
Suitability of Turkey Oak, European Beech, and Hornbeam to Gypsy Moth Feeding

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Host Plants
2.3. Leaf Characteristic
2.4. Preference Test
2.5. Performance Test
2.6. Statistical Analysis
3. Results
3.1. Leaf Characteristics
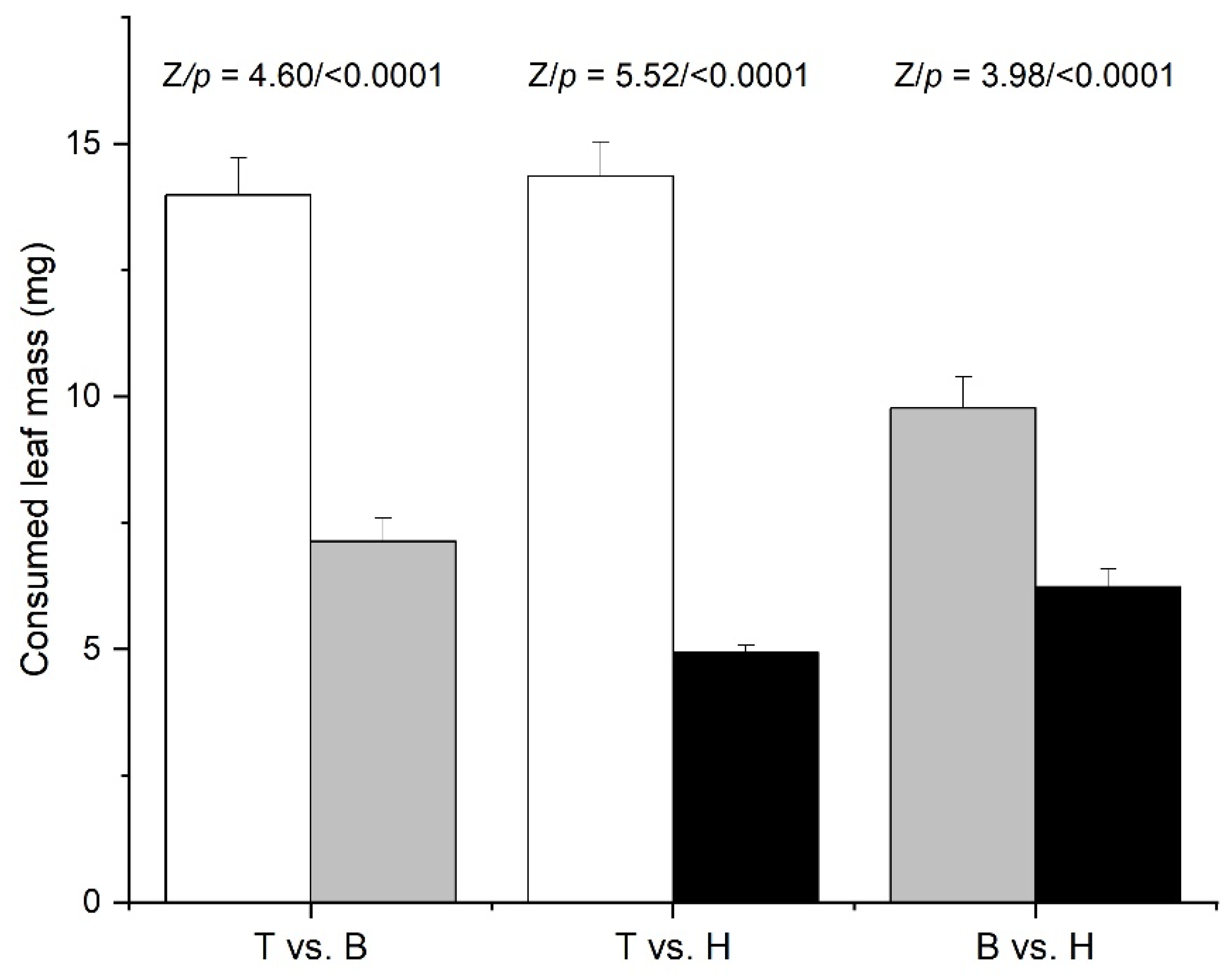
3.2. Larval Preference
3.3. Larval Performance
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. Insect-Plant Biology, 2nd ed.; Oxford University Press: Oxford, UK, 2005; p. 421. [Google Scholar]
- Despres, L.; David, J.P.; Gallet, C. The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol. Evol. 2007, 22, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Braga, M.P.; Janz, N. Host repertoires and changing insect–plant interactions. Ecol. Entomol. 2021, 46, 1241–1253. [Google Scholar] [CrossRef]
- Galway, K.E.; Duncan, R.P.; Syrett, P.; Emberson, R.M.; Sheppard, A.W. Insect Performance and Host-Plant Stress: A Review from a Biological Control Perspective. In Proceedings of the XI International Symposium on Biological Control of Weeds, Tifton, GA, USA, 27 April–2 May 2003. [Google Scholar]
- Andrew, N.R.; Roberts, I.R.; Hill, S.J. Insect herbivory along environmental gradients. Open J. Ecol. 2012, 2, 202–213. [Google Scholar] [CrossRef] [Green Version]
- Jactel, H.; Koricheva, J.; Castagneyrol, B. Responses of forest insect pests to climate change: Not so simple. Curr. Opin. Insect Sci. 2019, 35, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Łukowski, A.; Giertych, M.J.; Adamczyk, D.; Mąderek, E.; Karolewski, P. Preference and performance of the pine-tree lappet Dendrolimus pini on various pine species. Forests 2021, 12, 1261. [Google Scholar] [CrossRef]
- Haynes, K.J.; Liebhold, A.M.; Lefcheck, J.S.; Morin, R.S.; Wang, G. Climate affects the outbreaks of a forest defoliator indirectly through its tree hosts. Oecologia 2022, 198, 407–418. [Google Scholar] [CrossRef] [PubMed]
- de Rigo, D.; Enescu, C.M.; Houston Durrant, T.; Caudullo, G. Quercus cerris in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. e01b479+. [Google Scholar]
- Houston Durrant, T.; de Rigo, D.; Caudullo, G. Fagus sylvatica and other beeches in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. e012b90+. [Google Scholar]
- Dittmar, C.; Zech, W.; Elling, W. Growth variations of common beech (Fagus sylvatica L.) under different climatic and environmental conditions in Europe—a dendroecological study. For. Ecol. Manag. 2003, 173, 63–78. [Google Scholar] [CrossRef]
- Bresson, C.C.; Vitasse, Y.; Kremer, A.; Delzon, S. To what extent is altitudinal variation of functional traits driven by genetic adaptation in European oak and beech? Tree Physiol. 2011, 31, 1164–1174. [Google Scholar] [CrossRef] [Green Version]
- Allevato, E.; Saulino, L.; Cesarano, G.; Chirico, G.B.; D'Urso, G.; Bolognesi, S.F.; Rita, A.; Rossi, S.; Saracino, A.; Bonanomi, G. Canopy damage by spring frost in European beech along the Apennines: Effect of latitude, altitude and aspect. Remote Sens. Environ. 2019, 225, 431–440. [Google Scholar] [CrossRef]
- Sikkema, R.; Caudullo, G.; de Rigo, D. Carpinus betulus in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of European Union: Luxembourg, 2016; p. e01d8cf+. [Google Scholar]
- Kramer, P.J. Drought tolerance and water use efficiency. In Water Relations of Plants; Kramer, P.J., Ed.; Academic Press: Cambridge, MA, USA, 1983; pp. 390–415. [Google Scholar]
- Tognetti, R.; Cherubini, P.; Marchi, S.; Raschi, A. Leaf traits and tree rings suggest different water-use and carbon assimilation strategies by two co-occurring Quercus species in a Mediterranean mixed-forest stand in Tuscany, Italy. Tree Physiol. 2007, 27, 1741–1751. [Google Scholar] [CrossRef] [Green Version]
- Köcher, P.; Gebauer, T.; Horna, V.; Leuschner, C. Leaf water status and stem xylem flux in relation to soil drought in five temperate broad-leaved tree species with contrasting water use strategies. Ann. For. Sci. 2009, 66, 101. [Google Scholar] [CrossRef] [Green Version]
- Pšidová, E.; Ditmarová, Ľ.; Jamnická, G.; Kurjak, D.; Majerová, J.; Czajkowski, T.; Bolte, A. Photosynthetic response of beech seedlings of different origin to water deficit. Photosynthetica 2015, 53, 187–194. [Google Scholar] [CrossRef]
- van der Maaten-Theunissen, M.; Bümmerstede, H.; Iwanowski, J.; Scharnweber, T.; Wilmking, M.; van der Maaten, E. Drought sensitivity of beech on a shallow chalk soil in northeastern Germany—A comparative study. For. Ecosyst. 2016, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Liebhold, A.M.; Gottschalk, K.W.; Muzika, R.M.; Montgomery, M.E.; Yong, R.; O’Day, K.; Kelley, B. Suitability of North American Tree Species to the Gypsy Moth: A Summary of Field and Laboratory Tests; USDA For. Serv. Gen. Tech. Rep. NE-211: Radnor, PA, USA, 1995; pp. 1–34. [Google Scholar]
- Lazarević, J.M.; Perić-Mataruga, V.D. Nutritive stress effects on growth and digestive physiology of Lymantria dispar larvae. Jugoslov. Med. Biohem. 2003, 22, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Hättenschwiler, S.; Schafellner, C. Gypsy moth feeding in the canopy of a CO2-enriched mature forest. Glob. Chang. Biol. 2004, 10, 1899–1908. [Google Scholar] [CrossRef]
- Milanović, S.; Lazarević, J.; Popović, Z.; Miletić, Z.; Kostić, M.; Radulović, Z.; Karadzić, D.; Vuleta, A. Preference and performance of the larvae of Lymantria dispar (Lepidoptera: Lymantriidae) on three species of European oaks. Eur. J. Entomol. 2014, 111, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Hlásny, T.; Turčáni, M. Insect pests as climate change driven disturbances in forest ecosystems. In Bioclimatology and Natural Hazards; Střelcová, K., Mátyás, C., Kleidon, A., Lapin, M., Matejka, F., Blaženec, M., Škvarenina, J., Holécy, J., Eds.; Springer: Berlin, Germany, 2009; pp. 165–178. [Google Scholar]
- Wittman, J.T.; Aukema, B.H. Foliage type and deprivation alters the movement behavior of late instar European gypsy moth Lymantria dispar (Lepidoptera: Erebidae). J. Insect Behav. 2019, 32, 24–37. [Google Scholar] [CrossRef]
- Lechowicz, M.J. Leaf Quality and the Host Preferences of Gypsy Moth in the Northern Deciduous Forest; USDA For. Serv. Gen. Tech. Rep. NE-85: Radnor, PA, USA, 1983; pp. 67–82. [Google Scholar]
- Milanović, S.D.; Popović, M.M.; Dobrosavljević, J.N.; Kostić, I.M.; Lazarević, J.M. Desperate times call for desperate measures: Short-term use of the common ash tree by gypsy moth larvae (Lepidoptera: Erebidae) under density and starvation stress. Arch. Biol. Sci. 2020, 72, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Milanović, S.; Mihajlović, L.; Karadžić, D.; Jankovsky, L.; Aleksić, P.; Janković-Tomanić, M.; Lazarević, J. Effects of pedunculate oak tree vitality on gypsy moth preference and performance. Arch. Biol. Sci. 2014, 66, 1659–1672. [Google Scholar] [CrossRef]
- Elkinton, J.S.; Liebhold, A.M. Population dynamics of gypsy moth in North America. Annu. Rev. Entomol. 1990, 35, 571–596. [Google Scholar] [CrossRef]
- Rossiter, M.C. Genetic and phenotypic variation in diet breadth in a generalist herbivore. Evol. Ecol. 1987, 1, 272–282. [Google Scholar] [CrossRef]
- Barbosa, P.; Martinat, P.; Waldvogel, M. Development, fecundity and survival of the herbivore Lymantria dispar and the number of plant species in its diet. Ecol. Entomol. 1986, 11, 1–6. [Google Scholar] [CrossRef]
- Barbosa, P.; Krischik, V.A. Influence of alkaloids on feeding preference of eastern deciduous forest trees by the gypsy moth Lymantria dispar. Am. Nat. 1987, 130, 53–69. [Google Scholar] [CrossRef]
- Osier, T.L.; Lindroth, R.L. Effects of genotype, nutrient availability, and defoliation on aspen phytochemistry and insect performance. J. Chem. Ecol. 2001, 27, 1289–1313. [Google Scholar] [CrossRef]
- Barbehenn, R.V.; Jaros, A.; Lee, G.; Mozola, C.; Weir, Q.; Salminen, J.P. Tree resistance to Lymantria dispar caterpillars: Importance and limitations of foliar tannin composition. Oecologia 2009, 159, 777–788. [Google Scholar] [CrossRef]
- Barbehenn, R.V.; Haugberg, N.; Kochmanski, J.; Menachem, B.; Miller, C. Physiological factors affecting the rapid decrease in protein assimilation efficiency by a caterpillar on newly-mature tree leaves. Physiol. Entomol. 2014, 39, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Milanović, S.; Janković-Tomanić, M.; Kostić, I.; Kostić, M.; Morina, F.; Živanović, B.; Lazarević, J. Behavioural and physiological plasticity of gypsy moth larvae to host plant switching. Entomol. Exp. Appl. 2016, 158, 152–162. [Google Scholar] [CrossRef]
- Rossiter, M.C. Environmentally-based maternal effects: A hidden force in insect population dynamics? Oecologia 1991, 87, 288–294. [Google Scholar] [CrossRef]
- Ponomarieva, V.V.; Plotnikova, T.A. Determination of the Content and Composition of Organic Matter in Peat Soils. Methods of the Determination of Humus Content and Composition in the Soils (Mineral and Peat); Dokuchaev Central Soil Science Museum: St. Petersburg, Russia, 1975. (In Russian) [Google Scholar]
- Rhee, K.C. Unit B1.2. Determination of total nitrogen. In Handbook of Food Analytical Chemistry, Pigments, Colorants, Flavors, Texture and Bioactive Food Components; Wrolstad, R.E., Acree, T.E., Decker, E.A., Penne, M.H., Reid, D.S., Schwartz, S.J., Shoemaker, C.F., Smith, D., Sporns, P., Eds.; John Wiley and Sons Inc.: New York, NY, USA, 2001; pp. 95–113. [Google Scholar]
- Foss, L.K.; Rieske, L.K. Species-specific differences in oak foliage affect preference and performance of gypsy moth caterpillars. Entomol. Exp. Appl. 2003, 108, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Waldbauer, G.P. The consumption and utilization of food by insects. Adv. Insect Physiol. 1968, 5, 229–288. [Google Scholar]
- Farrar, R.R.; Barbour, J.D.; Kendedy, G.G. Quantifying food consumption and growth in insects. Ann. Entomol. Soc. Am. 1989, 82, 593–598. [Google Scholar] [CrossRef]
- Raubenheimer, D.; Simpson, S.J. Analysis of covariance:an alternative to nutritional indicies. Entomol. Exp. Appl. 1992, 62, 221–231. [Google Scholar] [CrossRef]
- Horton, D.R.; Redak, R.A. Further comments on analysisof covariance in insect dietary studies. Entomol. Exp. Appl. 1993, 69, 263–275. [Google Scholar] [CrossRef]
- Haägele, B.F.; Rowell-Rahier, M. Dietary mixing in three generalist herbivores: Nutrient complementation or toxin dilution? Oecologia 1999, 119, 521–533. [Google Scholar]
- Carrasco, D.; Larsson, M.C.; Anderson, P. Insect host plant selection in complex environments. Curr. Opin. Insect Sci. 2015, 8, 1–7. [Google Scholar] [CrossRef]
- Quintero, C.; Bowers, M.D. Plant and herbivore ontogeny interact to shape the preference, performance and chemical defense of a specialist herbivore. Oecologia 2018, 187, 401–412. [Google Scholar] [CrossRef]
- Rieske, L.K.; Rhoades, C.C.; Miller, S.P. Foliar chemistry and gypsy moth, Lymantria dispar (L.), herbivory on pure American chestnut, Castanea dentata (Fam: Fagaceae), and a disease-resistant hybrid. Environ. Entomol. 2003, 32, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Couture, J.J.; Meehan, T.D.; Lindroth, R.L. Atmospheric change alters foliar quality of host trees and performance of two outbreak insect species. Oecologia 2012, 168, 863–876. [Google Scholar] [CrossRef]
- Morrow, C.J.; Jaeger, S.J.; Lindroth, R.L. Intraspecific variation in plant economic traits predicts trembling aspen resistance to a generalist insect herbivore. Oecologia 2022, 199, 119–128. [Google Scholar] [CrossRef]
- Roden, D.B.; Mattson, W.J. Rapid induced resistance and host species effects on gypsy moth, Lymantria dispar (L.): Implications for outbreaks on three tree species in the boreal forest. For. Ecol. Manag. 2008, 255, 1868–1873. [Google Scholar] [CrossRef]
- Johnson, D.M.; Liebhold, A.M.; Bjørnstad, O.N. Geographical variation in the periodicity of gypsy moth outbreaks. Ecography 2006, 29, 367–374. [Google Scholar] [CrossRef]
- Lazarević, J.; Perić-Mataruga, V.; Ivanović, J.; Andjelković, M. Host plant effects on the genetic variation and correlations in the individual performance of the gypsy moth. Funct. Ecol. 1998, 12, 141–148. [Google Scholar] [CrossRef]
- Clancy, K.M.; Price, P.W. Rapid herbivore growth enhances enemy attack: Sublethal plant defenses remain a paradox. Ecology 1987, 68, 733–737. [Google Scholar] [CrossRef]
- Barbosa, P. Host plant exploitation by the gypsy moth, Lymantria dispar. Entomol. Exp. Appl. 1978, 24, 228–237. [Google Scholar] [CrossRef]
- Montgomery, M.E. Variation in the Suitability of Tree Species for the Gypsy Moth; USDA For. Serv. Gen. Tech. Rep. NE-146: Radnor, PA, USA, 1991; pp. 1–13. [Google Scholar]
- Schopf, A.; Hoch, G.; Klaus, A.; Novotny, J.; Zúbrik, M.; Schafellner, C. Influence of food quality of two oak species on the development and growth of gypsy moth larvae. In Physiology and Genetics of Tree Phytophage Interactions; Lieutier, F., Mattson, W.J., Wagner, M.R., Eds.; Les Colloques INRA 90: Gujan, France, 1999; pp. 231–247. [Google Scholar]
- Yanar, O.; Bilgener, M.; Altun, N. The effects of the water, protein and polyphenolic contents of four host plant species on the development and egg yield of female larvae of gypsy moth (Lymantria dispar). Int. J. Nat. Eng. Sci. 2007, 1, 93–98. [Google Scholar]
- Zúbrik, M.; Kunca, A.; Kulfan, J.; Rell, S.; Nikolov, C.; Galko, J.; Vakula, J.; Gubka, A.; Leontovyč, R.; Konôpka, B.; et al. Occurrence of gypsy moth (L.) in the Slovak Republic and its outbreaks during 1945–2020. Cent. Eur. For. J. 2021, 67, 55–71. [Google Scholar] [CrossRef]
- Schafellner, C.; Schopf, A. The greenhouse diet: Gypsy moth performance in a CO2-enriched world. Mitt. Dtsch. Ges. Allg. Angew. Entomol. 2008, 16, 71–76. [Google Scholar]
- Barbehenn, R.V.; Niewiadomski, J.; Pecci, C.; Salminen, J.P. Physiological benefits of feeding in the spring by Lymantria dispar caterpillars on red oak and sugar maple leaves: Nutrition versus oxidative stress. Chemoecology 2013, 23, 59–70. [Google Scholar] [CrossRef]
- Hoffmann, T.; Nebehaj, E.; Albert, L. Antioxidant properties and detailed polyphenol profiling of European hornbeam (Carpinus betulus L.) leaves by multiple antioxidant capacity assays and high-performance liquid chromatography/multistage electrospray mass spectrometry. Ind. Crops Prod. 2016, 87, 340–349. [Google Scholar] [CrossRef] [Green Version]
- Tálos-Nebehaj, E.; Hofmann, T.; Albert, L. Seasonal changes of natural antioxidant content in the leaves of Hungarian forest trees. Ind. Crops Prod. 2017, 98, 53–59. [Google Scholar] [CrossRef]
- Steen, J.S.; Asplund, J.; Lie, M.H.; Nybakken, L. Environment rather than provenance explains levels of foliar phenolics in European beech (Fagus sylvatica L.) seedlings. Trees 2021, 35, 1555–1569. [Google Scholar] [CrossRef]
- Shields, V.D.; Broomell, B.P.; Salako, J.O. Host selection and acceptability of selected tree species by gypsy moth larvae, Lymantria dispar (L.). Ann. Entomol. Soc. Am. 2003, 96, 920–926. [Google Scholar] [CrossRef]
- Shields, V.D.; Smith, K.P.; Arnold, N.S.; Gordon, I.M.; Shaw, T.E.; Waranch, D. The effect of varying alkaloid concentrations on the feeding behavior of gypsy moth larvae, Lymantria dispar (L.) (Lepidoptera: Lymantriidae). Arthropod-Plant Interact 2008, 2, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gripenberg, S.; Mayhew, P.J.; Parnell, M.; Roslin, T. A meta-analysis of preference–performance relationships in phytophagous insects. Ecol. Lett. 2010, 13, 383–393. [Google Scholar] [CrossRef]
- Jones, L. Behavioural Adaptations of the Generalist Herbivore Helicoverpa punctigera (Lepidoptera: Noctuidae) with Respect to Primary and Secondary Hosts. Doctoral Dissertation, University of Queensland, Queensland, Australia, 2020. [Google Scholar]
- Miller, J.C.; Hanson, P.E. Laboratory feeding tests on the development of gypsy moth larvae with reference to plant taxa and allelochemicals. Stn. Bull. Agric. Exp. Stn. State Univ. 1989, 63. [Google Scholar]
- Miller, F.; Wiegrefe, S. Susceptibility, preference, and suitability of Carpinus and Ostrya taxa for gypsy moth larvae (Lepidoptera: Lymantriidae). Great Lakes Entomol. 2021, 54, 5. [Google Scholar]
- Barbosa, P.; Greenblatt, J.; Withers, W.; Cranshaw, W.; Harrington, E.A. Host-plant preferences and their induction in larvae of the gypsy moth, Lymantria dispar. Entomol. Exp. Appl. 1979, 26, 180–188. [Google Scholar] [CrossRef]
- Mauffette, Y.; Lechowicz, M.J. Differences in the utilization of tree species as larval hosts and pupation sites by the gypsy moth, Lymantria dispar (Lepidoptera: Lymantriidae). Can. Entomol. 1984, 116, 685–690. [Google Scholar] [CrossRef]
- Clavijo McCormick, A.; Arrigo, L.; Eggenberger, H.; Mescher, M.C.; De Moraes, C.M. Divergent behavioural responses of gypsy moth (Lymantria dispar) caterpillars from three different subspecies to potential host trees. Sci. Rep. 2019, 9, 8953. [Google Scholar] [CrossRef] [Green Version]
- Stockhoff, B.A. Protein intake by gypsy moth larvae on homogeneous and heterogeneous diets. Physiol. Entomol. 1993, 18, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Baghery, F.; Fathipour, Y.; Naseri, B. Nutritional indices of Helicoverpa armigera (Lep.: Noctuidae) on seeds of five host plants. Appl. Ent. Phytopath. 2013, 80, 19–27. [Google Scholar]
- Mariottini, Y.; Lange, C.E.; Cepeda, R.; De Wysiecki, M.L. Efficiency of food utilization by Dichroplus maculipennis (Orthoptera: Acrididae: Melanoplinae) on four crop plants under controlled conditions. Stud. Neotrop. Fauna Environ. 2019, 54, 206–216. [Google Scholar] [CrossRef]
- Couture, J.J.; Mason, C.J.; Habeck, C.W.; Lindroth, R.L. Behavioral and morphological responses of an insect herbivore to low nutrient quality are inhibited by plant chemical defenses. Arthropod-Plant Interact 2016, 10, 341–349. [Google Scholar] [CrossRef]
- Adel, M.M.; El-Hawary, F.M.; Abdel-Aziz, N.F.; Sammour, E.A. Some physiological, biochemical and histopathological effects of Artemisia monosperma against the cotton leafworm, Spodoptera littoralis. Arch. Phytopathol Pflanzenschutz 2010, 43, 1098–1110. [Google Scholar] [CrossRef]
- Mason, C.J.; Rubert-Nason, K.F.; Lindroth, R.L.; Raffa, K.F. Aspen defense chemicals influence midgut bacterial community composition of gypsy moth. J. Chem. Ecol. 2015, 41, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Wang, Y.; Zou, H.; Ding, N.; Geng, N.; Cao, C.; Zhang, G. Sanguinarine in Chelidonium majus induced antifeeding and larval lethality by suppressing food intake and digestive enzymes in Lymantria dispar. Pestic. Biochem. Physiol. 2019, 153, 9–16. [Google Scholar] [CrossRef]
- Lazarević, J. Physiological and Genetic Mechanisms of Adaptation to Unsuitable Nutrition in the Gypsy Moth Lymantria dispar L. Ph.D. Dissertation, Faculty of Biology, University of Belgrade, Belgrade, Serbia, 2000. [Google Scholar]
- Lazarević, J.; Perić-Mataruga, V.; Stojković, B.; Tucić, N. Adaptation of the gypsy moth to an unsuitable host plant. Entomol. Exp. Appl. 2002, 102, 75–86. [Google Scholar] [CrossRef]
- Perić-Mataruga, V.; Lazarević, J.; Vlahović, M.; Mrdaković, M.; Ilijin, L. Note: Histology of the midgut and peritrophic membrane in Lymantria dispar caterpillars fed on leaves of Quercus cerris or Robinia pseudoacacia. Phytoparasitica 2006, 34, 49–53. [Google Scholar] [CrossRef]
- Perić-Mataruga, V.; Blagojević, D.; Spasić, M.B.; Ivanović, J.; Janković-Hladni, M. Effect of the host plant on the antioxidative defence in the midgut of Lymantria dispar L. caterpillars of different population origins. J. Insect Physiol. 1997, 43, 101–106. [Google Scholar] [CrossRef]
- Lazarević, J.; Janković-Tomanić, M.; Savković, U.; Đorđević, M.; Milanović, S.; Stojković, B. Host-associated divergence in the activity of digestive enzymes in two populations of the gypsy moth Lymantria dispar (Lepidoptera: Erebidae). Entomol. Sci. 2017, 20, 189–194. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Indices | Formulae | Units |
|---|---|---|
| Mass gain | mt − m0 | mg |
| Amount of consumed food | l0 − lt | mg |
| Amount of assimilated food | l0 − lt − me | mg |
| Relative growth rate (RGR) | (mt − m0)/(t × m0) | mg/mg/day |
| Relative consumption rate (RCR) | (l0 − lt)/(t × m0) | mg/mg/day |
| Efficiency of conversion of ingested food (ECI) | (mt − m0)/(l0 − lt) × 100 | % |
| Approximate digestibility (AD) | (l0 − lt − me)/(l0 − lt) × 100 | % |
| Efficiency of conversion of digested food (ECD) | (mt − m0)/(l0 − lt − me) × 100 | % |
| Leaf Traits | Turkey Oak | Beech | Hornbeam | F | p |
|---|---|---|---|---|---|
| C (%) | 34.6 ± 0.56 a | 33.8 ± 0.34 a | 31.2 ± 0.13 b | 22.4 | 0.0003 |
| N (%) | 2.4 ± 0.05 a | 1.9 ± 0.04 b | 2.3 ± 0.12 ab | 8.96 | 0.0072 |
| C/N | 15.3 ± 0.98 ab | 17.4 ± 0.45 a | 14.0 ± 0.81 b | 5.03 | 0.0342 |
| W (%) | 65.5 ± 0.53 b | 63.6 ± 0.40 c | 69.8 ± 0.55 a | 38.68 | <0.0001 |
| SLM (mg cm−2) | 4.5 ± 0.14 a | 3.4 ± 0.11 b | 2.3 ± 0.04 c | 113.41 | <0.0001 |
| Performance Indices | Turkey Oak | Beech | Hornbeam |
|---|---|---|---|
| Duration of feeding trial (days) | 5.2 ± 0.09 b | 5.7 ± 0.15 a | 5.8 ± 0.14 a |
| Initial mass (mg) | 7.9 ± 0.25 a | 7.2 ± 0.12 a | 7.6 ± 0.21 a |
| Final mass (mg) | 23.3 ± 1.06 a | 17.8 ± 0.82 b | 16.1 ± 0.85 b |
| Mass gain (mg) | 15.4 ± 0.94 a | 10.6 ± 0.78 b | 8.5 ± 0.73 b |
| Food consumed (mg) | 108.0 ± 4.72 b | 119.4 ± 6.22 b | 142.1 ± 6.33 a |
| Food assimilated (mg) | 38.9 ± 2.62 b | 39.6 ± 3.63 b | 63.1 ± 4.81 a |
| RGR (mg/mg/day) | 0.38 ± 0.02 a | 0.25 ± 0.02 b | 0.19 ± 0.02 c |
| RCR (mg/mg/day) | 2.7 ± 0.09 a | 2.9 ± 0.15 a | 3.4 ± 0.14 a |
| ECI (%) | 14.2 ± 0.56 a | 8.8 ± 0.39 b | 5.7 ± 0.38 c |
| AD (%) | 35.4 ± 1.53 b | 32.2 ± 1.62 b | 45.8 ± 2.20 a |
| ECD (%) | 44.4 ± 3.35 a | 31.3 ± 2.61 b | 14.1 ± 1.41 b |
| Performance Indices | Source of Variation | MS | F | p |
|---|---|---|---|---|
| Initial mass | Host plant | 0.108 | 2.70 | 0.0717 |
| Error | 0.040 | |||
| Final mass | Host plant | 6.253 | 16.60 | <0.0001 |
| Error | 0.377 | |||
| Mass gain | Host plant | 9.634 | 18.30 | <0.0001 |
| Error | 0.527 | |||
| Food consumed | Host plant | 23.490 | 9.74 | 0.0001 |
| Error | 2.410 | |||
| Food assimilated | Host plant | 40.394 | 19.14 | <0.0001 |
| Error | 2.110 |
| Performance Indices | Source of Variation | MS | F | p |
|---|---|---|---|---|
| Growth rate~RGR | Host plant | 1.949 | 30.10 | <0.0001 |
| Covariate (a) | 0.812 | 12.55 | 0.0006 | |
| Error | 0.065 | |||
| Consumption rate~RCR | Host plant | 1.162 | 2.16 | 0.1204 |
| Covariate (a) | 4.113 | 7.64 | 0.0067 | |
| Error | 0.538 | |||
| Mass gain~ECI | Host plant | 16.585 | 54.86 | <0.0001 |
| Covariate (b) | 23.832 | 78.83 | <0.0001 | |
| Error | 0.302 | |||
| Assimilation~AD | Host plant | 9.377 | 10.74 | 0.0001 |
| Covariate (b) | 129.535 | 148.43 | <0.0001 | |
| Error | 0.873 | |||
| Mass gain ~ ECD | Host plant | 12.590 | 26.63 | <0.0001 |
| Covariate (c) | 6.077 | 12.86 | 0.0005 | |
| Error | 0.473 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milanović, S.; Miletić, Z.; Marković, Č.; Šešlija Jovanović, D.; Trailović, Z.; Jankovský, L.; Lazarević, J. Suitability of Turkey Oak, European Beech, and Hornbeam to Gypsy Moth Feeding. Forests 2022, 13, 1006. https://doi.org/10.3390/f13071006
Milanović S, Miletić Z, Marković Č, Šešlija Jovanović D, Trailović Z, Jankovský L, Lazarević J. Suitability of Turkey Oak, European Beech, and Hornbeam to Gypsy Moth Feeding. Forests. 2022; 13(7):1006. https://doi.org/10.3390/f13071006
Chicago/Turabian StyleMilanović, Slobodan, Zoran Miletić, Čedomir Marković, Darka Šešlija Jovanović, Zoran Trailović, Libor Jankovský, and Jelica Lazarević. 2022. "Suitability of Turkey Oak, European Beech, and Hornbeam to Gypsy Moth Feeding" Forests 13, no. 7: 1006. https://doi.org/10.3390/f13071006