1. Introduction
Human activities, such as slash-and-burn agriculture and logging, have severely damaged tropical rainforest ecosystems and biodiversity [
1,
2]. Some restored forests look superficially like mature rainforests, yet are still greatly lacking biodiversity, which could take centuries to fully recover [
3]. In recent years, a plethora of research has been conducted on the restoration of aboveground plant diversity and underground microorganisms. Studies have found that the plant species composition [
4,
5] and soil microbial communities [
6,
7] of secondary forests were significantly different from that of old growth forest. Han et al. [
8] showed that logging disturbance significantly changed the ecological strategy spectrum, and that the strategy number increased with logging intensity. Chen et al. [
7] found that the ecological network structure of the microbial community in landscapes with selective logging was more complex than those with clear cutting. Despite such work, there are few studies on interkingdom plant–microbial ecological networks during the restoration period of secondary forests.
Plants and their associated diverse soil microorganisms, such as bacteria, fungi, archaea and protists, as well as microbe–microbe pairs, interact with each other (competitive, mutualistic, neutral, commensal) and form complex relationships [
9,
10]. Microorganisms can protect host plants from the invasion of pathogenic microorganisms [
11], boost the growth of host plants [
12] and help host plants adapt to salt, drought and other adverse environments [
13,
14]. In exchange, the large amount of sugars, amino acids and organic acids secreted by plants in the rhizosphere are nutrient sources for soil microorganisms [
15]. The structure and function of soil microbial communities is generated by the complex interactions among host plants, the surrounding plant community, the soil environment and microbial co-associations with the microbiome [
9]. Interactions between these species change with the environment [
16]. In this study, we took samples in non-rhizosphere soil loci. The composition of the rhizosphere microbiome is extremely different from the structure of the native soil microbiome [
17], but the plant rhizosphere-specific fungal mycelia may extend into the bulk soil, affecting the non-rhizosphere microbial community [
18]. Understanding the relationships between aboveground plants and belowground microorganisms under different disturbance types in natural ecosystems will aid in better understanding of the ecosystem functions of ecological networks.
With the rapid development of high-throughput technologies, microbiologists increasingly recognize that species interactions are more important than species richness in complex ecosystems [
19,
20]. These interactions can be visualized as a series of ecological networks in which species are directly or indirectly linked [
19]. The ecological network of microbial communities has been extensively studied. For example, Deng et al. [
21] constructed molecular ecological networks using Random Matrix Theory methods to provide better understanding of network interactions in microbial communities and their responses to environmental changes. Ecosystem functions of ecological networks can be identified based on network topology [
20]. In order to better understand the associations between aboveground plants and belowground microorganisms, Feng et al. [
22] proposed a workflow to construct interdomain ecological networks between plants and microbes. Chen et al. [
23] demonstrated that interdomain ecological networks between plants and microbes can reflect changes in tree composition and soil nutrients during forest restoration.
Hainan Island hosts the largest tropical forest in China and is one of the world’s biodiversity hotspots. Due to the interference of human activities, such as slash-and-burn agriculture and logging, a variety of secondary forests have been formed during the natural recovery process, making the composition of plant species significantly different from that of old growth forests [
5], which provides an ideal experimental platform for our research. In this study, using old growth forest (OG) as a control, three types of disturbed forest land (slash-and-burn (SB), clear cutting (CC) and selective cutting (SL)) were used to construct ecological networks between bacteria and fungi as well as plants and microbes to address the response of soil bacterial–fungal and plant–microbe ecological networks to different disturbance types.
4. Discussion
Molecular ecological network analysis simplifies the complex process of analyzing large amounts of data and aids in better understanding the relationships between microorganisms. The stability of a molecular network is closely related to the complexity of the network—the more complex the microbial community, the more stable the microbial community [
35,
36]. Diversity of interaction types (moderate mixture of antagonistic and mutualistic interactions) can stabilize population dynamics [
35], indicating that soil microbial community stability is higher in the selective cutting and clear cutting stands than in old growth forest. In contrast, the stability of the soil microbial community was lower in the slash and burn stands than in old growth forest. In this study, the positive proportion of bacterial–fungal networks in slash-and-burn stands was as high as 93%, indicating an increased mutualism between bacteria and fungi. The increased proportion of negative correlation in selective cutting stands indicates an increase in antagonistic interactions between bacteria and fungi, which reflect the competition between soil bacteria and fungi for limited resources within a unique environmental niche [
36]. Modularity prevailed in the molecular ecological network [
21]. A network module is a group of species that are highly associated with each other, but had much fewer associations with other modules species [
21], which could be perceived as niches [
37]. In the bacterial–fungal networks, a higher modularity was observed in the clear cutting and selective cutting stands, suggesting niche differentiation (i.e., the microbial community segregated into finer niches and functional units). Such niche differentiation is an important factor in shaping interactive species and maintaining biodiversity at different scales [
38].
Different from bacterial–fungal networks, the plant–bacteria and plant–fungi ecological networks of the clear cutting stand was relatively simple. Clear cutting drastically changes the stand structure and removes the forest canopy [
39], leading to the plant community’s slow recovery, suggesting that plant–microbe associations lag behind bacterial–fungal associations. Coexistence of nested and modularity properties is crucial for the resilience and structural stability of ecosystem communities [
40]. Nestedness depends on the neutral theory of community construction, while modularity depends on local adaptation and competition [
41]. Studies have found that modularity is strong in trophic networks, while nested structures are strong in mutualistic networks [
42]. Low nestedness is very important for resilience and structural stability of ecological communities [
43]. Non-nested structures have been observed both in plant–bacteria and plant–fungi ecological networks, which may be due to the both plant–bacteria and plant–fungi ecological network belonging to trophic networks [
42], and this may be a typical feature of interdomain ecological networks [
22], indicating that species preferences are divided to avoid competition and thus favor system-wide resource allocation [
44].
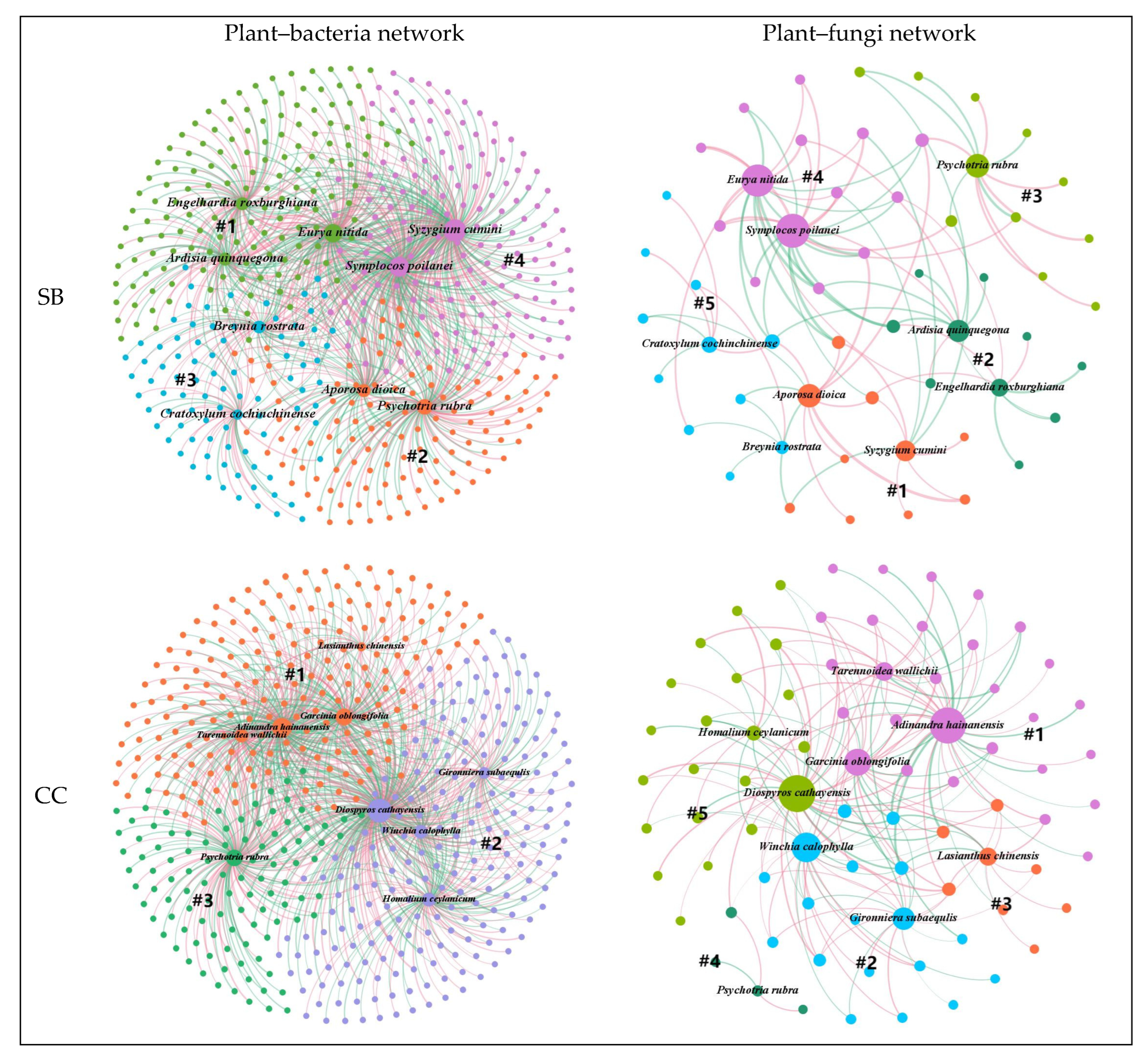
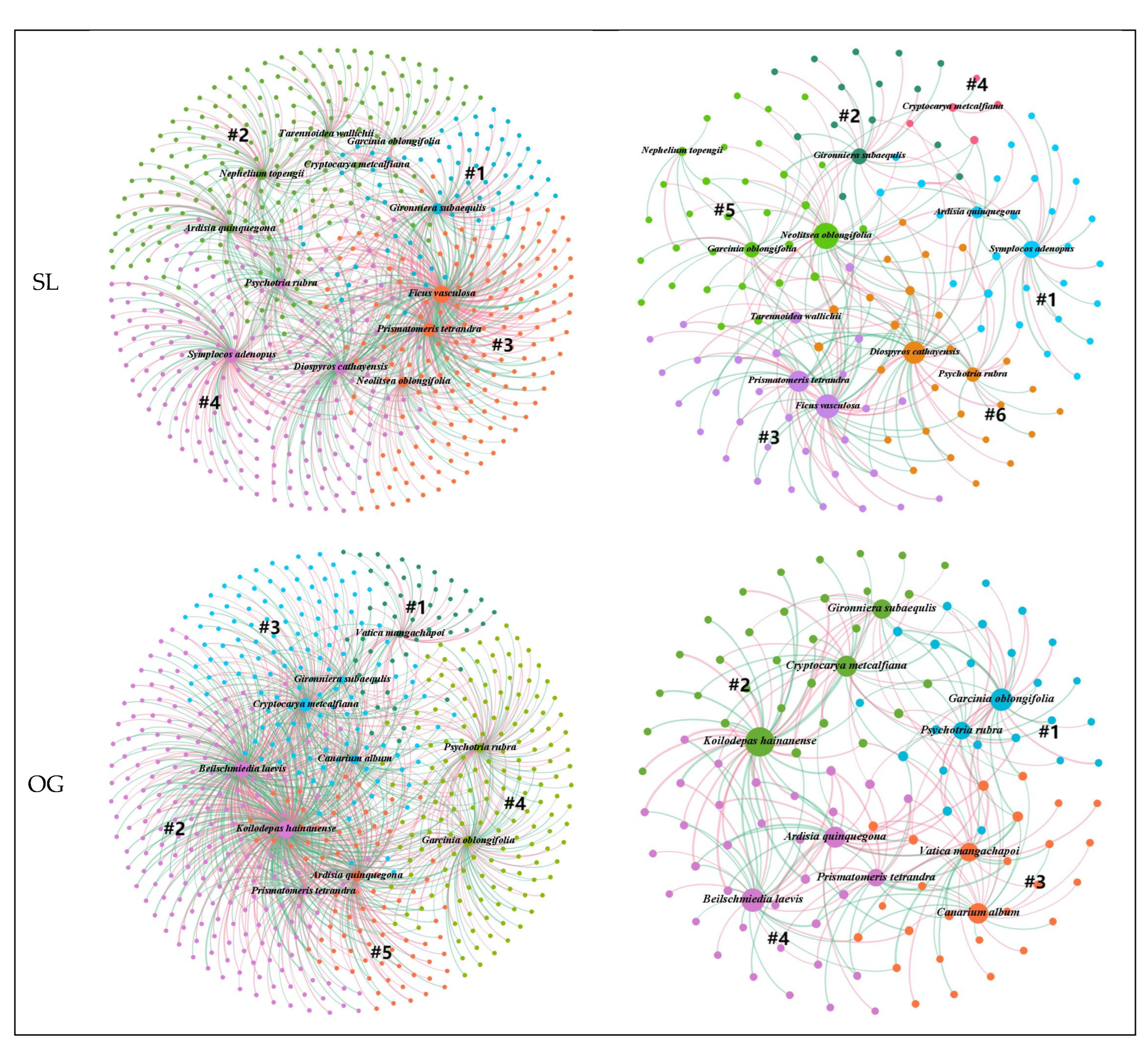
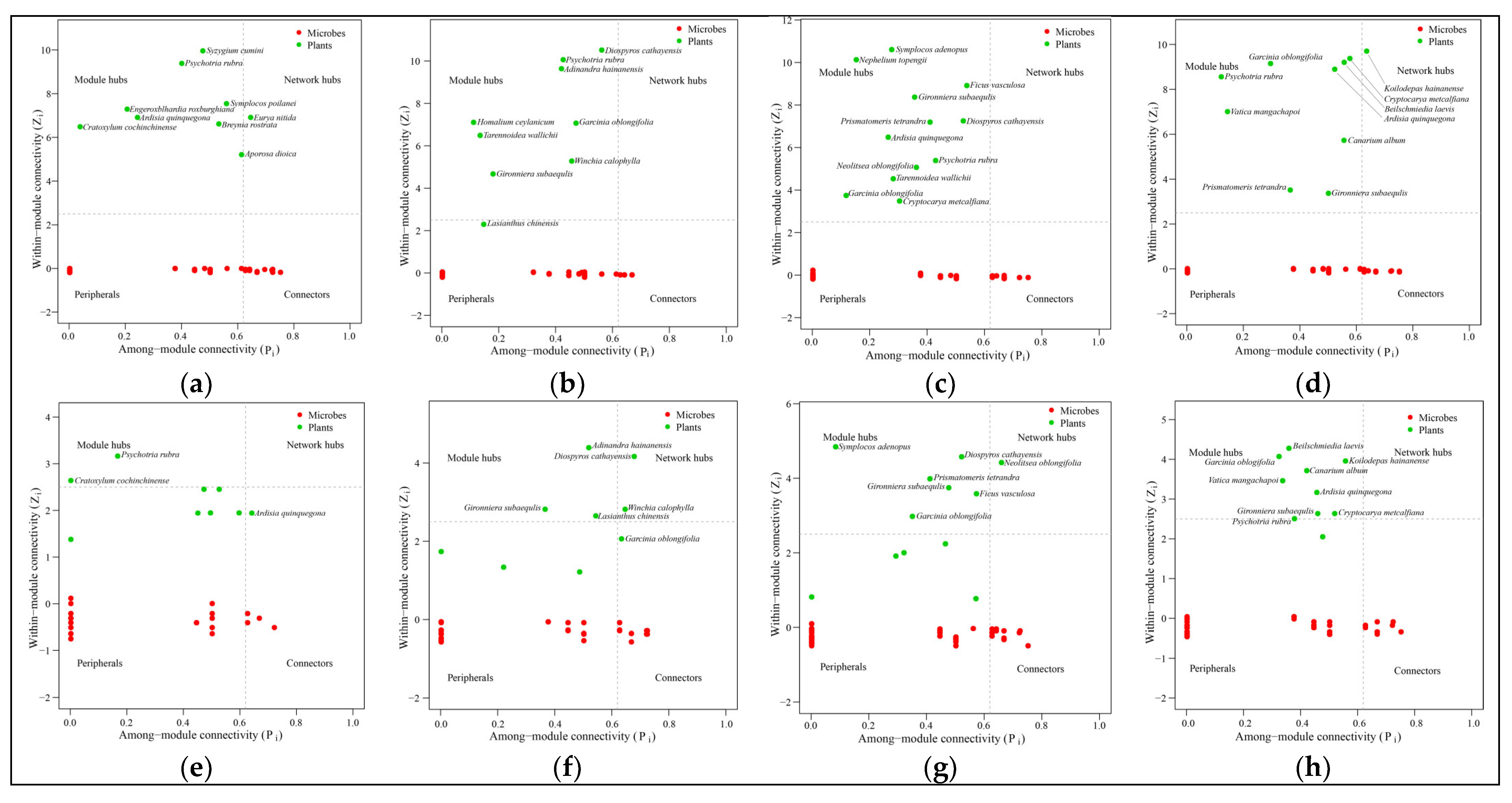
Ecological networks are usually divided into modules of closely interacting species, which are connected by some supergeneralist that serves as a hub or connector species [
45]. Most microorganisms are within plant-related modules rather than across modules, suggesting a specialized association between plants and microorganisms [
22]. A species defined as a hub is highly associated with other species and has a strong influence on community structures [
21]. Removing the hub (keystone species/taxa) can cause drastic changes in community structure and function [
46]. The loss of a keystone species leads to a loss of function, and in the case of functional redundancy, observed differences in the taxonomic composition of microbial communities do not imply a loss of function [
47]. In this study, different keystone species were found in four forest stands, suggesting that there may be functional redundancy, leading to an alternative stable state [
46]. The significant reduction in plant species and the change in dominant species will drastically change the relationship between plants and microorganisms [
48]. Some different plant species are associated with different microbial genera in the four different forest stands, suggesting that plants adapt dynamically by regulating their microbiomes. In addition, it appears that keystone species can use differing strategies to shape their microbiomes to their advantage [
46,
47]. Different plant species had more preferential associations with specific fungal genera than bacterial genera, suggesting that bacteria may also have functional redundancy.
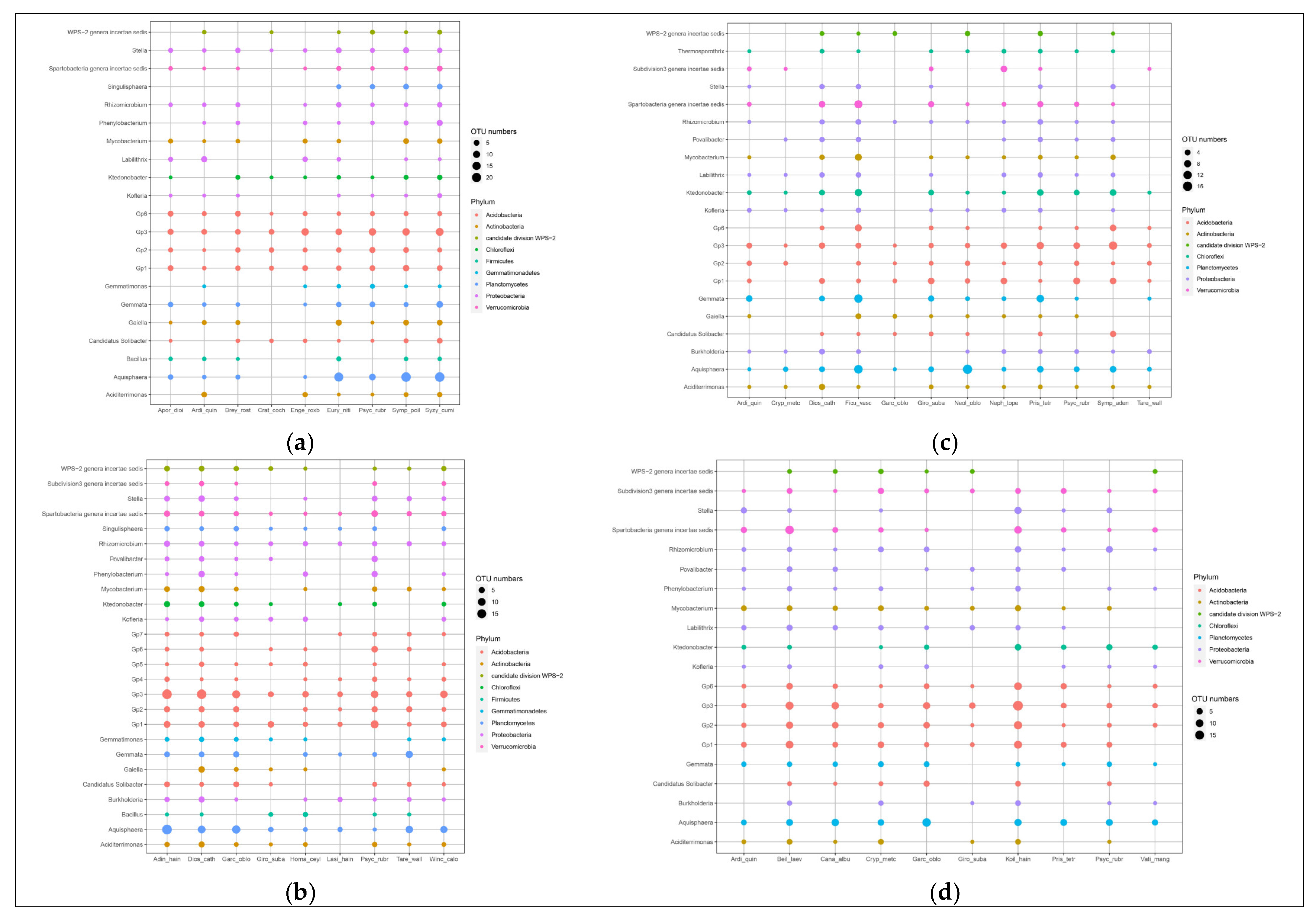
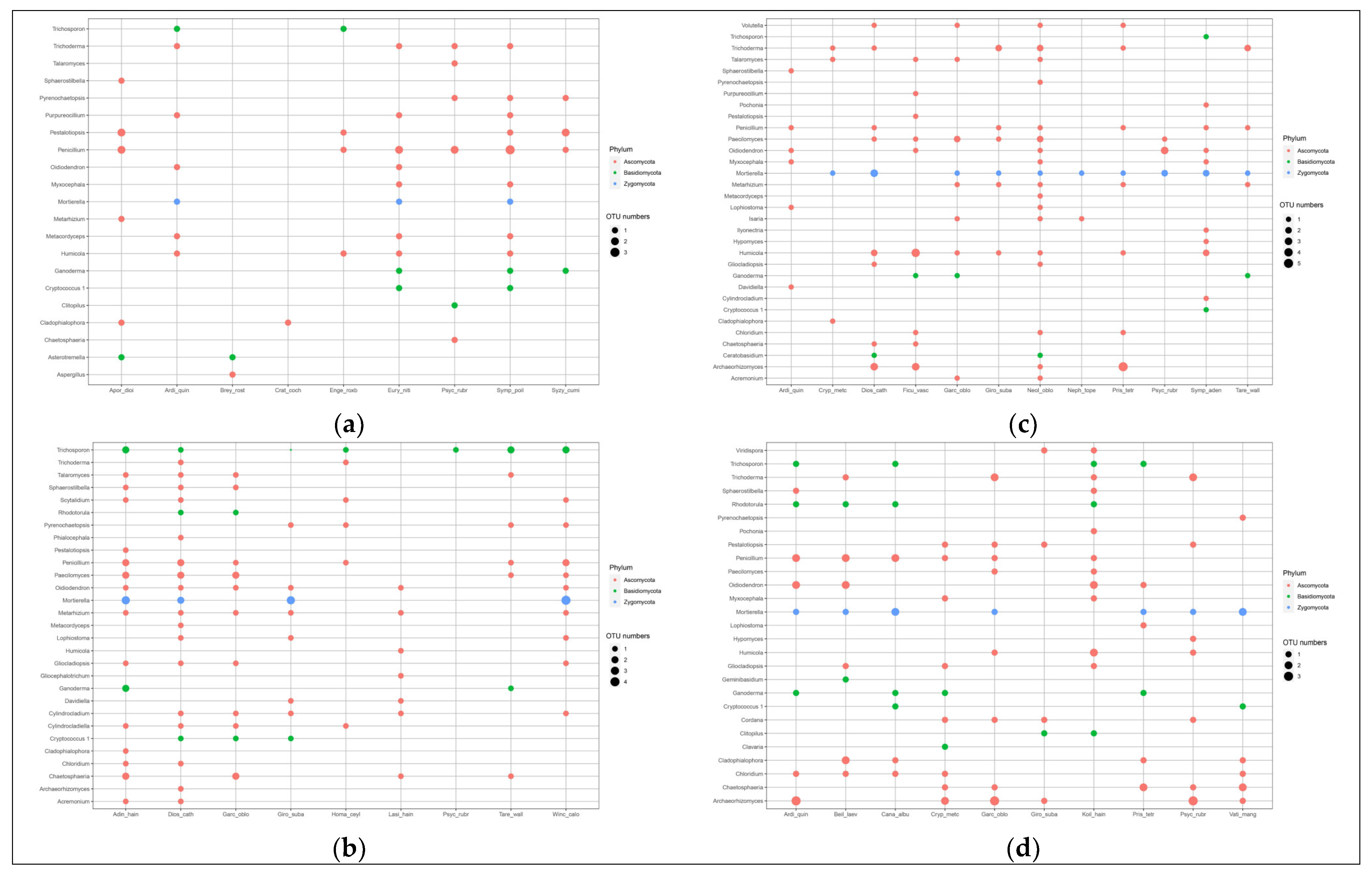
The genera of bacteria associated with plants belonged primarily to Proteobacteria and Acidobacteria, while the genera of fungi mainly belonged to Ascomycota, which are dominant phyla in forest soils [
49,
50,
51], indicating that these taxa are widely involved in ecological processes. The relationship between different plants and specific fungal communities are stronger than that between different plants and bacterial communities, which may be because the soil bacterial community structure is influenced by time (succession), and the characteristics of plant species and functional groups are the driving factors affecting soil fungal community structure, which can explain the large proportion of changes in fungal community composition [
52]. Urbanová et al. [
53] showed that 35–37% of fungal OTUs showed a preference for specific plant species, while 80% of bacterial OTUs shared multiple plant species, which was consistent with our results, indicating that different plants have a preference for fungi over bacteria. Ectomycorrhizal fungi OTUs usually show strong host preference [
53]. Chen et al. [
23] found that the dominant plant species (Cyclobalanopsis, Lithocarpus and Castanopsis) characterized by ectomycorrhizal fungi increased after clear cutting and selective cutting, indicating that the accumulation of mycorrhizal fungi are beneficial to plant community restoration. Genre et al. [
54] showed that mycorrhizal is one of the most important biosphere interactions, providing host plants with mineral nutrients, such as phosphorus and nitrogen, important nutrients to consider for forest management after forest fire disturbance and restoration of mining areas. However, in this study, the genera of fungi associated with plants mainly belonged to Ascomycota, which may be because most ectomycorrhizal plants associate with many unrelated ectomycorrhizal fungi, and many unrelated ectomycorrhizal fungi associate with different plant hosts [
55]. For example, Morris et al. [
56] found that Ascomycota were more frequent and diverse on
Quercus douglasii, which can increases the habitats in which the plants can live, because seedlings dispersing will not be limited in new settings, and may find compatible unrelated ectomycorrhizal fungi [
57].




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




