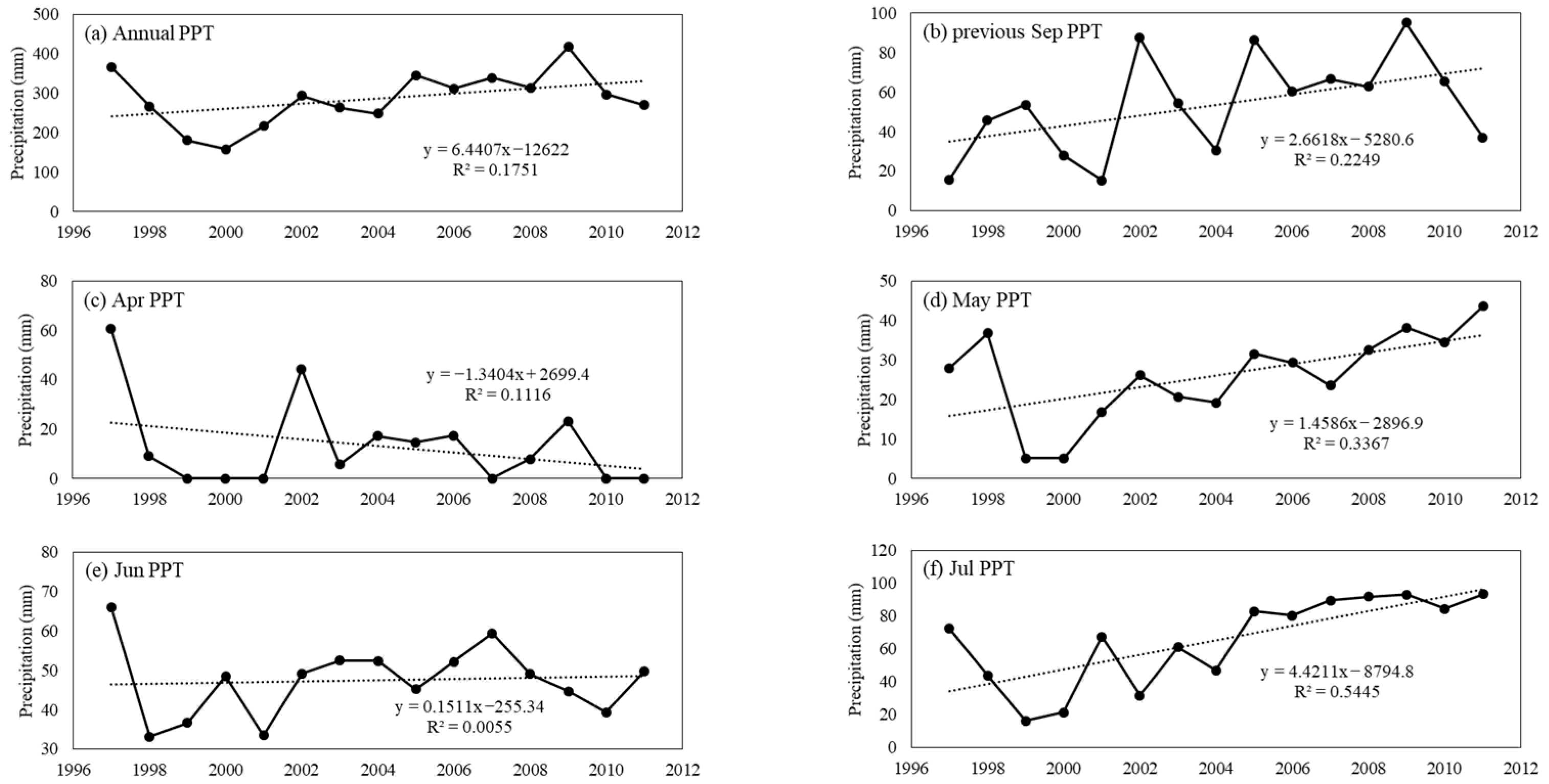
2.2. Data Collection
2.3. Statistical Analysis
2.4. Random Forest
3. Results
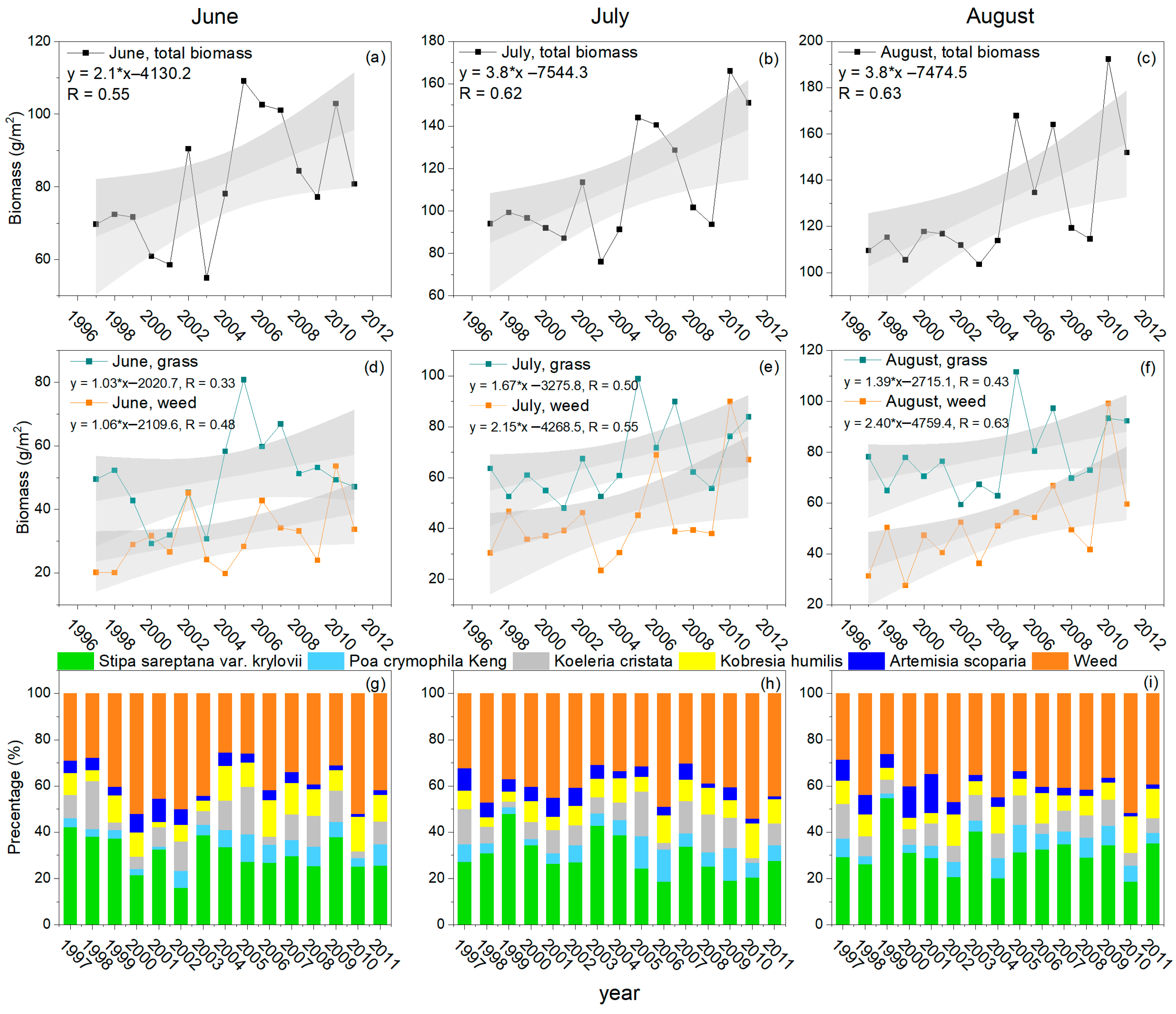
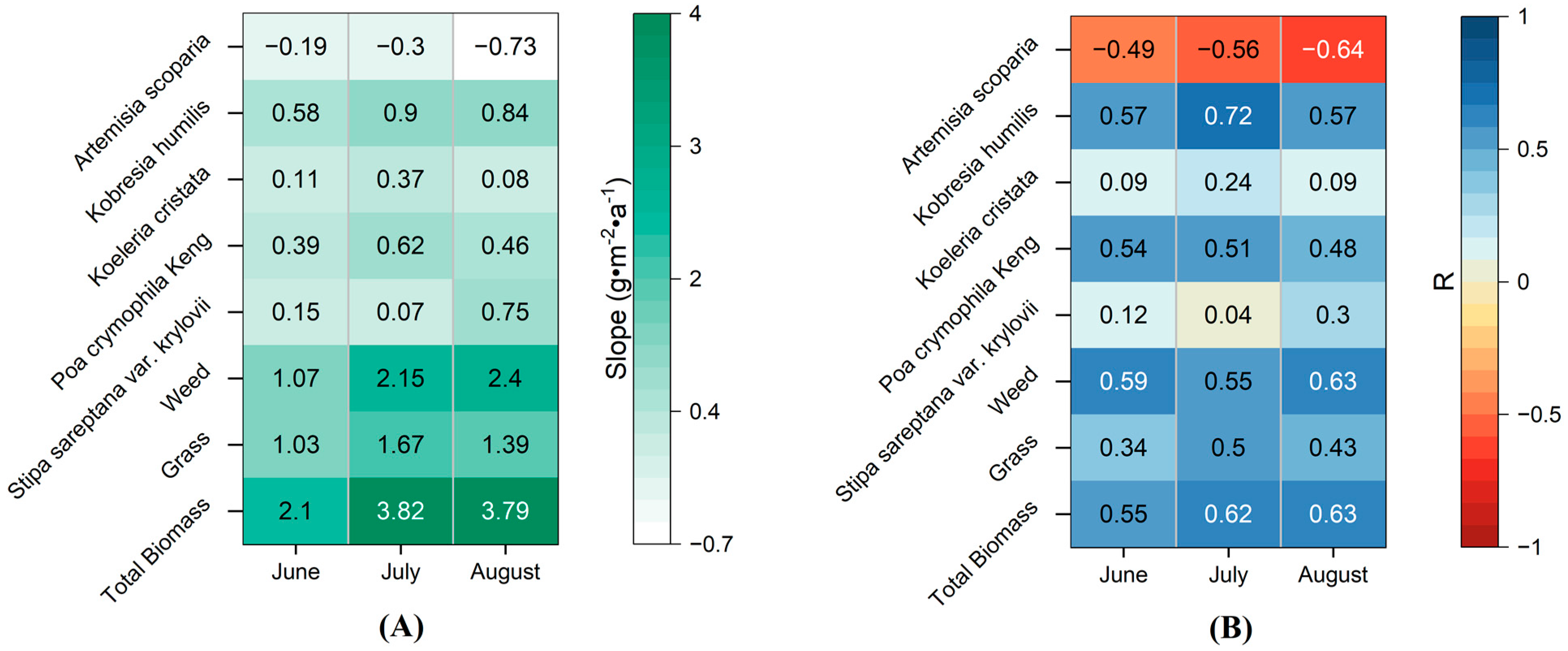
3.1. Variations in Aboveground Biomass of Different Species in Alpine Grassland
3.2. Response of Vegetation Biomass to Precipitation in Different Periods
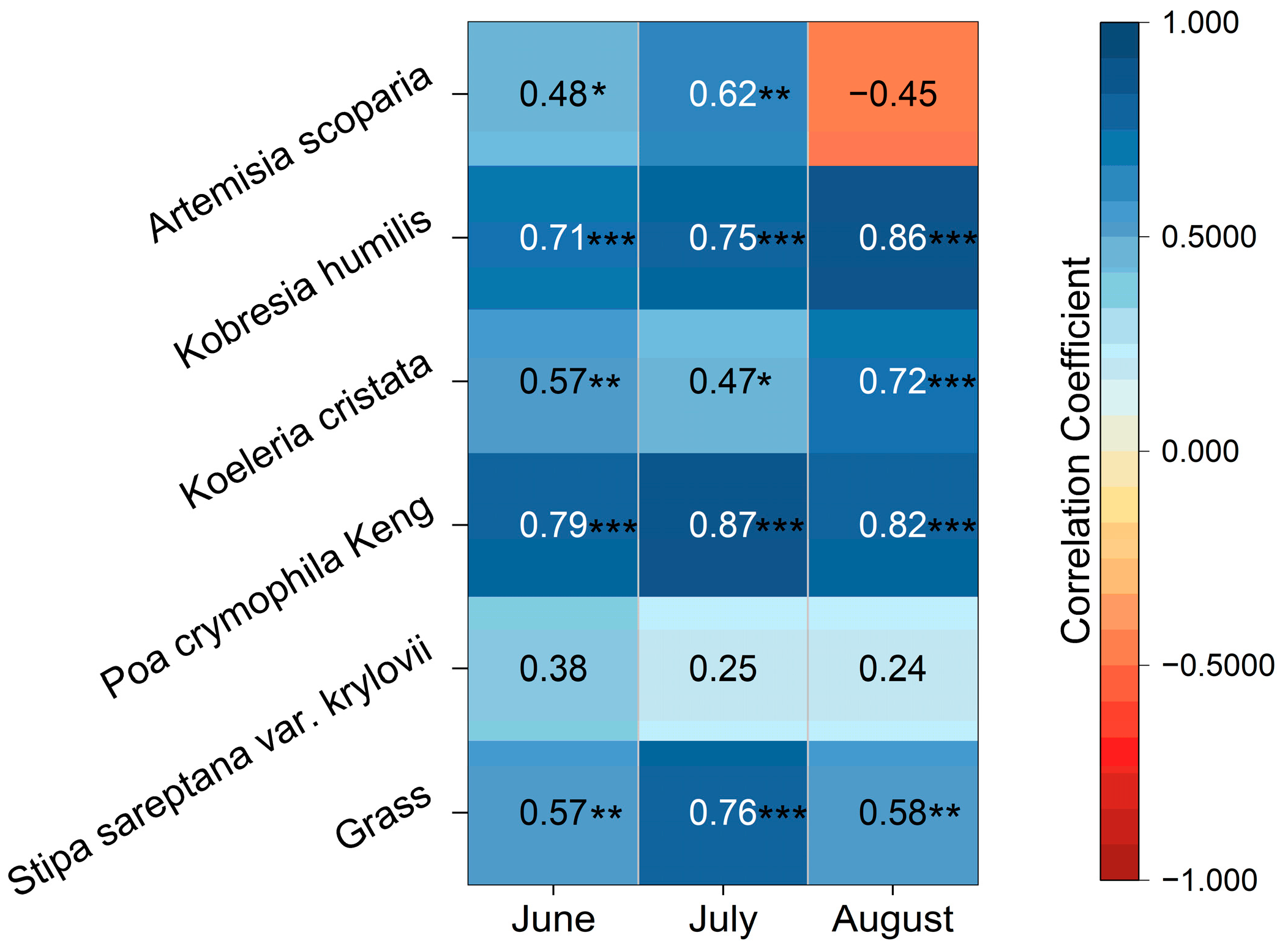
3.2.1. Correlation between Vegetation Biomass and Multi-Period Precipitation
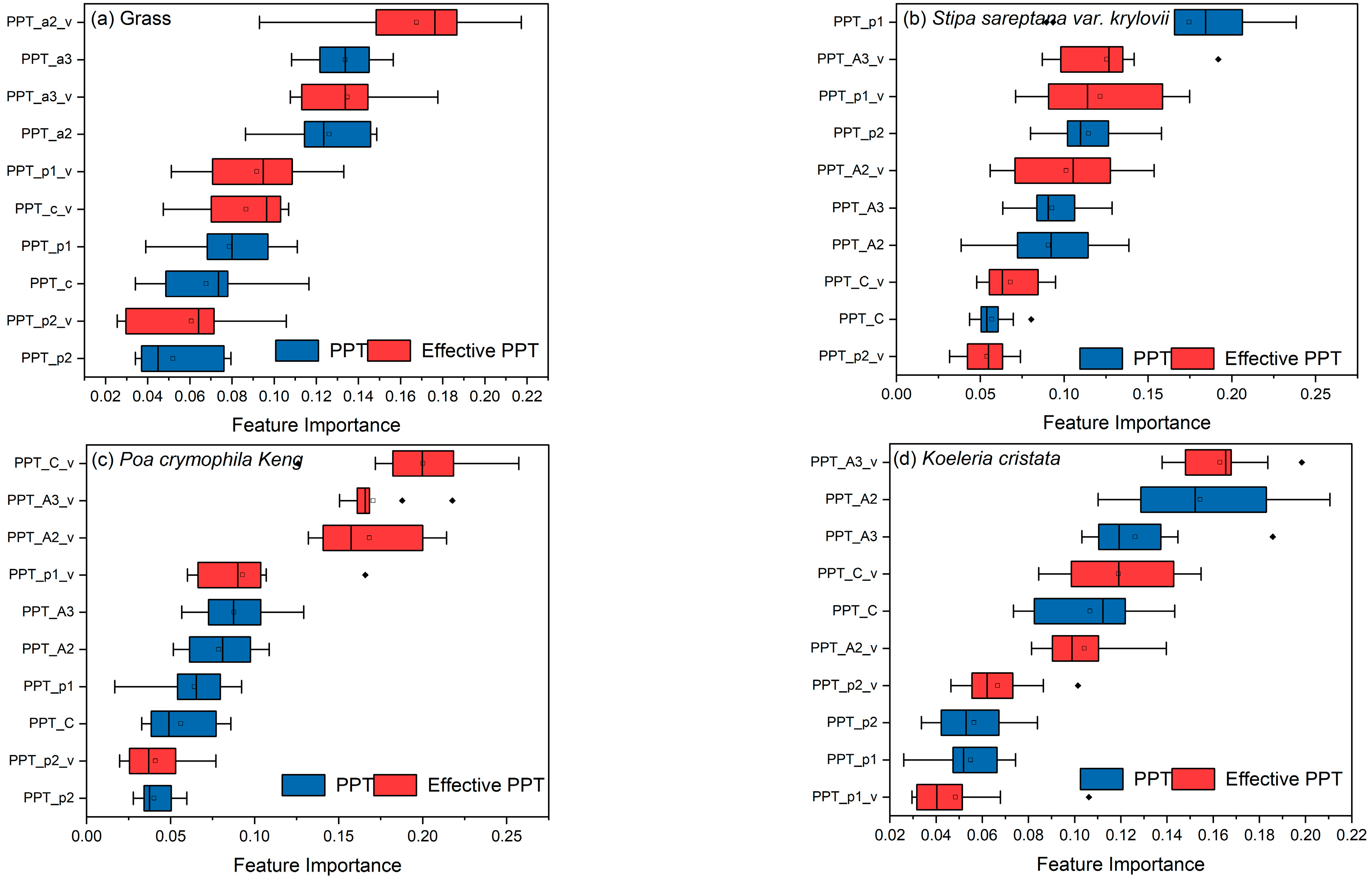
3.2.2. Contributions of Precipitation in Different Periods on Vegetation Biomass
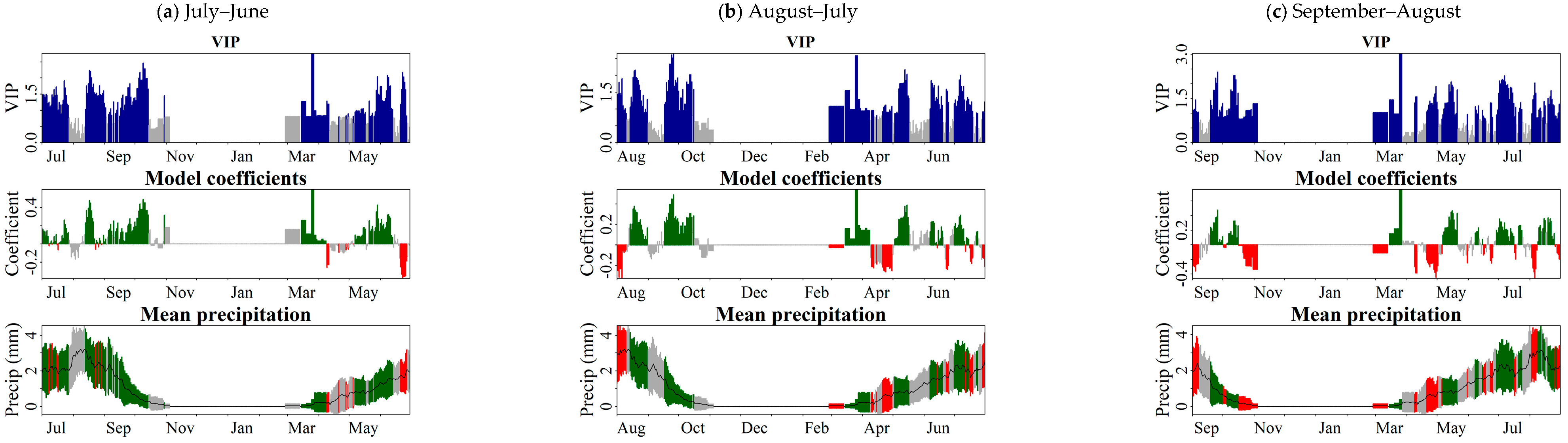
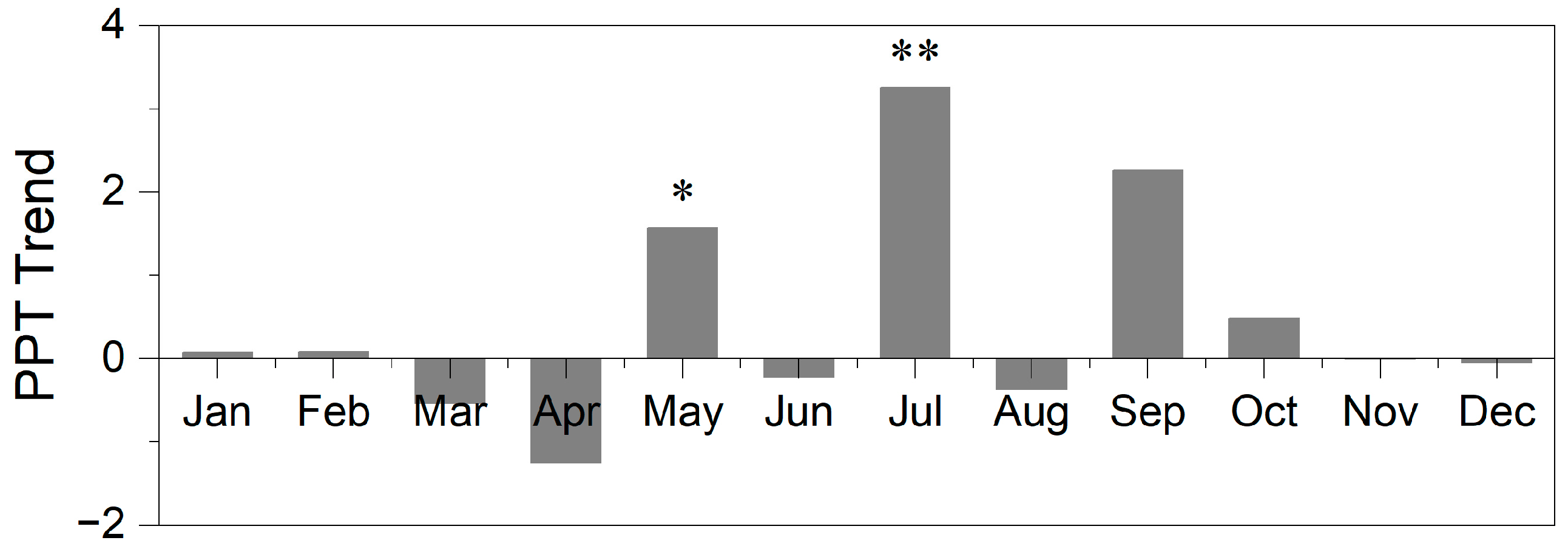
3.3. Crucial Periods of Effective Precipitation for Aboveground Biomass
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A




References
- Jiao, W.; Wang, L.; Smith, W.K.; Chang, Q.; Wang, H.; D’Odorico, P. Observed Increasing Water Constraint on Vegetation Growth over the Last Three Decades. Nat. Commun. 2021, 12, 3777. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y.; Ju, W.; Chen, J.; Ciais, P.; Cescatti, A.; Sardans, J.; Janssens, I.; Wu, M.; Berry, J.; et al. Recent Global Decline of CO2 Fertilization Effects on Vegetation Photosynthesis. Science 2020, 370, 1295–1300. [Google Scholar] [CrossRef]
- Anderson, R.; Canadell, J.; Randerson, J.; Jackson, R.; Hungate, B.; Baldocchi, D.; Ban-Weiss, G.; Bonan, G.; Caldeira, K.; Cao, L.; et al. Biophysical Considerations in Forestry for Climate Protection. Front. Ecol. Environ. 2011, 9, 174–182. [Google Scholar] [CrossRef] [Green Version]
- Cramer, W.; Bondeau, A.; Woodward, F.I.; Prentice, I.C.; Betts, R.A.; Brovkin, V.; Cox, P.M.; Fisher, V.; Foley, J.A.; Friend, A.D.; et al. Global Response of Terrestrial Ecosystem Structure and Function to CO2 and Climate Change: Results from Six Dynamic Global Vegetation Models: Ecosystem dynamics, CO2 and climate change. Glob. Change Biol. 2001, 7, 357–373. [Google Scholar] [CrossRef] [Green Version]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-Driven Increases in Global Terrestrial Net Primary Production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardgett, R.D.; Bullock, J.M.; Lavorel, S.; Manning, P.; Schaffner, U.; Ostle, N.; Chomel, M.; Durigan, G.; Fry, E.L.; Johnson, D.; et al. Combatting Global Grassland Degradation. Nat. Rev. Earth Environ. 2021, 2, 720–735. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Y.; Ma, Y.; Ma, W.; Liang, C.; Flynn, D.F.B.; Schmid, B.; Fang, J.; He, J.-S. Field-Based Observations of Regional-Scale, Temporal Variation in Net Primary Production in Tibetan Alpine Grasslands. Biogeosciences 2014, 11, 2003–2016. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.A.; Duncanson, L.; Hancock, S.; Neuenschwander, A.; Thomas, N.; Hofton, M.; Fatoyinbo, L.; Simard, M.; Marshak, C.Z.; Armston, J.; et al. Fusing Simulated GEDI, ICESat-2 and NISAR Data for Regional Aboveground Biomass Mapping. Remote Sens. Environ. 2021, 253, 112234. [Google Scholar] [CrossRef]
- Scurlock, J.M.O.; Johnson, K.; Olson, R.J. Estimating Net Primary Productivity from Grassland Biomass Dynamics Measurements: Net primary productivity of grasslands. Glob. Change Biol. 2002, 8, 736–753. [Google Scholar] [CrossRef] [Green Version]
- Lovell, J.T.; MacQueen, A.H.; Mamidi, S.; Bonnette, J.; Jenkins, J.; Napier, J.D.; Sreedasyam, A.; Healey, A.; Session, A.; Shu, S.; et al. Genomic Mechanisms of Climate Adaptation in Polyploid Bioenergy Switchgrass. Nature 2021, 590, 438–444. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, H.; Mi, Z.; Zhang, Z.; Wang, Y.; Xu, W.; Jiang, L.; He, J.-S. Climate Warming Reduces the Temporal Stability of Plant Community Biomass Production. Nat. Commun. 2017, 8, 15378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapp, A.; Smith, M. Interannual Variability in Net Primary Production and Precipitation—Response. Science 2001, 293, 1723. [Google Scholar]
- Knapp, A.K.; Smith, M.D. Variation Among Biomes in Temporal Dynamics of Aboveground Primary Production. Science 2001, 291, 481–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Mi, Z.; Lin, L.; Wang, Y.; Zhang, Z.; Zhang, F.; Wang, H.; Liu, L.; Zhu, B.; Cao, G.; et al. Shifting Plant Species Composition in Response to Climate Change Stabilizes Grassland Primary Production. Proc. Natl. Acad. Sci. USA 2018, 115, 4051–4056. [Google Scholar] [CrossRef] [Green Version]
- Grime, J.P.; Brown, V.K.; Thompson, K.; Masters, G.J.; Hillier, S.H.; Clarke, I.P.; Askew, A.P.; Corker, D.; Kielty, J.P. The Response of Two Contrasting Limestone Grasslands to Simulated Climate Change. Science 2000, 289, 762–765. [Google Scholar] [CrossRef]
- Guo, Q.; Hu, Z.; Li, S.; Li, X.; Sun, X.; Yu, G. Spatial Variations in Aboveground Net Primary Productivity along a Climate Gradient in Eurasian Temperate Grassland: Effects of Mean Annual Precipitation and Its Seasonal Distribution. Glob. Change Biol. 2012, 18, 3624–3631. [Google Scholar] [CrossRef]
- Sala, O.E.; Gherardi, L.A.; Reichmann, L.; Jobbágy, E.; Peters, D. Legacies of Precipitation Fluctuations on Primary Production: Theory and Data Synthesis. Phil. Trans. R. Soc. B 2012, 367, 3135–3144. [Google Scholar] [CrossRef] [Green Version]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; de Luca, E.; et al. Biodiversity Increases the Resistance of Ecosystem Productivity to Climate Extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- Li, P.; Peng, C.; Wang, M.; Li, W.; Zhao, P.; Wang, K.; Yang, Y.; Zhu, Q. Quantification of the Response of Global Terrestrial Net Primary Production to Multifactor Global Change. Ecol. Indic. 2017, 76, 245–255. [Google Scholar] [CrossRef]
- Xia, M.; Jia, K.; Zhao, W.; Liu, S.; Wei, X.; Wang, B. Spatio-Temporal Changes of Ecological Vulnerability across the Qinghai-Tibetan Plateau. Ecol. Indic. 2021, 123, 107274. [Google Scholar] [CrossRef]
- Dong, S.; Shang, Z.; Gao, J.; Boone, R.B. Enhancing Sustainability of Grassland Ecosystems through Ecological Restoration and Grazing Management in an Era of Climate Change on Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2020, 287, 106684. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, X.; Wang, Q.; Wang, C.; Zhan, Z.; Chen, L.; Yan, J.; Qu, R. Vegetation Net Primary Productivity and Its Response to Climate Change during 2001–2008 in the Tibetan Plateau. Sci. Total Environ. 2013, 444, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Zhang, Y.; Zhu, J.; Liu, Y.; Zu, J.; Zhang, J. The Influences of Climate Change and Human Activities on Vegetation Dynamics in the Qinghai-Tibet Plateau. Remote Sens. 2016, 8, 876. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.-W.; Shao, Q.-Q.; Liu, J.-Y.; Wang, J.-B.; Harris, W.; Chen, Z.-Q.; Zhong, H.-P.; Xu, X.-L.; Liu, R.-G. Assessment of Effects of Climate Change and Grazing Activity on Grassland Yield in the Three Rivers Headwaters Region of Qinghai–Tibet Plateau, China. Environ. Monit. Assess. 2010, 170, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Wang, X.; Zhang, X. Impacts of Climate Change and Human Activities on the Aboveground Production in Alpine Grasslands: A Case Study of the Source Region of the Yellow River, China. Arab. J. Geosci. 2017, 10, 17. [Google Scholar] [CrossRef]
- Xu, H.; Wang, X.; Zhang, X. Alpine Grasslands Response to Climatic Factors and Anthropogenic Activities on the Tibetan Plateau from 2000 to 2012. Ecol. Eng. 2016, 92, 251–259. [Google Scholar] [CrossRef]
- Gao, Q.; Li, Y.; Wan, Y.; Qin, X.; Jiangcun, W.; Liu, Y. Dynamics of Alpine Grassland NPP and Its Response to Climate Change in Northern Tibet. Clim. Change 2009, 97, 515–528. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, C.; Wang, Z.; Chen, Y.; Gang, C.; An, R.; Li, J. Vegetation Dynamics and Its Driving Forces from Climate Change and Human Activities in the Three-River Source Region, China from 1982 to 2012. Sci. Total Environ. 2016, 563–564, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, J.; Liang, S.; Liu, S.; Zhou, Y. A Perception of the Nexus “Resistance, Recovery, Resilience” of Vegetations Responded to Extreme Precipitation Pulses in Arid and Semi-Arid Regions: A Case Study of the Qilian Mountains Nature Reserve, China. Sci. Total Environ. 2022, 843, 157105. [Google Scholar] [CrossRef]
- Guo, Z.; Lou, W.; Sun, C.; He, B. Trend Changes of the Vegetation Activity in Northeastern East Asia and the Connections with Extreme Climate Indices. Remote Sens. 2022, 14, 3151. [Google Scholar] [CrossRef]
- Wang, L.; Hu, F.; Miao, Y.; Zhang, C.; Zhang, L.; Luo, M. Changes in Vegetation Dynamics and Relations with Extreme Climate on Multiple Time Scales in Guangxi, China. Remote Sens. 2022, 14, 2013. [Google Scholar] [CrossRef]
- Heisler-White, J.L.; Knapp, A.K.; Kelly, E.F. Increasing Precipitation Event Size Increases Aboveground Net Primary Productivity in a Semi-Arid Grassland. Oecologia 2008, 158, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Hu, Z.; Li, S.; Yu, G.; Sun, X.; Zhang, L.; Mu, S.; Zhu, X.; Wang, Y.; Li, Y.; et al. Contrasting Responses of Gross Primary Productivity to Precipitation Events in a Water-Limited and a Temperature-Limited Grassland Ecosystem. Agric. For. Meteorol. 2015, 214–215, 169–177. [Google Scholar] [CrossRef]
- Parton, W.; Morgan, J.; Smith, D.; Del Grosso, S.; Prihodko, L.; LeCain, D.; Kelly, R.; Lutz, S. Impact of Precipitation Dynamics on Net Ecosystem Productivity. Glob. Change Biol. 2012, 18, 915–927. [Google Scholar] [CrossRef]
- Fernandes, V.M.C.; Machado de Lima, N.M.; Roush, D.; Rudgers, J.; Collins, S.L.; Garcia-Pichel, F. Exposure to Predicted Precipitation Patterns Decreases Population Size and Alters Community Structure of Cyanobacteria in Biological Soil Crusts from the Chihuahuan Desert: Changing Rainfall Effects on Soil Cyanobacteria. Environ. Microbiol. 2018, 20, 259–269. [Google Scholar] [CrossRef]
- Liu, W.J.; Li, L.F.; Biederman, J.A.; Hao, Y.B.; Zhang, H.; Kang, X.M.; Cui, X.Y.; Wang, Y.F.; Li, M.W.; Xu, Z.H.; et al. Repackaging Precipitation into Fewer, Larger Storms Reduces Ecosystem Exchanges of CO2 and H2O in a Semiarid Steppe. Agric. For. Meteorol. 2017, 247, 356–364. [Google Scholar] [CrossRef]
- Sun, Q.; Meyer, W.S.; Koerber, G.R.; Marschner, P. Prior Rainfall Pattern Determines Response of Net Ecosystem Carbon Exchange to a Large Rainfall Event in a Semi-Arid Woodland. Agric. Ecosyst. Environ. 2017, 247, 112–119. [Google Scholar] [CrossRef]
- Lehmann, J.; Coumou, D.; Frieler, K. Increased Record-Breaking Precipitation Events under Global Warming. Clim. Change 2015, 132, 501–515. [Google Scholar] [CrossRef]
- Spinoni, J.; Naumann, G.; Carrao, H.; Barbosa, P.; Vogt, J. World Drought Frequency, Duration, and Severity for 1951–2010: World drought climatologies for 1951–2010. Int. J. Climatol. 2014, 34, 2792–2804. [Google Scholar] [CrossRef] [Green Version]
- Siteur, K.; Eppinga, M.B.; Karssenberg, D.; Baudena, M.; Bierkens, M.F.P.; Rietkerk, M. How Will Increases in Rainfall Intensity Affect Semiarid Ecosystems? Water Resour. Res. 2014, 50, 5980–6001. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.-H.; Li, X.-R.; Zhang, F.; Zhang, Z.-S.; Chen, Y.-L. Effects of Rainfall Intensity and Intermittency on Woody Vegetation Cover and Deep Soil Moisture in Dryland Ecosystems. J. Hydrol. 2016, 543, 270–282. [Google Scholar] [CrossRef]
- Del Grosso, S.J.; Parton, W.J.; Derner, J.D.; Chen, M.; Tucker, C.J. Simple Models to Predict Grassland Ecosystem C Exchange and Actual Evapotranspiration Using NDVI and Environmental Variables. Agric. For. Meteorol. 2018, 249, 1–10. [Google Scholar] [CrossRef]
- Yu, L.; Li, K.; Tao, B.; Xu, M. Simulating and Assessing the Adaptability of Geographic Distribution of Vegetation to Climate Change in China. Prog. Geogr. 2010, 29, 1326–1332. [Google Scholar]
- Li, C.; Li, Q.; Zhao, L.; Ge, S.; Chen, D.; Dong, Q.; Zhao, X. Land-Use Effects on Organic and Inorganic Carbon Patterns in the Topsoil around Qinghai Lake Basin, Qinghai-Tibetan Plateau. Catena 2016, 147, 345–355. [Google Scholar] [CrossRef]
- Ali, M.; Mubarak, S. Effective Rainfall Calculation Methods for Field Crops: An Overview, Analysis and New Formulation. ARJA 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Guo, L.; Dai, J.; Wang, M.; Xu, J.; Luedeling, E. Responses of Spring Phenology in Temperate Zone Trees to Climate Warming: A Case Study of Apricot Flowering in China. Agric. For. Meteorol. 2015, 201, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Luedeling, E.; Gassner, A. Partial Least Squares Regression for Analyzing Walnut Phenology in California. Agric. For. Meteorol. 2012, 158–159, 43–52. [Google Scholar] [CrossRef]
- Yu, H.; Luedeling, E.; Xu, J. Winter and Spring Warming Result in Delayed Spring Phenology on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2010, 107, 22151–22156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Cheng, J.; Luedeling, E.; Koerner, S.E.; He, J.-S.; Xu, J.; Gang, C.; Li, W.; Luo, R.; Peng, C. Critical Climate Periods for Grassland Productivity on China’s Loess Plateau. Agric. For. Meteorol. 2017, 233, 101–109. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Amagai, Y.; Kudo, G.; Sato, K. Changes in Alpine Plant Communities under Climate Change: Dynamics of Snow-Meadow Vegetation in Northern Japan over the Last 40 Years. Appl. Veg. Sci. 2018, 21, 561–571. [Google Scholar] [CrossRef]
- Zeidler, M.; Duchoslav, M.; Banaš, M.; Lešková, M. Impacts of Introduced Dwarf Pine (Pinus mugo) on the Diversity and Composition of Alpine Vegetation. Community Ecol. 2012, 13, 213–220. [Google Scholar] [CrossRef]
- Matteodo, M.; Ammann, K.; Verrecchia, E.P.; Vittoz, P. Snowbeds Are More Affected than Other Subalpine-Alpine Plant Communities by Climate Change in the Swiss Alps. Ecol. Evol. 2016, 6, 6969–6982. [Google Scholar] [CrossRef] [PubMed]
- Batllori, E.; Blanco-Moreno, J.M.; Ninot, J.M.; Gutiérrez, E.; Carrillo, E. Vegetation Patterns at the Alpine Treeline Ecotone: The Influence of Tree Cover on Abrupt Change in Species Composition of Alpine Communities. J. Veg. Sci. 2009, 20, 814–825. [Google Scholar] [CrossRef]
- Hsu, J.S.; Adler, P.B. Anticipating Changes in Variability of Grassland Production due to Increases in Interannual Precipitation Variability. Ecosphere 2014, 5, art58. [Google Scholar] [CrossRef]
- Knapp, A.K.; Fay, P.A.; Blair, J.M.; Collins, S.L.; Smith, M.D.; Carlisle, J.D.; Harper, C.W.; Danner, B.T.; Lett, M.S.; McCarron, J.K. Rainfall Variability, Carbon Cycling, and Plant Species Diversity in a Mesic Grassland. Science 2002, 298, 2202–2205. [Google Scholar] [CrossRef] [Green Version]
- Knapp, A.K.; Burns, C.E.; Fynn, R.W.S.; Kirkman, K.P.; Morris, C.D.; Smith, M.D. Convergence and Contingency in Production–Precipitation Relationships in North American and South African C4 Grasslands. Oecologia 2006, 149, 456–464. [Google Scholar] [CrossRef]
- Koerner, S.E.; Collins, S.L. Interactive Effects of Grazing, Drought, and Fire on Grassland Plant Communities in North America and South Africa. Ecology 2014, 95, 98–109. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, X.; Guo, W.; Liu, T.; Li, S.; Zhang, J. Long-Term Effects of Altered Precipitation Patterns on Alpine Vegetation Species Composition on the Qinghai-Tibet Plateau. Forests 2023, 14, 47. https://doi.org/10.3390/f14010047
Ni X, Guo W, Liu T, Li S, Zhang J. Long-Term Effects of Altered Precipitation Patterns on Alpine Vegetation Species Composition on the Qinghai-Tibet Plateau. Forests. 2023; 14(1):47. https://doi.org/10.3390/f14010047
Chicago/Turabian StyleNi, Xiangnan, Wei Guo, Tong Liu, Shuheng Li, and Junzhe Zhang. 2023. "Long-Term Effects of Altered Precipitation Patterns on Alpine Vegetation Species Composition on the Qinghai-Tibet Plateau" Forests 14, no. 1: 47. https://doi.org/10.3390/f14010047