
Secondary Succession in the Tropical Lowland Rainforest Reduced the Stochasticity of Soil Bacterial Communities through the Stability of Plant Communities

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Experimental Design, Plant Community Surveying and Soil Sampling
2.3. Analysis of Soil Properties
2.4. Soil Samples DNA Extraction, PCR Amplification, and Sequencing
2.4.1. DNA Extraction
2.4.2. PCR Amplification
2.4.3. Sequence Processing
2.5. Data Analysis
3. Results
3.1. Environmental Factors Differences at Successional Stages
3.2. Plant Important Value and Niche Width
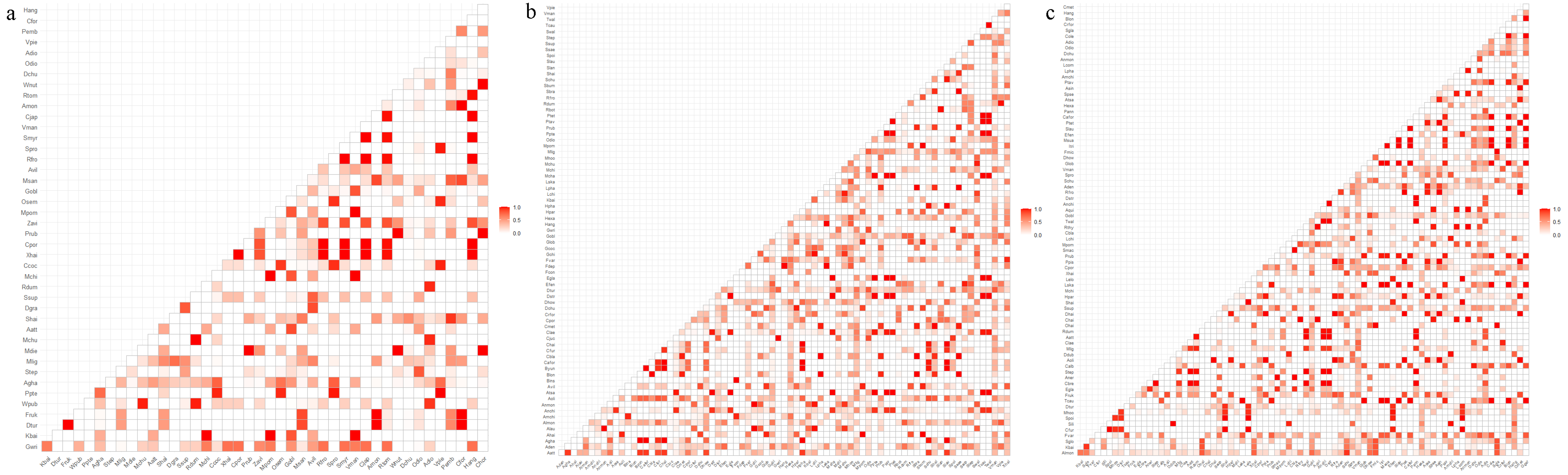
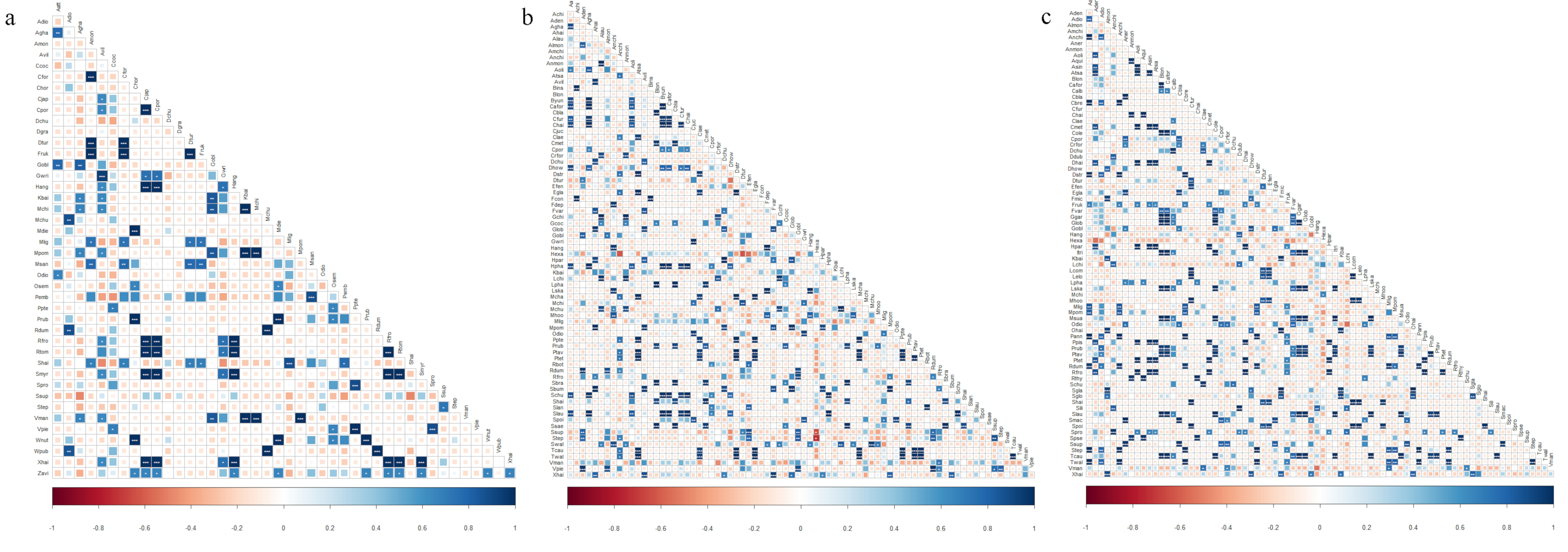
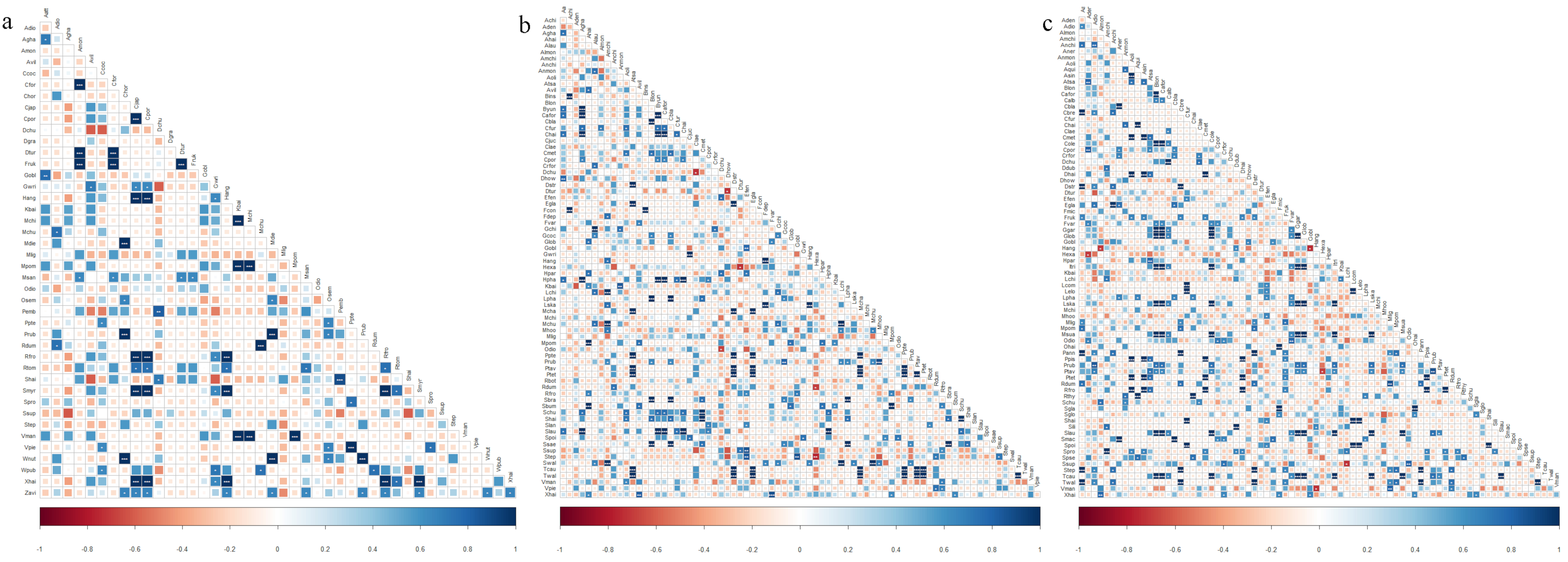
3.3. Plant Niche Overlap and Interspecific Associations
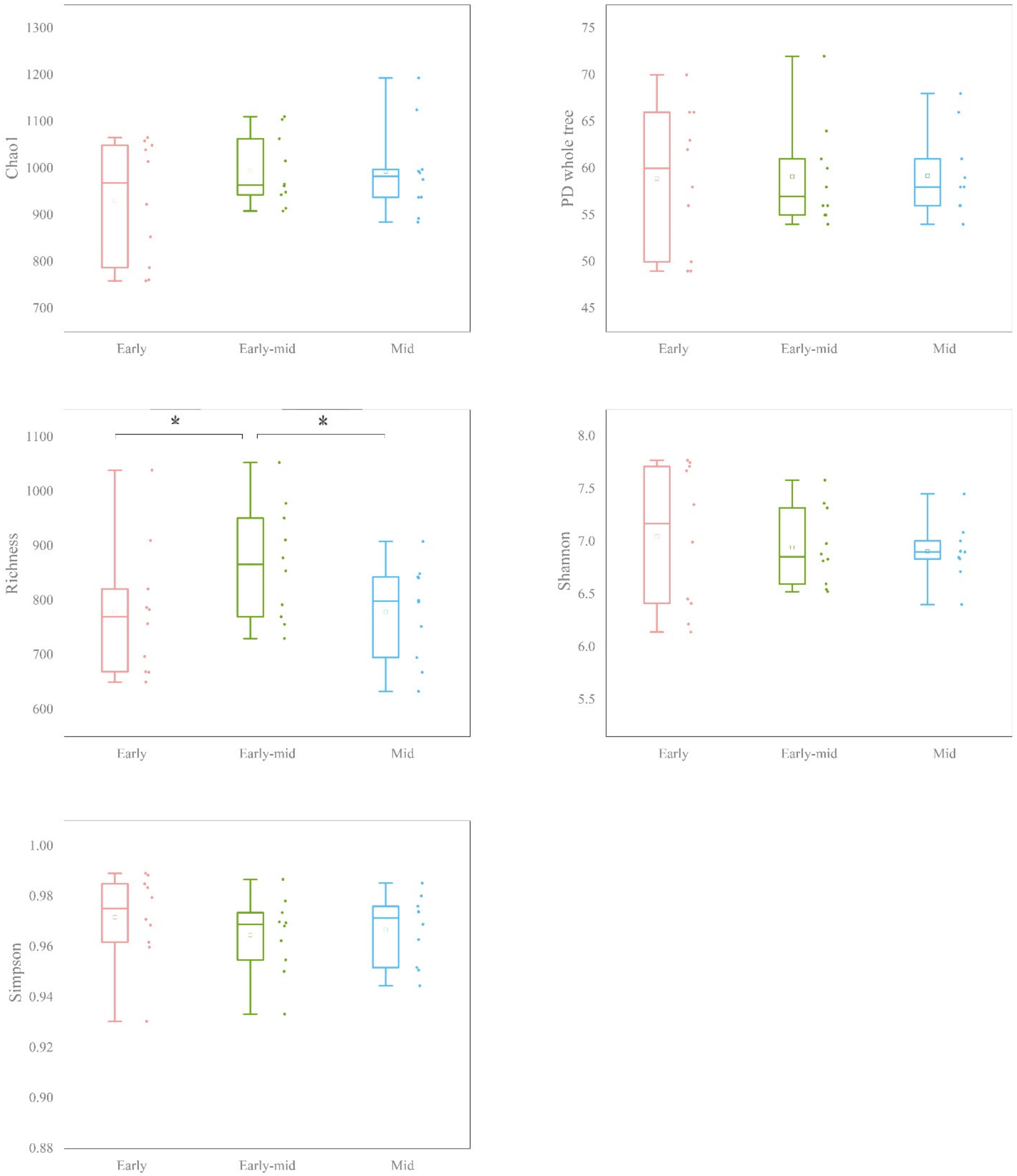
3.4. Soil Bacterial Diversity and the Influence of Successional Stage on Bacterial Community Structure
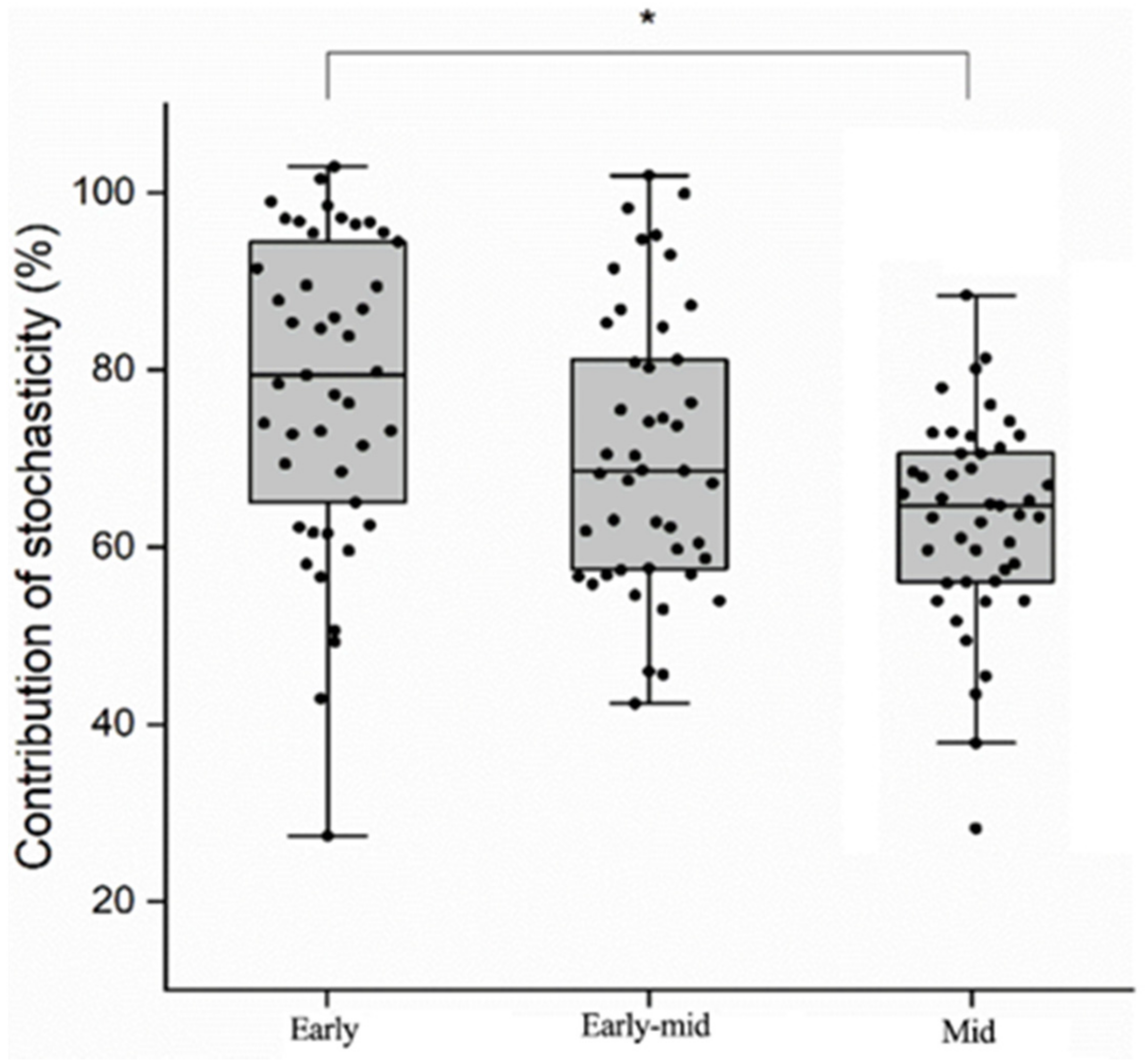
3.5. The Stochasticity of Soil Bacterial Communities
4. Discussion
4.1. The Development Direction of Plant Community Structure and Diversity
4.2. Changes in Plant Communities, Soil Environmental Factors and Soil Bacterial Communities
4.3. Succession of Forest Plant Communities and the Stochastic of Soil Bacterial Community Establishment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Townsend, A.R.; Cleveland, C.C.; Houlton, B.Z.; Alden, C.B.; White, J.W. Multi-element regulation of the tropical forest carbon cycle. Front. Ecol. Environ. 2011, 9, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Hayes, D. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Baccini, A.; Walker, W.; Carvalho, L.; Farina, M.; Sulla-Menashe, D.; Houghton, R.A. Tropical forests are a net carbon source based on aboveground measurements of gain and loss. Science 2017, 358, 230–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, J.E.; Evans, T.; Venter, O.; Williams, B.; Tulloch, A.; Stewart, C.; Lindenmayer, D. The exceptional value of intact forest ecosystems. Nat. Ecol. Evol. 2018, 2, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, G.; Xue, S.; Wang, G. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Herrera Paredes, S.; Lebeis, S.L. Giving back to the community: Microbial mechanisms of plant–soil interactions. Funct. Ecol. 2016, 30, 1043–1052. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Long, J.; Liao, H.; Zheng, C.; Li, J.; Liu, L.; Zhang, M. Dynamics of soil microbial communities following vegetation succession in a karst mountain ecosystem, Southwest China. Sci. Rep. 2019, 9, 2160. [Google Scholar]
- Liu, Y.; Zhu, G.; Hai, X.; Li, J.; Shangguan, Z.; Peng, C.; Deng, L. Long-term forest succession improves plant diversity and soil quality but not significantly increase soil microbial diversity: Evidence from the Loess Plateau. Ecol. Eng. 2020, 142, 105631. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, S.; Li, X.; Wang, J.; Ding, Q.; Wang, H.; Huang, Y. Changes of soil prokaryotic communities after clear-cutting in a karst forest: Evidences for cutting-based disturbance promoting deterministic processes. FEMS Microbiol. Ecol. 2016, 92, fiw026. [Google Scholar] [CrossRef]
- Hu, X.; Xu, R.; Shu, Q.; Guo, W.; Zhang, J.; Shang, Z.; Qi, L. Population Structure and Dynamics of Hopea reticulata, a Plant Endemic to Ganshiling, Hainan Island. Chin. J. Trop. Crops 2020, 41, 1939–1945. [Google Scholar]
- Qi, L.H.; Liang, C.Q.; Mao, C. Species composition and geographic elements of the tropical lowland secondary rain forest of Ganshiling, Hainan Island, China. Chin. J. Ecol. 2014, 33, 922. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy; United States of Agriculture, Soil Conservation Service: Washington, DC, USA, 2014.
- Yan, J.A. The Study on Evolutional History of Hainan Island’s Ecological Environment-From the Point of Animal and Plant; Nanjing Agricultural University: Nanjing, China, 2006. [Google Scholar]
- Xu, R.J.; Hu, X.; Qi, L.H.; Liu, G.L.; Peng, C.; Shu, Q. Effects of forest restoration on distribution and growth of rattan in lowland rainforest, Hainan Island. Chin. J. Ecol. 2019, 38, 3313–3319. [Google Scholar]
- Cole, L.E.S.; Bhagwat, S.A.; Willis, K.J. Recovery and resilience of tropical forests after disturbance. Nat. Commun. 2014, 5, 3906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, C. Study on Site Adaptability of Rattan to Environment in Lowland Secondary Rain Forest in Ganzhalin of Hainan Island, China; Chinese Academy of Forestry: Beijing, China, 2017. [Google Scholar]
- Krishna, M.; Gupta, S.; Delgado–Baquerizo, M.; Morriën, E.; Garkoti, S.C.; Chaturvedi, R.; Ahmad, S. Successional trajectory of bacterial communities in soil are shaped by plant-driven changes during secondary succession. Sci. Rep. 2020, 10, 9864. [Google Scholar]
- Li, J.Q. Forest Ecology; Higher Education Press: Beijing, China, 2010. [Google Scholar]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Ning, D.; Deng, Y.; Tiedje, J.M.; Zhou, J. A general framework for quantitatively assessing ecological stochasticity. Proc. Natl. Acad. Sci. USA 2019, 116, 16892–16898. [Google Scholar] [CrossRef] [Green Version]
- Lebrija-Trejos, E.; Meave, J.A.; Poorter, L.; Pérez-García, E.A.; Bongers, F. Pathways, mechanisms and predictability of vegetation change during tropical dry forest succession. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 267–275. [Google Scholar] [CrossRef]
- Myster, R.W. Plants replacing plants: The future of community modeling and research. Bot. Rev. 2012, 78, 2–9. [Google Scholar] [CrossRef]
- Letcher, S.G. Phylogenetic structure of angiosperm communities during tropical forest succession. Proc. R. Soc. B Biol. Sci. 2010, 277, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Kopecký, M.; Hédl, R.; Szabó, P. Non-random extinctions dominate plant community changes in abandoned coppices. J. Appl. Ecol. 2013, 50, 79–87. [Google Scholar] [CrossRef]
- Csecserits, A.; Halassy, M.; Lhotsky, B.; Rédei, T.; Somay, L.; Botta-Dukát, Z. Changing assembly rules during secondary succession: Evidence for non-random patterns. Basic Appl. Ecol. 2021, 52, 46–56. [Google Scholar] [CrossRef]
- Pastore, A.I.; Barabás, G.; Bimler, M.D.; Mayfield, M.M.; Miller, T.E. The evolution of niche overlap and competitive differences. Nat. Ecol. Evol. 2021, 5, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q. Population Structure and Community Characteristics of Vatica Mangachapoi in Ganshiling, Hainan Island; Chinese Academy of Forestry: Beijing, China, 2021. [Google Scholar]
- Zhang, H.; Chu, L.M. Plant community structure, soil properties and microbial characteristics in revegetated quarries. Ecol. Eng. 2011, 37, 1104–1111. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Bradford, M.A.; Pernilla Brinkman, E.; van de Voorde, T.F.; Veen, G.F. Where, when and how plant–soil feedback matters in a changing world. Funct. Ecol. 2016, 30, 1109–1121. [Google Scholar] [CrossRef]
- Pizano, C.; Kitajima, K.; Graham, J.H.; Mangan, S.A. Negative plant–soil feedbacks are stronger in agricultural habitats than in forest fragments in the tropical Andes. Ecology 2019, 100, e02850. [Google Scholar] [CrossRef]
- Leff, J.W.; Bardgett, R.D.; Wilkinson, A.; Jackson, B.G.; Pritchard, W.J.; De Long, J.R.; Fierer, N. Predicting the structure of soil communities from plant community taxonomy, phylogeny, and traits. ISME J. 2018, 12, 1794–1805. [Google Scholar] [CrossRef] [Green Version]
- Bardgett, R.D.; Wardle, D.A. Aboveground-Belowground Linkages: Biotic Interactions, Ecosystem Processes, and Global Change; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; DellaSala, D.A.; Hutto, R.L.; Swanson, F.J. The forgotten stage of forest succession: Early-successional ecosystems on forest sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Abrahão, A.; Ryan, M.H.; Laliberté, E.; Oliveira, R.S.; Lambers, H. Phosphorus-and nitrogen-acquisition strategies in two Bossiaea species (Fabaceae) along retrogressive soil chronosequences in south-western Australia. Physiol. Plant. 2018, 163, 323–343. [Google Scholar] [CrossRef]
- Ramos, S.J.; Gastauer, M.; Mitre, S.K.; Caldeira, C.F.; Silva, J.R.; Furtini Neto, A.E.; Siqueira, J.O. Plant growth and nutrient use efficiency of two native Fabaceae species for mineland revegetation in the eastern Amazon. J. For. Res. 2020, 31, 2287–2293. [Google Scholar] [CrossRef]
- Kuramae, E.; Gamper, H.; van Veen, J.; Kowalchuk, G. Soil and plant factors driving the community of soil-borne microorganisms across chronosequences of secondary succession of chalk grasslands with a neutral pH. FEMS Microbiol. Ecol. 2011, 77, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Jangid, K.; Whitman, W.B.; Condron, L.M.; Turner, B.L.; Williams, M.A. Soil bacterial community succession during long-term ecosystem development. Mol. Ecol. 2013, 22, 3415–3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, I.C.; Finzi, A.C.; Phillips, R.P. Root exudates increase N availability by stimulating microbial turnover of fast-cycling N pools. Soil Biol. Biochem. 2017, 106, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Zhao, J.; Yuan, J.; Hale, L.; Wen, T.; Huang, Q.; Shen, Q. Root exudates drive soil-microbe-nutrient feedbacks in response to plant growth. Plant Cell Environ. 2021, 44, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.F.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere interactions: Root exudates, microbes, and microbial communities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- Chase, J.M.; Myers, J.A. Disentangling the importance of ecological niches from stochastic processes across scales. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2351–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Deng, Y.E.; Zhang, P.; Xue, K.; Liang, Y.; Van Nostrand, J.D.; Arkin, A.P. Stochasticity, succession, and environmental perturbations in a fluidic ecosystem. Proc. Natl. Acad. Sci. USA 2014, 111, E836–E845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Ning, D. Stochastic community assembly: Does it matter in microbial ecology? Microbiol. Mol. Biol. Rev. 2017, 81, e00002-17. [Google Scholar] [CrossRef] [Green Version]
- Dini-Andreote, F.; Stegen, J.C.; Van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Early | Early-Mid | Mid | |
|---|---|---|---|
| Altitude (m) | 243.00 ± 8.17 a | 233.00 ± 23.11 a | 238.50 ± 29.19 a |
| pH | 4.69 ± 0.04 a | 4.74 ± 0.06 a | 4.79 ± 0.07 a |
| Organic matter (g/kg) | 10.54 ± 1.38 b | 24.27 ± 2.30 a | 19.29 ± 2.84 a |
| Total organic C (g/kg) | 6.11 ± 0.8 b | 14.08 ± 1.33 a | 11.19 ± 1.65 a |
| Total N (g/kg) | 0.45 ± 0.05 b | 1.19 ± 0.09 a | 0.98 ± 0.12 a |
| Total P (g/kg) | 0.12 ± 0.01 a | 0.14 ± 0.01 a | 0.18 ± 0.03 a |
| Alkali N (mg/kg) | 58.9 ± 7.03 b | 102.27 ± 9.16 a | 90.88 ± 13.62 a |
| Available P (mg/kg) | 0.11 ± 0.01 b | 0.53 ± 0.13 a | 0.54 ± 0.17 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, X.; Shu, Q.; Shang, Z.; Guo, W.; Qi, L. Secondary Succession in the Tropical Lowland Rainforest Reduced the Stochasticity of Soil Bacterial Communities through the Stability of Plant Communities. Forests 2022, 13, 348. https://doi.org/10.3390/f13020348
Hu X, Shu Q, Shang Z, Guo W, Qi L. Secondary Succession in the Tropical Lowland Rainforest Reduced the Stochasticity of Soil Bacterial Communities through the Stability of Plant Communities. Forests. 2022; 13(2):348. https://doi.org/10.3390/f13020348
Chicago/Turabian StyleHu, Xuan, Qi Shu, Zean Shang, Wen Guo, and Lianghua Qi. 2022. "Secondary Succession in the Tropical Lowland Rainforest Reduced the Stochasticity of Soil Bacterial Communities through the Stability of Plant Communities" Forests 13, no. 2: 348. https://doi.org/10.3390/f13020348