
Invasive Insect Pests of Forests and Urban Trees in Russia: Origin, Pathways, Damage, and Management

 ,
,  , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. The Western Coniferous Seed Bug, Leptoglossus occidentalis Heidemann, 1910 (Hemiptera: Heteroptera: Coreidae)
3.2. The Brown Marmorated Stink Bug, Halyomorpha halys (Stål, 1855) (Hemiptera: Heteroptera: Pentatomidae)
3.3. The Oak Lace Bug, Corythucha arcuata (Say, 1832) (Hemiptera: Heteroptera: Tingidae)
3.4. The Spotted Poplar Borer, Agrilus fleischeri Obenberger, 1925 (Coleoptera: Buprestidae)
3.5. The Apple Buprestid, Agrilus mali Matsumura, 1924 (Coleoptera: Buprestidae)
3.6. The Emerald Ash Borer, Agrilus planipennis Fairmaire, 1888 (Coleoptera: Buprestidae)
3.7. The Cypress Jewel Beetle, Lamprodila (Palmar) festiva (Linnaeus, 1767) (Coleoptera: Buprestidae)
3.8. The Small Spruce Bark Beetle, Ips amitinus (Eichhoff, 1872) (Coleoptera: Curculionidae: Scolytinae)
3.9. The Four-Eyed Fir Bark Beetle, Polygraphus proximus Blandford, 1894 (Coleoptera: Curculionidae: Scolytinae)
3.10. The Box Tree Moth, Cydalima perspectalis (Walker, 1859) (Lepidoptera: Crambidae)
3.11. The Leaf Blotch Miner Moth, Acrocercops brongniardella (Fabricius, 1798) (Lepidoptera: Gracillariidae)
3.12. The Horse-Chestnut Leaf Miner, Cameraria ohridella Deschka et Dimić, 1986 (Lepidoptera: Gracillariidae)
3.13. The Lime Leaf Miner, Phyllonorycter issikii (Kumata, 1963) (Lepidoptera: Gracillariidae)
3.14. The Poplar Leafminer, Phyllonorycter populifoliella (Treitschke, 1833) (Lepidoptera: Gracillariidae)
4. Discussion
4.1. Taxonomy
4.2. Directions of Invasions
- (1)
- Invasions from Asia to West or central Europe and then (or directly) to European Russia: This is the most numerous group, which consists of five species—Cydalima perspectalis, Phyllonorycter issikii, Halyomorpha halys (likely first invaded North America and only then Europe and European Russia; see above), and two species, which actually skipped West or central Europe and arrived directly to European Russia, namely, Agrilus planipennis and Polygraphus proximus;
- (2)
- Invasions from North America to West or central Europe and then to European Russia: Leptoglossus occidentalis and Corythucha arcuata;
- (3)
- Invasions from Europe to Asia: the case of Ips amitinus;
- (4)
- Range expansions within Europe and invasions to European Russia: Lamprodila festiva and Cameraria ohridella.
4.3. Causes and Pathways of Invasions
4.4. Probability of Further Invasions to Neighboring and Distant Countries
4.5. Role of the Trophic Factor in Insect Pest Range Expansion
- (1)
- A shift in an insect species (often not even a pest) within its native range from its usual host plant(s) to introduced and cultivated host plant(s) (usually from the same or close genus of woody plants and often non-resistant because of lack of co-evolution) (e.g., a shift in Agrilus planipennis in Russian Far East and China, from local Asian ash species to introduced North American ash species; a shift in Phyllonorycter populifoliella in European Russia and Siberia, from local poplars to widely cultivated introduced North American balsam poplar and hybrids; a shift in Lamprodila festiva from wild Cupressaceae to introduced Thuja and other cultivated representatives of this family, including their hybrids and cultivars in the Mediterranean region; a shift in Agrilus fleischeri to introduced poplars in China);
- (2)
- A local niche expansion due to exploration of cultivated, recently introduced host plants in anthropogenic (urban or agricultural) landscapes; local population build-up and outbreaks (e.g., recorded earlier in Agrilus planipennis, A. mali, Phyllonorycter populifoliella, and Lamprodila festiva, and currently seen in A. fleischeri);
- (3)
- A range expansion outside the limits of the native range through anthropogenic (urban or agricultural) landscapes, i.e., beginning of invasion (e.g., Agrilus mali and Lamprodila festiva);
- (4)
- A distant invasion (e.g., invasions of Agrilus planipennis to European Russia or North America or Ips amitinus to Siberia);
- (5)
- A secondary host plant shift to the native (for the invaded region) food plant(s) (e.g., shift in Agrilus planipennis to Chionanthus virginicus in North America [399] and to Fraxinus excelsior in European Russia; shift in A. mali from cultivated apples to the wild apple, Malus sieversii in China).
4.6. Monitoring and Control Measures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pimentel, D.; McNair, S.; Janecka, J.; Wightman, J.; Simmonds, C.; O’Connell, C.; Won, E.; Russel, L.; Zern, J.; Aquino, T.; et al. Economic and environmental threats of alien plant, animal, and microbe invasions. Agric. Ecosys. Environ. 2001, 84, 1–20. [Google Scholar] [CrossRef]
- Fei, S.; Phillips, J.; Shouse, M. Biogeomorphic impacts of invasive species. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 69–87. [Google Scholar] [CrossRef] [Green Version]
- Liebhold, A.M.; Brockerhoff, E.G.; Nunez, M.A. Biological invasions in forest ecosystems: A global problem requiring international and multidisciplinary integration. Biol. Invasions 2017, 19, 3073–3077. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; van Kleunen, M.; Winter, M.; et al. The global rise in emerging alien species results from increased accessibility of new source pools. Proc. Natl. Acad. Sci. USA 2018, 115, E2264–E2273. [Google Scholar] [CrossRef] [Green Version]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Paap, T.; Burgess, T.I.; Wingfield, M.J. Urban trees: Bridge-heads for forest pest invasions and sentinels for early detection. Biol. Invasions 2017, 19, 3515–3526. [Google Scholar] [CrossRef] [Green Version]
- Panzavolta, T.; Bracalini, M.; Benigno, A.; Moricca, S. Alien invasive pathogens and pests harming trees, forests, and plantations: Pathways, global consequences and management. Forests 2021, 12, 1364. [Google Scholar] [CrossRef]
- Maslyakov, V.Y.; Izhevsky, S.S. Alien Phytophagous Insects Invasions in the European Part of Russia; Institute of Geography ot Russian Academy of Sciences: Moscow, Russia, 2011; pp. 1–272. (In Russian) [Google Scholar]
- Deschka, G.; Dimić, N. Cameraria ohridella n. sp. (Lepidoptera, Lithocolletidae) aus Mazedonien, Jugoslawien. Acta Entomol. Jugosl. 1986, 22, 11–23. [Google Scholar]
- van Nieukerken, E.; Wagner, D.; Baldessari, M.; Mazzon, L.; Angeli, G.; Girolami, V.; Duso, C.; Doorenweerdet, C. Antispila oinophylla new species (Lepidoptera, Heliozelidae), a new North American grapevine leafminer invading Italian vineyards: Taxonomy, DNA barcodes and life cycle. ZooKeys 2012, 170, 29–77. [Google Scholar] [CrossRef] [Green Version]
- Mlynarek, J.J. Testing the enemy release hypothesis in a native insect species with an expanding range. PeerJ 2015, 3, e1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, Y.; Wei, K.; Wang, X.; Duan, J.J.; Jennings, D.E.; Poland, T.M. Introduced plants induce outbreaks of a native pest and facilitate invasion in the plants’ native range: Evidence from the emerald ash borer. J. Ecol. 2022; in press. [Google Scholar] [CrossRef]
- Venette, R.C.; Hutchison, W.D. Invasive insect species: Global challenges, strategies & opportunities. Front. Insect Sci. 2021, 1, 1. [Google Scholar] [CrossRef]
- Akulov, E.N.; Ponomarenko, M.G.; Kirichenko, N.I. Exploring fauna of Microlepidoptera in South Siberia: Novel regional records and interception of quarantine species. J. Asia-Pac. Biodivers. 2019, 12, 597–612. [Google Scholar] [CrossRef]
- Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. In The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L.; Kirichenko, N.I.; Selikhovkin, A.V. (Eds.) Saint Petersburg State Forest Technical University: St. Petersburg, Russia, 2020; p. 452. Available online: http://spbftu.ru/wp-content/uploads/2020/10/Kataev-Readings-XI-2020.pdf (accessed on 20 January 2022).
- ESRI. ArcGIS Desktop: Release 9.3; Environmental Systems Research Institute: Redlands, CA, USA, 2008; Available online: http://www.esri.com/software/arcgis/eval-help/arcgis-93 (accessed on 20 January 2022).
- Gapon, D.A. First records of the western conifer seed bug Leptoglossus occidentalis Heid. (Heteroptera, Coreidae) from Russia and Ukraine, regularities in its distribution and possibilities of its range expansion in the Palaearctic region. Entomol. Rev. 2013, 93, 174–181. [Google Scholar] [CrossRef]
- Tescari, G. Leptoglossus occidentalis coreide nearctico rinveuto in Italia (Heteroptera, Coreidae). Lavor. Societa Veneziana Scie. Nat. 2001, 26, 3–5. [Google Scholar]
- Gninenko, Y.I.; Cheplyansky, I.Y.; Chernova, U.A.; Rakov, A.G.; Khegai, I.V.; Latyshova, N.S.; Gimranov, R.I. Methodical Recommendations for Protection against Pine Seed Bug (for Industrial Trial); VNIILM: Pushkino, Russia, 2019; p. 28. [Google Scholar]
- Fent, M.; Kment, P. First record of the invasive western conifer seed bug Leptoglossus occidentalis (Heteroptera: Coreidae) in Turkey. North-West. J. Zool. 2011, 7, 72–80. Available online: https://www.researchgate.net/publication/228493133_First_record_of_the_invasive_western_conifer_seed_bug_Leptoglossus_occidentalis_Heteroptera_Coreidae_in_Turkey (accessed on 20 January 2022).
- Ahn, S.J.; Son, D.; Choo, P.Y.; Parka, C.G. The first record on Leptoglossus occidentalis (Hemiptera: Coreidae) in Korea, a potential pest of the pinaceous tree species. J. Asia-Pac. Entomol. 2013, 16, 281–284. [Google Scholar] [CrossRef]
- Aukema, B.; Rieger, C.; Rabitsch, W. (Eds.) Catalogue of the Heteroptera of the Palaearctic Region. Vol. 6. Supplement; The Netherlands Entomological Society: Wageningen, The Netherlands, 2013; 629p. [Google Scholar]
- Barclay, M.; Nikolaeva, S. Arrival in Kazakhstan of Leptoglossus occidentalis (Hemiptera: Heteroptera: Coreidae); a North American invasive species expands 2500 kilometres to the east. Klapalekiana 2018, 54, 1–3. [Google Scholar]
- van der Heyden, T. Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae: Coreinae: Anisoscelini) in Israel. Revista Chilena Entomol. 2019, 45, 435–437. [Google Scholar] [CrossRef]
- Kalashian, M.Y.; Ghrejyan, T.L.; Karagyan, G.H. First finding of Western conifer seed bug Leptoglossus occidentalis Heid. (Heteroptera, Coreidae) in Armenia. Russ. J. Biol. Invasions 2021, 12, 274–276. [Google Scholar] [CrossRef]
- Ben Jamâa, M.L.; Mejri, M.; Naves, P.; Sousa, E. Detection of Leptoglossus occidentalis Heidemann, 1910 (Heteroptera: Coreidae) in Tunisia. Afr. Entomol. 2013, 21, 165–167. [Google Scholar] [CrossRef]
- Gapon, D.A.; Busarova, N.V.; Komarov, Y.E. New records of the western conifer seed bug Leptoglossus occidentalis (Heteroptera: Coreidae) in Russia and in adjacent territories. Cauc. Entomol. Bull. 2016, 12, 221–222. (In Russian) [Google Scholar] [CrossRef]
- van der Heyden, T.; Faúndez, E.I. First records of Leptoglossus occidentalis Heidemann, 1910 (Hemiptera: Heteroptera: Coreidae) in Brazil and South Africa. Boletín Mus. Nac. Hist. Nat. Parag. 2020, 24, 28–30. Available online: https://www.researchgate.net/publication/341832223_First_records_of_Leptoglossus_occidentalis_Heidemann_1910_Hemiptera_Heteroptera_Coreidae_in_Brazil_and_South_Africa (accessed on 20 January 2022).
- Faúndez, E.I.; Rocca, J.; Villablanca, J. Detection of the invasive western conifer seed bug Leptoglossus occidentalis Heidemann, 1910 (Heteroptera: Coreidae: Coreinae) in Chile. Arq. Entomol. 2017, 17, 317–320. Available online: https://www.researchgate.net/publication/316277401_Detection_of_the_invasive_western_conifer_seed_bug_Leptoglossus_occidentalis_Heidemann_1910_Heteroptera_Coreidae_Coreinae_in_Chile (accessed on 20 January 2022).
- Carpintero, D.L.; Farina, J.L.; De Biase, S. Reporte para la provincia de Buenos Aires de tres especies de Heteroptera (Hemiptera) introducidas en Argentina. Hist. Nat. Terc. Ser. 2019, 9, 63–70. [Google Scholar]
- Faúndez, E.I.; Silvera, M. Sobre la presencia de la chinche de las coníferas occidental Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae) en Uruguay. Rev. Chil. Entomol. 2019, 45, 549–551. [Google Scholar] [CrossRef] [Green Version]
- Gildenkov, M.Y. Invasive bug species Leptoglossus occidentalis Heidemann, 1910 (Heteroptera: Coreidae) in ecosystems of the Smolensk area. In Modern Scientific Research: Trends and Prospects, Proceedings of the 2nd All-Russia Conference with International Participation, Chistopol, Tatarstan, Russia, 26 February 2021; Nazarov, E.A., Ed.; Astor i Ya Publisher: Kazan, Russia, 2021; pp. 5–8. (In Russian) [Google Scholar]
- Golub, V.B.; Aksenenko, E.V.; Soboleva, V.A.; Kornev, I.I. New data on the distribution of the tropical bed bug Cimex hemipterus and the western conifer seed bug Leptoglossus occidentalis (Heteroptera: Cimicidae, Coreidae) in the European part of Russia. Russ. J. Biol. Invasions 2020, 11, 97–100. [Google Scholar] [CrossRef]
- Golub, V.B. Personal Communication; Voronezh State University: Voronezh, Russia, 2021. [Google Scholar]
- Gninenko, Y.I.; Gapon, D.A.; Shchurov, V.I.; Bondarenko, A.S. A western conifer seed bug Leptoglossus occidentalis (Heteroptera, Coreidae) emerged in Russia. Plant Prot. Quar. 2014, 6, 38–40. Available online: https://www.elibrary.ru/download/elibrary_21561963_71479165.pdf (accessed on 20 January 2022). (In Russian).
- van der Heyden, T. New data on the distribution of Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae: Coreinae: Anisoscelini), including the first record of the species in Georgia. Rev. Chil. Entomol. 2018, 44, 433–435. [Google Scholar]
- Koerber, T.W. Leptoglossus occidentalis (Hemiptera, Coreidae), a newly discovered pest of coniferous seed. Ann. Entomol. Soc. Am. 1963, 56, 229–234. [Google Scholar] [CrossRef]
- Werner, D.J. Die amerikanische Koniferen-Samen-Wanze Leptoglossus occidentalis (Heteroptera: Coreidae) als Neozoon in Europa und in Deutschland: Ausbreitung und Biologie. Entomol. Heute 2011, 23, 31–68. Available online: https://www.zobodat.at/pdf/Entomologie-heute_23_0031-0068.pdf (accessed on 20 January 2022).
- Rice, R.E.; Uyemoto, J.K.; Ogawa, J.M.; Pemberton, W.M. New findings on pistachio problems. Calif. Agric. 1985, 39, 15–18. [Google Scholar]
- Barta, M. Biology and temperature requirements of the invasive seed bug Leptoglossus occidentalis (Heteroptera: Coreidae) in Europe. J. Pest. Sci. 2016, 89, 31–44. [Google Scholar] [CrossRef]
- Hedlin, A.F.; Yates, H.O., III; Tovar, D.C.; Ebel, B.H.; Koerber, T.W.; Merkel, E.P. Cone and Seed Insects of North American Conifers; Environment Canada, Forestry Service, Pacific Forestry Centre: Victoria, BC, Canada; The United States Forest Service: Wahington, DC, USA; Secretaría de Agricultura y Recursos Hidráulicos: Tulla de Allende, Mexico, 1981; pp. 1–122. Available online: https://cfs.nrcan.gc.ca/publications?id=2026 (accessed on 2 January 2022).
- Bernardinelli, I.; Rovato, M.; Zandigiacomo, P. Life history and laboratory rearing of Leptoglossus occidentalis. In IUFRO Working Party 7.III.10, Proceedings of the Workshop, Gmunden, Austria, 11–14 September 2006; Federal Research and Training Centre for Forests, Natural Hazards and Landscape Forest Training Centre Ort: Gmunden, Austria, 2006; p. 225. [Google Scholar]
- Rabitsch, W. Alien true bugs of Europe (Insecta: Hemiptera: Heteroptera). Zootaxa 2008, 1827, 1–44. [Google Scholar] [CrossRef] [Green Version]
- Luchi, N.; Mancini, V.; Feducci, M.; Santini, A.; Capretti, P. Leptoglossus occidentalis and Diplodia pinea: A new insect-fungus association in Mediterranean forests. For. Pathol. 2012, 42, 246–251. [Google Scholar] [CrossRef]
- Tamburini, M.; Maresi, G.; Salvadori, C.; Battisti, A.; Zottele, F.; Pedrazzoli, F. Adaptation of the invasive western conifer seed bug Leptoglossus occidentalis to Trentino, an alpine region (Italy). Bull. Insectol. 2012, 65, 161–170. Available online: https://www.researchgate.net/publication/233100158_Adaptation_of_the_invasive_western_conifer_seed_bug_Leptoglossus_occidentalis_to_Trentino_an_alpine_region_Italy (accessed on 20 January 2022).
- Cargnus, E.; Buian, F.M.; Zandigiacomo, P. Presenza di Trichopoda pennipes (Diptera, Tachinidae) nell’Italia nord-orientale. Boll. Soc. Nat. Silvia Zenari 2011, 35, 123–130. [Google Scholar]
- Maltese, M.; Caleca, V.; Guerrieri, E.; Strong, W.B. Parasitoids of Leptoglossus occidentalis Heidemann (Heteroptera: Coreidae) recovered in western North America and first record of its egg parasitoid Gryon pennsylvanicum (Ashmead) (Hymenoptera: Platygastridae) in California. Pan-Pac. Entomol. 2012, 88, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef]
- Haye, T.; Gariepy, T.; Hoelmer, K.; Rossi, J.-P.; Streito, J.-C.; Tassus, X.; Desneux, N. Range expansion of the invasive brown marmorated stink bug, Halyomorpha halys: An increasing threat to field, fruit and vegetable crops worldwide. J. Pest Sci. 2015, 88, 665–673. [Google Scholar] [CrossRef]
- Hoebeke, E.R.; Carter, M.E. Halyomorpha halys (Stål) (Heteroptera: Pentatomidae): A polyphagous plant pest from Asia newly detected in North America. Proc. Entomol. Soc. Wash. 2003, 105, 225–237. Available online: https://www.researchgate.net/publication/279897494_Halyomorpha_halys_Stal_Heteroptera_Pentatomidae_A_polyphagous_plant_pest_from_Asia_newly_detected_in_North_America (accessed on 20 January 2022).
- Lee, D.-H. Current status of research progress on the biology and management of Halyomorpha halys (Hemiptera: Pentatomidae) as an invasive species. Appl. Entomol. Zool. 2015, 50, 277–290. [Google Scholar] [CrossRef]
- Hamilton, G.C.; Ahn, J.J.; Bu, W.; Leskey, T.C.; Nielsen, A.L.; Park, Y.-L.; Rabitsch, W.; Hoelmer, K.A. Halyomorpha halys (Stål). In Invasive Stink Bugs and Related Species (Pentatomoidea): Biology, Higher Systematics, Semiochemistry and Management; McPherson, J.E., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 243–292. [Google Scholar] [CrossRef]
- Fogain, R.; Graff, S. First records of the invasive pest, Halyomorpha halys (Hemiptera: Pentatomidae), in Ontario and Quebec. J. Entomol. Soc. Ont. 2011, 142, 45–48. [Google Scholar]
- Faúndez, E.I.; Rider, D.A. The brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae) in Chile. Arquivos Entomol. 2017, 17, 305–307. Available online: https://www.researchgate.net/publication/316277383_The_brown_marmorated_stink_bug_Halyomorpha_halys_Stal_1855_Heteroptera_Pentatomidae_in_Chile (accessed on 12 December 2021).
- Arnold, K. Halyomorpha halys (Stål, 1855), eine für die europäische Fauna neu nachgewiesene Wanzenart (Insecta: Heteroptera: Pentatomidae: Cappaeini). Mitt. Des. Thüring. Entomol. 2009, 16, 19. [Google Scholar]
- Gariepy, T.D.; Musolin, D.L.; Konjević, A.; Karpun, N.N.; Zakharchenko, V.Y.; Zhuravleva, E.N.; Tavella, L.; Bruin, A.; Haye, T. Diversity and distribution of cytochrome oxidase I (COI) haplotypes of the brown marmorated stink bug, Halyomorpha halys Stål (Hemiptera, Pentatomidae), along the eastern front of its invasive range in Eurasia. NeoBiota 2021, 68, 53–77. [Google Scholar] [CrossRef]
- Harris, A.C. Halyomorpha halys (Hemiptera: Pentatomidae) and Protaetia brevitaris (Coleoptera: Scarabaeidae: Cetoniinae) intercepted in Dunedin. Weta 2010, 40, 42–44. Available online: https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.694.5502&rep=rep1&type=pdf (accessed on 20 January 2022).
- Malumphy, C. Second interception of Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) in Britain. Het News 2014, 21, 4–5. [Google Scholar]
- Mityushev, I.M. First record of Halyomorpha halys in Russia. Plant Prot. Quar. 2016, 3, 48. Available online: https://www.elibrary.ru/download/elibrary_25606587_18888986.pdf (accessed on 2 December 2021). (In Russian).
- Musolin, D.L.; Konjević, A.; Karpun, N.N.; Protsenko, V.Y.; Ayba, L.Y.; Saulich, A.K. Invasive brown marmorated stink bug Halyomorpha halys (Stål) (Heteroptera: Pentatomidae) in Russia, Abkhazia, and Serbia: Range expansion, early stages of establishment and first records of damage to local crops. Arthropod-Plant Interact. 2018, 12, 517–529. [Google Scholar] [CrossRef]
- Borisov, B.A.; Karpun, N.N.; Protsenko, V.Y. New data on trophic relationships of the invasive brown marmorated stink bug Halyomorpha halys Stål (Heteroptera: Pentatomidae) in the subtropical zone of the Black Sea coast of the Caucasus. In Monitoring and Biological Control Methods of Woody Plant Pests and Pathogens: From Theory to Practice, Proceedings of the 2nd International Conference, Moscow, Russia, 22–26 April 2019; Baranchikov, Y.N., Ed.; SIF SN RASc.: Krasnoyarsk, Russia, 2019; pp. 33–35. (In Russian) [Google Scholar]
- Karpun, N.N.; Grebennikov, K.A.; Protsenko, V.E.; Ayba, L.Y.; Borisov, B.A.; Mityushev, I.M.; Zhimerkin, V.N.; Ponomaryov, V.L.; Chekmaryov, P.A.; Dolzhenko, V.I.; et al. Brown marmorated stink bug monitoring and identification methods Halyomorpha halys Stål, 1855. Plant Quar. Sci. Pract. 2018, 2, 7–11. Available online: https://elibrary.ru/download/elibrary_35209732_42561776.pdf (accessed on 18 January 2022). (In Russian).
- Karpun, N.N. The Structure of Complexes of Harmful Organisms of Woody Plants in the Humid Subtropics of Russia and the Biological Basis of Protection Measures. Ph.D. Thesis, Russian State Agricultural University, Moscow, Russia, 2018. Available online: http://www.old.timacad.ru/catalog/disser/dd/karpun/disser.pdf (accessed on 20 January 2022). (In Russian).
- Zamotajlov, A.S.; Belyi, A.I.; Esipenko, L.P. Harmfulness of herbivorous bugs of the family Pentatomidae (Insecta, Heteroptera) on tomatoes. In Results of Research Work for 2017, Proceedings of the 73th Research Conference of the Kuban’ State Agricultural University, Krasnodar, Russia, 14 March 2018; Koshaev, A.G., Ed.; Kuban State Agricultural University: Krasnodar, Russia, 2018; pp. 43–44. (In Russian) [Google Scholar]
- Stryukova, N.M.; Stryukov, A.A. The first detection of the brown marmorated stink bug in Crimea. In Sustainable Neospheric Development, Proceedings of the Scientific Inter-university Conference Devoted to the 156th Anniversary of V.I. Vernadsky, Simferopol, Russia, 15 March 2019; Bashta, A.I., Ed.; IP Zueva, T.V. Publisher: Simferopol, Russia, 2019; pp. 68–70. (In Russian) [Google Scholar]
- Zhuravleva, E.N.; Karpun, N.N. On the discovery of the brown marmorated stink bug Halyomorpha halys Stål (Heteroptera: Pentatomidae) in Sevastopol. In Monitoring and Biological Control Methods of Woody Plant Pests and Pathogens: From Theory to Practice, Proceedings of the 2nd International Conference, Moscow, Russia, 22–26 April 2019; Baranchikov, Y.N., Ed.; SIF SN RASc.: Moscow, Russia; Krasnoyarsk, Russia, 2019; pp. 74–75. (In Russian) [Google Scholar]
- Gapon, D.A. The brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae): Expansion of its range in the European part of Russia, description of the imago and larvae, and the diagnostics of the species. Cauc. Entomol. Bull. 2019, 15, 241–247. [Google Scholar] [CrossRef]
- Bulgakov, T.S.; Federal Research Centre the Subtropical Scientific Centre of the Russian Academy of Sciences, Sochi, Russia. Personal communication, 2020.
- Chenikalova, E.V. Marmorated stink bug Halyomorpha halys Stål in Stavropol. Proc. Stavropol Branch Russ. Entomol. Soc. 2020, 16, 46–49. [Google Scholar]
- Petrov, A.V.; Institute of Forest Science of Russian Academy of Science, Moscow, Russia. Personal communication, 2020.
- Watanabe, K. Specific characteristics of damage to cherry caused by Lycogoris (Apolygus) lucorum (Meyer-Dür) (Heteroptera: Miridae) and Halyomorpha halys (Stål) (Heteroptera: Pentatomidae). Annu. Rep. Plant Prot. Soc. N. Jpn. 1996, 1996, 143–144. [Google Scholar]
- Bergmann, E.; Bernhard, K.M.; Bernon, G.; Bickerton, M.; Gill, S.; Gonzales, C.; Hamilton, G.C.; Hedstrom, C.; Kamminga, K.; Koplinka-Loehr, C.; et al. Host Plants of the Brown Marmorated Stink Bug in the U.S. Available online: https://www.stopbmsb.org/where-is-bmsb/host-plants (accessed on 2 January 2022).
- Zakharchenko, V.; Karpun, N.; Borisov, B. Trophic connections of the brown marmorated stink bug Halyomorpha halys Stål in the conditions of the invasive area on the Black Sea coast of the Caucasus. BIO Web. Conf. 2020, 21, 7. [Google Scholar] [CrossRef]
- Zakharchenko, V.Y. Bioecological Features of the Brown Marmorated Stink Bug (Halyomorpha halys Stål) in the Humid Subtropics of Russia and Control Measures. Ph.D. Thesis, Russian State Agricultural University, Moscow, Russia, 2021. Available online: http://www.old.timacad.ru/catalog/disser/kd/zaharchenko/zaharchenko_disser.pdf (accessed on 10 January 2022). (In Russian).
- Musolin, D.L.; Dolgovskaya, M.Y.; Protsenko, V.Y.; Karpun, N.N.; Reznik, S.Y.; Saulich, A.K. Photoperiodic and temperature control of nymphal growth and adult diapause induction in the invasive Caucasian population of the Brown marmorated stink bug, Halyomorpha halys. J. Pest Sci. 2019, 92, 621–631. [Google Scholar] [CrossRef]
- Khrimian, A.; Zhang, A.; Weber, D.C.; Ho, H.-Y.; Aldrich, J.R.; Vermillion, K.E.; Siegler, M.A.; Shirali, S.; Guzma, F.; Leskey, T.C. Discovery of the aggregation pheromone of the Brown marmorated stink bug (Halyomorpha halys) through the creation of stereoisomeric libraries of 1-bisabolen-3-ols. J. Nat. Prod. 2014, 77, 1708–1717. [Google Scholar] [CrossRef] [PubMed]
- Sinitsyna, E.V.; Protsenko, V.E.; Karpun, N.N.; Mityushev, I.M.; Lobur, A.Y.; Todorov, N.G. First field trials of Russia-produced pheromone preparations for monitoring and control of the Brown marmorated stink bug, Halyomorpha halys Stål. Bull. Timiryazev Agricult. Acad. 2019, 3, 60–79. [Google Scholar] [CrossRef]
- Gouli, V.; Gouli, S.; Skinner, M.; Hamilton, G.; Kim, J.S.; Parker, B.L. Virulence of select entomopathogenic fungi to the brown marmorated stink bug, Halyomorpha halys (Stål) (Heteroptera: Pentatomidae). Pest Manag. Sci. 2012, 68, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.L.; Skinner, M.; Gouli, S.; Gouli, V.; Kim, J.S. Virulence of BotaniGard® to second instar brown marmorated stink bug, Halyomorpha halys (Stål) (Heteroptera: Pentatomidae). Insects 2015, 6, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Gorgadze, O.; Bakhtadze, G.; Kereselidze, M.; Lortkhipanidze, M. The efficacy of entomopathogenic agents against Halyomorpha halys (Hemiptera: Pentatomidae). Int. J. Current Res. 2017, 9, 62177–62180. Available online: https://www.journalcra.com/sites/default/files/issue-pdf/27484.pdf (accessed on 10 January 2022).
- Borisov, B.A.; Protsenko, V.Y.; Karpun, N.N. Potential use of entomotrophic bacteria and fungi to contain the brown marmorated stink bug Halyomorpha halys Stål (Heteroptera: Pentatomidae). In Biological Plant Protection as a Basis for Sustainable Agroecosystems, Proceedings of the X International Scientific and Practical Conference, Krasnodar, Russia, 11–13 September 2018; All-Russia Research Institute of Biological Plant Protection: Krasnodar, Russia, 2018; pp. 165–168. (In Russian) [Google Scholar]
- Protsenko, V.Y.; Borisov, B.A.; Karpun, N.N. The results of evaluating the insecticidal action of some species and strains of entomoparasitic fungi on the marmorated bug (Halyomorpha halys). In New and Unconventional Plants and Prospects of their Application, Proceedings of the XIII International Conference; Sochi, Russia, 4–8 June 2018; RUDN University Publishing: Moscow, Russia, 2018; pp. 591–594. (In Russian) [Google Scholar]
- Kulava, L.D.; Karpun, N.N.; Zhuravleva, E.N.; Shoshina, E.I.; Ayba, L.Y. The effectiveness of pheromones of a brown marmorated stink bug and traps of various designs in mandarin agrocenoses in Abkhazia. Subtrop. Ornam. Hortic. 2021, 77, 161–169. [Google Scholar] [CrossRef]
- Fujisawa, T. Damage and control of the brown marmorated stink bug in apple orchards. Jpn. Agric. Technol. 2001, 45, 42–47. [Google Scholar]
- Willrich, M.M.; Leonard, B.R.; Cook, D.R. Arthropod management laboratory and field evaluations of insecticide toxicity to stink bugs (Heteroptera: Pentatomidae). J. Cotton Sci. 2003, 7, 156–163. Available online: https://www.cotton.org/journal/2003-07/4/upload/jcs07-156.pdf (accessed on 10 January 2022).
- Nielsen, A.L.; Shearer, P.W.; Hamilton, G.C. Toxicity of insecticides to Halyomorpha halys (Hemiptera: Pentatomidae) using glass vial bioassays. J. Econ. Entomol. 2008, 101, 1439–1442. [Google Scholar] [CrossRef]
- Leskey, T.C.; Lee, D.-H.; Short, B.D.; Wright, S.E. Impact of insecticides on the invasive Halyomorpha halys (Hemiptera: Pentatomidae): Analysis of insecticide lethality. J. Econ. Entomol. 2012, 105, 1726–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funayama, K. Control effect on the brown-marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae), by combined spraying of pyrethroid and neonicotinoid insecticides in apple orchards in northern Japan. Appl. Entomol. Zool. 2012, 47, 75–78. [Google Scholar] [CrossRef]
- Mooneyham, K.L.; Aigner, J.D., Jr.; Kuhar, T.P. Control of brown marmorated stink bug with insecticide-treated window screens. Arthropod Manag. Tests 2016, 41, tsw021. [Google Scholar] [CrossRef] [Green Version]
- Kuhar, T.P.; Kamminga, K.L. Review of the chemical control research on Halyomorpha halys in the USA. J Pest. Sci. 2017, 90, 1021–1031. [Google Scholar] [CrossRef]
- Osborn, H.; Drake, C.J. Notes on American Tingidae with descriptions of new species. Ohio J. Sci. 1917, 17, 295–307. [Google Scholar]
- Torres-Miller, L. Additions to the West Virginia tingid fauna (Hemiptera: Heteroptera: Tingidae). Insecta Mundi 1995, 9, 281–282. [Google Scholar]
- Bernardinelli, I.; Zandigiacomo, P. First record of the oak lace bug Corythucha arcuata (Say) (Heteroptera, Tingidae) in Europa. Inf. Fitopatol. 2000, 50, 47–49. [Google Scholar]
- Mutun, S. First report of the oak lace bug, Corythucha arcuata (Say, 1832) (Heteroptera: Tingidae) from Bolu, Turkey. Isr. J. Zool. 2003, 49, 323–324. [Google Scholar]
- Mutun, S.; Ceyhan, Z.; Sözen, C. Invasion by the oak lace bug, Corythucha arcuata (Say) (Heteroptera: Tingidae), in Turkey. Turk. J. Zool. 2009, 33, 263–268. [Google Scholar] [CrossRef]
- Samin, N.; Linnavuori, R.E. A contribution to the Tingidae (Heteroptera) from north and northwestern Iran. Entomofauna. Z. Entomol. 2011, 32, 373–380. [Google Scholar]
- Shchurov, V.I.; Bondarenko, A.S.; Skvortsov, M.M.; Shchurova, A.V. Alien forest insect pests revealed in the Northwest Caucasus in 2010–2016 and consequences of their uncontrolled dispersal. Izv. St.-Peterbg. Lesoteh. Akad. 2017, 220, 212–228. (In Russian) [Google Scholar] [CrossRef]
- Neimorovets, V.V.; Shchurov, V.I.; Bondarenko, A.S.; Skvortsov, M.M.; Konstantinov, F.V. First documented outbreak and new data on the distribution of Corythucha arcuata (Say, 1832) (Hemiptera: Tingidae) in Russia. Acta Zool. Bulg. 2017, 9, 139–142. Available online: https://www.researchgate.net/publication/321496929_First_Documented_Outbreak_and_New_Data_on_the_Distribution_of_Corythucha_arcuata_Say_1832_Hemiptera_Tingidae_in_Russia (accessed on 10 January 2022).
- Karpun, N.N.; Protsenko, V.Y.; Borisov, B.A.; Shiryaeva, N.V. Discovery of oak lace bug Corythucha arcuata (Say) (Heteroptera: Tingidae) in the subtropical zone of the Black Sea coast of the Caucasus. Eurasian Entomol. J. 2018, 17, 113–119. [Google Scholar] [CrossRef]
- Stryukova, N.M.; Omel’yanenko, T.Z.; Golub, V.B. Oak lace bug in the Republic of Crimea. Plant Prot. Quar. 2019, 9, 43–44. Available online: https://www.elibrary.ru/download/elibrary_39322104_97303903.pdf (accessed on 10 January 2022). (In Russian).
- Martynov, V.V.; Nikulina, T.V. Oak lace bug Corythucha arcuata (Say, 1832) (Hemiptera: Tingidae)—A new invasive pest in the forests of the southwestern part of mountain Crimea. Subtrop. Ornam. Hortic. 2020, 72, 124–138. (In Russian) [Google Scholar] [CrossRef]
- Golub, V.B.; Golub, N.V.; Soboleva, V.A. Distribution and trophic relations of the oak lace bug Corythucha arcuata (Say) (Heteroptera: Tingidae) in Crimea. Field Biol. J. 2020, 2, 179–184. Available online: https://www.elibrary.ru/download/elibrary_44184062_33929269.pdf (accessed on 10 January 2022). (In Russian).
- Gninenko, Y.I.; Chernova, U.A.; Gimranov, R.I.; Rakov, A.G.; Khegay, I.V. Oak lace bug expanses its area on the territory of Russia. Plant Prot. Quar. 2020, 10, 37–38. Available online: https://www.elibrary.ru/download/elibrary_43992021_73556954.pdf (accessed on 10 January 2022). (In Russian).
- Blyummer, A.G. Invasive species of not-Arctic lace bugs of the genus Corythucha (Heteroptera, Tingidae) in Eurasia: Features of distribution and injuriousness. In Ecological and Economic Consequences of Invasions of Dendrophilous Insects, Proceedings of the All-Russian Conference with International Participation, Krasnoyarsk, Russia, 25–27 September 2012; Baranchikov, Y.N., Ed.; Forest Institute SB RAS: Krasnoyarsk, Russia, 2012; pp. 139–143. (In Russian) [Google Scholar]
- Drake, C.J.; Ruhoff, F.A. Lacebugs of the World: A Catalog (Hemiptera: Tingidae). Bull. U.S. Natl. Mus. 1965, 243, 1–634. [Google Scholar] [CrossRef]
- Borisov, B.A.; Karpun, N.N.; Bibin, A.R.; Grabenko, Y.A.; Shiryaeva, N.V.; Lyanguzov, M.Y. New data on trophic relations of the invasive oak lace bug Corythucha arcuata (Heteroptera: Tingidae) in the Krasnodar region and in the Republic of Adygea based on the research findings for the year 2018. Subtrop. Ornam. Hortic. 2018, 67, 188–203. (In Russian) [Google Scholar] [CrossRef]
- Karpun, N.N.; Russian Research Institute of Floriculture and Subtropical Crops, Sochi, Russia. Personal communication, 2021.
- Paulin, M.; Hirka, A.; Eötvös, C.B.; Gáspár, C.; Fürjes-Mikó, Á.; Csóka, G. Known and predicted impacts of the invasive oak lace bug (Corythucha arcuata) in European oak ecosystems—A review. Folia Oecologica 2020, 47, 131–139. [Google Scholar] [CrossRef]
- Shchurov, V.I.; Zamotajlov, A.S.; Shchurova, A.V. Assessment of climatic conditions for the expansion of Corythucha arcuata (Say, 1832) (Heteroptera: Tingidae) in the European part of Russia. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 379–380. (In Russian) [Google Scholar]
- Shchurov, V.I.; Zamotajlov, A.S.; Bondarenko, A.S.; Shchurova, A.V.; Skvortsov, M.M.; Glushchenko, L.S. The oak lace bug Corythucha arcuata (Say, 1832) (Heteroptera: Tingidae) in the Northwestern Caucasus: Phenology, biology, monitoring of the territorial expansion, and harmfulness. Izv. St.-Peterbg. Lesoteh. Akad. 2019, 228, 58–87. (In Russian) [Google Scholar] [CrossRef]
- Gninenko, Y.I.; Chernova, U.A.; Nalepin, V.P. Oak lace bug: Stages of formation of the secondary range in Russia. In Innovations in the Conservation and Sustainable Development of Forest Ecosystems, Proceedings of International and Practical Conference Dedicated to the 20th Anniversary of State National Park “Burabay” Creation, Burabay, Kazakhstan, 2–5 September 2020; Bykov, S.V., Ed.; State National Park “Burabay”: Burabay, Kazakhstan, 2020; pp. 66–68. [Google Scholar]
- Kovač, M.; Gorczak, M.; Wrzosek, M.; Tkaczuk, C.; Pernek, M. Identification of entomopathogenic fungi as naturally occurring enemies of the invasive oak lace bug, Corythucha arcuata (Say) (Hemiptera: Tingidae). Insects 2020, 11, 679. [Google Scholar] [CrossRef] [PubMed]
- Borisov, B.A.; AgroBioTechnology Ltd., Moscow, Russia. Personal communication, 2021.
- Chernova, U.A.; Rakov, A.G.; Khegay, I.V. Mixed insecticide against oak lace bug. Plant Prot. Quar. 2019, 11, 51. Available online: https://www.elibrary.ru/download/elibrary_41239385_98490426.pdf (accessed on 10 January 2022). (In Russian).
- Bălăcenoiu, F.; Nețoiu, C.; Tomescu, R.; Simon, D.C.; Buzatu, A.; Toma, D.; Petrițan, I.C. Chemical control of Corythucha arcuata (Say, 1832), an invasive alien species, in oak forests. Forests 2021, 12, 770. [Google Scholar] [CrossRef]
- Besedina, E.N.; Ismailov, V.Y.; Nastasiy, A.S. Field evaluation of the effectiveness of biological and biorational insecticides against oak lace bug Corythucha arcuata Say (Hemiptera, Tingidae). Agrochemistry 2021, 3, 45–50. (In Russian) [Google Scholar] [CrossRef]
- EPPO. Pest Risk Analysis for Agrilus fleischeri. 2019. Available online: https://gd.eppo.int/taxon/AGRLFL/documents (accessed on 2 January 2022).
- Volkovitsh, M.G. On the invasive potential of buprestid beetles (Coleoptera: Buprestidae) damaging woody plants. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 113–116, (In Russian and English). [Google Scholar]
- Volkovitsh, M.G.; Kovalev, A.V.; Orlova-Bienkowskaja, M.J. Current distribution and diagnostic features of two potentially invasive Asian buprestid species: Agrilus mali Matsumura and A. fleischeri Obenberger (Coleoptera: Buprestidae). Insects 2020, 11, 493. [Google Scholar] [CrossRef] [PubMed]
- EPPO. A2 List of Pests Recommended for Regulation as Quarantine Pests (Version 2019-09). Available online: https://www.eppo.int/ACTIVITIES/plant_quarantine/A2_list (accessed on 15 August 2021).
- Obenberger, J. De novis Buprestidarum regionis palaearcticae speciebus V. Cas. Ceskoslovenske Spoiecnosti Entomol. 1925, 22, 30–34. [Google Scholar]
- Kubáň, V.; Jendek, E.; Kalashian, M.Y.; Volkovitsh, M.G. Superfamily Buprestoidea Leach, 1815. In Catalogue of Palaearctic Coleoptera. Scarabaeoidea, Scirtoidea, Dascilloidea, Buprestoidea and Byrrhoidea; Löbl, I., Löbl, D., Eds.; BRILL: Leiden, The Netherlands, 2016; Volume 3, pp. 19–32, 432–574. [Google Scholar]
- Zang, K.; Wang, X.-Y.; Yang, Z.-Q.; Wei, K.; Duan, J.J. Biology and natural enemies of Agrilus fleischeri (Coleoptera: Buprestidae), a newly emerging destructive buprestid pest in Northeast China. J. Asia-Pac. Entomol. 2017, 20, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.S.; Shang, P.H.; Li, F. Preliminary observations on the life history of Agrilus sp. Shaanxi For. Sci. Technol. 1995, 3, 40–42. [Google Scholar]
- Ji, Y.; Ji, R.; Huang, R.-X. Invasive species—Agrilus mali Matsumura and damage in Xinjiang. Xinjiang Agric. Sci. 2004, 41, 31–33. [Google Scholar]
- Cui, X.N.; Liu, D.G.; Li, A.H. Research progress in integrated management of Agrilus mali. Plant Prot. 2015, 41, 16–23. [Google Scholar]
- Bozorov, T.A.; Luo, Z.; Li, X.; Zhang, D. Agrilus mali Matsumura (Coleoptera: Buprestidae), a new invasive pest of wild apple in western China: DNA barcoding and life cycle. Ecol. Evol. 2018, 9, 1160–1172. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.-J.; Zhang, Y.-L.; Zhang, X.; Luo, Z.-H.; Zhang, P.; Golec, J.; Poland, T.M.; Zalucki, M.P.; Han, P.; Lu, Z.-Z. Life history and mortality factors of Agrilus mali Matsumura (Coleoptera: Buprestidae) in wild apples in Northwestern China. Agric. For. Entomol. 2019, 21, 309–317. [Google Scholar] [CrossRef]
- Zhang, P.; Cui, Z.; Xu, H.; Ali, A.; Zhang, X.; Liu, X.; Zhang, Y.; Zhou, X.; Lu, Z. Thirst or malnutrition: The impacts of invasive insect Agrilus mali on the physiological status of wild apple trees. Insects 2020, 11, 440. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, Y.-L.; Zhang, P.; Cui, Z.-J.; Han, P.; Gao, G.-Z.; Poland, T.M.; Zalucki, M.P.; Lu, Z.-Z. Agrilus mali Matsumura (Coleoptera: Buprestidae) density and damage in wild apple Malus sieversii (Rosales: Rosaceae) forests in Central Eurasia under four different management strategies. Entomol. Gen. 2021, 41, 257–266. [Google Scholar] [CrossRef]
- A Unified List of Quarantine Pests of the Eurasian Economic Union. (As Amended by 8 August 2019.) (Decision of the Council of the Eurasian Economic Commission of 8 August 2019 No. 74). Available online: https://vniikr.ru/edinyij-perechen-karantinnyix-obektov-evrazijskogo-ekonomicheskogo-soyuza (accessed on 22 August 2021).
- Agrilus mali. EPPO Global Database. 2021. Available online: https://gd.eppo.int/taxon/AGRLMA (accessed on 22 August 2021).
- Jendek, E.; Grebennikov, V. Agrilus (Coleoptera, Buprestidae) of East Asia; Jan Farkač: Prague, Czech Republic, 2011; p. 362. [Google Scholar]
- Matsumura, S. Life history of Agrilus mali Mats. (Buprestidae). In Bionomics on Apple Trees in China and Korea; Muramatsu, S., Ed.; Korea Agricultural Experiment Station: Seoul, Korea, 1924; pp. 1–21. (In Japanese) [Google Scholar]
- Sokolov, E.A.; Atanov, N.M.; Zhimerikin, V.N.; Gura, N.A.; Komarova, G.F.; Nikritin, L.M.; Shakhramanov, I.K. Quarantine pests with limited distribution on the territory of the Russian Federation. Plant Prot. 1995, 5, 37–41. (In Russian) [Google Scholar]
- Jendek, E.; Poláková, J. Host Plants of World Agrilus (Coleoptera, Buprestidae). A Critical Review; Springer: New York, NY, USA, 2014; pp. 1–706. [Google Scholar]
- Liu, Z.Q.; Chen, W.M.; Xu, Z.; Liang, Q.L. Malus sieversii forest distribution and Agrilus mali Matsumura status of damage in the west part of Tianshan Mountains. North. Hortic. 2014, 17, 121–124. [Google Scholar]
- Li, M.L.; Zhang, Z.Q. Discussion on biology and life history associated with Agrilus mali Matsumura. J. Northwest For. Univ. 2017, 32, 139–146. [Google Scholar]
- Nikritin, L.M.; Shutova, N.N. Apple buprestid. Plant Prot. 1985, 1, 44. (In Russian) [Google Scholar]
- Nikritin, L.M. Apple buprestid. Plant Prot. 1994, 3, 46. (In Russian) [Google Scholar]
- Cao, L.M.; Zhang, Y.L.; van Achterberg, C.; Wang, Z.Y.; Wang, X.Y.; Zhao, W.X.; Yang, Z.Q. Notes on braconid wasps (Hymenoptera, Braconidae) parasitising on Agrilus mali Matsumura (Coleoptera, Buprestidae) in China. ZooKeys 2019, 867, 97–121. [Google Scholar] [CrossRef]
- Fairmaire, L. Notes sur les Coléoptères des environs de Pékin (2e Partie). Revue Ent. Caen 1888, 7, 111–160. [Google Scholar]
- Chamorro, M.L.; Jendek, E.; Haack, R.A.; Petrice, T.R.; Woodley, N.E.; Konstantinov, A.S.; Volkovitsh, M.G.; Yang, X.K.; Grebennikov, V.V. Illustrated Guide to the Emerald Ash Borer, Agrilus planipennis Fairmaire and Related Species (Coleoptera, Buprestidae); Pensoft Publishers: Sofia, Bulgaria; Moscow, Russia, 2015; pp. 1–198. [Google Scholar]
- Baranchikov, Y.; Mozolevskaya, E.; Yurchenko, G.; Kenis, M. Occurrence of the emerald ash borer, Agrilus planipennis in Russia and its potential impact on European forestry. EPPO Bull. 2008, 38, 233–238. [Google Scholar] [CrossRef]
- Musolin, D.L.; Selikhovkin, A.V.; Shabunin, D.A.; Zviagintsev, V.B.; Baranchikov, Y.N. Between ash dieback and emerald ash borer: Two Asian invaders in Russia and the future of ash in Europe. Balt. For. 2017, 23, 316–333. [Google Scholar]
- Orlova-Bienkowskaja, M.J.; Volkovitsh, M.G. Are native ranges of the most destructive invasive pests well known? A case study of the native range of the emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae). Biol. Invasions 2018, 20, 1275–1286. [Google Scholar] [CrossRef]
- Meshkova, V.L.; Kucheryavenko, T.V.; Zinchenko, O.V.; Borysenko, A.I. Beginning of the spread of Agrilus planipennis Fairmaire (Coleoptera: Buprestidae) on the territory of Ukraine. Izv. St.-Peterbg. Lesoteh. Akad. 2021, 236, 163–184. (In Russian) [Google Scholar] [CrossRef]
- Volkovitsh, M.G.; Bieńkowski, A.O.; Orlova-Bienkowskaja, M.J. Emerald ash borer approaches the borders of the European Union and Kazakhstan and is confirmed to infest European ash. Forests 2021, 12, 691. [Google Scholar] [CrossRef]
- Volkovitsh, M.G.; Orlova-Bienkowskaja, M.J.; Kovalev, A.V.; Bieńkowski, A.O. An illustrated guide to distinguish emerald ash borer (Agrilus planipennis) from its congeners in Europe. Forestry 2020, 93, 316–325. [Google Scholar] [CrossRef]
- Haack, R.A.; Jendek, E.; Liu, H.; Marchant, K.R.; Petrice, T.R.; Poland, T.M.; Ye, H. The emerald ash borer: A new exotic pest in North America. Newsl. Mich. Entomol. Soc. 2002, 47, 1–5. Available online: https://www.researchgate.net/publication/228544207_The_Emerald_Ash_Borer_A_New_Exotic_Pest_in_North_America (accessed on 10 January 2022).
- Izhevskii, S.S. Threatening Findings of the Emerald Ash Borer Agrilus planipennis in the Moscow Region. 2007. Available online: http://www.zin.ru/Animalia/Coleoptera/rus/agrplaiz.htm (accessed on 6 July 2021). (In Russian).
- Volkovitsh, M.G. Emerald Ash Borer Agrilus planipennis—New Extremely Dangerous Pest of Ash in the European Part of Russia. 2007. Available online: http://www.zin.ru/Animalia/Coleoptera/rus/eab_2007.htm (accessed on 7 July 2021).
- Emerald Ash Borer Informative Network. Available online: http://www.emeraldashborer.info/ (accessed on 6 July 2021).
- Orlova-Bienkowskaja, M.J.; Drogvalenko, A.N.; Zabaluev, I.A.; Sazhnev, A.S.; Peregudova, E.Y.; Mazurov, S.G.; Komarov, E.V.; Struchaev, V.V.; Martynov, V.V.; Nikulina, T.V.; et al. Current range of Agrilus planipennis Fairmaire, an alien pest of ash trees, in European Russia and Ukraine. Ann. For. Sci. 2020, 77, 1–14. [Google Scholar] [CrossRef]
- Meshkova, V.L.; Skrylnik, Y.Y.; Terekhova, V.V.; Kucheryavenko, T.V. Emerald ash borer (Agrilus planipennis) in Kharkiv region. In Modern Problems of Forestry and Ecology and Ways of Their Solution, Proceedings of the International Scientific and Practical Conference “Faculty of Forestry and Ecology—20 years”, Zhytomyr, Ukraine, 7–8 October 2021; Skidan, O.V., Romanchuk, L.D., Vishnevskiy, A.V., Siruk, Y.V., Kratiuk, O.L., Zhitova, O.P., Andreeva, O.Y., Shvets, M.V., Ishuk, O.V., Eds.; Poliskiy National University: Zhytomyr, Ukraine, 2021; pp. 125–126. [Google Scholar]
- Musolin, D.L.; Selikhovkin, A.V.; Peregudova, E.Y.; Popovichev, B.G.; Mandelshtam, M.Y.; Baranchikov, Y.N.; Vasaitis, R. North-westward expansion of the invasive range of Emerald ash borer, Agrilus planipennis Fairmaire (Coleoptera: Buprestidae) towards the EU: From Moscow to Saint Petersburg. Forests 2021, 12, 502. [Google Scholar] [CrossRef]
- Selikhovkin, A.V.; Musolin, D.L.; Popovichev, B.G.; Merkuryev, S.A.; Volkovitsh, M.G.; Vasaitis, R. Invasive populations of the Emerald ash borer Agrilus planipennis Fairmaire, 1888 (Coleoptera: Buprestidae) in Saint Petersburg, Russia: A hitchhiker? Insects 2022, 13, 191. [Google Scholar] [CrossRef] [PubMed]
- Volkovitsh, M.G.; Mozolevskaya, E.G. The tenth «anniversary» of the invasion of emerald ash borer Agrilus planipennis Fairm. (Coleoptera: Buprestidae) in Russia: Results and prospects. Izv. St.-Peterbg. Lesoteh. Akad. 2014, 207, 8–19. (In Russian) [Google Scholar]
- Baranchikov, Y.N.; Seraya, L.G.; Grinash, M.N. All European ash species are susceptible to emerald ash borer Agrilus planipennis Fairmaire (Coleoptera: Buprestidae)—A Far Eastern invader. Siberian Forest J. 2014, 6, 80–85. Available online: https://www.researchgate.net/publication/272822249_Baranchikov_Yu_N_Seraya_L_G_Grinash_M_N_All_European_ash_species_are_susceptible_to_emerald_ash_borer_Agrilus_planipennis_Fairmaire_Coleoptera_Buprestidae_-_a_Far_Eastern_invader_Sibirskiy_Lesnoy_Zurn (accessed on 10 January 2022). (In Russian).
- Volkovitsh, M.G. Buprestidae. In Inventory of Alien Beetles of European Russia; Orlova-Bienkowskaja, M.J., Ed.; Mukhametov, G.V. Publisher: Livny, Russia, 2019; pp. 84–95. (In Russian) [Google Scholar]
- Volkovitsh, M.G.; Suslov, D.V. The first record of the emerald ash borer, Agrilus planipennis Fairmaire (Coleoptera: Buprestidae), in St. Petersburg signals a real threat to the palace and park ensembles of Peterhof and Oranienbaum. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 119–122, (In Russian and English). [Google Scholar]
- Haubrock, P.J.; Turbelin, A.J.; Cuthbert, R.N.; Novoa, A.; Taylor, N.G.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.W.; Capinha, C.; Diagne, C.; et al. Economic costs of invasive alien species across Europe. NeoBiota 2021, 67, 153–190. [Google Scholar] [CrossRef]
- Gninenko, Y.I.; Kliukin, M.S.; Khegai, I.V. Emerald ash borer: Catastrophe postponed? Plant Health Res. Pract. 2016, 3, 38–45. Available online: https://elibrary.ru/download/elibrary_28379702_35791451.pdf (accessed on 10 January 2022). (In Russian).
- Kirichenko, N.; Haubrock, P.J.; Cuthbert, R.N.; Akulov, E.; Karimova, E.; Shneider, Y.; Liu, C.; Angulo, E.; Diagne, C.; Courchamp, F. Economic costs of biological invasions in terrestrial ecosystems in Russia. NeoBiota 2021, 67, 103–130. [Google Scholar] [CrossRef]
- Commission Delegated Regulation (EU) 2019/1702 of 1 August 2019 Supplementing Regulation (EU) 2016/2031 of the European Parliament and of the Council by Establishing the List of Priority Pests. C/2019/5637. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?qid=1570788497660&uri=CELEX:32019R1702 (accessed on 16 January 2022).
- Orlova-Bienkowskaja, M.J.; Bienkowski, A.O. Low heat availability could limit the potential spread of the emerald ash borer to Northern Europe (Prognosis based on growing degree days per year). Insects 2022, 13, 52. [Google Scholar] [CrossRef] [PubMed]
- EPPO. PM 9/14 (1) Agrilus planipennis: Procedures for official control. Bull. OEPP/EPPO Bull. 2013, 43, 499–509. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Strazanac, J.S.; Marsh, P.M.; Van Achterberg, C.; Choi, W.Y. First recorded parasitoid from China of Agrilus planipennis: A new species of Spathius (Hymenoptera: Braconidae: Doryctinae). Ann. Entomol. Soc. Am. 2005, 98, 636–642. [Google Scholar] [CrossRef] [Green Version]
- Orlova-Bienkowskaja, M.J.; Belokobylskij, S.A. Discovery of the first European parasitoid of the emerald ash borer Agrilus planipennis (Coleoptera: Buprestidae). Eur. J. Entomol. 2014, 111, 594–596. [Google Scholar] [CrossRef]
- Bauer, L.S.; Duan, J.J.; Gould, J.R. Emerald ash borer (Agrilus planipennis Fairmaire) (Coleoptera: Buprestidae). In The Use of Classical Biological Control to Preserve Forests in North America; Van Driesche, R., Reardon, R., Eds.; FHTET-2013-02; U.S. Department of Agriculture, Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2014; pp. 189–209. Available online: https://www.fs.usda.gov/treesearch/pubs/48051 (accessed on 16 January 2022).
- Volkovitsh, M.G.; Zykov, I.E.; Karpun, N.N.; Zakharchenko, V.Y.; and Kovalev, A.V. A description of the larva of the cypress jewel beetle, Lamprodila (Palmar) festiva (L.), with notes on the larval characters of Poecilonotini and Dicercini (Coleoptera, Buprestidae). Entomol. Rev. 2020, 99, 1304–1317. [Google Scholar] [CrossRef]
- Volkovitsh, M.G.; Karpun, N.N. A new invasive species of Buprestid beetles in the Russian fauna: Lamprodila (Palmar) festiva (L.) (Coleoptera, Buprestidae), a pest of Cupressaceae. Entomol. Rev. 2017, 97, 425–437. [Google Scholar] [CrossRef]
- Karpun, N.N.; Gnezdilov, A.A. Why do Arborvitae and False Cypresses Dry Up on the Black Sea Coast? 2016. Available online: http://www.vniisubtrop.ru/novosti/731-pochemu-usykhayut-tui-i-kiparisoviki-na-cheromorskompoberezhe.html (accessed on 3 August 2021). (In Russian).
- Karpun, N.N.; Zhuravleva, Y.N.; Volkovitsh, M.G.; Procenko, V.Y.; Musolin, D.L. To the fauna and biology of new alien insect pest species of woody plants in humid subtropics of Russia. Izv. St.-Peterbg. Lesoteh. Akad. 2017, 220, 169–185. (In Russian) [Google Scholar] [CrossRef]
- Gubin, A.I.; Martynov, V.V.; Nikulina, T.V. The first record of Cypress jewel beetle Lamprodila (Palmar) festiva (Linnaeus, 1767) (Coleoptera, Buprestidae) in the Donbass. Subtrop. Ornam. Hortic. 2020, 75, 96–107. [Google Scholar] [CrossRef]
- Sinelnikov, K.Y. Cypress jewel beetle Lamprodila festiva (L.) on the Southern Coast of the Crimea (2019). Available online: https://vitusltd.ru/blog/lesozaschita/17147 (accessed on 3 August 2021). (In Russian).
- Skvortsov, M.M. The Cypress Jewel Beetle is a New Object of State Forest Pathology Monitoring in Krasnodar Territory. 2017. Available online: http://www.czl23.ru/print.php?news.221 (accessed on 3 August 2021). (In Russian).
- Zhuravleva, Y.N.; Research Centre the Subtropical Scientific Centre of RAS, Sochi, Russia. Personal communication, 2021.
- Wermelinger, B. Der Grüne Wacholder-Prachtkäfer. G’plus Gart.-Fachz. 2011, 3, 30. [Google Scholar]
- Razinger, J.; Zerjav, M.; Modic, S. Thuja occidentalis L. is commonly a host for Cypress jewel beetle (Ovalisia festiva L.) in Slovenia. In Zbornik Predavanj in Referatov 11. Slovenskega Posvetovanja o Varstvu Rastlin z Mednarodno Udeleibo, Proceedings of Conference, Bled, Ljubljana, Slovenia, 5–6 March 2013; Društvo za varstvo rastlin Slovenije: Ljubljana, Slovenia, 2013; pp. 359–365. Available online: http://www.dvrs.bf.uni-lj.si/spvr/2013/61Razinger.pdf (accessed on 18 January 2022).
- Schmidt, G.; Diószegi, M.S.; Szabó, V.; Hrotkó, K. Cypress borer (Lamprodila festiva), a new urban pest in Hungary. In Plants in Urban Areas and Landscape, Proceedings of International Symposium, Nitra, Slovakia, 14–15 May 2014; Slovak University of Agriculture: Nitra, Slovakia, 2014; pp. 32–34. [Google Scholar]
- Thoma, J.; Eickermann, M. Erstauftreten des Wacholderprachtkafers Ovalisia festiva (Linnaeus, 1767) in Luxemburg. Bull. Soc. Nat. Luxemb. 2014, 115, 227–229. [Google Scholar]
- Nitzu, E.; Dobrin, I.; Dumbravă, M.; Gutue, M. The range expansion of Ovalisia festiva (Linnaeus, 1767) (Coleoptera: Buprestidae) in Eastern Europe and its damaging potential for Cupressaceae. Trav. Muséum Natl. Hist. Nat. 2016, 58, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Ruseva, S.; Todorov, I.; Pencheva, A. New data on Ovalisia (Palmar) festiva (Linnaeus) (Coleoptera: Buprestidae) and its natural enemies reported from Bulgaria. Ecol. Montenegrina 2020, 28, 53–60. Available online: https://www.researchgate.net/publication/339539961_New_data_on_Ovalisia_Palmar_festiva_Linnaeus_Coleoptera_Buprestidae_and_its_natural_enemies_reported_from_Bulgaria (accessed on 18 January 2022). [CrossRef] [Green Version]
- Karpun, N.N.; Volkovitsh, M.G. Cypress jewel-beetle Lamprodila (Palmar) festiva (L.) (Coleoptera: Buprestidae)—A new invasive pest on the Black Sea coast of the Caucasus. In The Kataev Memorial Readings–IX. Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems, Proceedings of the International Conference, St. Petersburg, Russia, 23–25 November 2016; Musolin, D.L., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2016; pp. 45–46. Available online: https://www.researchgate.net/publication/310832354_Cypress_jewel-beetle_Lamprodila_Palmar_festiva_L_Coleoptera_Buprestidae_-_a_new_invasive_pest_on_the_Black_Sea_Coast_of_the_Caucasus_in_Russian (accessed on 18 January 2022). (In Russian)
- Volkovitsh, M.G.; Karpun, N.N. Lamprodila (Palmar) festiva (L.), a New Invasive Pest of Cupressaceae in the Russian Fauna (Coleoptera: Buprestidae: Poecilonotini). 2016. Available online: http://www.zin.ru/animalia/coleoptera/pdf/Volkovitsh_Karpun_2016_Lamprodila_festiva.pdf (accessed on 5 August 2021). (In Russian).
- Pfeffer, A. Zentral- und Westpaläarktishe Borken- und Kernkäfer (Coleoptera: Scolytidae, Platypodidae). Entomol. Basiliensia 1994, 17, 5–310. [Google Scholar]
- Knížek, M. Curculionidae: Scolytinae, In Catalogue of Palaearctic Coleoptera; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2011; Volume 7, Curculionoidea I; pp. 86–88, 204–251. [Google Scholar]
- Økland, B.; Flø, D.; Schroeder, M.; Zach, P.; Cocos, D.; Martikainen, P.; Siitonen, J.; Mandelshtam, M.Y.; Musolin, D.L.; Neuvonen, S.; et al. Range expansion of the small spruce bark beetle Ips amitinus: A newcomer in northern Europe. Agric. For. Entomol. 2019, 21, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Mandelshtam, M.Y.; Selikhovkin, A.V. Bark and ambrosia beetles (Coleoptera, Curculionidae: Scolytinae) of Northwest Russia: History of the study, composition and genesis of the fauna. Entomol. Rev. 2020, 100, 800–826. [Google Scholar] [CrossRef]
- Mandelshtam, M.Y.; Selikhovkin, A.V. Comparative characteristics of species richness of the bark beetles (Coleoptera: Curculionidae, Scolytinae) fauna in north-west subregions of Russia. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 214–215. (In Russian) [Google Scholar]
- Mandelshtam, M.Y.; Musolin, D.L.; Økland, B.; Flø, D.; Schroeder, M.; Zach, P.; Cocos, D.; Martikainen, P.; Siitonen, J.; Neuvonen, S.; et al. Range expansion of the small spruce bark beetle Ips amitinus (Coleoptera, Curculionidae: Scolytinae) in Northern Europe and Western Siberia. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 211–212. (In Russian) [Google Scholar]
- Mikutowicz, J.M. Zur Koleopterenfauna der Ostseeprovinzen Russlands. Korresp.-Bl. Des Nat.-Ver. Zu Riga 1905, 48, 73–92. [Google Scholar]
- Leius, K. Taiendavaid andmeid kodumaa urasklaste (Ipidae) fauna kohta. Erganzende Angaben uber die Borkenkafer-Fauna (Ipidae) in Estland (Zusammenfassung). Eest. Metsanduse Aastaraam. (Estländisches Forstwirtsch. Jahrb. Tartu) 1939, 9, 317–328. (In Estonian) [Google Scholar]
- Voolma, K.; Estonian University of Life Sciences, Tartu, Estonia. Personal communication, 2021.
- Zolk, K. Kodumaa ürasklased (Ipidae) ühes lühikese ülevaatega nende bionoomiast ja levimisest Eestis [Die Borkenkäfer (Ipidae) Estlands mit kurzer Berücksichtigung ihrer Bionomie und Verbreitung]. Eest. Metsanduse Aastaraam. (Estländisches Forstwirtsch. Jahrb. Tartu) 1932, 6, 127–176. (In Estonian) [Google Scholar]
- Zolk, K. Metsakahjurite esinemine Eestis 1934 [Das Vorkommen der Forstschädlinge in Estland im Jahre 1934]. Eest. Metsanduse Aastaraam. (Estländisches Forstwirtsch. Jahrb. Tartu) 1935, 7, 614–638. (In Estonian) [Google Scholar]
- Nuorteva, M. Fennoskandialle uusi kaarnakuoriaislaji Ips amitinus Eichh. tavattu Suomesta [Ips amitinus Eichh. (Col., Scolytidae) neu für Fennoscandien]. Ann. Entomol. Fenn. 1955, 21, 30–32. [Google Scholar]
- Koponen, M. Distribution of Ips amitinus Eichh. (Coleoptera, Scolytidae) in Finland in 1950–1973. Ann. Entomol. Fenn. 1975, 41, 65–69. [Google Scholar]
- Koponen, M. Distribution of Ips amitinus Eichh. (Coleoptera, Scolytidae) in Finland in 1974–1979. Not. Entomol. 1980, 60, 223–225. [Google Scholar]
- Muona, J.; Viramo, J. The Coleoptera of the Koillismaa area (Ks), North-East Finland. Oulanka Rep. 1986, 6, 3–50. [Google Scholar]
- Hansen, V. Catalogus Coleopterorum Fennoscandiae et Daniae; Lindroth, C.H., Ed.; Entomologiska Sällskapet i Lund: Lund, Sweden, 1960; pp. 1–476. [Google Scholar]
- Lindelöw, Å. Väntad barkborre funnen i Sverige-fynd av Ips amitinus (Coleoptera, Scolytinae) [Ips amitinus (Coleoptera, Scolytinae) expected and found in Sweden]. Entomol. Tidskr. 2013, 134, 203–206. [Google Scholar]
- Stark, V.N. Materials to knowledge of bark beetles of Briansk Gouvernement. Zashchita Rasteniy [Plant Protection]. 1926, III, 330–339. (In Russian) [Google Scholar]
- Mandelshtam, M. Notes on the current status of Ips amitinus Eichh. (Coleoptera, Scolytidae) in North-West Russia. Entomol. Fenn. 1999, 10, 29–34. [Google Scholar] [CrossRef]
- Voolma, K.; Mandelshtam, M.Y.; Shcherbakov, A.N.; Yakovlev, E.B.; Õunap, H.; Süda, I.; Popovichev, B.G.; Sharapa, T.V.; Galasjeva, T.V.; Khairetdinov, R.R.; et al. Distribution and spread of bark beetles (Coleoptera: Scolytidae) around the Gulf of Finland: A comparative study with notes on rare species of Estonia, Finland and North-Western Russia. Entomol. Fenn. 2004, 15, 198–210. [Google Scholar] [CrossRef]
- Titova, E.V. Bark beetles associated with coniferous coppice in forest clearings of Karelia. In Studies of Forest Regeneration in Karelia, Proceedings of the Karelian Branch of the USSR Academy of Sciences, Petrozavodsk, Russia, 1959; Shiperovich, V.Y., Kishenko, T.I., Lisenkov, A.F., Eds.; Karelian Branch of the USSR Academy of Sciences: Petrozavodsk, Russia, 1959; Volume 16, pp. 110–126. (In Russian) [Google Scholar]
- Yakovlev, E.B.; Shorokhov, V.V.; Gorbunova, V.N. Materials on the fauna of xylophagous beetles of Karelia. In The Fauna and Ecology of Arthropods in Karelia; Yakovlev, E.B., Uzenbaev, S.D., Eds.; Forest Institute: Petrozavodsk, Russia, 1986; pp. 40–43. (In Russian) [Google Scholar]
- Mozolevskaya, E.G.; Sharapa, T.V. Species composition of xylophagous insects of Murmansk Province. Entomol. Obozr. 1996, 75, 558–566. (In Russian) [Google Scholar]
- Shcherbakov, A.N.; Nikitsky, N.B.; Polevoi, A.V.; Humala, A.E. On the fauna of beetles (Insecta, Coleoptera) of Pasvik Nature Reserve. Lesn. Vestn. 2013, 6, 16–21. (In Russian) [Google Scholar]
- Mandelshtam, M.Y.; Musolin, D.L. Bark beetle Ips amitinus (Eichhoff, 1872) (Coleoptera: Curculionidae: Scolytinae) continues to expand its range in north-western and northern Russia. In Monitoring and Biological Control Methods of Woody Plant Pests and Pathogens: From Theory to Practice, Proceedings of International Conference, Moscow, Russia, 18–22 April 2016; Baranchikov, Y.N., Ed.; SIF SN RASc.: Moscow, Russia; Krasnoyarsk, Russia, 2016; pp. 129–130. (In Russian) [Google Scholar]
- Mandelshtam, M.Y.; Khairetdinov, R.R. Additions to the check-list of bark beetles (Coleoptera, Curculionidae: Scolytinae) from Leningrad province, Russia. Entomol. Rev. 2017, 97, 893–899. [Google Scholar] [CrossRef]
- Kerchev, I.A.; Mandelshtam, M.Y.; Krivets, S.A.; Ilinsky, Y.Y. Small spruce bark beetle Ips amitinus (Eichhoff, 1872) (Coleoptera, Curculionidae: Scolytinae): A new alien species in Western Siberia. Entomol. Rev. 2019, 99, 639–644. [Google Scholar] [CrossRef]
- Skorokhodov, S.N. Save the neighboring stone pine woods. Yashkinsky Vestn. 2017, 37, 8564. (In Russian) [Google Scholar]
- Krivets, S.A. Problems of protection of Siberian stone pine forests from the small spruce bark beetle Ips amitinus (Coleoptera: Curculionidae, Scolytinae). In Dendrobiotic Invertebrates and Fungi and Their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 183–184. (In Russian) [Google Scholar]
- Kerchev, I.A.; Krivets, S.A.; Smirnov, N.A. Analysis of the population characteristics of the small spruce bark beetle Ips amitinus (Coleoptera: Curculionidae: Scolytinae) in the areas of invasion in Western Siberia. In Dendrobiotic Invertebrates and Fungi and Their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 170–171. (In Russian) [Google Scholar]
- Kerchev, I.A.; Krivets, S.A. An attack of Ips amitinus (Coleoptera: Curculionidae: Scolytinae) on arboretum in West Siberia: New host of invasive bark beetle among exotic conifers. J. Asia-Pac. Entomol. 2021, 24, 148–152. [Google Scholar] [CrossRef]
- Kerchev, I.A.; Krivets, S.A.; Bisirova, E.M.; Smirnov, N.A. Distribution of the small spruce bark beetle Ips amitinus (Eichhoff, 1872) in the Western Siberia. Russ. J. Biol. Invasions 2021, 4, 77–84. (In Russian) [Google Scholar] [CrossRef]
- Holuša, J.; Lukasova, K.; Grodzki, W.; Kula, E.; Matousek, P. Is Ips amitinus (Coleoptera: Curculionidae) abundant in wide range of altitudes? Acta Zool. Bulg. 2012, 64, 219–228. Available online: https://www.researchgate.net/publication/236260984_Is_Ips_amitinus_Coleoptera_Curculionidae_Abundant_in_Wide_Range_of_Altitudes (accessed on 18 January 2022).
- Bisirova, E.M.; Kerchev, I.A. Assessment of the state of the Siberian stone pine Pinus sibirica in the outbreak foci of the small spruce bark beetle Ips amitinus (Coleoptera: Curculionidae, Scolytinae) a new invasive pest in Siberia. In Dendrobiotic Invertebrates and Fungi and Their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 82–83. (In Russian) [Google Scholar]
- Baranchikov, Y.N.; Krivets, S.A. About professional skills in determination of insects: How the occurrence of new aggressive invasive fir pest in Siberia was missed. Ecol. South. Sib. Adjac. Territ. 2010, 1, 50–52. (In Russian) [Google Scholar]
- Baranchikov, Y.N.; Pet’ko, V.M.; Astapenko, S.A.; Akulov, E.N.; Krivets, S.A. Four-eyed fir bark beetle as a new aggressive pest of fir in Siberia. Lesn. Vestn. 2011, 80, 78–81. (In Russian) [Google Scholar]
- Kurentsov, A.I. The Bark Beetles of the USSR Far East; Academy of Sciences of the USSR: Moscow, Russia; St. Petersburg, Russia, 1941; pp. 1–234. (In Russian) [Google Scholar]
- Krivolutskaya, G.O. Fam. Scolytidae—The bark beetles. In Key for Identification of Insects of the Russian Far East; Ler, P.A., Ed.; Dal’nauka: Vladivostok, Russia, 1996; Volume 3, Part 3. Coleoptera, pp. 312–373. (In Russian) [Google Scholar]
- Kerchev, I.A. Ecology of four-eyed fir bark beetle Polygraphus proximus Blandford (Coleoptera: Curculionidae, Scolytinae) in the West Siberian region of invasion. Russ. J. Biol. Invasion 2014, 5, 176–185. [Google Scholar] [CrossRef]
- Chilakhsaeva, E.A. First record of Polygraphus proximus (Coleoptera: Scolytidae) in Moscow Province. Bull. Mosc. Soc. Nat. Biol. 2008, 113, 39–42. (In Russian) [Google Scholar]
- Baranchikov, Y.N.; Seraya, L.G. For whom the bell tolls: Evaluation of potential invasive organisms and their role in regional biota. In Monitoring and Biological Control Methods of Woody Plant Pests and Pathogens: From Theory to Practice. Proceedings of International Conference, Moscow, Russia, 18–22 April 2016; Baranchikov, Y.N., Ed.; SIF SN RASc.: Moscow, Russia; Krasnoyarsk, Russia, 2016; pp. 25–26. (In Russian) [Google Scholar]
- Mandelshtam, M.Y.; Popovichev, B.G. Annotated list of bark beetles (Coleoptera, Scolytidae) of Leningrad Province. Entomol. Rev. 2000, 80, 887–903. [Google Scholar]
- Baranchikov, Y.N.; Efremenko, A.A.; Demidko, D.A.; Titova, V.V. Four-eyed fir bark beetle Polygraphus proximus Blandford (Coleoptera: Curculionidae, Scolytinae) in the western piedmont of the Ural Mountains: Where, whence and when? In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 74–75. (In Russian) [Google Scholar]
- Dedyukhin, S.V.; Titova, V.V. Finding of the bark beetle Polygraphus proximus Blandford, 1894 (Coleoptera, Curculionidae: Scolytinae) in Udmurtia. Russ. J. Biol. Invasions 2021, 12, 258–263. [Google Scholar] [CrossRef]
- Krivets, S.A.; Kerchev, I.A.; Kizeev, Y.M.; Kozhurin, M.A.; Kozak, R.G.; Filimonov, M.N.; Chemodanov, A.V.; Chugin, B.S. Four-eyed fir bark beetle Polygraphus proximus Blandf. (Coleoptera, Scolytidae) in fir forests of Tomsk Oblast. In Diseases and Pests in the Forests of Russia: XXI Century, Proceedings of the All-Russian Conference and V Annual Kataev Memorial Reading, Yekaterinburg, Russia, 20–25 September 2011; Selikhovkin, A.V., Shavnin, S.A., Zalesov, S.V., Onuchin, A.A., Isaev, A.S., Mozolevskaya, E.G., Ponomarev, V.I., Baranchikov, Y.N., Petrova, I.V., Meshkova, V.L., et al., Eds.; Institute of Forest, SB RAS: Krasnoyarsk, Russia, 2011; pp. 53–55. (In Russian) [Google Scholar]
- Krivets, S.A.; Bisirova, E.M.; Kerchev, I.A.; Pashenova, N.V.; Demidko, D.A.; Petko, V.M.; Baranchikov, Y.N. Four-eyed Fir Bark Beetle in Siberian Forests (Distribution, Biology, Ecology, Detection and Survey of Damaged Stands). Manual; Umium Publisher: Tomsk, Russia; Krasnoyarsk, Russia, 2015; pp. 1–48. (In Russian) [Google Scholar]
- Krivets, S.A.; Bisirova, E.M.; Kerchev, I.A.; Pats, E.N.; Chernova, N.A. Transformation of taiga ecosystems in the Western Siberian invasion focus of four-eyed fir bark beetle Polygraphus proximus Blandford (Coleoptera: Curculionidae, Scolytinae). Russ. J. Biol. Invasions 2015, 6, 94–108. [Google Scholar] [CrossRef]
- Krivets, S.A.; Kerchev, I.A.; Bisirova, E.M.; Demidko, D.A.; Pet’ko, V.M.; Baranchikov, Y.N. Distribution of the four-eyed fir bark beetle Polygraphus proximus Blandf. (Coleoptera, Curculionidae: Scolytinae) in Siberia. Izv. St.-Peterbg. Lesoteh. Akad. 2015, 211, 33–45. (In Russian) [Google Scholar]
- Krivets, S.A.; Kerchev, I.A.; Bisirova, E.M.; Debkov, N.M. Current distribution and forecasted invasive area expansion of the four-eyed fir bark beetle Polygraphus proximus Blandford, 1894 in Tomsakaya Oblast’ (Western Siberia). Eurasian Entomol. J. 2018, 17, 53–60. (In Russian) [Google Scholar]
- Bystrov, S.O.; Antonov, I.A. First record of the four-eyed beetle Polygraphus proximus Blandford, 1894 (Coleoptera: Curculionidae, Scolytinae) from Irkutsk Province, Russia. Entomol. Rev. 2019, 98, 54–55. [Google Scholar] [CrossRef]
- Baranchikov, Y.N.; Demidko, D.A.; Laptev, A.B.; Pet’ko, V.M. The dynamics of Siberian fir death in the outbreak focus of four-eyed fir bark beetle. Lesn. Vestn. 2014, 18, 78–81. (In Russian) [Google Scholar]
- Demidko, D.A. The dates of invasion of four-eyed fir bark beetle Polygraphus proximus Blandford (Coleoptera: Curculionidae, Scolytinae) in Tomsk Oblast. Izv. St.-Peterbg. Lesoteh. Akad. 2014, 207, 225–234. Available online: https://www.researchgate.net/publication/273321170_Dating_of_Four-Eyed_Fir_Bark_Beetle_Polygraphus_proximus_Blandford_Coleoptera_Curculionidae_Scolytinae_Invasion_into_Tomsk_Region (accessed on 18 January 2022). (In Russian).
- Bykov, R.; Kerchev, I.; Demenkova, M.; Ryabinin, A.; Ilinsky, Y. Sex-specific Woldbachia infection patterns in populations of Polygraphus proximus Blandford (Coleoptera: Curculionidae: Scolytinae). Insects 2020, 11, 547. [Google Scholar] [CrossRef]
- Kononov, A.; Ustyantsev, K.; Blinov, A.; Fet, V.; Baranchikov, Y.N. Genetic diversity of aboriginal and invasive populations of four-eyed fir bark beetle Polygraphus proximus Blandford (Coleoptera, Curculionidae, Scolytinae). Agric. For. Entomol. 2016, 18, 294–301. [Google Scholar] [CrossRef]
- Kerchev, I.A. Experimental study of the possibility of the emergence of new trophic connections of four-eyed fir bark beetle Polygraphus proximus Blandf. (Coleoptera: Curculionidae, Scolytinae) in the West Siberian region. Vestn. Tomsk. Gos. Univ. Biol. (Tomsk State Univ. J. Biol.) 2012, 3, 169–177. (In Russian) [Google Scholar]
- Pashenova, N.V.; Kononov, A.V.; Ustyantsev, K.V.; Blinov, A.G.; Pertsovaya, A.A.; Baranchikov, Y.N. Ophiostomatoid fungi associated with the four-eyed fir bark beetle on the territory of Russia. Russ. J. Biol. Invasion 2018, 9, 63–74. [Google Scholar] [CrossRef]
- Pashenova, N.V.; Petko, V.M.; Babichev, N.S.; Kerchev, I.A. Transfer of ophiostomatoid fungi by a four-eyed fir bark beetle Polygraphus proximus Blandf. (Coleoptera, Scolytidae) in Siberia. Izv. St.-Peterbg. Lesoteh. Akad. 2012, 210, 114–120. Available online: https://www.researchgate.net/publication/286452586_Transfer_of_Ophiostomatales_fungi_by_Polygraphus_proximus_Blandford_Coleoptera_Sco_lytidae_in_Sibera (accessed on 20 January 2022). (In Russian).
- Pashenova, N.V.; Demidko, D.A.; Pertsovaya, A.A.; Baranchikov, Y.N. Phloem sensibility of Siberian conifers towards Grosmannia aoshimae (Ohtaka, Masuya & Yamaoka) Masuya & Yamaoka, a symbiotic fungus of the four-eyed fir bark beetle. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 247–248. (In Russian) [Google Scholar]
- Bisirova, E.M.; Krivets, S.A. Dynamics of the state of Siberian fir tree stands damaged by the four-eyed fir bark beetle Polygraphus proximus Blandf. in Tomsk Oblast. Vestn. Tomsk. Gos. Univ. Biol. (Tomsk State Univ. J. Biol.) 2018, 44, 118–140. (In Russian) [Google Scholar] [CrossRef] [PubMed]
- Mukhortova, L.V.; Sergeeva, O.V.; Demidko, D.A.; Krivobokov, L.V.; Baranchikov, Y.N. Dynamics of coarse woody debris stocks in the fir forests damaged by the bark beetle Polygraphus proximus Blandf. (Coleoptera: Curculionidae, Scolytinae). In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 231–232. (In Russian) [Google Scholar]
- Kerchev, I.A.; Krivets, S.A. Composition and number of xylophagous consort’s members of Siberian fir associated with four-eyed fit bark beetle in Tomsk Oblast. In Environmental and Economic Consequences of Invasions of Dendrophilous Insects, Proceedings of All-Russian Conference with International Participation, Krasnoyarsk, Russia, 25–27 September 2012; Baranchikov, Y.N., Ed.; Institute of Forest, SB RAS: Krasnoyarsk, Russia, 2012; pp. 57–59. (In Russian) [Google Scholar]
- Bisirova, E.M.; Kerchev, I.A.; Krivets, S.A.; Pats, E.N. The state of fir forests of Tashtagol District of Kemerovo Province, Russia, damaged by the four-eyed fir bark beetle Polygraphus proximus Blandf. (Coleoptera: Curculionidae, Scolytinae). In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 84–85. (In Russian) [Google Scholar]
- Krivets, S.A.; Kerchev, I.A. Insects inhabiting the galleries of the four-eyed fir bark beetle Polygraphus proximus Blandf. (Coleoptera, Curculionidae: Scolytinae) in Siberia. Entomol. Rev. 2016, 96, 545–558. [Google Scholar] [CrossRef]
- Krivosheina, M.G.; Krivosheina, N.P.; Kerchev, I.A. Flies (Diptera) associated with Polygraphus proximus Blandford, 1894 (Coleoptera, Curculionidae) in Siberia and the Russian Far East. Entomol. Rev. 2018, 98, 156–164. [Google Scholar] [CrossRef]
- Kerchev, I.A.; Kryukov, V.Y.; Yaroslavtseva, O.N.; Polovinko, G.P.; Tokarev, Y.S.; Glupov, V.V. The first data on fungal pathogens (Ascomycota, Hypocreales) in the invasive populations of four-eyed fir bark beetle Polygraphus proximus Blandf. Russ. J. Biol. Invasions 2017, 8, 34–40. [Google Scholar] [CrossRef]
- Efremenko, A.A.; Demidko, D.A.; Baranchikov, Y.N. An old remedy for a new problem: Submersion of logs to prevent adult emergence of the invasive bark beetle Polygraphus proximus Blandford (Coleoptera: Curculionidae, Scolytinae). In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; p. 159. (In Russian) [Google Scholar]
- Seraya, L.G.; Pashenova, N.V.; Demidko, D.A.; Kozhenkova, A.A.; Efremenko, A.A.; Gninenko, Y.I.; Baranchikov, Y.N. Attempts of chemical control of invasive populations of the bark beetle Polygraphus proximus (Coleoptera: Curculionidae). In The Kataev Memorial Readings—X, Proceedings of the International Conference, St. Petersburg, Russia, 22–25 October 2018; Musolin, D.L., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2018; Volume 1, pp. 97–98. (In Russian) [Google Scholar]
- Xiang, Q.P.; Wei, R.; Shao, Y.Z.; Yang, Z.Y.; Wang, X.Q.; Zhang, X.C. Phylogenetic relationships, possible ancient hybridization, and biogeographic history of Abies (Pinaceae) based on data from nuclear, plastid, and mitochondrial genomes. Mol. Phylogenet. Evol. 2015, 82, 1–14. [Google Scholar] [CrossRef]
- Kirpichnikova, V.A. Pyralidae. In Key for Identification of Insects of the Russian Far East; Ler, P.A., Ed.; Dal’nauka: Vladivostok, Russia, 2005; Volume 5, Part 2, pp. 526–539. (In Russian) [Google Scholar]
- Mally, R.; Nuss, M. Phylogeny and nomenclature of the box tree moth, Cydalima perspectalis (Walker, 1859) comb. n., which was recently introduced into Europe (Lepidoptera: Pyraloidea: Crambidae: Spilomelinae). Eur. J. Entomol. 2010, 107, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Kruger, E.O. Glyphodes perspectalis (Walker, 1859)—Neu für die Fauna Europas (Lepidoptera: Crambidae). Entomol. Z. 2008, 118, 81–83. [Google Scholar]
- USDA Confirms Box Tree Moth and Takes Action to Contain and Eradicate the Pest. 2021. Available online: https://www.aphis.usda.gov/aphis/newsroom/stakeholder-info/sa_by_date/sa-2021/sa-05/box-tree-moth (accessed on 2 January 2022).
- Gninenko, Y.I.; Shiryaeva, N.V.; Shchurov, V.I. Box tree moth—A new invasive organism in the forests of the Russian Caucasus. Plant Quar. Sci. Prac. 2014, 7, 32–36. Available online: https://elibrary.ru/download/elibrary_25051637_65094735.pdf (accessed on 18 January 2022).
- Karpun, N.N.; Ignatova, Y.A. The first report about Cydalima perspectalis Walker on Black Sea Coast of Russia. In Zprávy Vědecké Ideje—2013 (19), Proceedings of the IX International Scientific and Practical Conference, Prague, Czech Republic, 27 October—5 November 2013; Publishing House “Education and Science” s.r.o.: Prague, Czech Republic, 2013; pp. 29–32. Available online: http://www.rusnauka.com/31_NNM_2013/Biologia/7_146134.doc.htm (accessed on 18 January 2022).
- Proklov, V.V.; Karayeva, S.Z. New and interesting Lepidoptera records from Chechen Republic (Russia). Cauc. Entomol. Bull. 2013, 9, 281–282. Available online: https://www.ssc-ras.ru/files/files/11_%20Proklov.pdf (accessed on 18 January 2022). [CrossRef]
- Karpun, N.N.; Ignatova, Y.A.; Zhuravleva, E.N. New species of harmful entomofauna on ornamental woody plants in humid subtropics of Krasnodar region. Izv. St.-Peterbg. Lesoteh. Akad. 2015, 211, 187–203. Available online: https://elibrary.ru/download/elibrary_23815865_37414681.pdf (accessed on 18 January 2022). (In Russian).
- Shchurov, V.I.; Kuchmistaya, E.V.; Vibe, E.N.; Bondarenko, A.S.; Skvortsov, M.M. The box-tree moth Cydalima perspectalis (Walker, 1859)—The real threat to biological diversity of natural forests of the Northwest Caucasus. Proc. Kuban State Agrar. Univ. 2015, 53, 178–198. (In Russian) [Google Scholar]
- Balykina, E.B.; Trikoz, N.N. Changes in the taxonomic structure of the ornamental plants phytophages complex in Crimean parks. Veles 2017, 43, 59–63. [Google Scholar]
- Dobronosov, V.V. New Data on the Box Tree Moth Cydalima perspectalis (Walker, 1859) in the Central Caucasus. 2017. Available online: http://aeconomy.ru/news/agro/novye-dannye-o-samshitovoy-ognevke-.html (accessed on 2 January 2022). (In Russian).
- Poltavskiy, A.N. Annotated catalog of insect pests of introduced plants in the Botanical Garden of the Southern Federal University. Proc. Bot. Gard. South. Fed. Univ. 2017, 2, 120–138. (In Russian) [Google Scholar]
- Tuniyev, B.S.; Aliev, K.U.; Timukhin, I.N. Colchis boxwood and the new pest Cydalima perspectalis Walker, 1859 (Lepidoptera, Crambidae) in South Ossetia. Bot. Her. North Cauc. 2018, 1, 30–36. Available online: http://gorbotsad.ru/files/Tuniev_2018_1.pdf (accessed on 18 January 2022). (In Russian).
- Il’ina, E.V.; Gasanova, N.M.-S. New Information about Invasive Insects in the Republic of Dagestan. Plant Prot. Quar. 2020, 4, 39–40. Available online: https://elibrary.ru/download/elibrary_42673425_76973702.pdf (accessed on 18 January 2022). (In Russian).
- Rozhina, V.I.; Volodina, L.S.; Drotikova, A.V. First detection of Cydalima perspectalis (Walker, 1859) (Lepidoptera: Crambidae: Pyraustinae) in Kaliningrad region. In ChemBioSeasons, Proceedings of the Abstracts of the Forum of Young Researchers, Kaliningrad, Russia, 23 April 2021; Baltic Federal University Named after I. Kant: Kaliningrad, Russia, 2021; pp. 67–68. Available online: https://elibrary.ru/item.asp?id=46359179 (accessed on 4 January 2022).
- Cydalima perspectalis. EPPO Global Database. 2021. Available online: https://gd.eppo.int/taxon/DPHNPE (accessed on 1 July 2021).
- Karpun, N.N.; Trochov, E.S.; Ignatova, Y.A.; Zhuravleva, E.N.; Kaurova, Z.G. Analysis of the food specialization of the box tree moth (Cydalima perspectalis Walker). Regul. Issues Vet. Med. 2015, 4, 173–176. Available online: https://elibrary.ru/item.asp?id=24825731 (accessed on 18 January 2022).
- Kawazu, K.; Honda, H.; Nakamura, S.; Adati, T. Identification of sex pheromone components of the box tree pyralid, Glyphodes perspectalis. J. Chem. Ecol. 2007, 33, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Park, I.-K. Female sex pheromone components of the box tree pyralid, Glyphodes perspectalis, in Korea: Field test and development of film-type lure. J. Asia-Pacific Entomol. 2013, 16, 473–477. [Google Scholar] [CrossRef]
- Nesterenkova, A.E.; Ponomarev, V.L.; Karpun, N.N.; Protsenko, V.E.; Glebov, V.E.; Danilenko, E.A.; Rastegaeva, V.M. Field testing of biological activity of pheromone box tree moth Cydalima perspectalis Walker. Lesn. Vestn. 2017, 21, 70–77. (In Russian) [Google Scholar] [CrossRef]
- Göttig, S.; Herz, A. The box tree pyralid Cydalima perspectalis: New results of the use of biological control agents and pheromone traps in the field. J. Plant Dis. Prot. 2014, 121, 98–99. [Google Scholar]
- Göttig, S.; Herz, A. Susceptibility of the box tree pyralid Cydalima perspectalis Walker (Lepidoptera: Crambidae) to potential biological control agents Neem (NeemAzal®-T/S) and entomopathogenic nematodes (Nemastar®) assessed in laboratory bioassays and field trials. J. Plant Dis. Prot. 2018, 125, 365–375. [Google Scholar] [CrossRef]
- Borisov, B.A.; Karpun, N.N.; Zhuravleva, E.N.; Borisova, I.P. Evaluation of the possibility of biological control of box tree moth (Cydalima perspectalis) by entomoparasitic fungi. In Monitoring and Biological Control Methods of Woody Plant Pests and Pathogens: From Theory to Practice, Proceedings of International Conference, Moscow, Russia, 18–22 April 2016; Baranchikov, Y.N., Ed.; SIF SN RASc.: Moscow, Russia; Krasnoyarsk, Russia, 2016; pp. 41–43. (In Russian) [Google Scholar]
- Kenis, M.; Nacambo, S.; Leuthardt, F.L.G.; Di Domenico, F.; Haye, T. The box tree moth, Cydalima perspectalis, in Europe: Horticultural pest or environmental disaster? Aliens 2013, 33, 38–41. Available online: https://www.researchgate.net/publication/271483672_The_box_tree_moth_Cydalima_perspectalis_in_Europe_horticultural_pest_or_environmental_disaster (accessed on 18 January 2022).
- Agasieva, I.S.; Ismailov, V.Y.; Fedorenko, E.V.; Nefedova, M.V. Biological control of box tree moth. Plant Prot. Quar. 2017, 8, 21–23. Available online: https://elibrary.ru/download/elibrary_29879952_31932504.pdf (accessed on 18 January 2022).
- Fora, C.G.; Sasu, L.; Poşta, D.; Berar, C. Chemical possibilities of Cydalima perspectalis Walk. (Lepidoptera: Crambidae) control. J. Hortic. For. Biotechnol. 2016, 20, 31–34. Available online: https://journal-hfb.usab-tm.ro/romana/2016/Lucrari%20PDF/Vol%2020(3)%20PDF/7Fora%20George.pdf (accessed on 18 January 2022).
- Somsai, A.P.; Lukacs, L.; Florian, T.; Sestraş, A.F.; Bunea, C.I.; Hoble, A.; Vlaşin, H. Chemical control of box tree moth Cydalima perspectalis (Walker, 1859) (Lepidoptera: Crambidae) in North-Western Romania. Agricultura 2019, 111, 210–216. [Google Scholar] [CrossRef]
- Plugutar, Y.V.; Sharmagy, A.K.; Shishkin, V.A. Effectiveness of bioinsecticides against the caterpillar of Cydalima perspectalis (Walker, 1859) on the Southern Coast of the Crimea. Bull. State Nikitsk. Bot. Gard. 2020, 137, 7–15. (In Russian) [Google Scholar] [CrossRef]
- Baryshnikova, S.V. Gracillariidae. In Catalogue of the Lepidoptera of Russia, 2nd ed.; Sinev, S.Y., Ed.; Zoological Institute RAS: St. Petersburg, Russia, 2019; pp. 36–43. (In Russian) [Google Scholar]
- De Prins, J.; De Prins, W. Global Taxonomic Database of Gracillariidae (Lepidoptera). World Wide Web Electronic Publication. 2006–2021. Available online: http://www.gracillariidae.net) (accessed on 10 November 2021).
- Caradja, A. Beitrag zur Kenntnis der geographischen Verbreitungder Mikrolepidopteren des palaearktischen Faunengebietes nebst Beschreibung neuer Formen. Dt. Ent. Z. Ires 1920, 34, 75–179. [Google Scholar]
- Baryshnikova, S.V.; Dubatolov, V.V. To the knowledge of small moths (Microlepidoptera) of the Bolshekhekhtsirskii Nature Reserve (Khabarovsk District). 2nd report. Bucculatricidae, Gracillariidae, Lyonetiidae]. Anim. World Far East 2007, 6, 47–50. Available online: https://elibrary.ru/download/elibrary_20453720_27411630.pdf (accessed on 18 January 2022). (In Russian).
- Selikhovkin, A.V.; Drenkhan, R.; Mandelstam, M.Y.; Musolin, D.L. Invasions of insect pests and fungal pathogens of woody plants into the North-Western part of the European Russia. Vestn. St. Petersburg Univ. Earth Sci. 2020, 65, 263–283. (In Russian) [Google Scholar] [CrossRef]
- Selikhovkin, A.; Merkuriev, S.; Khodachek, A. Native and alien tree insect pests: Climate change impact and economic losses in Northwestern Russia. In Proceedings of the 1st International Electronic Conference on Entomology, Basel, Switzerland, 1–15 July 2021; Available online: https://sciforum.net/paper/view/10412 (accessed on 4 January 2021).
- Buy, D.D.; Baryshnikova, S.V.; Denisova, N.V.; Mamaev, N.A.; Musolin, D.L.; Selikhovkin, A.V. Changes in the complex of dendrobiotic phyllophagous insects in Saint Petersburg. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. In The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 98–99. (In Russian) [Google Scholar]
- Buy, D.D.; Baryshnikova, S.V.; Denisova, N.V.; Shevchenko, S.V.; Selikhovkin, A.V. Actual changes in the species composition and population density of phyllophagous insects in St. Petersburg. Izv. St.-Peterbg. Lesoteh. Akad. 2020, 230, 73–99. (In Russian) [Google Scholar] [CrossRef]
- Chursina, V.A.; Vokhtantseva, K.V.; Gayvas, A.A. The main pest of English oak on the territory of the city of Omsk is the leaf blotch miner moth. In Innovative Technologies in Agriculture, Proceedings of the II International Scientific Conference. St. Petersburg, Russia, 20–23 July 2012; Akhmetov, I.G., Akhmetova, M.N., Ivanova, Y.V., Kalenskiy, A.V., Kutashov, V.A., Laktionova, K.S., Saraeva, N.M., Abdrasilov, T.K., Avdejuk, O.A., Aidarov, O.T., et al., Eds.; Svoye Izdatel’stvo Publisher: St. Petersburg, Russia, 2016; pp. 21–25. (In Russian) [Google Scholar]
- Baryshnikova, S.V.; Zoological Institute RAS, Saint Petersburg, Russia. Personal communication, 2021.
- Galich, D.E.; Siberian Forest Experiment Station, Tyumen, Russia. Personal communication, 2021.
- Heckford, R.J. Previously unrecorded foodplants of three species of Microlepidoptera. Entomol. Rec. J. Var. 1993, 105, 93. [Google Scholar]
- Nikitenko, H.M.; Fursov, V.M.; Hershenzon, Z.S.; Svyrydov, S.V. Oak broad-leaved moths and other mining insects on the oak. 2. Morphobiological and ecological characteristics of oak widespread moth and other mining pests of oak. Vestn. Zool. 2004, 38, 53–61. (In Ukrainian) [Google Scholar]
- Apostolov, L.G. Harmful Entomofauna of Forest Biogeocenoses in the Vicinity of the Central Dnieper; Vishcha Shkola Publisher: Kiyev, Ukraine, 1981; pp. 1–232. (In Russian) [Google Scholar]
- Utkina, I.A.; Rubtsov, V.V. The leaf blotch miner moth Acrocercops brongniardella F. (Lepidoptera: Gracillariidae) in Tellerman oakwood. In The Kataev Memorial Readings—X, Proceedings of the International Conference, St. Petersburg, Russia, 22–25 October 2018; Musolin, D.L., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2018; Volume 1. Insects and Other Invertebrates, pp. 104–105. Available online: https://elibrary.ru/download/elibrary_36373591_75292794.pdf (accessed on 18 January 2022). (In Russian)
- Utkina, I.A.; Rubtsov, V.V. The leaf blotch miner moth—Long known, but still little studied species. Izv. St.-Peterbg. Lesoteh. Akad. 2019, 228, 42–56. (In Russian) [Google Scholar]
- Golub, V.B.; Soboleva, V.A.; Aksenenko, E.V. Damage of oak by the leaf blotch miner moth Acrocercops brongniardella F. (Lepidoptera: Gracillariidae) in the Usman’ Forest (Voronezh Province, Russia) 10 years after the fire. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 136–137. (In Russian) [Google Scholar]
- Utkina, I.A.; Rubtsov, V.V. Relationship of different species of oak and phyllophages as an object of biogeocenotic research. Lesovedenie 2021, 5, 547–554. (In Russian) [Google Scholar] [CrossRef]
- Golub, V.B.; Berezhnova, O.N.; Kornev, I.I. Outbreak of the leaf blotch miner moth (Acrocercops brongniardella F., Lepidoptera, Gracillariidae) in the Voronezh Region. Izv. St.-Peterbg. Lesoteh. Akad. 2009, 187, 96–102. Available online: https://elibrary.ru/download/elibrary_13020131_77590310.pdf (accessed on 18 January 2022). (In Russian).
- Eaton, E.; Caudullo, G.; Oliveira, S.; de Rigo, D. Quercus robur and Quercus petraea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publisher Off. EU: Luxembourg, Belgium, 2016; pp. 160–163. [Google Scholar]
- Ermolaev, I.V.; Vasil’ev, A.A. Phytophagous insects associated with the pedunculate oak (Quercus robur L.) in the Siva River valley. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 155–156. (In Russian) [Google Scholar]
- Ermolaev, I.V.; Ponomarev, V.I.; Vasil’ev, A.A.; Kumaeva, M.S. Phytophagous insects of the pedunculate oak (Quercus robur) in the northeast of its distribution area. Zool. J. 2021, 100, 640–651. (In Russian) [Google Scholar] [CrossRef]
- Marion, D.F.; Larew, H.G.; Knodel, J.J.; Natoli., W. Systemic activity of neem extract against birch leafminer. J. Arboric. 1990, 16, 12–16. [Google Scholar]
- Golosova, M.A.; Gninenko, Y.I.; Golosova, E.I. Ohrid Leaf-Miner Cameraria ohridella is a Dangerous Quarantine Pest on Objects of Urban Ggreening; VNIILM: Moscow, Russia, 2008; pp. 1–26. (In Russian) [Google Scholar]
- Gous, S.; Richardson, B. Stem injection of insecticides to control herbivorous insects on Eucalyptus niten. New Zealand Plant Prot. 2008, 61, 174–178. [Google Scholar] [CrossRef]
- Šefrová, H. Invasions of Lithocolletinae species in Europe—Causes, kinds, limits and ecological impact (Lepidoptera, Gracillariidae). Ekológia 2003, 22, 132–142. Available online: https://www.researchgate.net/publication/297685678_Invasions_of_Lithocolletinae_species_in_Europe_-_Causes_kinds_limits_and_ecological_impact_Lepidoptera_Gracillariidae (accessed on 18 January 2022).
- Kirichenko, N.I.; Triberti, P.; Akulov, E.N.; Ponomarenko, M.G.; Lopez-Vaamonde, C. Novel data on the taxonomic diversity, distribution, and host plants of leafmining moths of the family Gracillariidae (Lepidoptera) in Siberia, based on DNA barcoding. Entomol. Rev. 2019, 99, 796–819. [Google Scholar] [CrossRef]
- Hellrigl, K. Neue Erkenntnisse und Untersuchungen über die Roßkastanien-Miniermotte Cameraria ohridella Deschka & Dimić, 1986 (Lepidoptera, Gracillariidae). Gredleriana 2001, 1, 9–81. [Google Scholar]
- Grabenweger, G.; Avtzis, N.; Girardoz, S.; Hrasovec, B.; Tomov, R.; Kenis, M. Parasitism of Cameraria ohridella (Lepidoptera, Gracillariidae) in natural and artificial horse-chestnut stands in the Balkans. Agric. For. Entomol. 2005, 7, 291–296. [Google Scholar] [CrossRef]
- Valade, R.; Kenis, M.; Hernandez-Lopez, A.; Augustin, S.; Mari Mena, N.; Magnoux, E.; Rougerie, R.; Lakatos, F.; Roques, A.; Lopez-Vaamonde, C. Mitochondrial and microsatellite DNA markers reveal a Balkan origin for the highly invasive horse-chestnut leaf miner Cameraria ohridella (Lepidoptera, Gracillariidae). Mol. Ecol. 2009, 18, 3458–3470. [Google Scholar] [CrossRef] [PubMed]
- Lees, D.C.; Lack, H.W.; Rougerie, R.; Hernandez-Lopez, A.; Raus, T.; Avtzis, N.; Augustin, S.; Lopez-Vaamonde, C. Tracking origins of invasive herbivores using herbaria and archival DNA: The case of the horse-chestnut leafminer. Front. Ecol. Environ. 2011, 9, 322–328. [Google Scholar] [CrossRef]
- Gninenko, Y.I.; Shepelev, S.V. New phytophages and diseases of tree species. Lesn. Khozyaistvo 2004, 3, 48. (In Russian) [Google Scholar]
- Gninenko, Y.I.; Orlinski, A.D. New insect pests of forest plantations. Plant Prot. Quar. 2004, 4, 33. (In Russian) [Google Scholar]
- Golosova, M.A.; Gninenko, Y.I. The appearance of the horse-chestnut leafminer on horse chestnut in Moscow. Lesn. Vestn. 2006, 2, 43–46. Available online: https://elibrary.ru/download/elibrary_9273118_72290780.pdf (accessed on 18 January 2022). (In Russian).
- Akimov, I.A.; Zerova, M.D.; Gershenson, Z.S.; Narolsky, N.B.; Kochanez, O.M.; Sviridov, S.V. First record of the horse-chestnut leafminer Cameraria ohridella (Lepidoptera, Gracillariidae) on Aesculus hippocastanum (Hippocastanaceae) in Ukraine. Vestn. Zool. 2003, 37, 3–12. (In Russian) [Google Scholar]
- Rakov, A.G. The Horse-chestnut Leafminer and other Invasive Phyllophagous Species under the Conditions of Formation of their Ranges in the European Part of Russia. Ph.D. Thesis, Russian State Agricultural University, Moscow, Russia, 2015. (In Russian). [Google Scholar]
- Zhuravleva, E.N. The first appearance of the Ohrid leaf miner Cameraria ohridella (Lepidoptera: Gracillariidae) on the horse chestnut in the Greater Sochi. In The Kataev Memorial Readings—VIII, Proceedings of the International Conference, St. Petersburg, Russia, 18–20 November 2014; Musolin, D.L., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2014; p. 32. (In Russian) [Google Scholar]
- Karpun, N.N. Features of formation of dendrophagous invasive pest fauna in the humid subtropics of Russia at the beginning of the XXI century. Izv. St.-Peterbg. Lesoteh. Akad. 2019, 228, 104–119. (In Russian) [Google Scholar] [CrossRef]
- Chenikalova, E.V. Balkan leafminer moth—A threat to the horse chestnut in the Stavropol Region. Plant Prot. Quar. 2018, 11, 38–39. Available online: https://elibrary.ru/download/elibrary_36498157_20089242.pdf (accessed on 18 January 2022). (In Russian).
- Chenikalova, E.V. The chestnut moth Cameraria ohridella (Deschka et Dimić, 1986) and boxwood moth Cydalima perspectalis (Walker, 1859) in the tree stands of Stavropol and its environs. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 347–348. (In Russian) [Google Scholar]
- Martynov, V.V.; Nikulina, T.V.; Shokhin, I.V.; Terskov, E.N. Contributions to the fauna of invasive insects of Ciscaucasia. Field Biol. J. 2020, 2, 99–122. (In Russian) [Google Scholar] [CrossRef]
- Selikhovkin, A.V.; Popovichev, B.G.; Denisova, N.V.; Timofeeva, Y.A. Invasive mining moths (Lepidoptera: Gracillariidae) of woody plantations in St. Petersburg. In XV Congress of the Russian Entomological Society, Proceedings of the Congress, Novosibirsk, Russia, 31 July—7 August 2017; Azarkina, G.N., Baranchikov, Y.N., Barkalov, A.V., Belokobilskiy, S.A., Glupov, V.V., Grichanov, I.Y., Danilov, Y.N., Dubatolov, V.V., Dudko, R.Y., Kireychuk, A.G., et al., Eds.; Garamond Publisher: Novosibirsk, Russia, 2017; pp. 446–447. (In Russian) [Google Scholar]
- Anikin, V.V. Present day bio-invasions in the Volga-Ural Region: From the South to the North or from the East to the West? Cameraria ohridella (Lepidoptera: Gracillariidae) in the Lower and Middle Volga. Zootaxa 2019, 4624, 583–588. [Google Scholar] [CrossRef]
- Anikin, V.V.; Zolotuhin, V.V.; Polumordvinov, O.A. Mass damage of horse chestnut’s leaves (Aesculus hippocastanum) by Ohrid leafminer (Cameraria ohridella) on the territory of Penza in 2019. Bull. Botanic Garden Saratov State Univ. 2019, 17, 235–241. (In Russian) [Google Scholar] [CrossRef]
- Kryukova, A.V.; Nikolaeva, Z.V. Harmfulness of chestnut mining moth in Pskov region. Bull. Velikoluk. State Agric. Acad. 2018, 1, 1–7. Available online: https://elibrary.ru/download/elibrary_35034180_72824990.pdf (accessed on 18 January 2022). (In Russian).
- Grigoriev, M.A.; Grigoriev, A.I. Ecological and biological characteristic of horse chestnut (Aesculus hippocastanum L.) in the conditions of the south of Western Siberia. Bull. Omsk State Agric. Univ. 2017, 3, 61–68. Available online: https://elibrary.ru/download/elibrary_30467968_44356667.pdf (accessed on 18 January 2022). (In Russian).
- Triebel, S.A.; Gamanova, O.N. Monitoring chestnut miner moth. Plant Prot. Quar. 2009, 2, 45–47. [Google Scholar]
- Lack, H.W. The discovery and rediscovery of the horse chestnut. Arnoldia 2002, 61, 15–19. [Google Scholar] [CrossRef]
- Kenis, M.; Tomov, R.; Svatos, A.; Schlinsog, P.; Lopez-Vaamonde, C.; Heitland, W.; Grabenweger, G.; Girardoz, S.; Freise, J.; Avtzis, N. The horse-chestnut leaf miner in Europe—Prospects and constraints for biological control. In Proceedings of the Second International Symposium on Biological Control of Arthropods, Davos, Switzerland, 12–16 September 2005; Hoddle, M., Ed.; Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2005; pp. 77–90. [Google Scholar]
- D’Costa, L.; Koricheva, J.; Straw, N.; Simmonds, M.S.J. Oviposition patterns and larval damage by the invasive horse-chestnut leaf miner Cameraria ohridella on different species of Aesculus. Ecol. Entomol. 2013, 38, 456–462. [Google Scholar] [CrossRef]
- Davydova, I.A.; Gradusov, V.M.; Rybinceva, A.L. Cameraria ohridella Deschka et Dimić (Lepidoptera: Gracillariidae) on different species of horse chestnut in Moscow landscaping. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November, 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 141–142. (In Russian) [Google Scholar]
- Pschorn-Walcher, H. Freiland-Biologie der eingeschleppten Roßkastanien-Miniermotte Cameraria ohridella Deschka and Dimić (Lep., Gracillariidae) im Wienerwald. Linz. Biol. Beiträge 1994, 26, 633–642. [Google Scholar]
- Gilbert, M.; Grégoire, J.-C.; Freise, J.F.; Heitland, W. Long-distance dispersal and human population density allow the prediction of invasive patterns in the horse chestnut leafminer Cameraria ohridella. J. Anim. Ecol. 2004, 73, 459–468. [Google Scholar] [CrossRef]
- Augustin, S.; Reynaud, P. Un nouveau ravageur pour le marronnier: Cameraria ohridella. PHM Rev. Hortic. 2000, 418, 44–45. [Google Scholar]
- Kirichenko, N.; Augustin, S.; Kenis, M. Invasive leafminers on woody plants: A global review of pathways, impact and management. J. Pest Sci. 2019, 92, 93–106. [Google Scholar] [CrossRef] [Green Version]
- The Most Dangerous Invasive Species in Russia (TOP-100); Dgebuadze, Y.Y.; Petrosyan, V.G.; Khlyap, L.A. (Eds.) KMK Scientific Press Ltd.: Moscow, Russia, 2018; pp. 1–688. (In Russian) [Google Scholar]
- Cherpakov, V.V. About bacterial damage caused by Pseudomonas syringae pv. aesculi on horse chestnut in the Russian Federation. Actual Probl. For. Complex 2013, 36, 72–80. Available online: https://elibrary.ru/item.asp?id=21267711 (accessed on 20 January 2022). (In Russian).
- Straw, N.A.; Williams, D.T. Impact of the leaf miner Cameraria ohridella (Lepidoptera: Gracillariidae) and bleeding canker disease on horse-chestnut: Direct effects and interaction. Agric. For. Entomol. 2013, 15, 321–333. [Google Scholar] [CrossRef]
- Koskella, B.; Meaden, S.; Crowther, W.J.; Leimu, R.; Metcalf, C.J.E. A signature of tree health? Shifts in the microbiome and the ecological drivers of horse chestnut bleeding canker disease. New Phytol. 2017, 215, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Freise, J.; Heitland, W.; Tosevski, I. Parasitism of the horse chestnut leafminer, Cameraria ohridella Deschka and Dimić (Lep., Gracillariidae), in Serbia and Macedonia. Anz. Schädlingskunde 2002, 75, 152–157. [Google Scholar] [CrossRef]
- Bhatti, I.; Shaw, O.; Shaw, P. Parasitoids and parasitism rates of the horse chestnut leaf miner Cameraria ohridella Deschka and Dimić [Lepidoptera: Gracillariidae] across four sites in south-west London. Arboricultural J. 2013, 35, 147–159. [Google Scholar] [CrossRef]
- Volter, L.; Kenis, M. Parasitoid complex and parasitism rates of the horse chestnut leafminer, Cameraria ohridella (Lepidoptera: Gracillariidae) in the Czech Republic, Slovakia and Slovenia. Eur. J. Entomol. 2006, 103, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Girardoz, S.; Volter, L.; Tomov, R.; Quicke, D.L.J.; Kenis, M. Variations in parasitism in sympatric populations of three invasive leaf miners. J. Appl. Entomol. 2007, 131, 603–612. [Google Scholar] [CrossRef]
- Grabenweger, G.; Kehrli, P.; Zweimuller, I.; Augustin, S.; Avtzis, N.; Bacher, S.; Freise, J.; Girardoz, S.; Guichard, S.; Heitland, W.; et al. Temporal and spatial variations in the parasitoid complex of the horse chestnut leafminer during its invasion of Europe. Biol. Invasions 2010, 12, 2797–2813. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Lopez, A.; Rougerie, R.; Augustin, S.; Lees, D.C.; Tomov, R.; Kenis, M.; Cota, E.; Kullaj, E.; Hansson, C.; Grabenweger, G.; et al. Host tracking or cryptic adaptation? Phylogeography of Pediobius saulius (Hymenoptera, Eulophidae), a parasitoid of the highly invasive horse-chestnut leafminer. Evol. Appl. 2012, 5, 256–269. [Google Scholar] [CrossRef]
- Kostyukov, V.V.; Kosheleva, O.V.; Nakonechnaya, I.V.; Gunasheva, Z.M. First report on parasitoids of the horse chestnut moth in Russia. Plant Prot. Quar. 2014, 9, 41–42. Available online: https://elibrary.ru/download/elibrary_21841455_76822640.pdf (accessed on 18 January 2022). (In Russian).
- Měsić, A.; Barčić, J.; Barčić, J.I.; Miličević, T.; Duralija, B.; Čuljak, T.G. A low environmental impact method to control horse chestnut leaf miner Cameraria ohridella (Deschka & Dimic). J. Food Agric. Environ. 2008, 6, 421–427. Available online: https://www.researchgate.net/publication/231167581_A_low_environmental_impact_method_to_control_horse_chestnut_leaf_miner_Cameraria_ohridella_Deschka_Dimic (accessed on 18 January 2022).
- De Prins, J.; De Prins, W. The occurrence of Cameraria ohridella in Belgium (Lepidoptera: Gracillariidae). Phegea 2001, 29, 81–88. Available online: https://www.researchgate.net/publication/250928533_The_occurrence_of_Cameraria_ohridella_in_Belgium_Lepidoptera_Gracillariidae (accessed on 18 January 2022).
- Jagiełło, R.; Walczak, U.; Iszkuło, G.; Karolewski, P.; Baraniak, E.; Giertych, M.J. Impact of Cameraria ohridella on Aesculus hippocastanum growth and long-term effects of trunk injection with pesticides. Int. J. Pest Manag. 2019, 65, 33–43. [Google Scholar] [CrossRef]
- Larina, G.E.; Seraya, L.G.; Kashtanova, O.A.; Dymovich, A.M.; Baranchikov, Y.N. The effectiveness of endotherapy of trunks in protection of horse-chestnut Aesculus hippocastanum L. against mining insects. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 187–188. (In Russian) [Google Scholar]
- Svatos, A.; Kalinová, B.; Hoskovec, M.; Kindl, J.; Hrdy, I. Chemical communication in horse-chestnut leafminer Cameraria ohridella Deschka & Dimic. Plant. Prot. Sci. 1999, 35, 10–13. [Google Scholar]
- Atanov, N.M.; Kuzina, N.P.; Kuzin, A.A.; Chirskaya, M.V. Impact of modifiers on the attractiveness of the horse-chestnut leaf-miner’s pheromone Cameraria ohridella Deschka et Dimič, 1986. Plant Health Res. Pract. 2016, 4, 47–49. Available online: http://www.rsn-msk.ru/files/karantin-18-2016.pdf (accessed on 18 January 2022).
- Kehrli, P.; Lehmann, M.; Bacher, S. Mass-emergence devices: A biocontrol technique for conservation and augmentation of parasitoids. Biol. Control 2005, 32, 191–199. [Google Scholar] [CrossRef]
- Kumata, T. Taxonomic studies on the Lithocolletinae of Japan. Insecta Matsumurana 1963, 25, 53–90. [Google Scholar]
- Kirichenko, N.; Ryazanova, M.; Akulov, E.; Ponomarenko, M. Can native parasitoids control the invasive lime leaf-miner Phyllonorycter issikii (Lepidoptera: Gracillariidae) in Western Siberia? In Proceedings of the 1st International Electronic Conference on Entomology, Basel, Switzerland, 1–15 July 2021; Available online: https://sciforum.net/paper/view/10537 (accessed on 4 January 2021).
- Kumata, T.; Kuroko, H.; Park, K. Some Korean species of the subfamily Lithocolletinae Gracillariidae, Lepidoptera. Korean J. Plant Prot. 1983, 22, 213–227. [Google Scholar]
- Kirichenko, N.I.; Akulov, E.N.; Babichev, N.S.; Mikhailova, I.A.; Ponomarenko, M.G.; Lopez-Vaamonde, C. Past distribution of Tilia-feeding Phyllonorycter micromoth (Lepidoptera: Gracillariidae) in the Russian Far East based on survey of historical herbarium. Far Eastern Entomol. 2019, 390, 19–32. [Google Scholar] [CrossRef]
- Kirichenko, N.I. Trophic Associations and Invasion Patterns of the Dendrophilous Gracillariid Moths (Lepidoptera: Gracillariidae) in the Asian Part of Russia. Ph.D. Thesis, Siberian Federal University, Krasnoyarsk, Russia, 2020. Available online: https://research.sfu-kras.ru/sites/research.sfu-kras.ru/files/Dissertaciya_Kirichenko.pdf (accessed on 18 January 2022). (In Russian).
- Bednova, O.V.; Belov, D.A. Lime leaf miner (Lepidoptera, Gracillariidae) in green spaces of Moscow and Moscow Region. Lesn. Vestn. 1999, 2, 172–177. (In Russian) [Google Scholar]
- Kozlov, M.V. Leaf-mining moth is a pest of limes. Plant Prot. Quar. 1991, 4, 46. (In Russian) [Google Scholar]
- Šefrová, H. Phyllonorycter issikii (Kumata, 1963)—Bionomics, ecological impact and spread in Europe (Lepidoptera, Gracillariidae). Acta Univ. Agric. Silvic. Mendel. Brun. 2002, 50, 99–104. [Google Scholar]
- Ermolaev, I.V.; Rubleva, E.A. History, rate and factors of invasion of lime leafminer Phyllonorycter issikii (Kumata, 1963) (Lepidoptera, Gracillariidae) in Eurasia. Russ. J. Biol. Invasions 2017, 1, 2–19. (In Russian) [Google Scholar] [CrossRef]
- Gninenko, Y.I.; Kozlova, E.I. Lime leaf miner in Russia and problems of biological control. In Proceedings of the International Organization for Biological Control of Harmful Animals and Plants, East Palaearctic Regional Section, Abstracts, Poznan, Poland, 15–19 May 2006; IOBC: Poznan, Poland, 2006; p. 16. [Google Scholar]
- Kirichenko, N.; Triberti, P.; Ohshima, I.; Haran, J.; Byun, B.-K.; Li, H.; Augustin, S.; Roques, A.; Lopez-Vaamonde, C. From east to west across the Palearctic: Phylogeography of the invasive lime leaf miner Phyllonorycter issikii (Lepidoptera: Gracillariidae) and discovery of a putative new cryptic species in East Asia. PLoS ONE 2017, 12, e0171104. [Google Scholar] [CrossRef]
- Ermolaev, I.V.; Rubleva, E.A.; Rysin, S.L.; Ermolaeva, M.V. Forage plants of lime leafminer Phyllonorycter issikii (Kumata, 1963) (Lepidoptera, Gracillariidae). Russ. J. Biol. Invasions 2018, 2, 2–13. Available online: https://elibrary.ru/download/elibrary_35166252_58174722.pdf (accessed on 18 January 2022). (In Russian).
- Kirichenko, N. The lime leafminer Phyllonorycter issikii in Western Siberia: Some ecological characteristics of the population of the recent invader. Contemp. Probl. Ecol. 2014, 7, 114–121. [Google Scholar] [CrossRef]
- Pigott, C.D. Lime-trees and Basswoods. A Biological Monograph of the Genus Tilia; Cambridge University Press: New York, NY, USA, 2012; pp. 1–395. [Google Scholar]
- Zolotukhin, V.V. About some invasive arthropods on the territory of the Ulyanovsk region. Nat. Simbirsk Volga Reg. 2002, 2, 200–203. [Google Scholar]
- Ermolaev, I.V.; Zorin, D.A. Ecological consequences of invasion of Phyllonorycter issikii (Lepidoptera, Gracillariidae) in lime forests in Udmurtia. Entomol. Rev. 2011, 91, 592–598. [Google Scholar] [CrossRef]
- Kirichenko, N.I.; Zakharov, E.V.; Lopez-Vaamonde, C. Revealing the invasion history of the lime leafminer Phyllonorycter issikii (Lepidoptera: Gracillariidae) using historical herbaria and next generation sequencing. In Dendrobiotic Invertebrates and Fungi and their Role in Forest Ecosystems. The Kataev Memorial Readings—XI, Proceedings of the All-Russia Conference with International Participation, St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; SPbFTU: St. Petersburg, Russia, 2020; pp. 172–173, 175. (In Russian) [Google Scholar]
- Ermolaev, I.V.; Yefremova, Z.A.; Izhboldina, N.V. Parasitoids as a mortality factor for the lime leafminer (Phyllonorycter issikii, Lepidoptera, Gracillariidae). Entomol. Rev. 2011, 91, 326–334. [Google Scholar] [CrossRef]
- Szőcs, L.; George, M.; Thuróczy, C.; Csóka, G. Parasitoids of the lime leaf miner Phyllonorycter issikii (Lepidoptera: Gracillariidae) recorded throughout the area it recently colonized. Eur. J. Entomol. 2015, 112, 591–598. [Google Scholar] [CrossRef] [Green Version]
- Ermolaev, I.V.; Yefremova, Z.A.; Gerasimova, N.A.; Koryoleva, E.A.; Lushnikov, N.N.; Petrov, A.I.; Pchel’nikov, A.A. Parasitoids (Hymenoptera) of the lime leafminer (Phyllonorycter issikii, Lepidoptera, Gracillariidae) in different cities of the Russian Federation and their role in the mortality of the invasive pest. Entomol. Rev. 2019, 99, 485–493. [Google Scholar] [CrossRef]
- Yefremova, Z.A.; Mishchenko, A.V. The parasitoid complex (Hymenoptera, Eulophidae) of the leafminer Phyllonorycter issikii (Kumata) (Lepidoptera, Gracillariidae) from the Middle Volga Basin. Entomol. Rev. 2008, 88, 178–185. [Google Scholar] [CrossRef]
- Mishchenko, A.V. Method of mass rearing ichneumons Minotetrastichus frontalis (Nees). Bull. Russ. Fed. Fed. Serv. Intellect. Property 2015, 10, 1–8. [Google Scholar]
- Baryshnikova, S.V.; Bolshakov, L.V. Microlepidoptera of Tula Province. 15. Families Bucculatricidae, Gracillariidae, and Lyonetiidae (Hexapoda: Lepidoptera). In Biodiversity of Tula Region at the Turn of Centuries; Bolshakov, L.V., Ed.; Grif & K: Tula, Russia, 2004; Volume 4, pp. 31–37. (In Russian) [Google Scholar]
- Ermolaev, I.V.; Rubleva, E.A.; Rysin, S.L.; Kozhenkova, A.A.; Ermolaeva, M.V. Trophic specialization of the poplar leafminer Phyllonorycter populifoliella (Treitschke, 1833) (Lepidoptera, Gracillariidae). Entomol. Rev. 2020, 100, 287–300. [Google Scholar] [CrossRef]
- Shashank, P.R.; Singh, N.; Harshana, A.; Sinha, T.; Kirichenko, N. First report of the poplar leaf miner, Phyllonorycter populifoliella (Treitschke) (Lepidoptera: Gracillariidae) from India. Zootaxa 2021, 4915, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Kozhanchikov, I.V.; Danilevskiy, A.S.; Djakonov, A.M. Order Lepidoptera. In Pests of Forest (Reference Book); Shtakelberg, A.A., Ed.; Publishing House of the Academy of Sciences of the USSR: Moscow, Russia; St. Petersburg, Russia, 1955; Volume 1, pp. 35–287. (In Russian) [Google Scholar]
- Tarasova, O.V.; Kovalev, A.V.; Sukhovolsky, V.G.; Khlebopros, R.G. Phylophagous Insects of Green Plantations of Cities: Species Compositions and Population Dynamics; Nauka: Novosibirsk, Russia, 2004; pp. 1–180. (In Russian) [Google Scholar]
- Ermolaev, I.V. Ecological mechanisms of nonperiodical population wave: A case study of the poplar leafminer—Phyllonorycter populifoliella (Lepidoptera, Gracillariidae). J. General Biol. 2019, 80, 451–476. [Google Scholar]
- Polezhaev, V.G. Struggle for existence in the poplar leafminer Lithocolletis populifoliella Tr. Zool. Zhurnal. 1934, 13, 67–75. (In Russian) [Google Scholar]
- Frolov, D.N. The poplar leafminer as a landscaping pest in Irkutsk. Proceedings Irkutsk State Univ. Ser. Biol. 1948, 3, 19. (In Russian) [Google Scholar]
- Selikhovkin, A.V. Population dynamics of Microlepidoptera under the conditions of industrial air pollution: A follow-up. Izv. St.-Peterbg. Lesoteh. Akad. 1996, 162, 26–38. (In Russian) [Google Scholar]
- Selikhovkin, A.V.; Baryshnikova, S.V.; Denisova, N.V.; Timofeeva, Y.A. Species composition and population dynamics of dominant dendrophagous moths (Lepidoptera) in St. Petersburg and its environs. Entomol. Rev. 2018, 98, 963–978. [Google Scholar] [CrossRef]
- Selikhovkin, A.V.; Egorov, A.A.; Sitnikova, D.D.; Mamaev, N.A. Occurrence of leaf miners Phyllonorycter populifoliella (Treitschke) and Ph. pastorella (Zeller) (Lepidoptera, Gracillariidae) on different species of poplars. Entomol. Rev. 2020, 100, 301–306. [Google Scholar] [CrossRef]
- Rumyantsev, P.D. Biology of the poplar leaf miner Lithocolletis populifoliella in the conditions of Moscow. Zool. Zhurnal. 1934, 13, 257–279. (In Russian) [Google Scholar]
- Mamaev, N.A.; Buy, D.D.; Selikhovkin, A.V. The second outbreak of the poplar leafminer Phyllonorycter populifoliella in St. Petersburg. Izv. St.-Peterbg. Lesoteh. Akad. 2020, 233, 81–94. (In Russian) [Google Scholar] [CrossRef]
- Lvovsky, A.L. Lepidoptera (Insecta) within the city limits of St. Petersburg. Proceedings Kharkiv Entomol. Soc. 1994, 2, 4–48. (In Russian) [Google Scholar]
- Bondarenko, E.A. An outbreak of the poplar leafminer Phyllonorycter populifoliella Tr. (Lepidoptera, Gracillariidae) in St. Petersburg. Izv. St.-Peterbg. Lesoteh. Akad. 2008, 182, 45–55. (In Russian) [Google Scholar]
- Selikhovkin, A.V. Peculiarities of the population dynamics of the poplar mining moth Phyllonorycter populifoliella Tr. (Gracillariidae). Izv. St.-Peterbg. Lesoteh. Akad. 2010, 192, 220–235. Available online: https://elibrary.ru/download/elibrary_17047133_73905770.pdf (accessed on 18 January 2022). (In Russian).
- Ermolaev, V.P. Ecologo-faunistic review of leafmining moths (Lepidoptera, Gracillariidae) of southern Primorye. Tr. Zool. Inst. Akad. Nauk SSSR 1977, 70, 98–116. (In Russian) [Google Scholar]
- Ðức, B.Đ.; Leont’ev, L.L.; Baryshnikova, S.V.; Selikhovkin, A.V. Consequences of the outbreaks of the poplar leaf miner and other mining Microlepidoptera in St. Petersburg. Contemp. Probl. Ecol. 2021, 14, 822–826. [Google Scholar] [CrossRef]
- Sulkhanov, A.V. Ecology of urban populations of the poplar moth Lithocolletis populifoliella Tr. In Dendrobiont Insects of Trees and Shrubs of Moscow; Krivosheina, N.P., Striganova, B.R., Eds.; Nauka Publisher House: Moscow, Russia, 1992; pp. 70–97. (In Russian) [Google Scholar]
- Chaika, S.Y. Insects. In Great Russian Encyclopedia; Electronic Version; 2017; Available online: https://bigenc.ru/biology/text/2250439 (accessed on 2 November 2021).
- Kerzhner, I.M. An approach and perspective of study of Insecta in the territory of the former USSR. In Biological Diversity: The Level of Taxonomic Study; Sokolov, V.E., Reshetnikov, Y.S., Eds.; Nauka Publisher House: Moscow, Russia, 1994; pp. 65–69. (In Russian) [Google Scholar]
- Cippolini, D. White fringetree as a novel larval host for emerald ash borer. J. Econ. Entomol. 2015, 108, 370–375. [Google Scholar] [CrossRef] [Green Version]
- Orlova-Bienkowskaja, M.J. Cascading ecological effects caused by the establishment of the emerald ash borer Agrilus planipennis (Coleoptera: Buprestidae) in European Russia. Eur. J. Entomol. 2015, 112, 778–789. [Google Scholar] [CrossRef]
- Eschen, R.; O’Hanlon, R.; Santini, A.; Vannini, A.; Roques, A.; Kirichenko, N.; Kenis, M. Safeguarding global plant health: The rise of sentinels. J. Pest Sci. 2018, 62, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Morales-Rodríguez, C.; Anslan, S.; Auger-Rozenberg, M.-A.; Augustin, S.; Baranchikov, Y.; Bellahirech, A.; Burokienė, D.; Čepukoit, D.; Çota, E.; Davydenko, K.; et al. Forewarned is forearmed: Preventive detection of potentially invasive pests and pathogens in sentinel plantings. NeoBiota 2019, 47, 95–123. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]





















{kind=link}
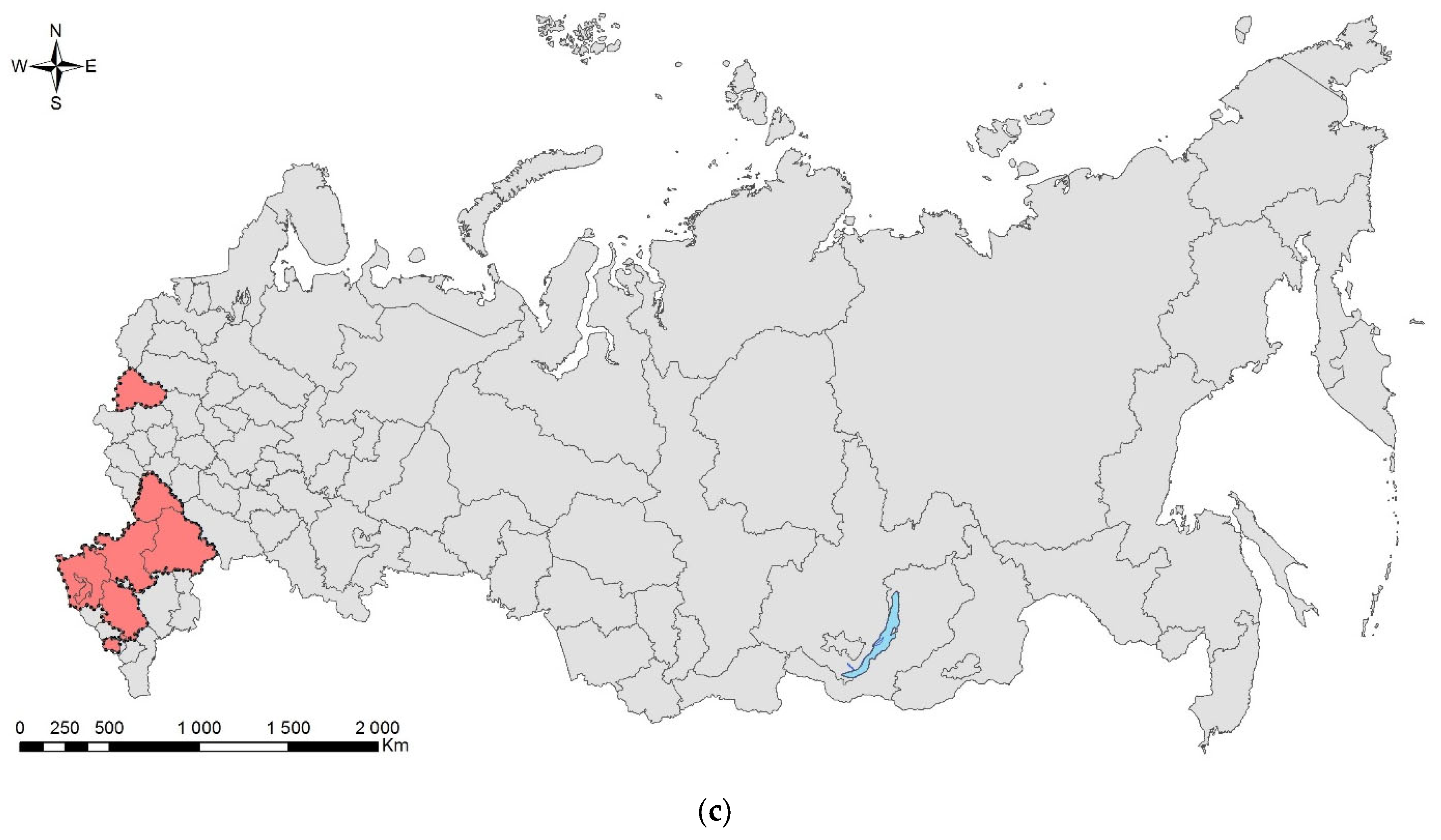{kind=link}
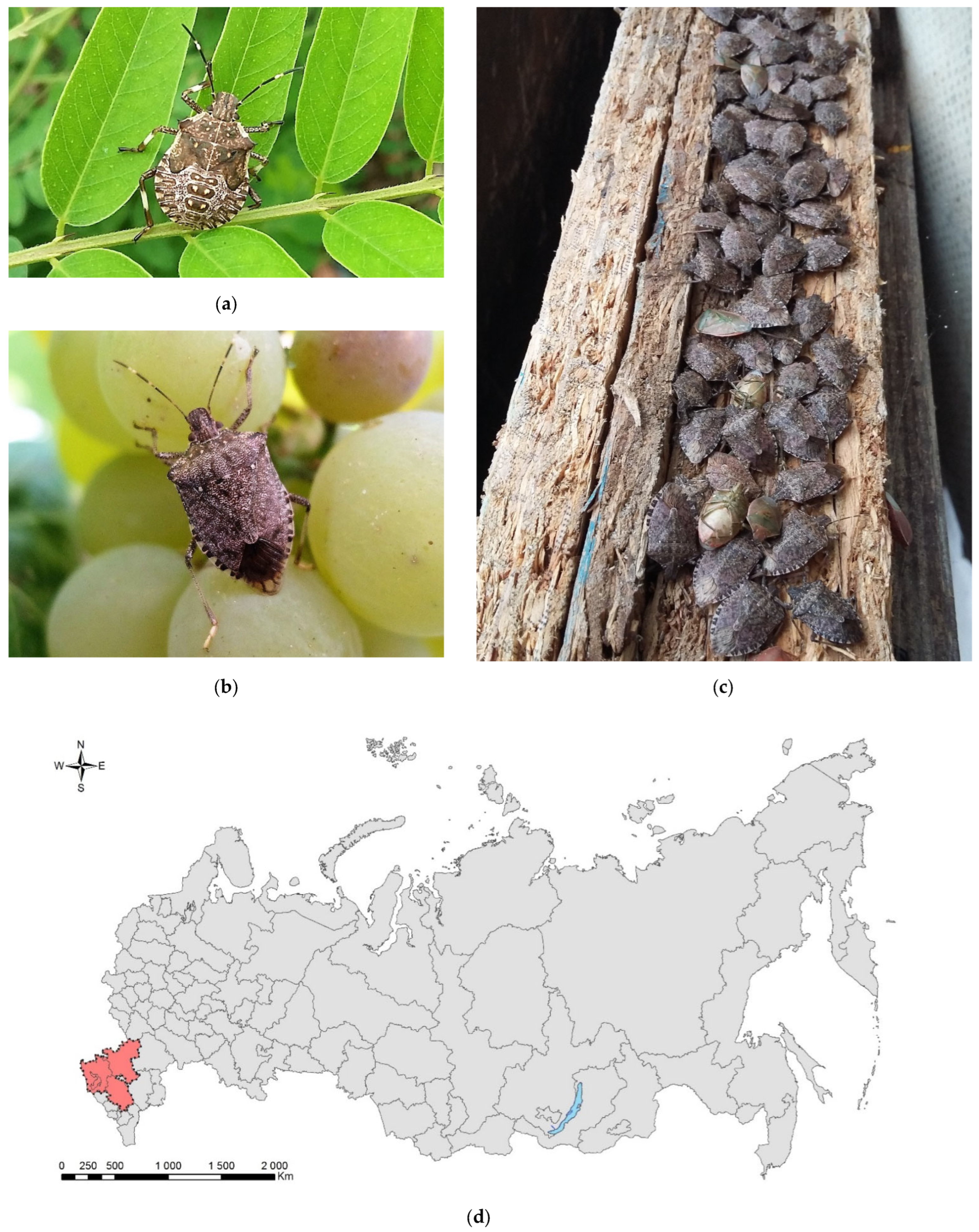{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
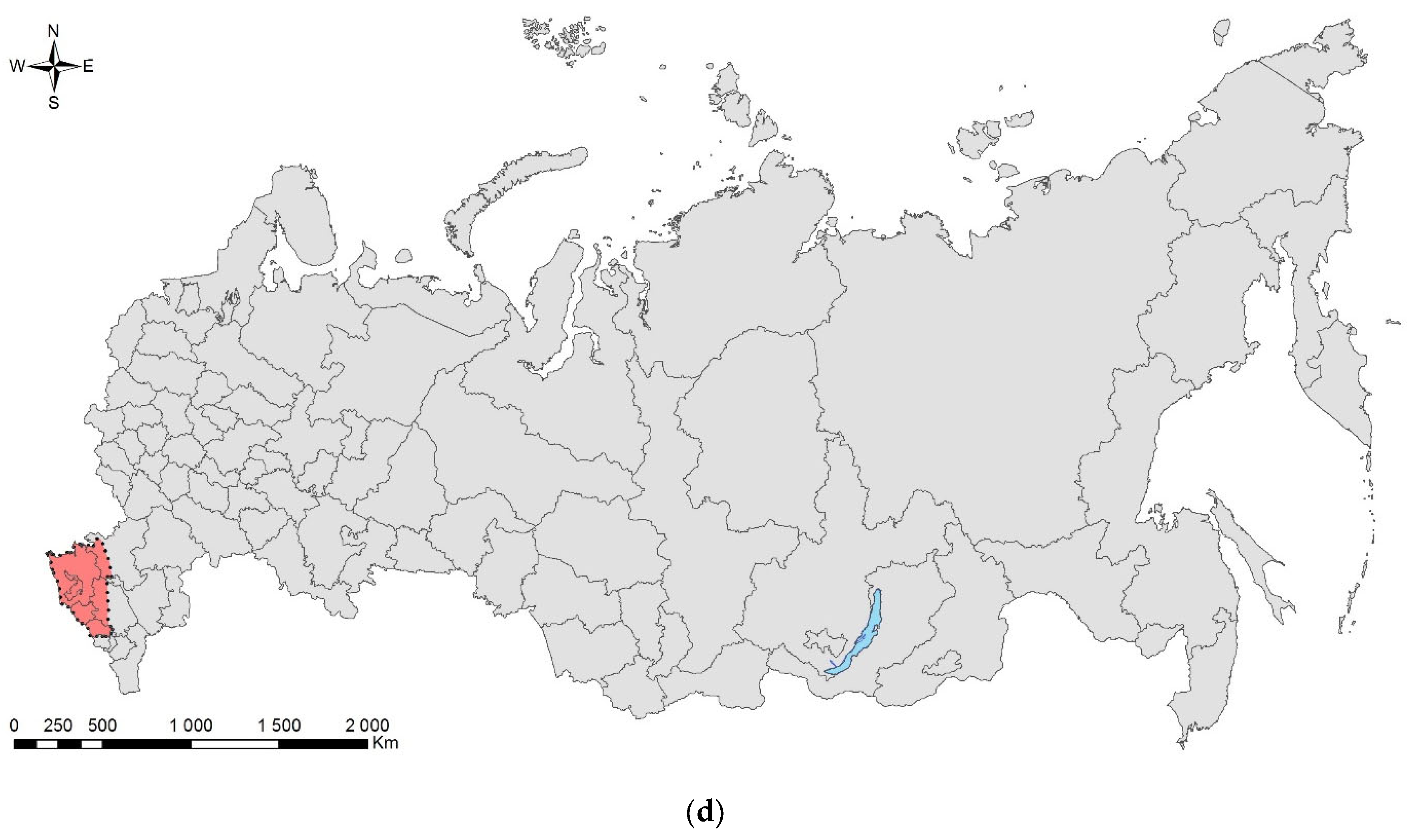{kind=link}
{kind=link}
{kind=link}
{kind=link}
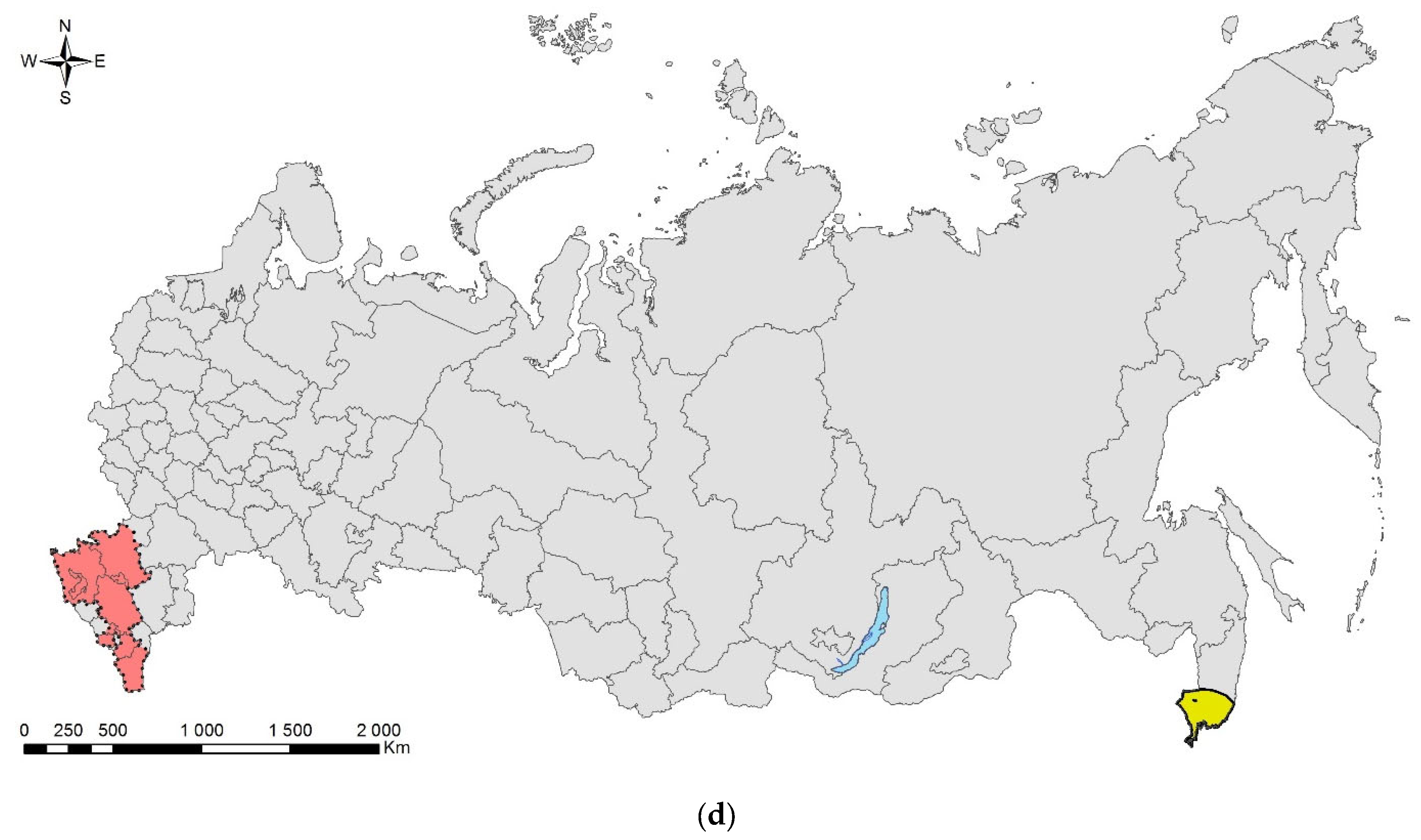{kind=link}
{kind=link}
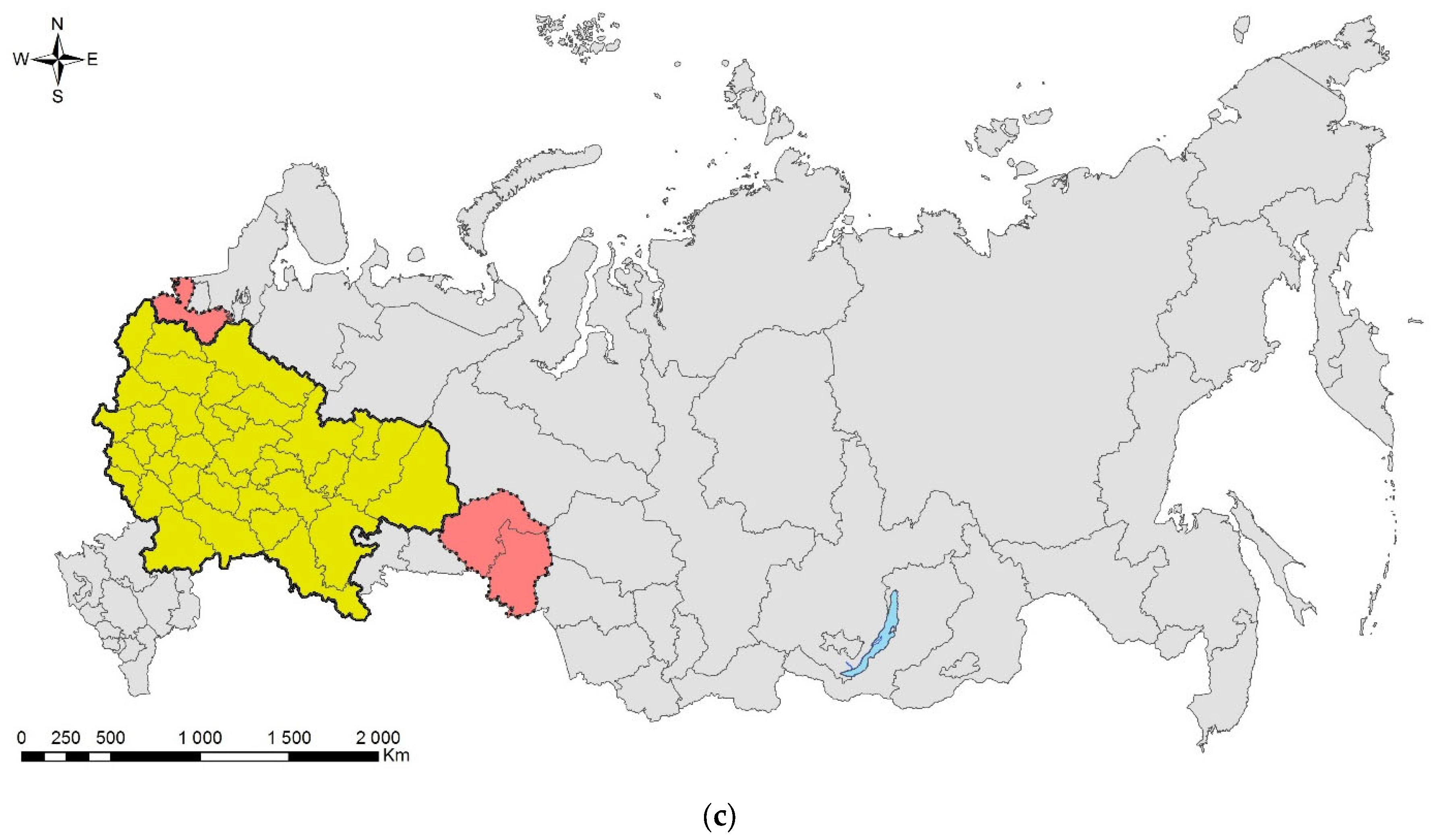{kind=link}
{kind=link}
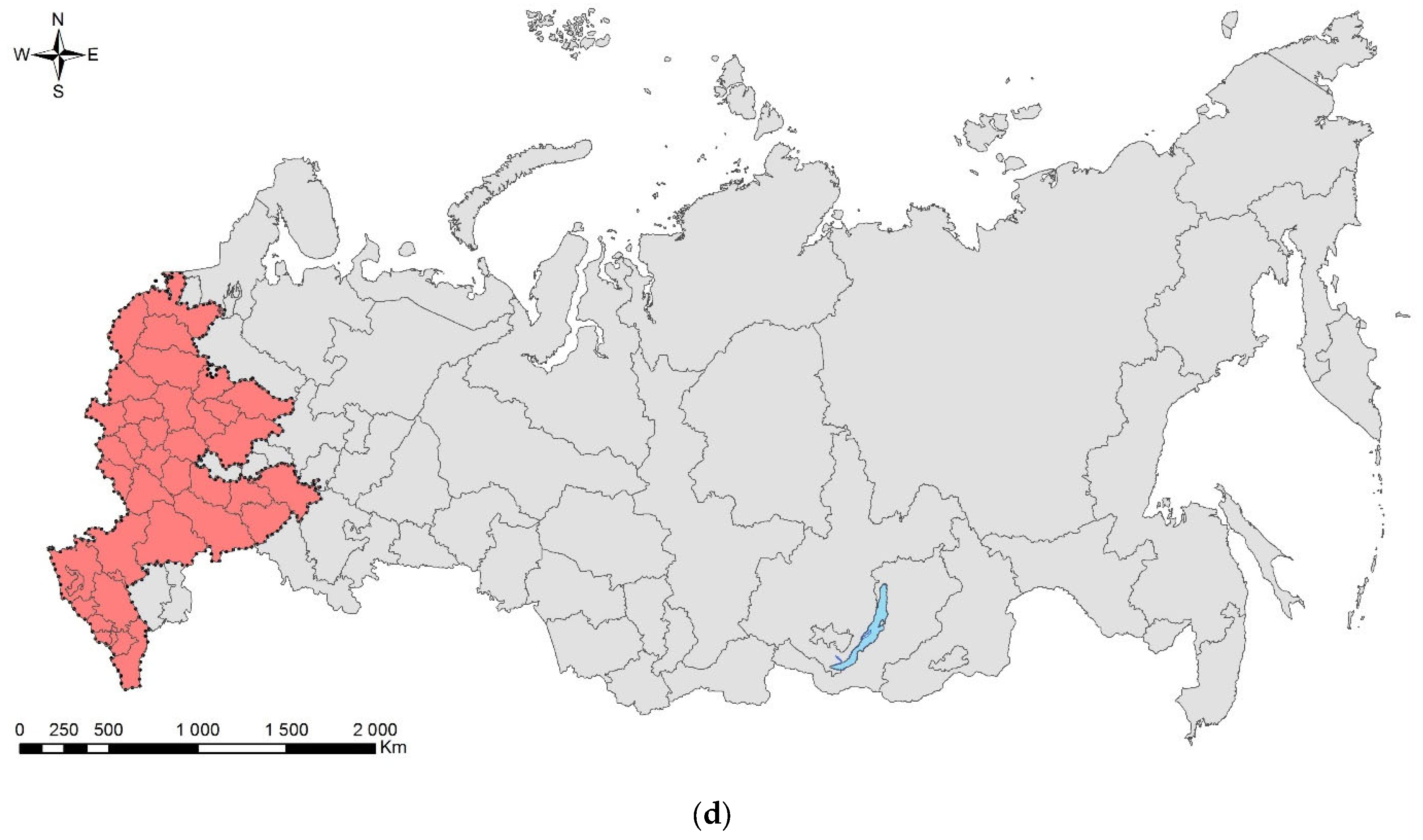{kind=link}
{kind=link}
{kind=link}
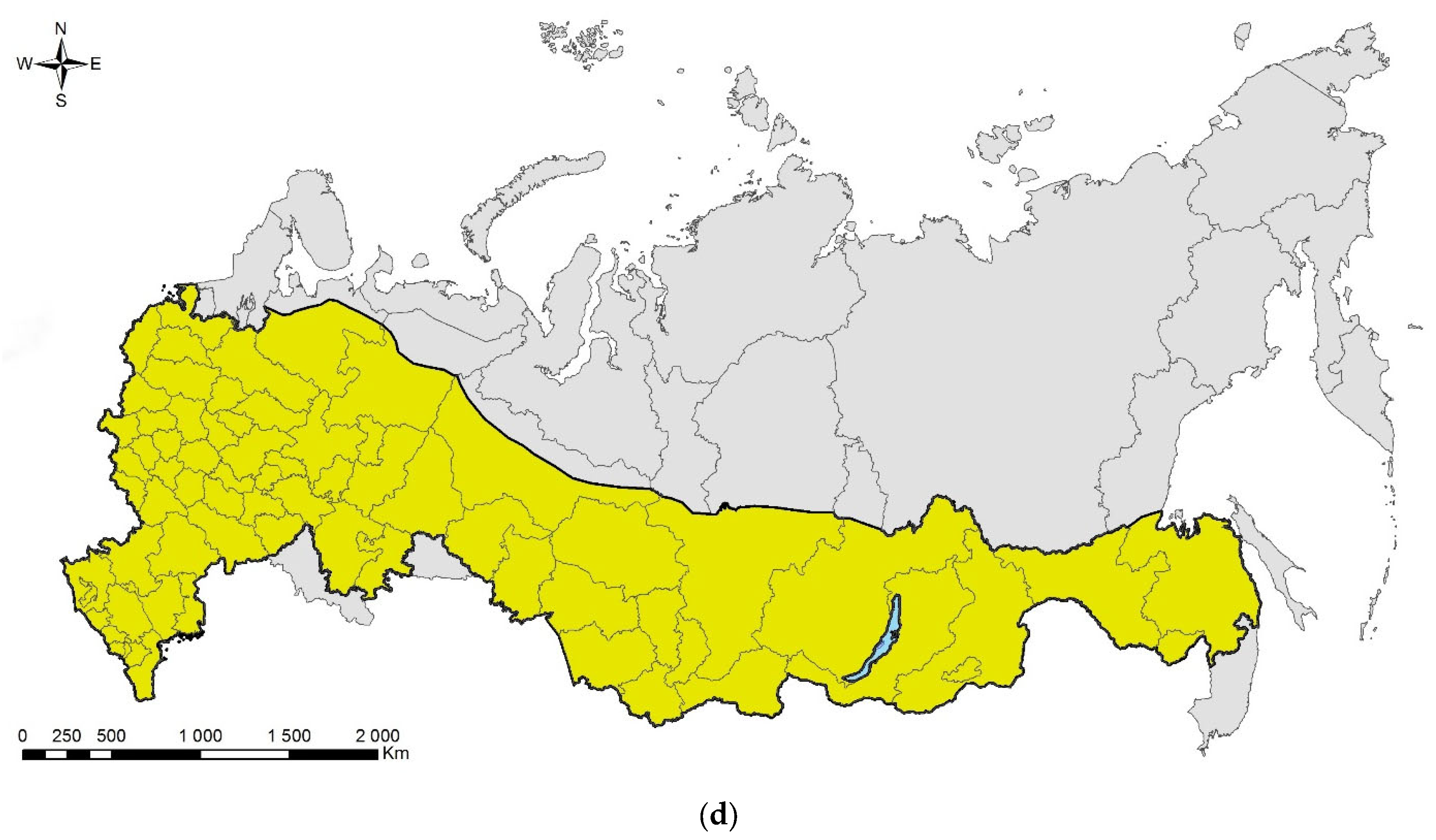{kind=link}
| No. | Species | Native Range | Invasive Range | Availability of Host Plants Outside Russia | The Regions and/or Countries to Where the Invasion Is Possible from Russia | Damage Level (in Forests/ Urban Areas) | Monitoring Measures * | Control Measures * | |
|---|---|---|---|---|---|---|---|---|---|
| General and in Forests | Specific in Urban Areas | ||||||||
| 1 | Conifer seed bug, Leptoglossus occidentalis (Hemiptera: Heteroptera: Coreidae) | West of North America: from South of Canada to Mexico | Most of Europe: from South of Norway to Mediterranean Sea, from British Isles to European Russia; Asia: Kazakhstan, Northeast China, Japan, South Korea; Africa: North and South; South America: Chile, Argentina, Uruguay | All conifer, native and introduced Pinaceae | Mongolia, Kyrgyzstan, Asian Turkey | Moderate to high level of damage in forests, coniferous tree nurseries, and urban areas | Visual inspection; analysis of seed damage and germination of seeds from infested trees | Biological and chemical control | Biological control |
| 2 | Brown marmorated stink bug, Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae) | Southeast Asia: China, Taiwan, Korea, Japan, Vietnam | North America; Europe (including South of European Russia); Asia: Kazakhstan | 300+ species of vascular plants | Transcaucasia, Middle Asia | Major damage in fruit, berry, and nut plantations | Inspections of overwintering quarters; visual inspection of host plants; pheromone traps | Quarantine measures; pheromone traps; biological and chemical control | Pheromone traps |
| 3 | Oak lace bug, Corythucha arcuata (Hemiptera: Heteroptera: Tingidae) | North America | Europe (including South of European Russia); Asia Minor | Quercus spp. and other Fagaceae, some species of Salicaceae, Rosaceae, Fabaceae | Middle Asia, Transcaucasia, Ukraine, Moldova, Belarus | Major in urban areas; moderate in forests | Inspections of overwintering quarters; visual inspection of host plants | Biological and chemical control | Biological control |
| 4 | Spotted poplar borer, Agrilus fleischeri (Coleoptera: Buprestidae) | North Asia: Russia (West and East Siberia, Far East), East Kazakhstan, Mongolia, Northeast and central China, North Korea, South Korea, Japan | unclear | Native and introduced Populus spp. and Salix spp. | West Kazakhstan, Europe (including east of European Russia), North America | In urban areas and forests (minor; during outbreaks—major) | Visual, color, and pheromone traps | Quarantine measures; sanitation felling; chemical control should be developed; biological control (egg and larval parasitoids) should be developed | Elimination of individual infested trees; chemical control should be developed (stem injections with insecticides); biological control (egg and larval parasitoids) should be developed |
| 5 | Apple buprestid, Agrilus mali (Coleoptera: Buprestidae) | East Asia: Russia (East Siberia, Far East), Mongolia, Northeast and central China, North Korea (**), South Korea (**) | Asia: China (**): Xinjiang, North Korea (**), South Korea (**) | Cultivated and wild Malus spp., Cydonia spp., Prunus spp., Pyrus spp. | Kazakhstan and other Central Asian countries; entire apple orchard cultivation area | In agricultural areas and forests (major) | Visual, color, and pheromone traps | Quarantine measures; biological control (egg and larval parasitoids) and chemical control should be developed | Pruning; elimination of individual infested trees; chemical control (stem injections with insecticides) and biological control (egg and larval parasitoids) should be developed |
| 6 | Emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae) | East Asia: Russian Far East, Northeast China, Japan, Mongolia, North Korea, South Korea | Europe: Central, Northwest, South and Southeast of European Russia, Eastern Ukraine; North America | Native and introduced Fraxinus spp., Chionanthus ssp., potentially other Oleaceae | Ukraine, Belarus, Poland, Baltic countries, Finland, and other European countries | In urban areas and forests (major) | Visual, color and pheromone traps; checking nests of Cerceris spp. (Hymenoptera) | Quarantine measures; sanitation felling; chemical control should be developed; biological control (egg and larval parasitoids) | Elimination of individual infested trees; chemical control (stem injections with insecticides); biological control (egg and larval parasitoids) |
| 7 | Cypress jewel beetle, Lamprodila (Palmar) festiva (Coleoptera: Buprestidae) | Europe: Mediterranean countries, South Europe, South of central Europe; North Africa; Southwestern Asia | Europe: Central and East Europe, South of European Russia, Eastern Ukraine | Native and introduced Cupressaceae | Transcaucasia, entire Ukraine, Moldova, Belorussia, Poland, Baltic countries, North America | In urban areas (major) and forests | Visual, color, and pheromone traps | Quarantine measures; sanitation felling; chemical control (insecticides during adult flight period); biological control (egg and larval parasitoids) should be developed | Elimination of infested trees; chemical control (pyrethroid insecticides during adult flight period, if allowed and stem injections with insecticides); biological control (egg and larval parasitoids) should be developed |
| 8 | Small spruce bark beetle, Ips amitinus (Coleoptera: Curculionidae: Scolytinae) | Europe: Central, South, and North Europe; Baltic countries; Northwest Russia | Asia: West Siberia | Different conifers, especially Pinus spp. and Picea spp. | China, North America | Major pest of all conifer forests; possible economic effect is unpredictable | Surveys; pheromone traps | Pheromone traps; sanitation felling | Pheromone traps; elimination of individual infested trees |
| 9 | Four-eyed fir bark beetle, Polygraphus proximus (Coleoptera: Curculionidae: Scolytinae) | Asia: Northeast China, Japan, Korea, Russian Far East | Europe: central Russia; Asia: West and East Siberia | Different conifers, especially Abies spp. | North and Central Europe, North America | Major pest of firs in forests and urban areas | Phytosanitary quarantine and surveys | Pheromone traps; sanitation felling | Pheromone traps; elimination of individual infested trees |
| 10 | Box tree moth, Cydalima perspectalis (Lepidoptera: Crambidae) | Asia: China, Japan, Korea, Russian Far East, India | Europe: South of European Russia, Georgia, Turkey | Buxus spp. | Natural forests across the Caucasus (Transcaucasia) and to countries located further south | Major in urban areas and forests | Visual, UV-light, or pheromone traps | Pheromone traps; biological and chemical control; sanitation felling | Pheromone traps; elimination of individual infested trees; biological and chemical control |
| 11 | Leaf blotch miner moth, Acrocercops brongniardella (Lepidoptera: Gracillariidae) | Europe | Europe: St. Petersburg and the Karelian Isthmus | Quercus spp. | Siberia, Far East, North America | Mainly in urban areas | Visual (mines); pheromone traps | Biological control (egg and larval parasitoids) should be developed | Stem injections with insecticides should be developed |
| 12 | Horse-chestnut leaf miner, Cameraria ohridella (Lepidoptera: Gracillariidae) | Europe: The Balkans | Europe: East and West Europe, European Russia (except some northern and eastern regions) | Native and introduced Aesculus spp., and possibly Acer spp. | East Asia, East and West of North America | Mainly in urban areas | Visual; pheromone traps (might be not efficient al low density) | Biological control (parasitoids, disease agents) | Stem injections with insecticides; removal of leaf litter (populations with overwintering of pupae) |
| 13 | Lime leaf miner, Phyllonorycter issikii (Lepidoptera: Gracillariidae) | East Asia: Japan, South Korea, China, Russian Far East | Europe: East and West Europe, European Russia (except some northern and southern regions); Asia: Western Siberia | Native and introduced Tilia spp. | North America | Mainly in urban areas, occasionally in forests | Visual; pheromone traps (might be not efficient al low density) | Biological control (parasitoids, disease agents) | Protection of natural enemies |
| 14 | Poplar leafminer, Phyllonorycter populifoliella (Lepidoptera: Gracillariidae) | Eurasia | Europe: North of European Russia; Asia: Siberia | Populus spp. | Far East, and North America | In urban areas | Visual (mines); pheromone traps | Biological control (egg and larval parasitoids) should be developed | Stem injections with insecticides should be developed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musolin, D.L.; Kirichenko, N.I.; Karpun, N.N.; Aksenenko, E.V.; Golub, V.B.; Kerchev, I.A.; Mandelshtam, M.Y.; Vasaitis, R.; Volkovitsh, M.G.; Zhuravleva, E.N.; et al. Invasive Insect Pests of Forests and Urban Trees in Russia: Origin, Pathways, Damage, and Management. Forests 2022, 13, 521. https://doi.org/10.3390/f13040521
Musolin DL, Kirichenko NI, Karpun NN, Aksenenko EV, Golub VB, Kerchev IA, Mandelshtam MY, Vasaitis R, Volkovitsh MG, Zhuravleva EN, et al. Invasive Insect Pests of Forests and Urban Trees in Russia: Origin, Pathways, Damage, and Management. Forests. 2022; 13(4):521. https://doi.org/10.3390/f13040521
Chicago/Turabian StyleMusolin, Dmitry L., Natalia I. Kirichenko, Natalia N. Karpun, Evgeniy V. Aksenenko, Viktor B. Golub, Ivan A. Kerchev, Michail Y. Mandelshtam, Rimvys Vasaitis, Mark G. Volkovitsh, Elena N. Zhuravleva, and et al. 2022. "Invasive Insect Pests of Forests and Urban Trees in Russia: Origin, Pathways, Damage, and Management" Forests 13, no. 4: 521. https://doi.org/10.3390/f13040521