
Effects of Wild Forest Fires on Genetic Diversity and Population Structure of a Boreal Conifer, White Spruce (Picea glauca (Moench) Voss): Implications for Genetic Resource Management and Adaptive Potential under Climate Change

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites, Experimental Design and Sampling
2.2. DNA Extraction and Genotyping
2.3. Data Analysis
2.3.1. Genetic Diversity, Inbreeding Coefficient and Effective Population Size
2.3.2. Genetic Differentiation and Genetic Structure
2.3.3. Selection Scan
3. Results
3.1. Genetic Diversity, Fixation Index and Effective Population Size
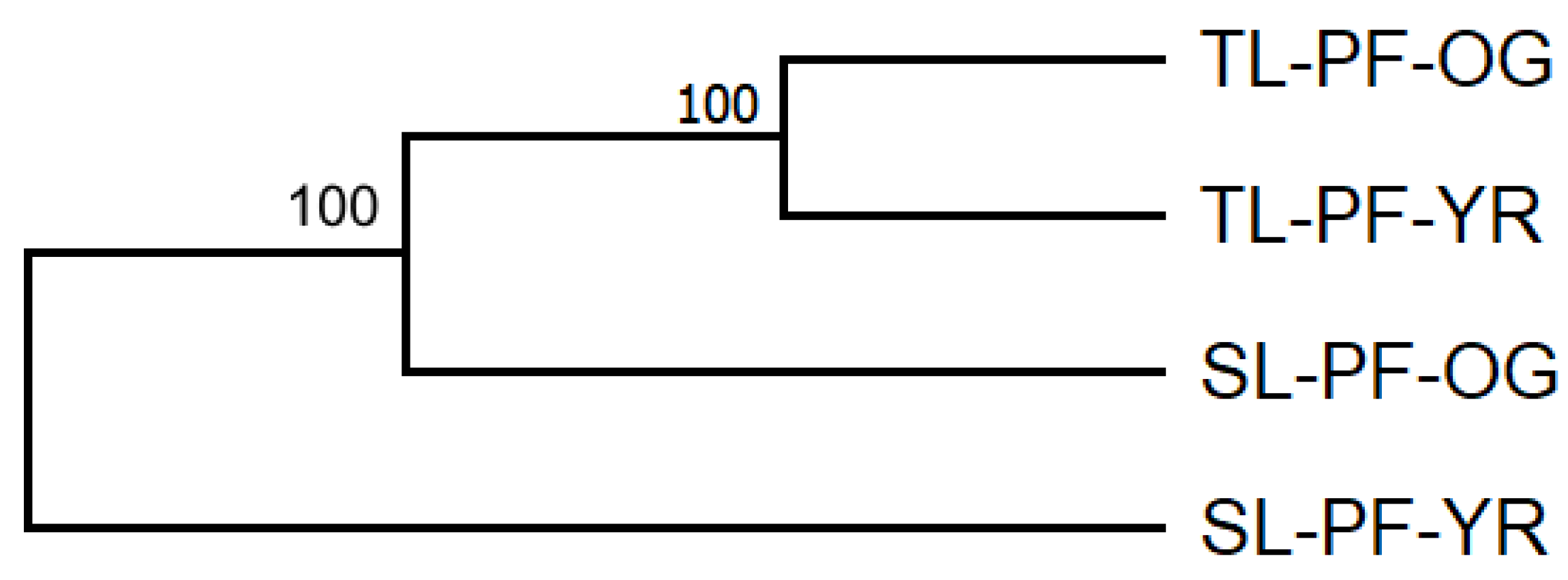
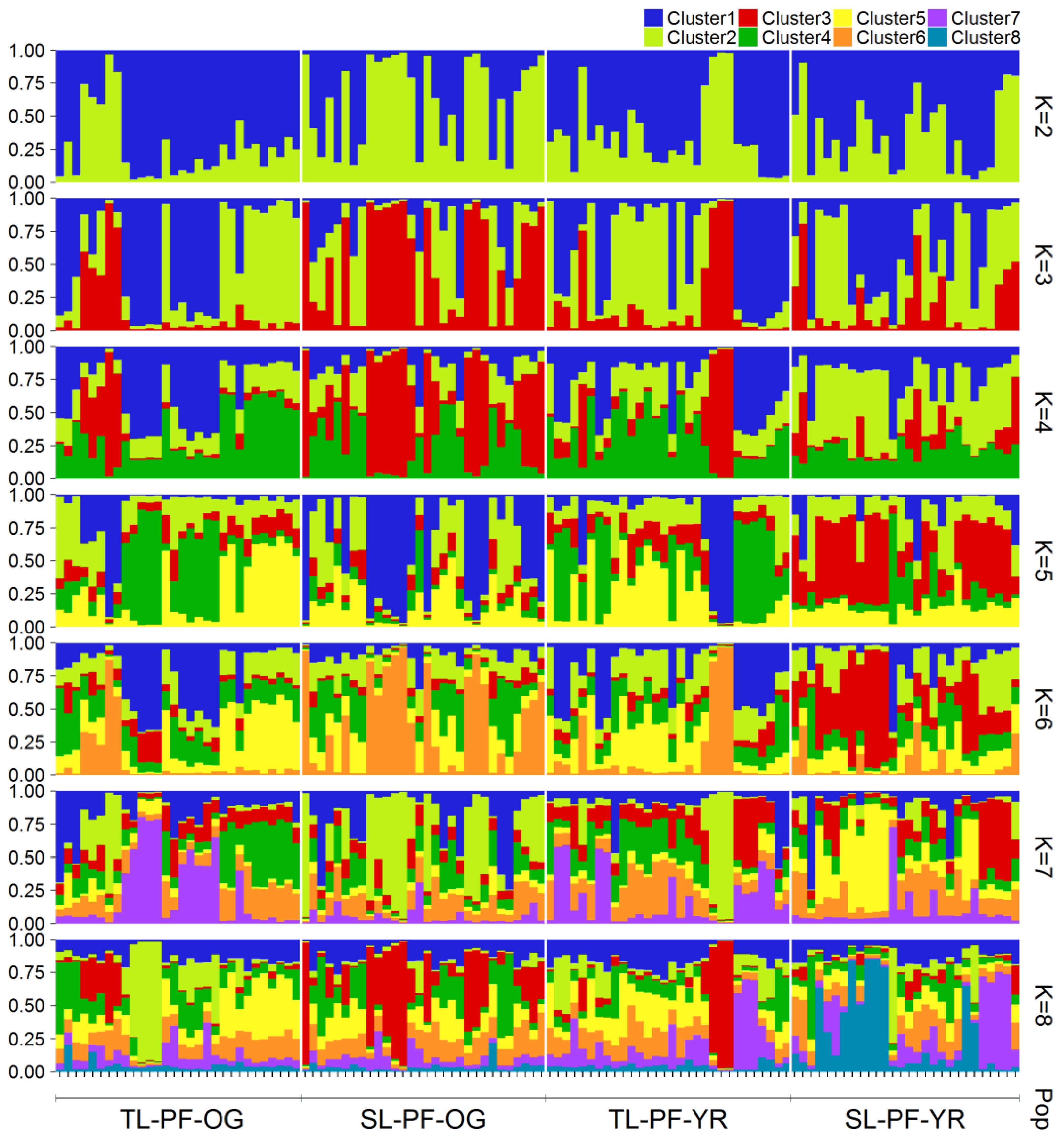
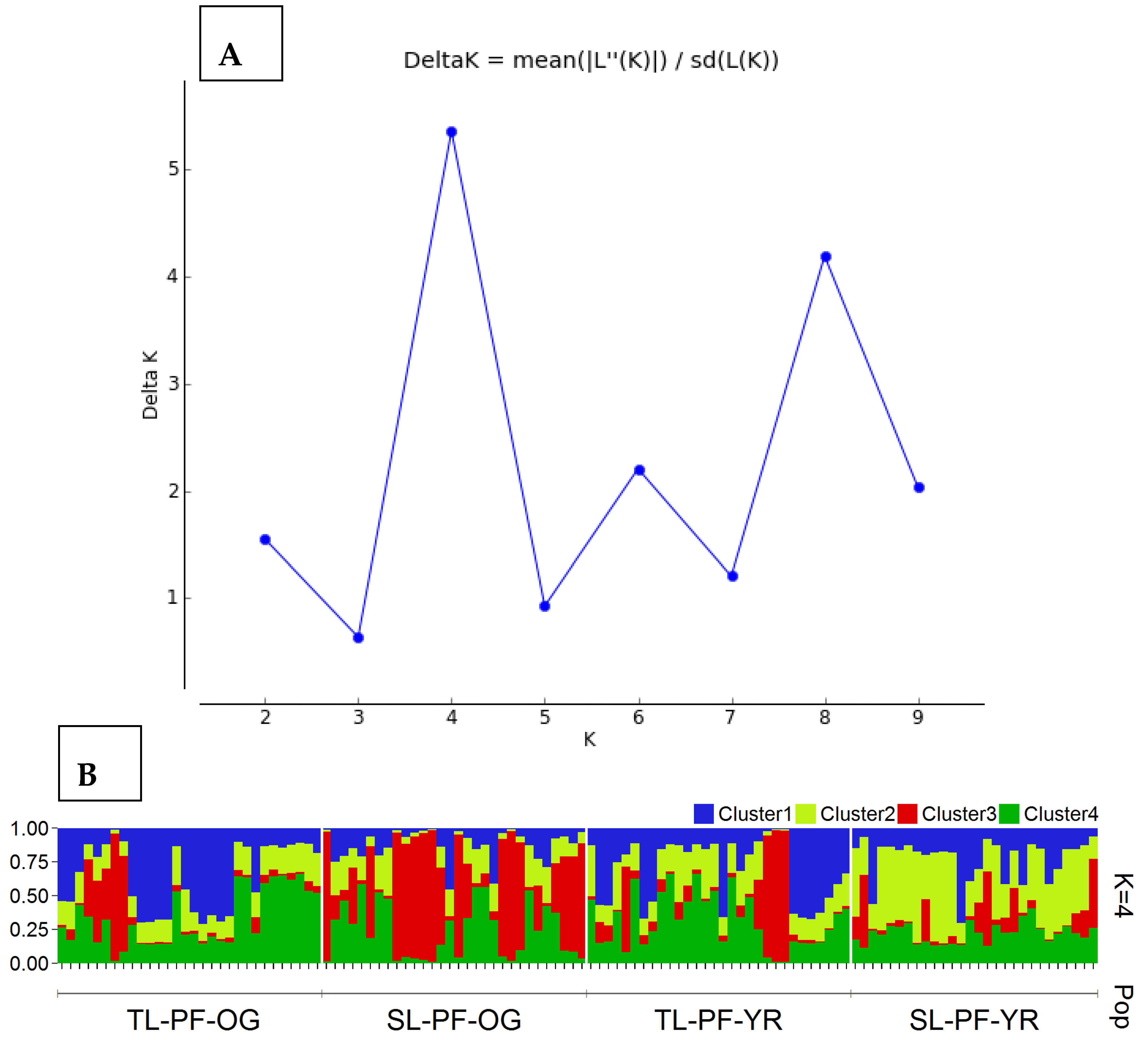
3.2. Genetic Differentiation and Population Genetic Structure
3.3. Selection Scan
4. Discussion
4.1. Genetic Diversity, Inbreeding and Effective Population Size
4.2. Genetic Differentiation and Genetic Structure
4.3. The Selection Effects of Fire
4.4. Implications for White Spruce Genetic Resources Management and Adaptive Potential under Climate Change
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reusch, T.B.H.; Ehlers, A.; Hammerli, A.; Worm, B. Ecosystem recovery after climate extremes enhanced by genotypic diversity. Proc. Natl. Acad. Sci. USA 2005, 102, 2826–2831. [Google Scholar] [CrossRef] [Green Version]
- Roger, F.; Godhe, A.; Gamfeldt, L. Genetic diversity and ecosystem functioning in the face of multiple stressors. PLoS ONE 2012, 7, e45007. [Google Scholar] [CrossRef]
- Salo, T.; Gustafsson, C. The effect of genetic diversity on ecosystem functioning in vegetated coastal ecosystems. Ecosystems 2016, 19, 1429–1444. [Google Scholar] [CrossRef]
- Rajora, O.P.; Zinck, J.W.R. Genetic diversity, structure and effective population size of old-growth versus second-growth populations of keystone and long-lived conifer, eastern white pine (Pinus strobus): Conservation value and climate adaptation potential. Front. Genet. 2021, 12, 650299. [Google Scholar] [CrossRef]
- Vandewoestijne, S.; Schtickzelle, N.; Baguette, M. Positive correlation between genetic diversity and fitness in a large, well-connected metapopulation. BMC Biol. 2008, 6, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markert, J.A.; Champlin, D.M.; Gutjahr-Gobell, R.; Grear, J.S.; Anne Kuhn, A.; McGreevy, T.J., Jr.; Roth, A.; Mark, J.; Bagley, M.J.; Nacci, D.E. Population genetic diversity and fitness in multiple environments. BMC Evol. Biol. 2010, 10, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.; Tanaka, R.; Yamamoto, D.; Noriyuki, S.; Kawata, M. Balanced genetic diversity improves population fitness. Proc. R. Soc. B 2018, 285, 2017–2045. [Google Scholar] [CrossRef] [PubMed]
- Buchert, G.P.; Rajora, O.P.; Hood, J.V.; Dancik, B.P. Effects of harvesting on genetic diversity in old-growth eastern white pine (Pinus strobus L.) in Ontario, Canada. Conserv. Biol. 1997, 11, 747–758. [Google Scholar] [CrossRef]
- Rajora, O.P. Genetic biodiversity impacts of silvicultural practices, phenotypic selection in white spruce. Theor. Appl. Genet. 1999, 99, 954–961. [Google Scholar] [CrossRef]
- Rajora, O.P.; Rahman, M.H.; Buchert, G.P.; Dancik, B.P. Microsatellite analysis of genetic effects of harvesting in old-growth eastern white pine (Pinus strobus) in Ontario, Canada. Mol. Ecol. 2000, 9, 339–348. [Google Scholar] [CrossRef]
- Banks, S.C.; Gary, G.J.; Smith, A.L.; Davies, I.D.; Driscoll, D.A.; Gill, A.M.; Lindenmayer, D.B.; Peakall, R. How does ecological disturbance influence genetic diversity? Trends Ecol. Evol. 2013, 28, 670–679. [Google Scholar] [CrossRef]
- Stocks, B.J.; Wotton, B.M.; Flannigan, M.D.; Fosbert, M.A.; Cahoon, D.R.; Goldammer, J.G. Boreal forest fire regimes and climate change. In Remote Sensing and Climate Modeling: Synergies and Limitations; Beniston, M., Verstraete, M.M., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 2001; pp. 233–246. [Google Scholar]
- Parchman, T.L.; Gompert, Z.; Mudge, J.; Schilkey, F.D.; Benkman, C.W.; Buerkle, C.A. Genome-wide association genetics of an adaptive trait in lodgepole pine. Mol. Ecol. 2012, 21, 2991–3005. [Google Scholar] [CrossRef] [PubMed]
- Rowe, S. Concepts of fire effects on plant individuals and species. In The Role of Fire in Northern Circumpolar Ecosystems; Wein, R.W., MacLean, D.A., Eds.; John Wiley & Sons Ltd.: Toronto, ON, Canada, 1983; pp. 135–1554. [Google Scholar]
- Ne’eman, G.; Goubitz, S.; Nathan, R. Reproductive traits of Pinus halepensis in the light of fire—A critical review. Plant Ecol. 2004, 171, 69–79. [Google Scholar] [CrossRef]
- Rajora, O.P.; Pluhar, S.A. Genetic diversity impacts of forest fires, forest harvesting, alternative reforestation practices in black spruce (Picea mariana). Theor. Appl. Genet. 2003, 106, 1213–1224. [Google Scholar] [CrossRef]
- Ayre, D.J.; Ottewell, K.M.; Krauss, S.L.; Whelan, R.J. Genetic structure of seedling cohorts following repeated wildfires in the fire-sensitive shrub Persoonia mollis ssp. nectens. J. Ecol. 2009, 97, 752–760. [Google Scholar] [CrossRef]
- Gershberg, A.; Neeman, G.; Ben-Shlomo, R. Genetic structure of a naturally regenerating post-fire seedling population: Pinus halepensis as a case study. Front. Plant Sci. 2016, 7, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budde, K.B.; Gonzalez-Martınez, S.C.; Navascue´s, M.; Burgarella, C.; Mosca, E.; Lorenzo, Z.; Zabal-Aguirre, M.; Vendramin, G.G.; Verdu, M.; Pausas, J.G.; et al. Increased fire frequency promotes stronger spatial genetic structure and natural selection at regional and local scales in Pinus halepensis Mill. Ann. Bot. 2017, 119, 1061–1072. [Google Scholar] [CrossRef] [Green Version]
- von Takach Dukai, B.; Peakall, R.; Lindenmayer, D.B.; Banks, S.C. The influence of fire and silvicultural practices on the landscape-scale genetic structure of an Australian foundation tree species. Conserv. Genet. 2020, 21, 231–246. [Google Scholar] [CrossRef]
- Flaniggan, M.D.; Van Wagner, C.E. Climate change and wild fires in Canada. Can. J. For. Res. 1991, 21, 66–72. [Google Scholar] [CrossRef]
- Flaniggan, M.D.; Stocks, B.J.; Wotton, B.M. Climate change and forest fires. Sci. Total Environ. 2000, 262, 221–229. [Google Scholar] [CrossRef]
- Amiro, B.D.; Flannigan, M.D.; Stocks, B.J.; Todd, J.B.; Wotton, M.B. Boreal forest fires: An increasing issue in a changing climate. In Proceedings of the XII World Forestry Congress, Quebec City, QC, Canada, 21–28 September 2003; Available online: http://www.fao.org/3/xii/0207-b3.htm (accessed on 12 April 2022).
- Bergeron, Y.; Leduc, A.; Harvey, B.D.; Gauthier, S. Natural fire regime: A guide for sustainable management of the Canadian boreal forest. Silva Fenn. 2002, 36, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Kuuluvainen, T.; Angelstam, P.; Frelich, L.; Jõgiste, K.; Koivula, M.; Kubota, Y.; Lafleur, B.; Macdonald, E. Natural disturbance-based forest management: Moving beyond retention and continuous-cover forestry. Front. For. Glob. Change 2021, 4, 629020. [Google Scholar] [CrossRef]
- Schiller, G.; Ne’eman, G.; Korol, L. Post-fire vegetation dynamics in a native Pinus halepensis Mill. forest on Mt. Carmel, Israel. Isr. J. Plant Sci. 1997, 45, 297–308. [Google Scholar] [CrossRef]
- Uchiyamaa, K.; Gotob, S.; Tsudaac, Y.; Takahashib, Y.; Yuji, I. Genetic diversity and genetic structure of adult and buried seed populations of Betula maximowicziana in mixed and post-fire stands. For. Ecol. Manag. 2006, 237, 119–126. [Google Scholar] [CrossRef]
- Worth, J.R.P.; Jordon, G.J.; Marthick, J.R.; Sakaguchi, S.; Colhoun, E.A.; Williamson, G.J.; Ito, M.; Bowman, M.J.S. Fire is a major driver of patterns of genetic diversity in two co-occurring Tasmanian paleoendemic confers. J. Biogeogr. 2017, 44, 1254–1267. [Google Scholar] [CrossRef]
- Hosie, R.C. Native Trees of Canada; Fitzhenry and Whiteside Ltd.: Don Mills, ON, Canada, 1979. [Google Scholar]
- Rajora, O.P.; Mann, I.K.; Shi, Y.Z. Genetic diversity, population structure of boreal white spruce (Picea glauca) in pristine conifer-dominated, mixed-wood forest stands. Can. J. Bot. 2005, 83, 1096–1105. [Google Scholar] [CrossRef]
- O’Connell, L.M.; Mosseler, A.; Rajora, O.P. Impacts of forest fragmentation on the mating system and genetic diversity of white spruce (Picea glauca) at the landscape level. Heredity 2006, 97, 418–426. [Google Scholar] [CrossRef] [Green Version]
- Fageria, M.S.; Rajora, O.P. Effects of harvesting of increasing intensities on genetic diversity and population structure of white spruce. Evol. Appl. 2013, 6, 778–794. [Google Scholar] [CrossRef]
- Fageria, M.S.; Rajora, O.P. Effects of silvicultural practices on genetic diversity and population structure of white spruce in Saskatchewan. Tree Genet. Genomes 2014, 10, 287–296. [Google Scholar] [CrossRef]
- O’Connell, L.M.; Mosseler, A.; Rajora, O.P. Extensive long-distance pollen dispersal in a fragmented landscape maintains genetic diversity in white spruce. J. Hered. 2007, 98, 640–645. [Google Scholar] [CrossRef]
- Stewart, J.D.; Hogg, E.H.; Hurdle, P.A.; Stadt, K.J.; Tollestrup, P.; Lieffers, V.J. Dispersal of white spruce seed in mature aspen stands. Can. J. Bot. 1998, 76, 181–188. [Google Scholar]
- Padbury, G.A.; Acton, D.F.; Stushnoff, C.T. The Ecoregions of Saskatchewan; University of Regina Press: Regina, SK, Canada, 1998. [Google Scholar]
- Weir, J.M.H.; Johnson, E.A.; Miyanishi, K. Fire frequency and the age mosaic of the mixed-wood boreal forest in western Canada. Ecol. Appl. 2000, 10, 1162–1177. [Google Scholar] [CrossRef]
- Weir, J.M.H. The Fire Frequency and Age Mosaic of a Mixed-Wood Boreal Forest. Master’s Thesis, University of Calgary: Calgary, AB, Canada, 1996. [Google Scholar]
- Rajora, O.P.; Mann, I.K. Development and characterization of Novel EST-based single-copy genic microsatellite DNA markers in white spruce and black spruce. Mol. Biol. Rep. 2021, 48, 2963–2971. [Google Scholar] [CrossRef]
- Rajora, O.P.; Rahman, M.H.; Dayanandan, S.; Mosseler, A. Isolation, characterization, inheritance and linkage of microsatellite DNA markers in white spruce (Picea glauca) and their usefulness in other spruce species. Mol. Genet. Genom. 2001, 264, 871–882. [Google Scholar] [CrossRef]
- Hodgetts, R.B.; Aleksiuk, M.A.; Brown, A.; Clark, C.; Macdonald, E.; Nadeem, S.; Khasa, D. Development of microsatellite markers for white spruce (Picea glauca) and related species. Theor. Appl. Genet. 2001, 102, 1252–1258. [Google Scholar] [CrossRef]
- van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-checker, software for identifying, correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX6, genetic analysis in Excel. Population genetic software for teaching, research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Rajora, O.P. Effects of forestry practices on genetic diversity: Implications for sustainable forest management and gene conservation. In Proceedings of the IUFRO Conference, Diversity and Adaptation in Forest Ecosystems in a Changing World, University of British Columbia, Vancouver, BC, Canada, 5–10 August 1996; Kluwer Academic Publishers: Amsterdam, The Netherlands, 1996. [Google Scholar]
- Bergmann, F.; Gregorius, H.R.; Larsen, J.B. Levels of genetic variation in European silver fir (Abies alba). Genetica 1990, 82, 1–10. [Google Scholar] [CrossRef]
- Waples, R.S.; Do, C. Linkage disequilibrium estimates of contemporary Ne using highly variable genetic markers: A largely untapped resource for applied conservation and evolution. Evol. Appl. 2010, 3, 244–262. [Google Scholar] [CrossRef] [PubMed]
- Do, C.; Waples, R.S.; Peel, D.; Macbeth, G.M.; Tillett, B.J.; Ovenden, J.R. NeEstimator V2: Reimplementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Resour. 2014, 14, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.; Rousset, F. GENEPOP version 1.2: Population genetics software for the exact tests and ecumenicism. J. Heredity 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity, genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction sites. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Felsenstein, J. PHYLIP (Phylogeny Inference Package) version 3.6; Distributed by the author; Department of Genome Sciences, University of Washington: Seattle, DC, USA, 2005. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Foll, M.; Gaggiotti, O. A genome-scan method to identify selected loci appropriate for both dominant and codominant marker: A Bayesian perspective. Genetics 2008, 180, 977–993. [Google Scholar] [CrossRef] [Green Version]
- Ledig, T.F.; Guries, R.P.; Bonefeld, B.A. The relation of growth to heterozygosity in pitch pine. Evolution 1983, 37, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- Fryer, J. Picea mariana. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory: Fort Collins, CO, USA, 2014. Available online: https://www.fs.usda.gov/database/feis/plants/tree/picmar/all.html (accessed on 7 April 2020).
- Templeton, A.R.; Levin, D.A. Evolutionary consequences of seed pools. Am. Nat. 1979, 114, 232–249. [Google Scholar] [CrossRef]
- Abrahamson, I. Picea glauca, white spruce. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory: Fort Collins, CO, USA, 2015. Available online: https://www.fs.fed.us/database/feis/plants/tree/picgla/all.html (accessed on 7 April 2020).
- Greene, D.F.; Zasada, J.C.; Sirois, L.; Kneeshaw, D.; Morin, H.; Charron, I.; Simard, M.J. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For. Res. 1999, 29, 824–839. [Google Scholar] [CrossRef]
- Bergeron, Y. Species and stand dynamics in the mixed woods of Quebec’ssouthern boreal forest. Ecology 2000, 81, 1500–1516. [Google Scholar] [CrossRef]
- Alden, J.; Loopstra, C. Genetic diversity and population structure of Picea glauca on an altitudinal gradient in interior Alaska. Can. J. For. Res. 1987, 17, 1519–1526. [Google Scholar] [CrossRef]
- Tremblay, N.; Simon, J.P. Genetic structure of marginal populations of white spruce as its northern limit of distribution in Nouveau-Quebec. Can. J. For. Res. 1989, 19, 1371–1379. [Google Scholar] [CrossRef]
- Rajora, O.P.; Dancik, B.P. Population genetic variation, structure, and evolution in Engelmann spruce, white spruce, and their natural hybrid complex in Alberta. Can. J. Bot. 2000, 78, 768–780. [Google Scholar]
- Gartner, F.M.; Lieffers, V.J.; Macdonald, S.E. Ecology and management of natural regeneration of white spruce in the boreal forest. Environ. Rev. 2011, 19, 461–478. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Population | Population ID | Longitude W | Latitude N | Age Years |
|---|---|---|---|---|---|
| Tibiska Lake (TL) | Post-fire natural old growth (PF-OG) | TL-PF-OG | 106°06′41″ | 54°15′16″ | 168 |
| Post-fire natural young regeneration (PF-YR) | TL-PF-YR | 34 | |||
| Sanctuary Lake (SL) | Post-fire natural old growth (PF-OG) | SL-PF-OG | 106°33′25″ | 54°09′50″ | 128 |
| Post-fire natural young regeneration (PF-YR) | SL-PF-YR | 12 |
| Microsatellite Locus * | Repeat Unit | Total No. of Alleles | Allele Size Range (bp) |
|---|---|---|---|
| EST-based genic microsatellites * | |||
| RPGSE2 | (CTG)3G(CTG)3G(CTG)3 | 5 | 170–185 |
| RPGSE5 | (GAA)6 | 2 | 242–245 |
| RPGSE17 | (TCG)6 | 3 | 154–160 |
| RPGSE34 | (GA)10 | 16 | 237–269 |
| RPGSE35 | (TA)26 | 8 | 151–171 |
| RPGSE44 | (TA)9 | 7 | 204–218 |
| Mean no. of alleles per locus | 6.83 | ||
| Genomic microsatellites | |||
| SPAG003 | (AG)n | 15 | 118–146 |
| PGL14 | (AG)22 | 23 | 130–182 |
| UAPgGT8 | (GT)22 | 12 | 192–228 |
| UAPgCA91 | (CA)20 | 28 | 102–190 |
| Mean no. of alleles per locus | 19.50 | ||
| Overall mean no. of alleles per locus | 11.20 | ||
| Population | AT | A | AP | AE | HO | HE | F | LGP | GAO | GAE | Ne (0.05) | Ne (0.01) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tibiska Lake | ||||||||||||
| PF-OG | 95.0 | 9.50 | 2 | 6.32 | 0.507 | 0.656 | 0.224 | 3.18 | 131.0 | 622.0 | 42 | 66 |
| (2.17) | (1.70) | (0.094) | (0.092) | (0.103) | (0.61) | (2.87) | (26.36) | (29–68) | (45–115) | |||
| PF-YR | 90.0 | 9.00 | 3 | 5.92 | 0.495 | 0.658 | 0.307 | 3.08 | 132.0 | 574.0 | 33 | 46 |
| (1.78) | (1.39) | (0.099) | (0.092) | (0.102) | (1.05) | (2.65) | (20.33) | (23–51) | (32–71) | |||
| Sanctuary Lake | ||||||||||||
| PF-OG | 91.0 | 9.10 | 2 | 4.95 | 0.473 | 0.644 | 0.271 | 4.15 | 114.0 | 613.0 | 74 | 83 |
| (1.98) | (1.09) | (0.095) | (0.089) | (0.093) | (0.53) | (2.50) | (16.92) | (37–424) | (51–197) | |||
| PF-YR | 91.0 | 9.10 | 3 | 5.34 | 0.404 | 0.635 | 0.366 | 3.76 | 128.0 | 589.0 | 108 | 193 |
| (1.85) | (1.20) | (0.082) | (0.095) | (0.093) | (0.71) | (2.55) | (17.23) | (46-infinite) | (78-infinite) | |||
| Mean stand types | ||||||||||||
| PF-OG | 93.00 | 9.30 | 2 | 5.64 | 0.490 | 0.650 | 0.247 | 3.67 | 122.5 | 617.5 | 58 | 75 |
| PF-YR | 90.05 | 9.05 | 3 | 5.63 | 0.450 | 0.647 | 0.336 | 3.42 | 130.0 | 581.5 | 71 | 120 |
| Overall mean | 91.75 | 9.18 | 2.5 | 5.63 | 0.470 | 0.648 | 0.292 | 3.52 | 126.25 | 599.5 | 64 | 97 |
| TL-PF-OG | SL-PF-OG | TL-PF-YR | SL-PF-YR | |
|---|---|---|---|---|
| TL-PF-OG | 0.022 | 0.011 | 0.021 | |
| SL-PF-OG | 0.064 | 0.028 | 0.032 | |
| TL-PF-YR | 0.015 | 0.070 | 0.024 | |
| SL-PF-YR | 0.068 | 0.115 | 0.067 |
| Population Groups | No. of Populations | FIS | FIT | FST |
|---|---|---|---|---|
| All populations | 4 | 0.290 (0.094) | 0.314 (0.091) | 0.035 (0.002) |
| PF-OG | 2 | 0.247 (0.095) | 0.262 (0.094) | 0.022 (0.004) |
| PF-YR | 2 | 0.336 (0.095) | 0.353 (0.093) | 0.024 (0.004) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajora, O.P.; Fageria, M.S.; Fitzsimmons, M. Effects of Wild Forest Fires on Genetic Diversity and Population Structure of a Boreal Conifer, White Spruce (Picea glauca (Moench) Voss): Implications for Genetic Resource Management and Adaptive Potential under Climate Change. Forests 2023, 14, 157. https://doi.org/10.3390/f14010157
Rajora OP, Fageria MS, Fitzsimmons M. Effects of Wild Forest Fires on Genetic Diversity and Population Structure of a Boreal Conifer, White Spruce (Picea glauca (Moench) Voss): Implications for Genetic Resource Management and Adaptive Potential under Climate Change. Forests. 2023; 14(1):157. https://doi.org/10.3390/f14010157
Chicago/Turabian StyleRajora, Om P., Manphool S. Fageria, and Michael Fitzsimmons. 2023. "Effects of Wild Forest Fires on Genetic Diversity and Population Structure of a Boreal Conifer, White Spruce (Picea glauca (Moench) Voss): Implications for Genetic Resource Management and Adaptive Potential under Climate Change" Forests 14, no. 1: 157. https://doi.org/10.3390/f14010157