1. Introduction
Soil stability strongly correlates to vegetation cover; when large areas are cleared, soil becomes vulnerable to erosion. As land is exposed to direct rainfall and surface water runoff, sedimentation increases, particularly along human-made structures such as roads and landing sites [
1,
2].
Surface runoff, sedimentation and soil erosion are natural processes that play an integral role in landscape formation [
3,
4]. Erosion-related changes in the landscape are highly dependent on site-specific soil properties and climate [
5,
6]. It has been established that runoff and sediment production in forested areas is determined by rainfall amount, rainfall intensity, topography, canopy density, current vegetation type, and it can be exacerbated by the construction of roads and land use management [
7]. In forested areas, litter cover acts as a protective layer to slow down runoff and prevent the loss of nutrients from the soil [
8]. The impact of wild ungulates on runoff and sedimentation is often overlooked, with only a few studies investigating this interaction [
2,
9,
10].
Mammalian herbivores influence ecosystem-level processes, but the effects and their extent are context-dependent, thus generalizations remain elusive [
11,
12]. Related to the soil properties, the impact of ungulates is highly dependent on vegetation type and availability, climatic conditions, soil macro- and microfauna composition, and even soil type [
13]. Impacts can be derived from feeding behavior, wallowing, trampling, or rooting (also commonly referred to as grubbing) that affects the litter or soil itself, which varies according to population density and the behavior of each species [
14].
Of particular interest in this regard is the wild boar or wild pig (
Sus scrofa; in this paper, wild boar refers to the populations present in the species’ native range, and wild pig, to those in its invasive range [
15]), due to its preference for rooting as a feeding behavior [
14,
16], and their current population density and distribution; wild boar populations have significantly increased worldwide in recent decades [
17], which has caused an increase in species-related conflicts [
18]. Its impact on forest regeneration, vegetation cover, soil macroinvertebrates, and soil composition has been analyzed in a varied array of climatic and topographic conditions [
8,
9,
18,
19,
20], although the relationship between wild boar rooting and sedimentation in forested areas remains widely under-studied [
2].
Wild boar rooting is usually recognizable by disturbance of the topsoil layer [
20] and can have varying effects: it can increase nutrient cycles and decomposition rates; nutrient loss through leaching; reduce hillslope hydrological connectivity; decrease sediment transport; act as litter and sediment traps [
3]; and increase richness, evenness, and diversity of species. It has been argued that the impacts of wild boar activities can be seen in the decrease in organic matter in the topsoil and an increase in soil compaction because of wallowing, trampling, or resting, all resulting in an increase in sedimentation flow through sloped terrain [
4]. Although studies in Germany [
19], Hawaii [
21], Australia [
3], and Argentina [
14] found that any effect of wild pig rooting on soil properties is either not significant or supplanted by more pressing factors, such as precipitation, road construction and use, and vegetation cover. Overall, wild boar rooting overturns extensive areas of soil, leaving them bare of vegetation and causing soil disturbance with important effects on the dynamics of native plant communities [
13]. It is possible that it either triggers erosion by exposing fresh soil or improves the conditions of the forest soil by acting as a sediment trap, as is the case for tree throws [
22].
Highways built near forested areas, like in our study area, create a greater problem since the precipitation running down the road through pronounced slopes intensifies the transport of sediments. We hypothesized that, in a situation of this kind, soil runoff will cause measurable differences between sedimented and non-sedimented forest soil patches, in terms of soil characteristics and rooting occurrence by wild boar. We also expected that wild boar rooting would cause differences in the soil properties of disturbed and intact patches, and this would modify the outcome of the sedimentation process by redistributing soil layers and increasing soil penetration resistance, thus enhancing runoff. In the background of our expectations, the aims of this study were to:
- -
Measure the extent and spatio-temporal pattern of wild boar rooting in a forested area where soil sedimentation is intense;
- -
Determine the preference of soil used by wild boar, based on sediment cover;
- -
Investigate the impact of wild boar rooting on soil compaction, and soil chemical composition.
2. Materials and Methods
2.1. Study Area
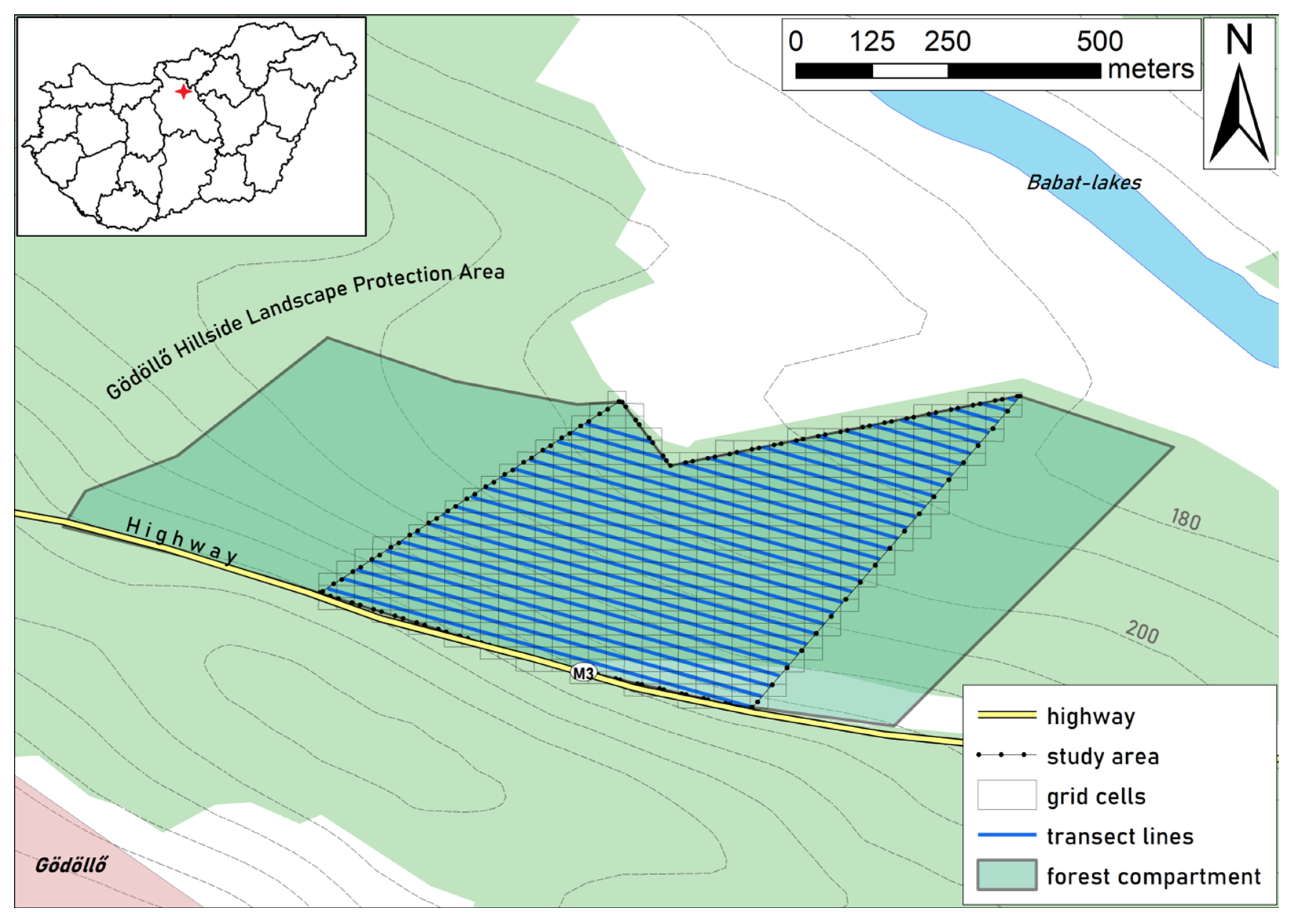
This study was conducted in the Babat Valley, forming part of the Gödöllő Hillside Landscape Protection Area and part of the Natura 2000 network. It is located near the city of Gödöllő in Pest County, Hungary (47°36′56.61″ N, 19°22′50.40″ E).
The Babat Valley is a transitional area between the Great Hungarian Plain and the North Hungarian Mountains. The average annual temperature is 9.5–10 °C. The amount of annual precipitation is 600 mm with most precipitation falling in early summer. During that time, significant erosion can occur due to the shallow topsoil cover, mainly comprised of loess mixed with sand [
23].
The study area includes 21.5 ha of coniferous (Scots pine:
Pinus sylvestris) and broadleaved deciduous (pedunculate oak:
Quercus robur and field maple:
Acer campestre) forest [
24] with eroded areas. This site was selected due to the presence of sedimented, non-sedimented, steep (>7°) and flat (<7°) areas within it. The soil type in the area is rusty brown forest soil. The highway built along the study area has exacerbated the pre-existing inclination of water erosion to shift sediments downward along the slope. The rainwater is directed into the protected area’s forested land and, as a result, causes severe soil erosion problems. In the examined area there are huge gullies, sometimes reaching 6–8 m in depth (
Supplementary Materials Figure S1).
The availability of sedimented and non-sedimented areas was estimated by walking along transects (
Figure 1) and classifying soil type at every 15 m (on 649 sampling points). Sites were classified as sedimented if the topsoil had a sandy, yellow texture and as non-sedimented where the topsoil had a typical loamy texture and a brown color. Based on the preliminary data, sedimented sites covered 22% of the study area (4% were flat and 18% steep) and non-sedimented sites covered 78% (56% flat and 22% steep).
In the spring of 2020, a yearly total of 5174 wild boar was hunted in Pest County [
25], so their continuous presence in the area is obvious.
2.2. Monitoring and Categorization of Wild Boar Rooting
To monitor wild boar rooting appropriately across time and location, we used a grid map system across the study area aligned to north–south and west–east directions (
Figure 1). The dimensions of each grid cell were 20 by 20 m (N = 610 cells in total) where new and persistent boar rooting was documented and monitored. We collected monthly data on the spatial distribution of rooted soil patches along 30 transects (50 to 500 m long), located 20 m from each other (approx. in parallel to the highway) from October 2019 to August 2020 (except February and March, due to bad weather conditions). When transects crossed rooted areas, the enter and exit GPS points of the rooting site were recorded. We used a Garmin GPSmap62 (Garmin International, Inc., Olathe, KS, USA) in-field, the Compass tool in a Huawei Y9 Smartphone (Huawei Technologies Co., Ltd., Shenzhen, Guangdong, China) and LocusMap application (version 3.38.6, Asamm Software s. r. o., Prague, Czech Republic) with Map1.eu (version Alpha 0.3, GitHub, Inc., San Francisco, CA, USA) installed, to geo-locate the rooting sites.
During monthly field work, grid cells were considered as rooted by wild boar if the soil surface was disturbed, litter moved away from the ground or mixed with the topsoil forming a microtopography of undulations as is common in pig rooting. This can be recognized by the bare soil stripped of vegetation, accumulation of grubbed soil along the edges, and the presence of wild boar footprints and other boar-related signs (i.e., feces and hair). The rooting occurrence recorded was classified according to three main factors: by depth as (1) shallow (<10 cm), (2) medium (10–30 cm), and (3) deep (>30 cm); actual soil sedimentation as (1) sedimented or (2) non-sedimented; and the steepness of the patch as (1) steep (>7°) or (2) flat (<7°).
2.3. Soil Sampling and Field Analyses
Soil sampling was performed at 3 specific locations of selected patches in rooted sites: (1) inside, in the deepest part of the patch disturbed by the wild boar, called “rooting”; (2) around, where the soil that had been removed from the rooting accumulated on top of the original topsoil, called the “ring”; and (3) outside, at 1 m distance from the edge of the rooting patch where the original soil conditions can be measured, called the “control” (
Supplementary Materials Figure S2).
To measure soil penetration resistance, we randomly chose 10 deep-rooted patches, 5 from sedimented and 5 from non-sedimented areas. In each of those, the compaction was determined in the rooting, ring, and control sections, with two repetitions, using an Eijkelkamp Penetrologger equipment at every 1 to 80 cm. A total of 30 measurements were taken, grouped by sedimentation (i.e., sedimented and non-sedimented) and location (i.e., rooting, ring, and control), categorized into 6 groups in total with equal sample sizes (n = 5).
For soil chemical analysis, we collected soil samples from the patches’ rooting, ring and control of another 20 randomly selected rooting patches, ensuring that sedimentation and rooting depth were equally represented in the sample, e.g., 5 rooted patches were selected for each combination of sedimentation (sedimented and non-sedimented) and rooting depth (deep and shallow) categories. Rooting with medium depth was not included in this comparison. The sampling procedure was done in April 2020, when the upper 25 cm of the soil on a 30 × 30 cm area was dug at each sampling point, the soil was extracted and mixed on a plastic surface and samples of 1 kg were taken for soil analysis.
Parameters of collected soil samples (pH, Total Organic Carbon (TOC—m/m %), including total nitrogen (g/kg), total phosphorus (g/kg), exchangeable potassium (mmol/kg), clay (m/m %) and Cation Exchange Capacity (CEC—mmol/kg) were evaluated by using a Near InfraRed (NIR) spectrometer (Wavelength Range: 1300–2600 nm MEMS technology) through the SoilCares mobile app.
An additional descriptive soil profile analysis was performed in the field with a 100 cm stainless-steel Pürckhauer-type soil probe on 6 deep rooting patches (
Tables S1 and S2). The soil horizons (A, B, and C) were identified based on color and texture (lighter to darker; sandy to loamy), based on the Munsell soil color chart and expert assessment in August 2020.
2.4. Data Analysis
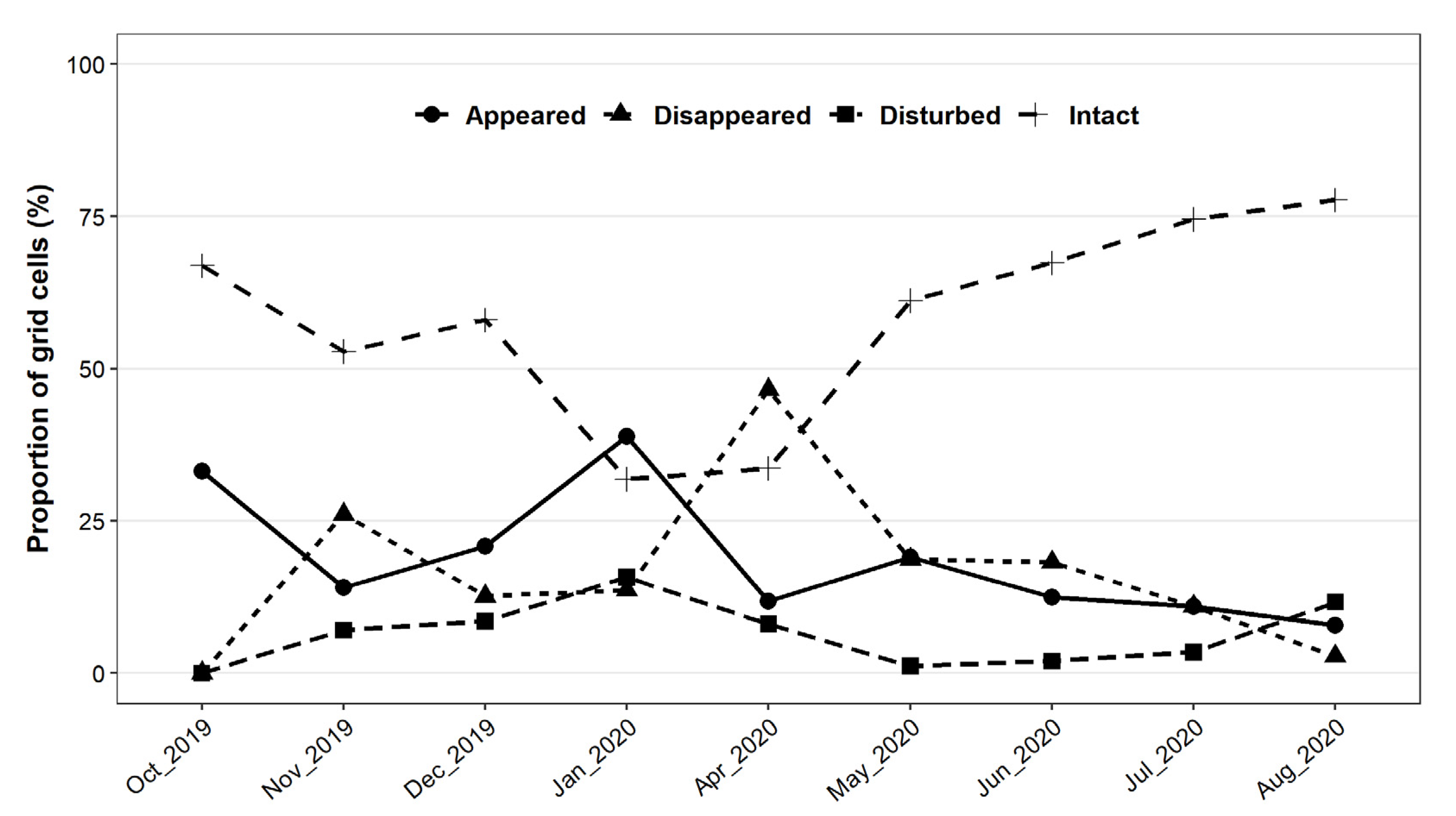
The temporal dynamics of rooting occurrence were described by the within-grid cell changes of wild boar disturbances noted between consecutive monthly assessments. These cells were categorized according to rooting intensity classifications, such as: (1) “Appeared”, if we found rooted soil surface within a grid cell, but there was no rooting there in the previous month; (2) “Disturbed”, if rooting was already found in the previous month in the cell and the area had been visibly disturbed again since then; (3) “Disappeared”, if no rooting was found in the grid cell, but it was rooted in the previous month; and (4) “Intact” if no rooting was observed during the survey or during the previous survey. The proportion of grid map cells within which rooting occurrence was observed were determined for each month.
Boar rooting preference for sedimentation and steepness was tested by the Chi-square goodness of fit test and Bonferroni confidence intervals [
26], comparing the proportions of all rooted patches in different sedimentation and steepness classes throughout the study against the proportion of the availability of those classes, based on the dataset of 649 sampling points along transects done to determine soil availability.
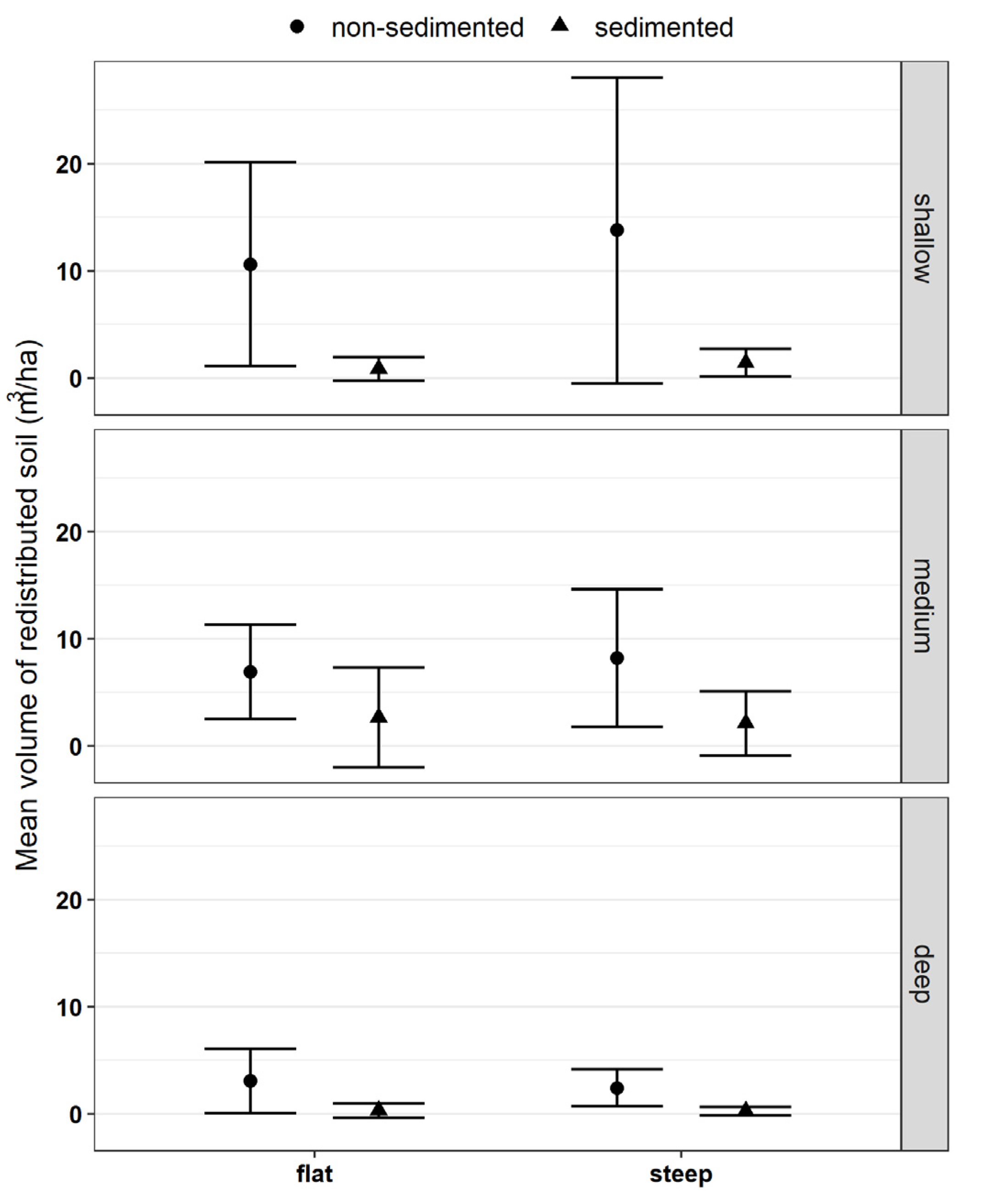
The amount of redistributed soil by rooting in different soils and steepness classes was also estimated for each sampling occasion (except for May due to data loss), where we considered the three-dimensional shape of the rooted depression as a spherical cap and calculated its volume by using the data on length and depth of rooting patches. The length (i.e., the diameter in our estimation) of the rooted soil patch was calculated as the distance between the “enter” and “exit” GPS points using the software QGIS 2.18.28 Las Palmas (QGIS.org, 2022. QGIS Geographic Information System. QGIS Association.
http://www.qgis.org) in HD72/EOV (EPSG: 23700 projected coordinate reference system for Hungary). The following formula was used for estimating the volume (V) of excavated soil:
where
h (height) is the depth of a rooted soil patch [m], and
r (radius) is half of the length of a rooted soil patch [m].
The direct correlation of soil penetration resistance with sedimentation and wild boar rooting (rooting–ring–control comparisons) were analyzed by Generalized Estimating Equations (GEE) in R [
27], with the “geepack” package [
28]. Since these compaction values were highly autocorrelated in similar depths, GEE was an appropriate method to handle issues related to any type of temporal or spatial autocorrelation [
29] by selecting the best working correlation structure by the QIC (quasi-likelihood under the independence model criterion) value of each equation.
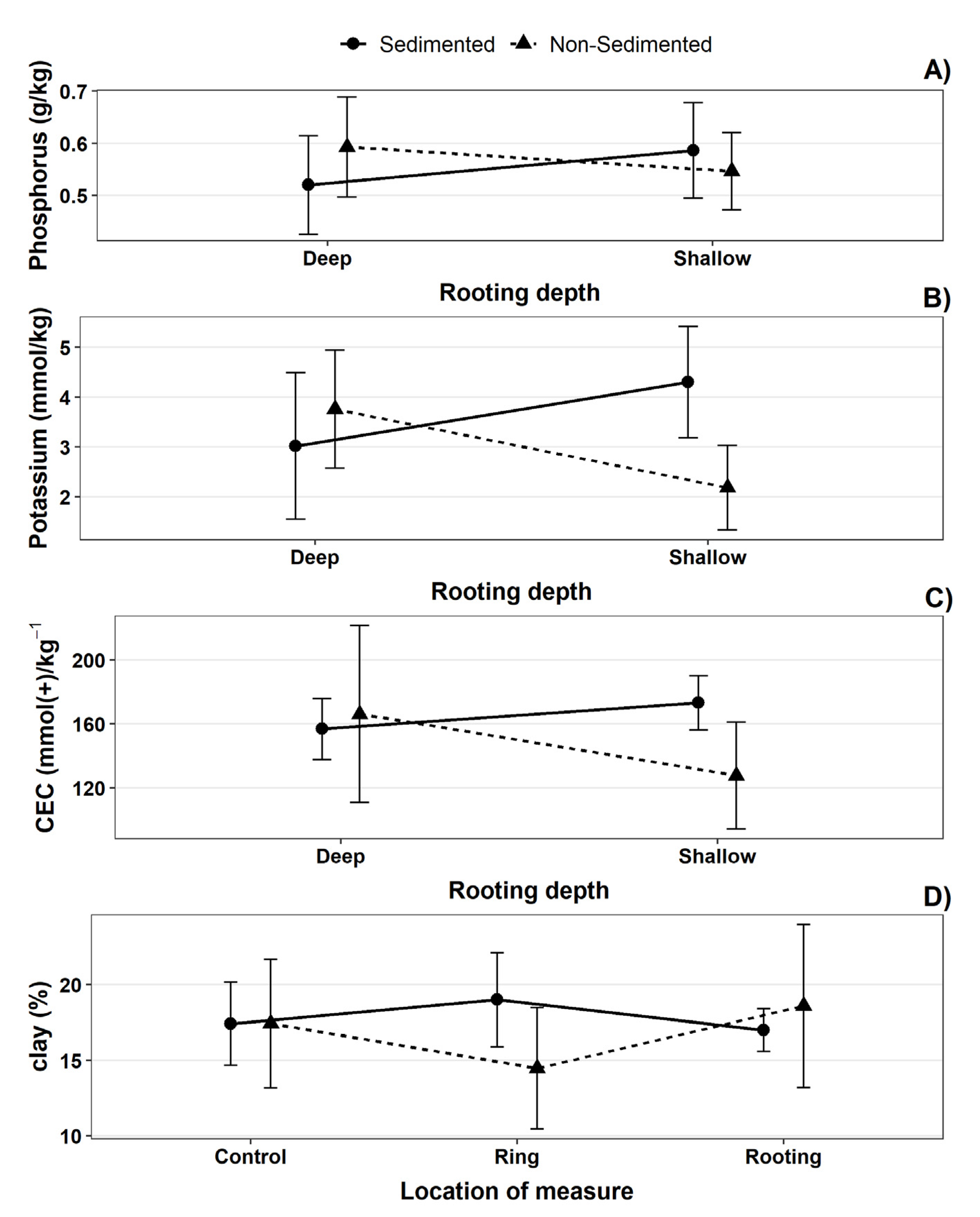
We implemented factorial MANOVA (Multivariate Analysis of Variance) in R to evaluate the effects of wild boar rooting and sedimentation on soil nutrient levels. We considered sedimentation, rooting depth and location of measurements (rooting—ring—control comparisons) as independent variables and NIR results (pH, Phosphorus, Potassium, TOC, CEC) as dependent variables excluding Nitrogen due to its high correlation with TOC (r = 0.88). All dependent variables were log-transformed. We used Pillai’s trace for test statistics in MANOVA.
4. Discussion
The impact of wild boar rooting on the soil was constant throughout the year. We found an increasing amount of rooting during the autumn and winter months and a gradual decrease in rooting activity in spring and summer. This can be due to the decreasing availability of green herbaceous biomass during winter and the need to search for acorns, seeds and roots below-ground. Therefore, wild boar foraging activity was variable across time and location but can be considered as a continuous disturbance factor for forest soil.
In our study, the area of soil disturbed by wild boar was proportionally low: 775 m
2/ha implies less than 1% of the total area is being affected and considering the shallowness of most of the rooting, its effects can be superseded by other factors. However, the volume of soil redistributed (on average 52.8 m
3/ha) might have long-term effects on soil characteristics [
3]. Accepting that wild boar rooting is a permanent disturbance in forest habitats, it becomes clear that the spatially and temporally variable rooted surfaces will cover the largest part of the ground within a relatively short period, thus being a fundamental driver of soil formation processes. Our results are comparable to those reported by Kotanen [
30], who studied the effects of invasive wild pigs in California and stated that the soil was overturned by this species at a 7.4% rate annually, following a seasonal pattern similar to our findings, i.e., more pronounced from mid-autumn to spring. Other authors report more significant disturbances related to soil redistributed by wild boar rooting in the introduced range of the species, where those ecosystems could be more resilient to its effects. Cooray and Mueller-Dombois [
31] found that wild pig digging could disturb up to 30% of the forest floor in Hawaii, whereas Singer [
32] described a plant cover reduction of 87%, with a 36% reduction in litter horizon depth due to wild pig-related disturbance in forested areas in North Carolina and Tennessee.
In Hungary, Sütő et al. [
17] found wild boar rooting in 42% of their study area in the outskirts of an urban area; meanwhile, in a near-natural dry oak forest, Sütő et al. [
20] measured an average proportion of the disturbed 20 × 20 m cells to be 28.11 ± 22.01% (ranging between 0 and 80.31%) throughout a three year study. There, the authors explained the high proportion of disturbances by referring to the vegetation composition and food availability both in the city outskirts and in the dry oak forest, similar to the results reported by Wilson [
33], who found a direct relationship between the distance to woodland and the reported disturbance events by a wild pig in west Dorset, UK. Further research in Hungary, by Fehér et al. [
34], registered that only 6% of the sampled forested areas were rooted during the summer, and the majority of rooting was observed in oak forests. In the Babat Valley, we revealed that the spatial variability in rooting distribution is likely due to the sedimentation processes, which were absent in the other study sites. However, local vegetation characteristics can also have an additional influence, but this requires further research.
The forested area in our research included a large proportion of non-sedimented soil, however, a significant amount of wild boar rooting was found in sedimented soil, even if it was not preferred. It is possibly due to the less-compacted nature of the sedimented particles, which makes it easier for boars to dig in the soil. Further analyses on the causes and impact of this rooting on sedimentation and runoff should be done since soil macrofauna and vegetation cover were already scarce in this type of soil. Additionally, further studies are needed to conclude whether rooting has different effects on macrofauna and vegetation cover in sedimented and non-sedimented areas, respectively.
These relationships could be quite complex, and wild pig disturbances can have the opposite effect, depending on habitat characteristics. Mohr et al. [
19] studied the impact of ungulates in enclosures on a low mountain range in Germany characterized by steep slopes, and found that although macrofauna was more abundant in the enclosures, the overall soil characteristics were better on the areas where ungulates were present. Another example comes from Strauch et al. [
9], who examined the independent and interactive effects of
Psidium cattleianum (a highly invasive tree in tropical islands) and wild pigs on runoff amount and soil erosion rates. They reported that wild pigs were less active in more invaded areas, but on those on which it was present, the resulting impact was a reduced canopy cover with consequently higher runoff volumes and soil erosion rates, and even a negative effect on native plant species. Similar results were reported by Tierney and Cushman [
35], who compared the effect of wild pigs on native and invasive plant species in a coastal grassland in northern California. They found that native species recover more slowly than invasive ones after a disturbance event. The presence of invasive species (e.g.,
Robinia pseudoacacia,
Asclepias syriaca,
Celtis occidentalis,
Acer negundo, etc.) in the Babat Valley can be analyzed in further studies to assess the impact of wild boar rooting on the vegetation cover.
Our main result shows that the rooting behavior had no significant impact on the measured soil characteristics, being overridden by the soil type, and other landscape features like microtopography. We differentiated between depths of rooting, given that shallower ditches occupy a larger area, whereas deeper excavations can excavate higher volumes of soil. Therefore, the impact could be significant from both perspectives. Nevertheless, we found that only soil compaction was altered by the rooting depth, due to the closeness of the C horizon in deeper ditches.
Our results indicate that, although the wild boar is considered a high-impact species, changes in pH, TOC and CEC are mainly related to the sedimentation processes in the forest, and not to wild boar rooting activity, which rejects the hypothesis of wild boar rooting having a significant impact on soil characteristics. It can be argued that wild boar rooting has a higher impact on the geomorphology of the soil, whereas the presence of other materials (such as sediments) is the main factor to consider while evaluating changes in a soil’s composition. These results are similar to those reported by Frank et al. [
36], who examined the impact of herds of native migratory ungulates in variable topo-edaphic conditions in the Yellowstone National Park grasslands, finding that soil turnover by herbivores might cause a long-term impact on the soil Carbon content, although topography plays the primary role on this characteristic. It is worth noting that although Frank et al. [
36] studied a grassland, the presence of slopes was considered while evaluating the gradient change in the C content, resulting in an increased amount of biomass at the bottom of the slope, which might be different from the Babat Valley, given the sediment presence at the bottom of the valley. It is also worth noting that in the Yellowstone National Park there are herds of herbivores moving together, whereas, in our study area of a deciduous forest, such movements are related to smaller family groups of wild boars or solitary individuals.
Related but quite distinct (from geographical and also from research question point of view) studies are from Hawaii, where native and non-native forests were compared [
9] and one of the main findings was that wild pig caused reduced canopy cover and increased runoff volumes and soil erosion rates in the native forest, while the non-native forest had higher stem density and lower soil erosion and runoff rates. This relation is well-known; however, in our research area the canopy cover is almost 100% and stem density could only be increased by the exclusion of the wild boar that would lead to a rather philosophical question of how a harsh intervention such as the exclusion of wild boar would relate to natural processes. However, this would be another article.
Nevertheless, our results show that wild boar rooting does not have a significant effect on soil penetration resistance or soil composition and that any changes in those soil properties are mainly enhanced by sedimentation or runoff to be observable. Furthermore, the depressions in the slopes of the forest (including the wild boar rooted areas) can inhibit sediment from moving forward down the valley, decreasing the speed of runoff, and enhancing the slow infiltration of accumulated water. Warren et al. [
5] analyzed the effect of livestock on soil hydrologic characteristics, under intensive rotation grazing management, and found a significant impact due to trampling on the soil; namely a lower infiltration rate (i.e., higher compaction) and higher sediment production. On the other hand, Frank et al. [
36] applied a similar amount of pressure on the soil resulting in different outcomes that were based not on the actions of the herbivores but on the characteristics and land management of the area. Analyzing those results, it is possible to assess that land management and use have a significant influence on the impact caused by ungulates on the soil. Therefore, in our study, it is the forest composition and the characteristics of the terrain that might influence the overall changes in the runoff and infiltration. Wild boar rooting can be, at least partly, beneficial in slowing down runoff and trapping sediment. Although more substantial natural or artificial dams are needed to address the current sedimentation problems in the Babat Valley.
Wild boars are regarded as being environmental engineers and the cause of continuous disturbance to soil. Assessing the importance of abiotic characteristics to boars can give a more accurate picture of the nature of these causes and extent of landscape transformations related to other disturbance sources. Further analyses can explore additional abiotic factors, including the impact of vegetation cover, to determine whether wild boar rooting mitigates soil runoff and sedimentation process by trapping washed soil particles or enhancing soil properties through soil disturbance and loosening particles, causing it to be washed down. Although it has improved recently, research on wild boar rooting and its impacts has been predominantly focused on agricultural lands and grasslands, without considering natural ecosystems with limited anthropogenic intervention. In this sense, this study contributes to a better understanding of these dynamics in deciduous forests in the interest of conservation.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}