
Upward Treeline Shifts in Two Regions of Subarctic Russia Are Governed by Summer Thermal and Winter Snow Conditions

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
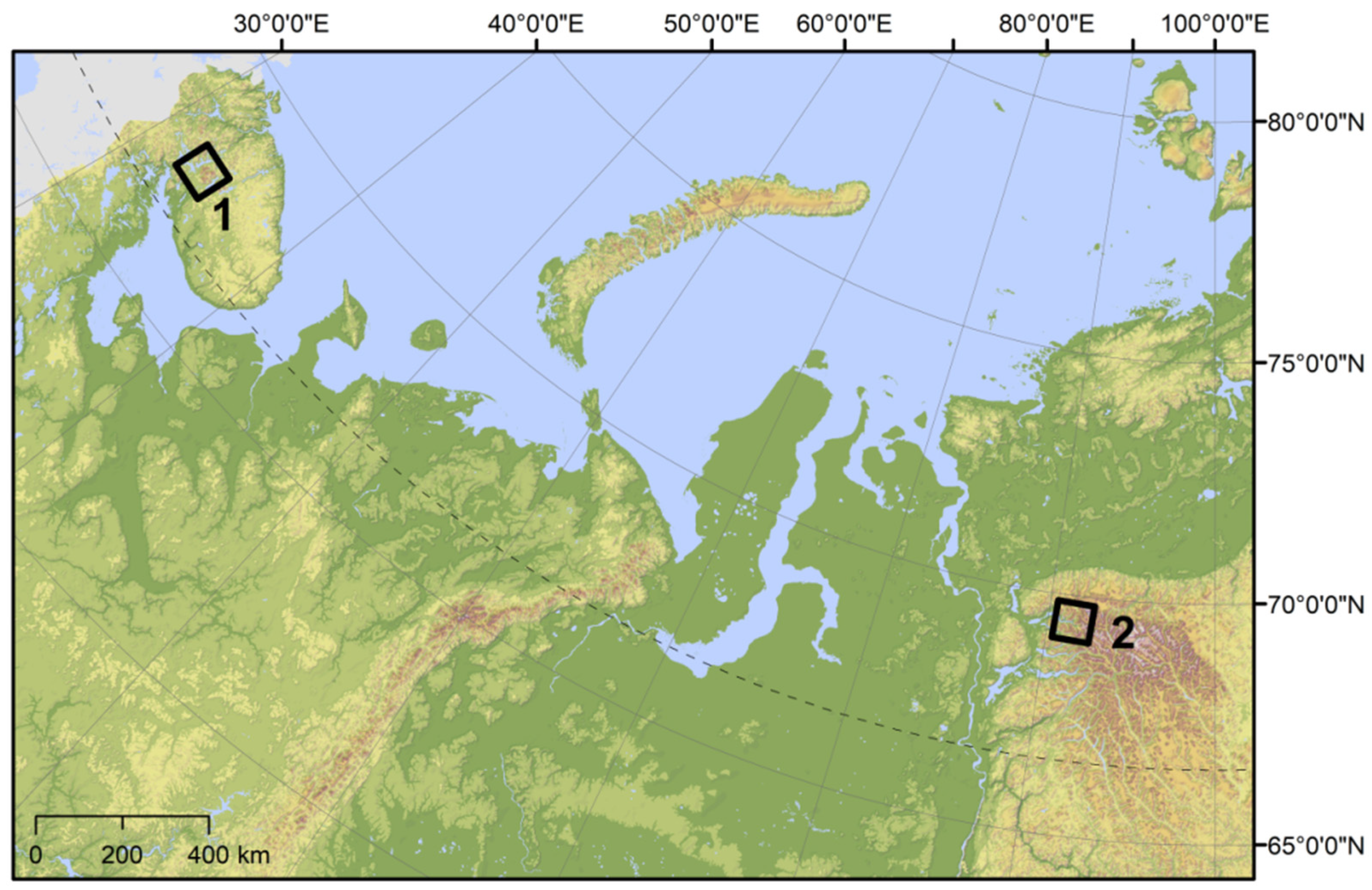
2.1. Study Sites
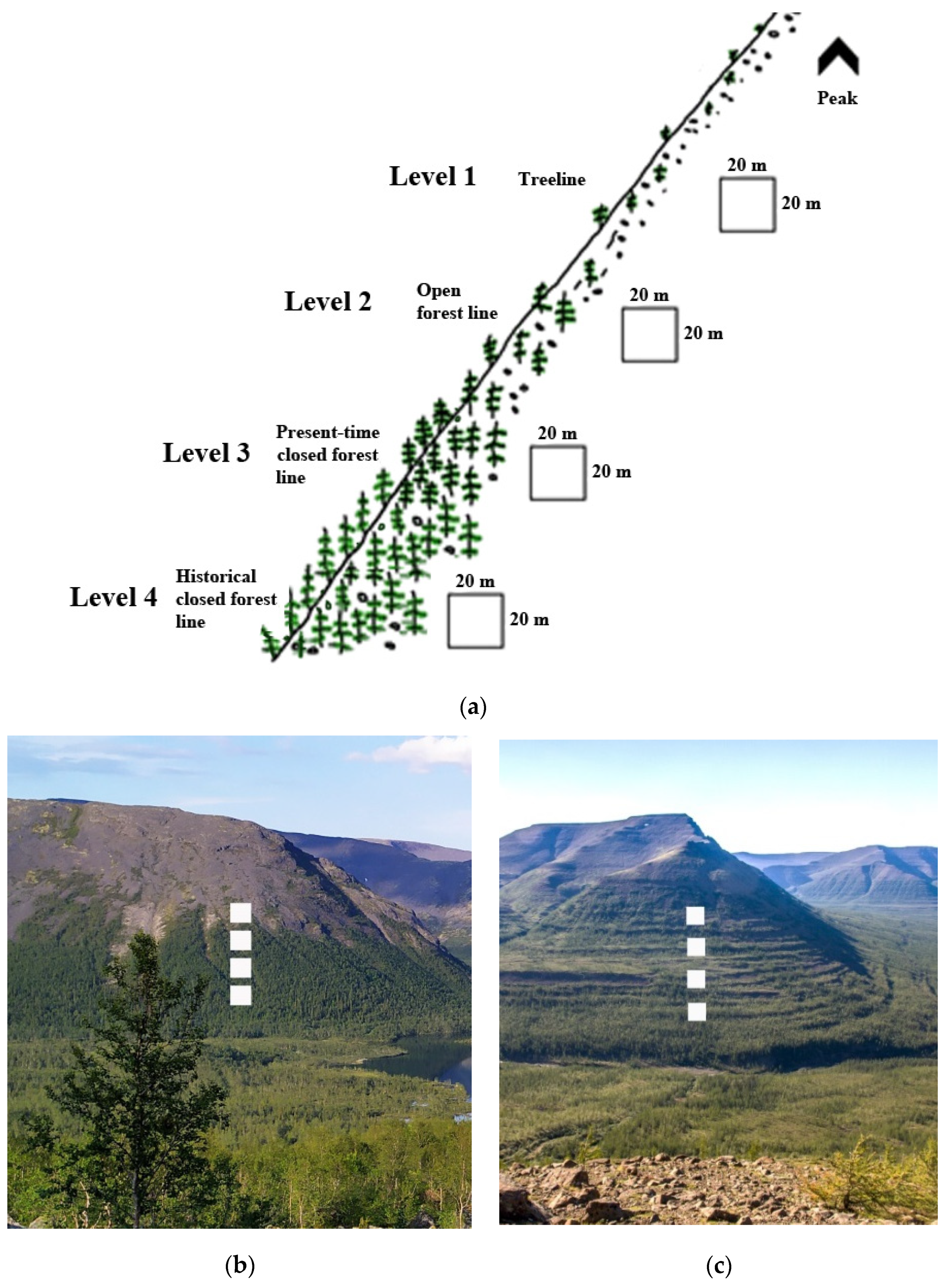
2.2. Studied Treelines
2.3. Field Sampling
2.4. Climate Data and Statistical Analysis
2.5. In Situ Air Temperature and Snow Measurements
2.6. Statistical Analyses
3. Results
3.1. Microclimatic Conditions
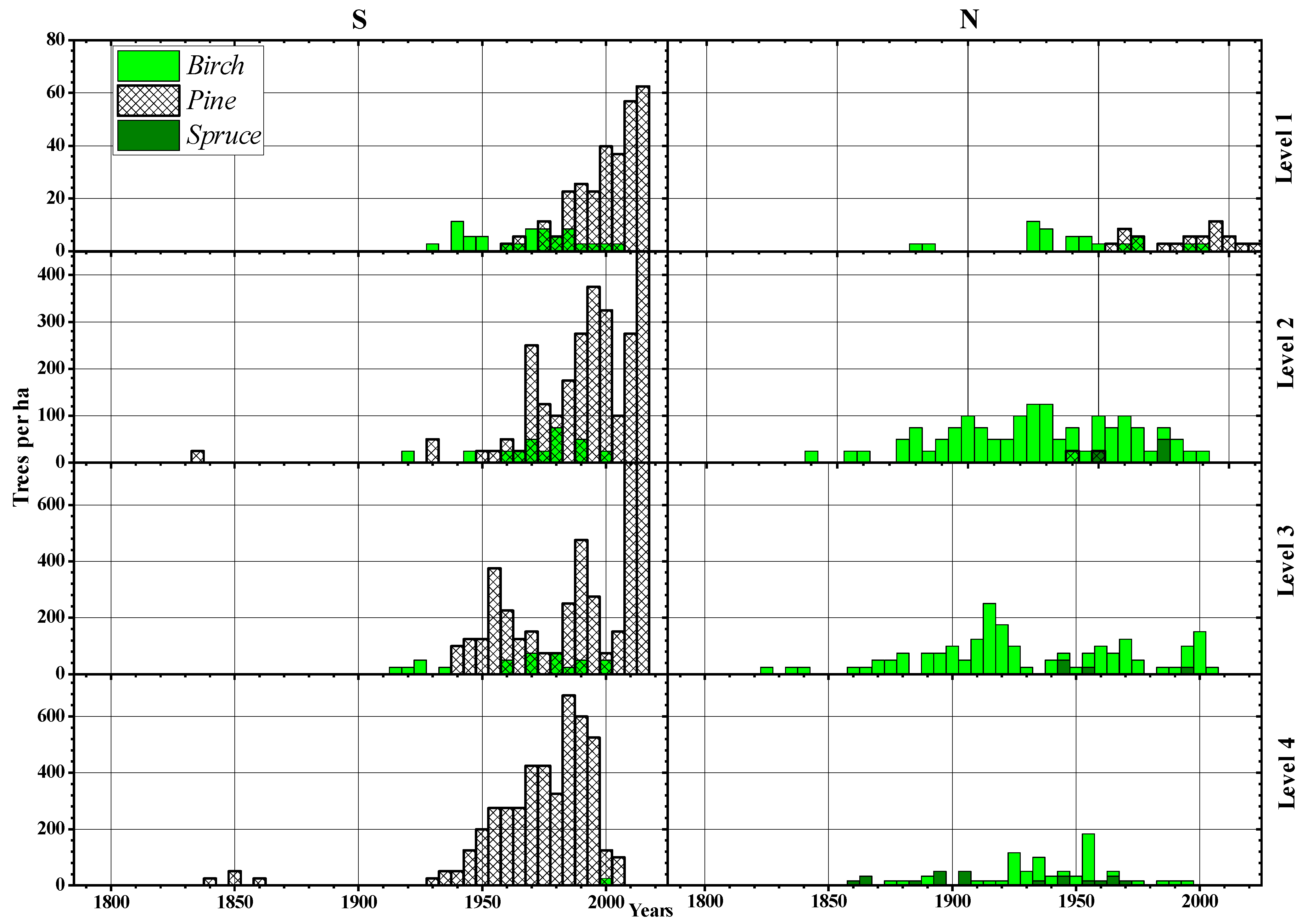
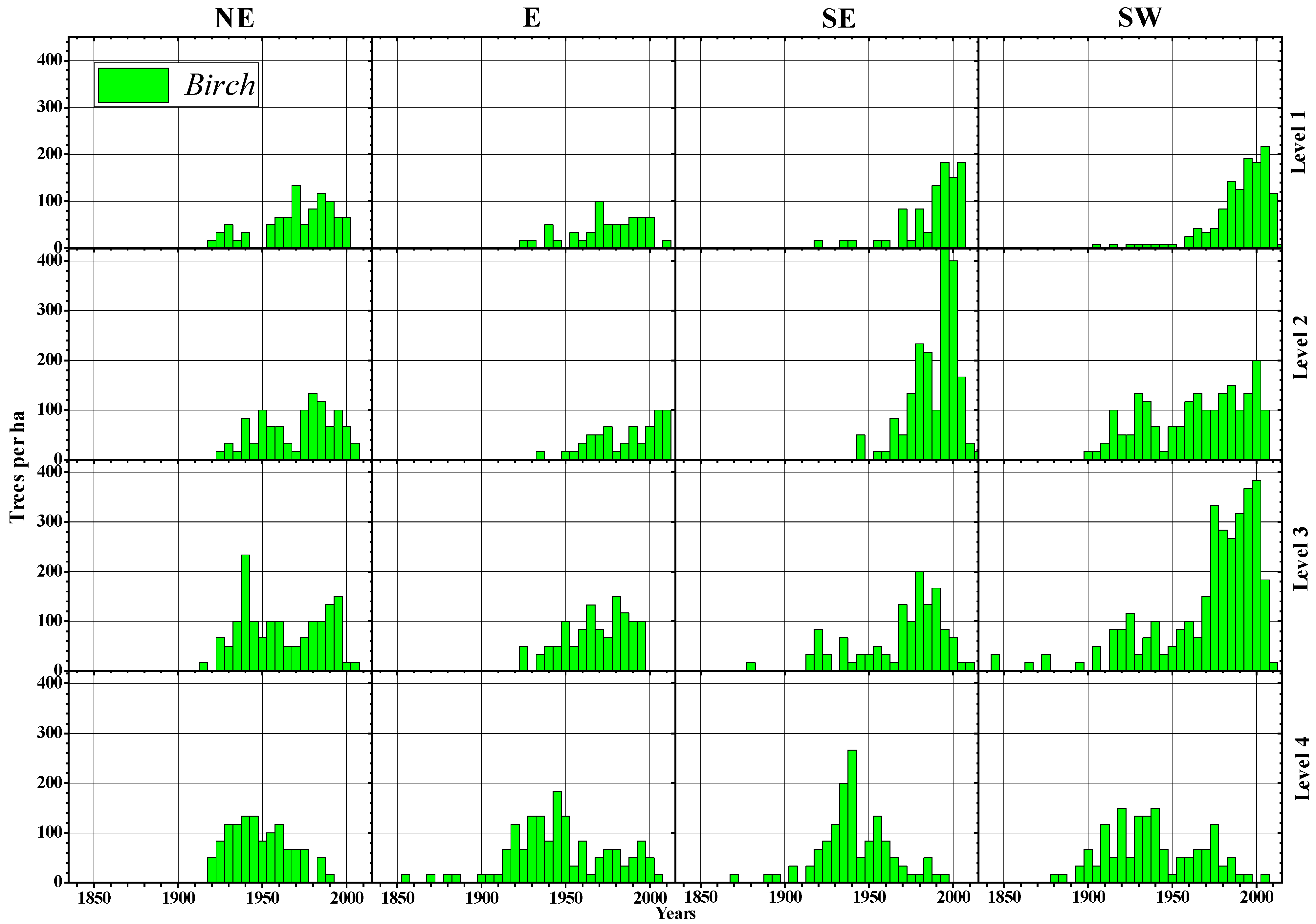
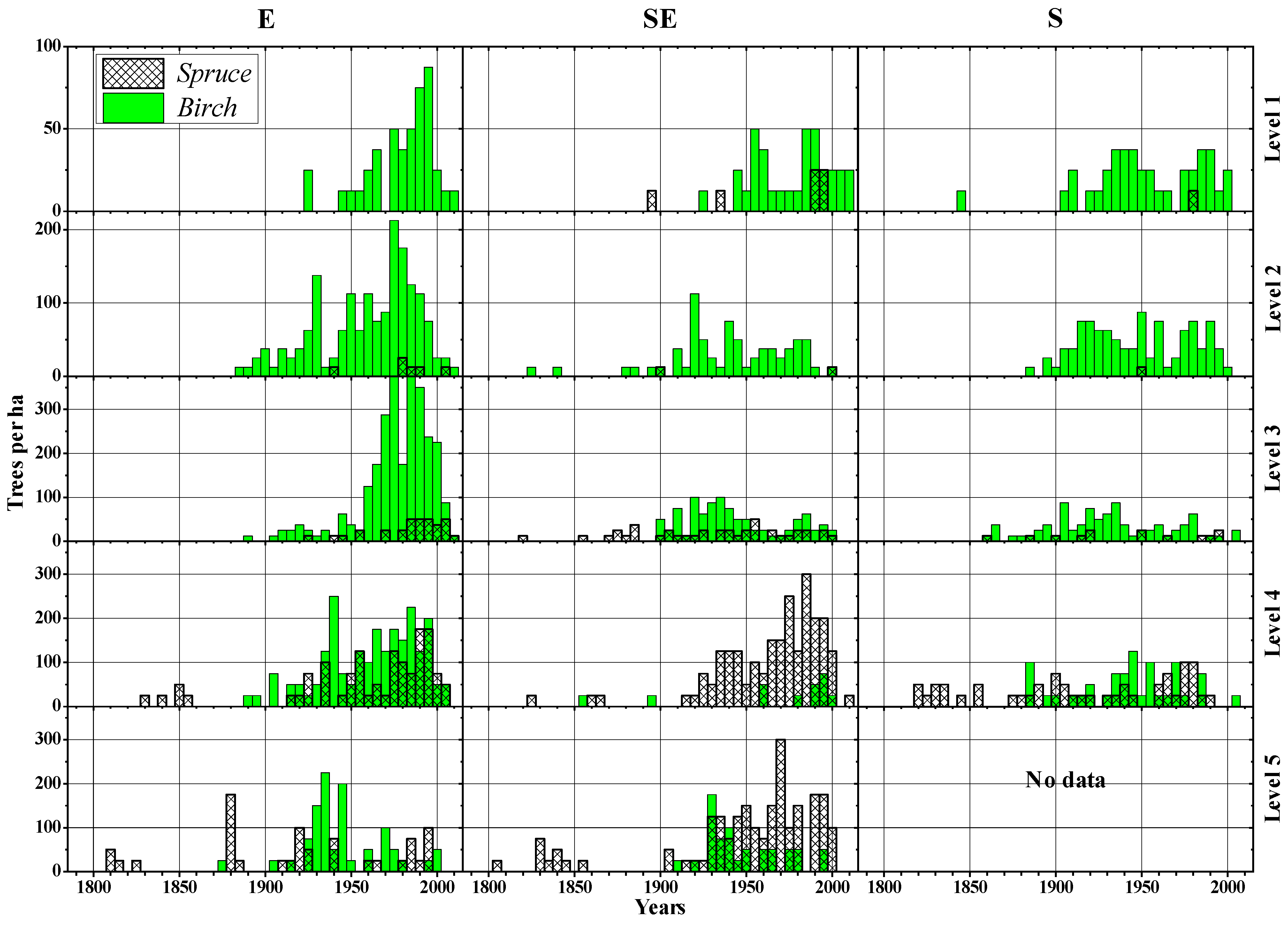
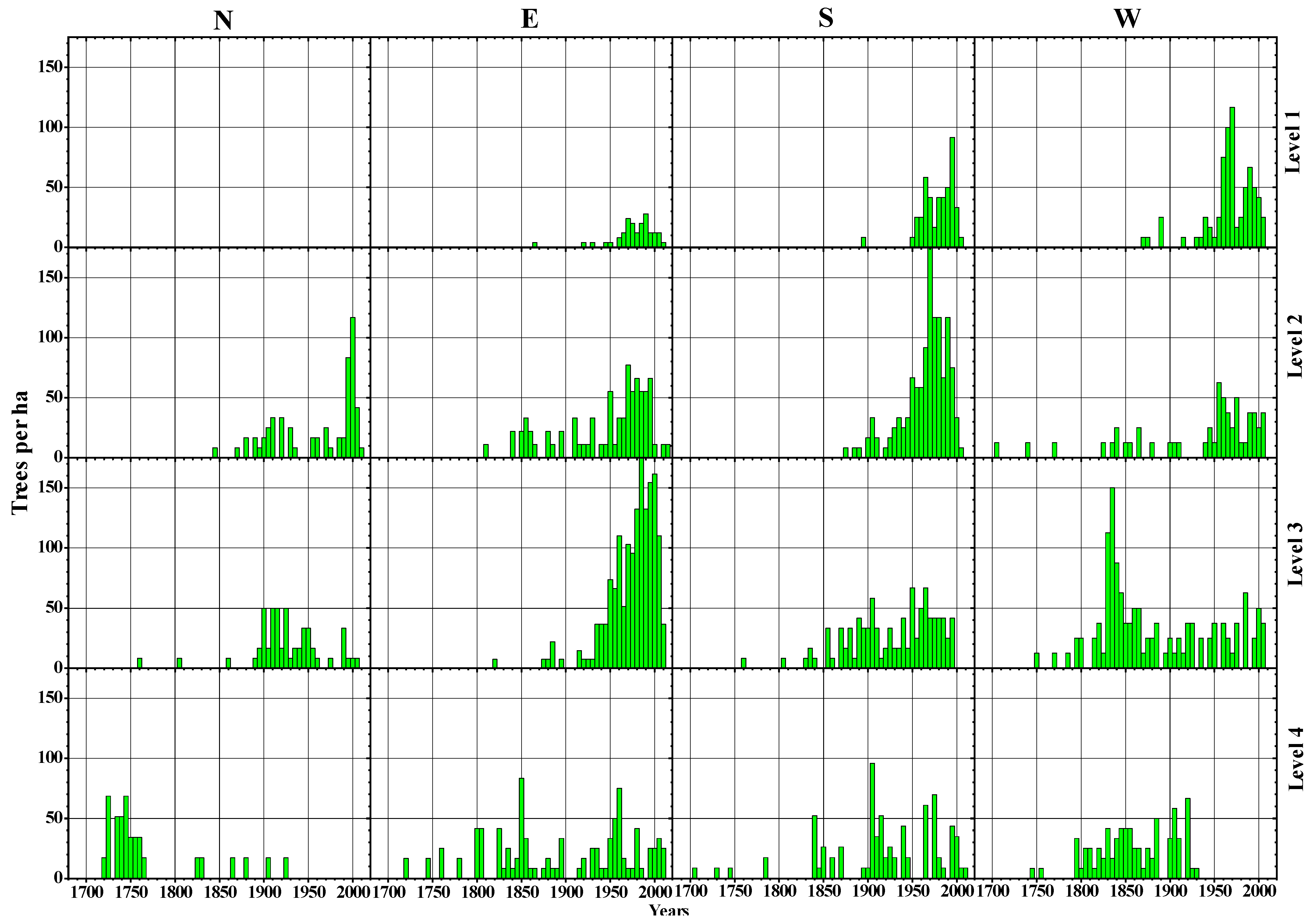
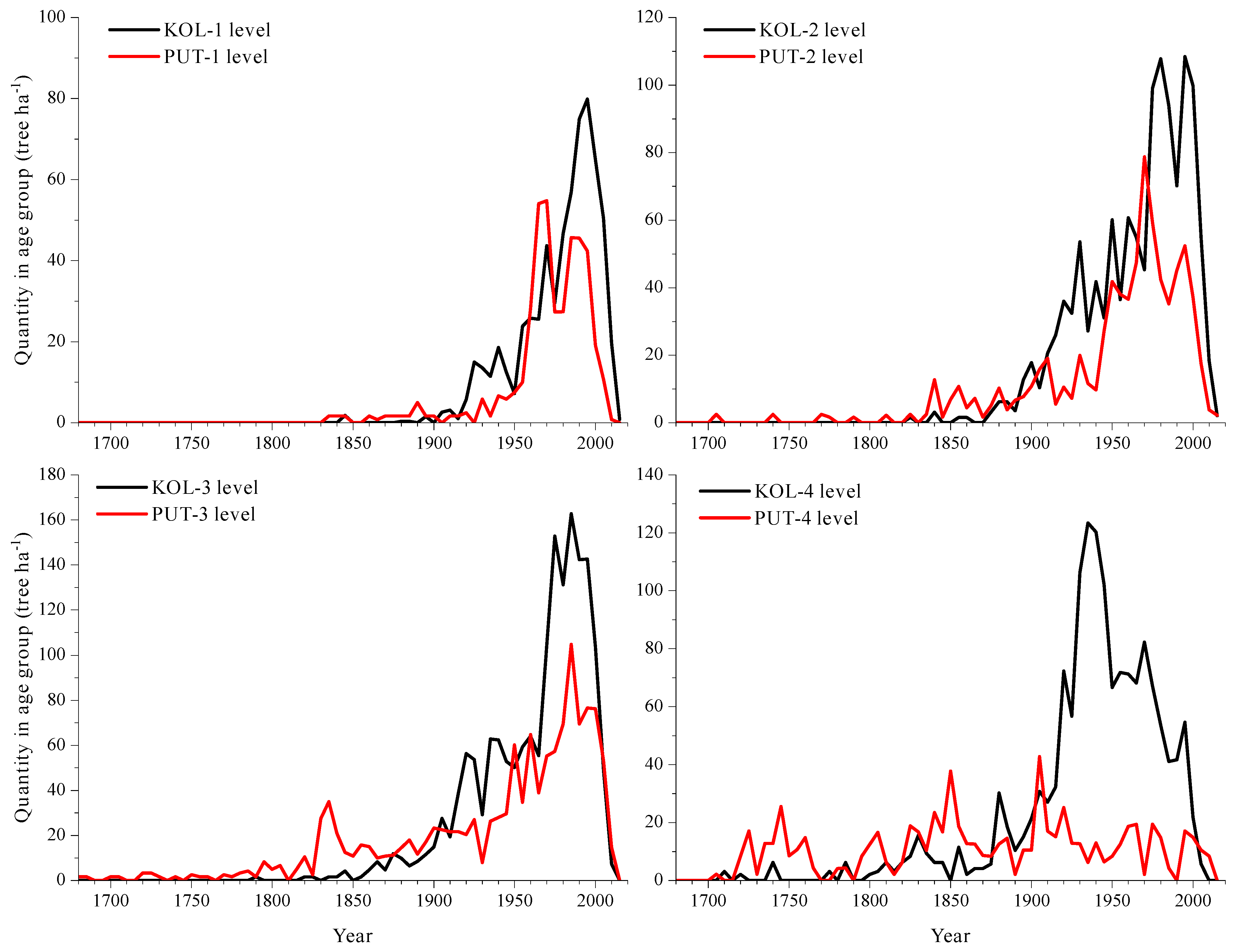
3.2. Morphological Features and Age Structures
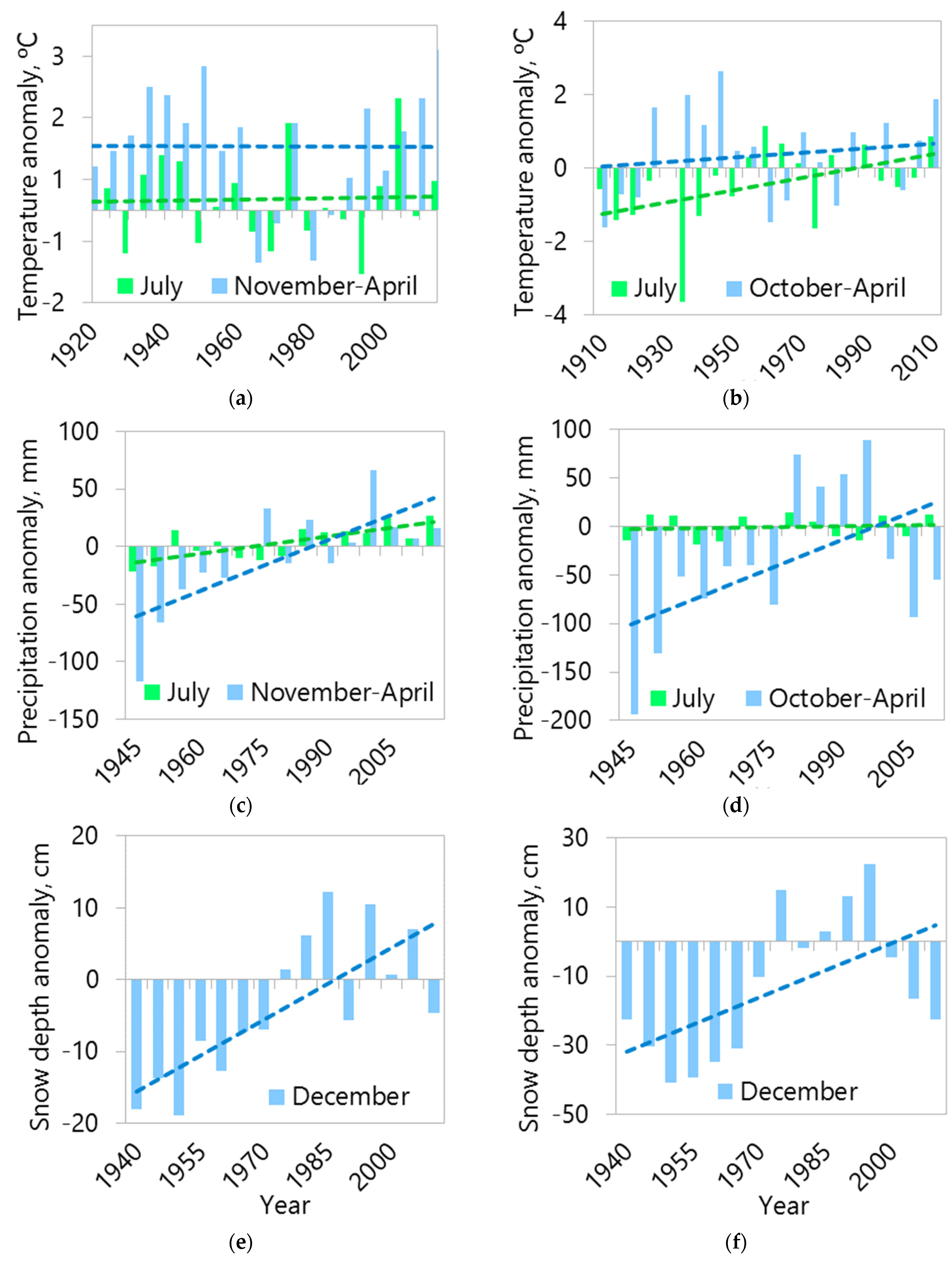
3.3. Analysis of Changes in Climatic Conditions
3.4. Relationship of Climate Variability with Tree Expansion: Roles of Altitude and Slope Exposure
4. Discussion
4.1. Treeline Characteristics and Slope Exposure
4.2. Drivers of Treeline Dynamics
4.3. Regional Climate and Treeline Features
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. 2021: Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; in press. [Google Scholar]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Körner, C. Alpine treelines. In Functional Ecology of the Global High Elevation Tree Limits; Springer: Berlin, Germany, 2012. [Google Scholar]
- Kullman, L.; Öberg, L. Post-little ice age treeline rise and climatic warming in the Swedish Scandes: A landscape ecological perspective. J. Ecol. 2009, 97, 415–429. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Shiyatov, S.G.; Kasishke, E.; Fedotova, E.V.; Naurzbaev, M.M. Response of the forest-tundra ecotone to climate change. Probl. Ecol. Monit. Model. Ecosyst. 2002, 18, 234–260. [Google Scholar]
- Callaghan, T.V.; Tweedie, C.E.; Akerman, J.; Andrews, C.; Bergstedt, J.; Butler, M.G.; Christensen, T.R.; Cooley, D.; Dahlberg, U.; Dunby, R.K.; et al. Multi-Decadal Changes in Tundra Environments and Ecosystems: Synthesis of the International Polar Year-Back to the Future Project (IPY-BTF). AMBIO 2011, 40, 705–716. [Google Scholar] [CrossRef] [Green Version]
- Gorchakovskiy, P.L.; Shiyatov, S.G. Phytoindication of Environmental Conditions and Natural Processes in High Mountain Regions; Nauka: Moscow, Russia, 1985. [Google Scholar]
- Hantemirov, R.M.; Shiyatov, S.G. The main stages of the development of woody vegetation in Yamal in the Holocene. Russ. J. Ecol. 1999, 30, 141–147. [Google Scholar]
- Mazepa, V.S. Stand density in the last millennium at the upper tree-line ecotone in the Polar Ural Mountains. Can. J. For. Res. 2005, 35, 2082–2091. [Google Scholar] [CrossRef]
- Kullman, L.; Kjallgren, L. Holocene pine tree-line evolution in the Swedish Scandes: Recent tree-line rise and climate in a long-term perspective. Boreas 2006, 35, 159–168. [Google Scholar] [CrossRef]
- MacDonald, G.M.; Kremenetski, K.V.; Beilman, D.W. Climate change and the northern Russian treeline zone. Philos. Trans. R. Soc. Biol. Sci. 2007, 363, 2285–2299. [Google Scholar] [CrossRef] [Green Version]
- Hansson, A.; Dargusch, P.; Shulmeister, J. A review of modern treeline migration, the factors controlling it and the implications for carbon storage. J. Mt. Sci. 2021, 18, 291–306. [Google Scholar] [CrossRef]
- Lu, X.; Liang, E.; Wang, Y.; Babst, F.; Camarero, J.J. Mountain treelines climb slowly despite rapid climate warming. Global Ecol. Biogeogr. 2020, 30, 305–315. [Google Scholar] [CrossRef]
- Payette, S.; Filion, L. White spruce expansion at the tree line and recent climatic change. Can. J. For. Res. 1985, 15, 241–251. [Google Scholar] [CrossRef]
- Holtmeier, F.-K. Mountain Timberlines—Ecology, Patchiness, and Dynamic; Kluwer: Dordrecht, The Netherlands, 2003. [Google Scholar]
- Kullman, L. Neoglacial climate control of subarctic Picea abies stand dynamics and range limit in Northern Sweden. Arct. Alp. Res. 1997, 29, 315–326. [Google Scholar] [CrossRef]
- Kullman, L. Wind-conditioned 20th century decline of birch treeline vegetation in the Swedish Scandes. Arctic 2005, 58, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, A.H. Ecological histories from Alaskan treelines provide insight into future change. Ecology 2005, 86, 1687–1695. [Google Scholar] [CrossRef] [Green Version]
- Aune, S.; Hofgaard, A.; Söderström, L. Contrasting climate- and land-use-driven tree encroachment patterns of subarctic tundra in northern Norway and the Kola Peninsula. Can. J. For. Res. 2011, 41, 437–449. [Google Scholar] [CrossRef]
- Hagedorn, F.; Shiyatov, S.G.; Mazepa, V.S.; Devi, N.M.; Grigor’ev, A.A.; Bartysh, A.A.; Fomin, V.V.; Kapralov, D.S.; Terent’ev, M.; Bugman, H.; et al. Treeline advances along the Urals mountain range—Driven by improved winter conditions? Global Change Biol. 2014, 20, 3530–3543. [Google Scholar] [CrossRef] [PubMed]
- Devi, N.M.; Kukarskih, V.V.; Galimova, A.A.; Mazepa, V.S.; Grigoriev, A.A. Climate change evidence in tree growth and stand productivity at the upper treeline ecotone in the Polar Ural Mountains. For. Ecosyst. 2020, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Shiyatov, S.G.; Terent’ev, M.M.; Fomin, V.V.; Zimmermann, N.E. Altitudinal and horizontal shifts of the upper boundaries of open and closed forests in the Polar Urals in the 20th century. Russ. J. Ecol. 2007, 38, 223–227. [Google Scholar] [CrossRef]
- Shiyatov, S.G. Dynamics of Woody and Shrubby Vegetation in the Mountains of the Polar Urals under the Influence of Modern Climate Changes; Yekaterinburg Publish: Yekaterinburg, Russia, 2009. [Google Scholar]
- Shiyatov, S.G.; Mazepa, V.S. The modern expansion of Siberian larch in to the mountain tundra of the Polar Urals. Russ. J. Ecol. 2015, 6, 403–410. [Google Scholar]
- Kirdyanov, A.V.; Hagedorn, F.; Knorre, A.A.; Fedotova, E.V.; Vaganov, E.A.; Naurzbaev, M.M.; Moiseev, P.A.; Rigling, A. 20th century tree-line advance and vegetation changes along an altitudinal transect in the Putorana Mountains, northern Siberia. Boreas 2012, 41, 56–67. [Google Scholar] [CrossRef]
- Im, S.T.; Kharuk, V.I. Climatically induced changes in the ecotone of the alpine forest-tundra of the Putorana plateau. Earth Res. Space 2013, 5, 32–44. [Google Scholar]
- Kharuk, V.I.; Ranson, K.J.; Im, S.I.; Oskorbin, P.A.; Dvinskaya, M.L.; Ovchinnikov, D.V. Tree-line structure and dynamics at the northern limit of the larch forest: Anabar Plateau, Siberia, Russia. Arct. Ant. Alp. Res. 2013, 4, 526–537. [Google Scholar] [CrossRef] [Green Version]
- Moiseev, P.A.; Galimova, A.A.; Bubnov, M.O.; Devi, N.M.; Fomin, V.V. The dynamics of stands and their productivity at the upper limit of growth in the Khibiny against the background of modern climate changes. Russ. J. Ecol. 2019, 5, 341–355. [Google Scholar]
- Grigoriev, A.A.; Devi, N.M.; Kukarskikh, V.V.; V’yukhin, V.V.; Galimova, A.A.; Moiseev, P.A.; Fomin, V.V. Structure and dynamics of stands of the upper border of the forest in the western part of the Putorana plateau. Russ. J. Ecol. 2019, 4, 243–254. [Google Scholar]
- Lukyanova, L.M.; Lokteva, T.N.; Bulycheva, T.M. Gas Exchange and the Pigment System of Plants in the Kola Subarctic (Khibiny Massif); KFAN USSR: Apatity, Russia, 1986. [Google Scholar]
- Parmuzin, Y.P. Modern relief-forming processes and the genesis of lake basins. In Putoranskaya Lake Province; Nauka: Novosibirsk, Russia, 1975; pp. 64–97. [Google Scholar]
- Atlas SSSR (Atlas of the Soviet Union); Tochenov, V.V.; Belyaeva, L.I. (Eds.) GUGK: Moscow, Russia, 1983. [Google Scholar]
- Holmes, R.L. Dendrochronological Program Library (Computer Program); The University of Arizona, Laboratory of Tree Ring Research: Tucson, Arizona, 1995. [Google Scholar]
- Bogdanova, E.G.; Golubev, V.S.; Ilyin, B.M.; Dragomilova, I.V. A new model for correcting measured precipitation and its application in the polar regions of the Russian Federation. Meteorol. Hydrol. 2002, 10, 68–93. [Google Scholar]
- Bogdanova, E.G.; Gavrilova, S.Y. Elimination of the inhomogeneity of precipitation time series caused by the replacement of the rain gauge with Nifer protection by the Tretyakov rain gauge. Meteorol. Hydrol. 2008, 8, 87–102. [Google Scholar]
- Koshkina, N.B.; Moiseev, P.A.; Goryaeva, A.V. Reproduction of the Siberian spruce in the timberline ecotone of the Iremel’ Massif. Russ. J. Ecol. 2008, 39, 83–91. [Google Scholar] [CrossRef]
- Grigoriev, A.A.; Moiseev, P.A.; Nagimov, Z.Y. Dynamics of the upper limit of woody vegetation in the highlands of the Subpolar Urals under the influence of modern climate change. Ecology 2013, 4, 284–295. [Google Scholar]
- Natural Conditions of the Khibiny Mountain Range; Myagkov, S.M. (Ed.) Moscow State University: Moscow, Russia, 1986. [Google Scholar]
- Holtmeier, F.K.; Broll, G. Wind as an Ecological Agent at Treelines in North America, the Alps, and the European Subarctic. Phys. Geogr. 2010, 31, 203–233. [Google Scholar] [CrossRef]
- Shrestha, K.B.; Hofgaard, A.; Vandvik, V. Recent treeline dynamics are similar between dry and mesic areas of Nepal, central Himalaya. J. Plant Ecol. 2015, 4, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sturm, M.; Schimel, J.; Michaelson, G.; Welker, J.M.; Oberbauer, S.F.; Liston, G.E.; Fahnestock, J.; Romanovsky, V.E. Winter biological processes could help convert arctic tundra to shrubland. BioScience 2005, 55, 17–26. [Google Scholar] [CrossRef]
- Elliott, G.P.; Cowell, C.M. Slope aspect mediates fine-scale tree establishment patterns at upper treeline during wet and dry periods of the 20th century. Arct. Antarct. Alp. Res. 2015, 47, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Elliott, G.P.; Kipfmueller, K.F. Multi-scale Influences of Slope Aspect and Spatial Pattern on Ecotonal Dynamics at Upper Treeline in the Southern Rocky Mountains, U.S.A. Arct. Antarct. Alp. Res. 2010, 42, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Ziaco, E.; Biondi, F.; Rossi, S.; Deslauriers, A. Climatic influences on wood anatomy and tree-ring Features of great basin conifers at a new mountain Observatory. Appl. Plant Sci. 2014, 2, 1400054. [Google Scholar] [CrossRef]
- Kuyek, N.J.; Thomas, S.C. Trees are larger on south slopes in late-seral conifer stands in northwestern British Columbia. Can. J. For. Res. 2019, 49, 1349–1356. [Google Scholar] [CrossRef] [Green Version]
- Danby, R.K.; Hik, D.S. Variability, contingency and rapid change in recent Subarctic alpine tree line dynamics. J. Ecol. 2007, 95, 352–363. [Google Scholar] [CrossRef]
- Reger, B.; Kölling, C.; Ewald, J. Modelling effective thermal climate for mountain forests in the Bavarian Alps: Which is the best model? J. Veg. Sci. 2011, 22, 677–687. [Google Scholar] [CrossRef]
- Demin, V.I. The main climatic trends on the Kola Peninsula for the period of instrumental meteorological measurements. Rep. Kola Sci. Cent. Russ. Acad. Sci. 2012, 3, 98–110. [Google Scholar]
- Holtmeier, F.-K.; Broll, G. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. Glob. Ecol. Biogeogr. 2005, 14, 395–410. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Ranson, K.J.; Im, S.T.; Vdovin, A.S. Spatial distribution and temporal dynamics of high-elevation forest stands in southern Siberia. Glob. Ecol. Biogeogr. 2010, 19, 822–830. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gutiérrez, E. Pace and pattern of recent treeline dynamics: Response of ecotones to climatic variability in the Spanish Pyrenees. Clim. Change 2004, 63, 181–200. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Dunca, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Grigoriev, A.A.; Shalaumova, Y.V.; Erokhina, O.V.; Sokovnina, S.Y.; Vatolina, E.I.; Wilmking, M. Expansion of Juniperus sibirica Burgsd. as a response to climate change and associated effect on mountain tundra vegetation in the Northern Urals. J. Mt. Sci. 2020, 17, 2339–2353. [Google Scholar] [CrossRef]
- Frost, G.V.; Epstein, H.E.; Walker, D.A.; Matyshak, G.; Ermohina, K. Seasonal and long-term changes to active-layer temperatures after tall shrubland expansion and succession in arctic tundra. Ecosystems 2018, 21, 507–520. [Google Scholar] [CrossRef]
- Sveinbjöornsson, B.; Kauhanen, H.; Nordell, O. Treeline ecology of mountain birch in the Tornetrask area. Ecol. Bull. 1996, 46, 65–70. [Google Scholar]
- Groffman, P.M.; Driscoll, C.T.; Fahey, T.J.; Hardy, J.P.; Fitzhugh, R.D.; Tierney, G.L. Colder soils in a warmer world: A snow manipulation study in a northern hardwood forest ecosystem. Biogeochemistry 2001, 56, 135–150. [Google Scholar] [CrossRef]
- Mamet, S.D.; Brown, C.D.; Trant, A.J.; Laroque, C.P. Shifting global Larix distributions: Northern expansion and southern retraction as species respond to changing climate. J. Biogeogr. 2019, 46, 30–44. [Google Scholar] [CrossRef] [Green Version]
- Moiseev, P.A.; Van der Meer, M.; Rigling, A.; Shevchenko, I.G. Effect of Climatic Changes on the Formation of Siberian Spruce Generations in Subglotsy Tree Stands of the Southern Urals. Russ. J. Ecol. 2004, 35, 135–143. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Khibiny Massif | Putorana Plateau |
|---|---|---|
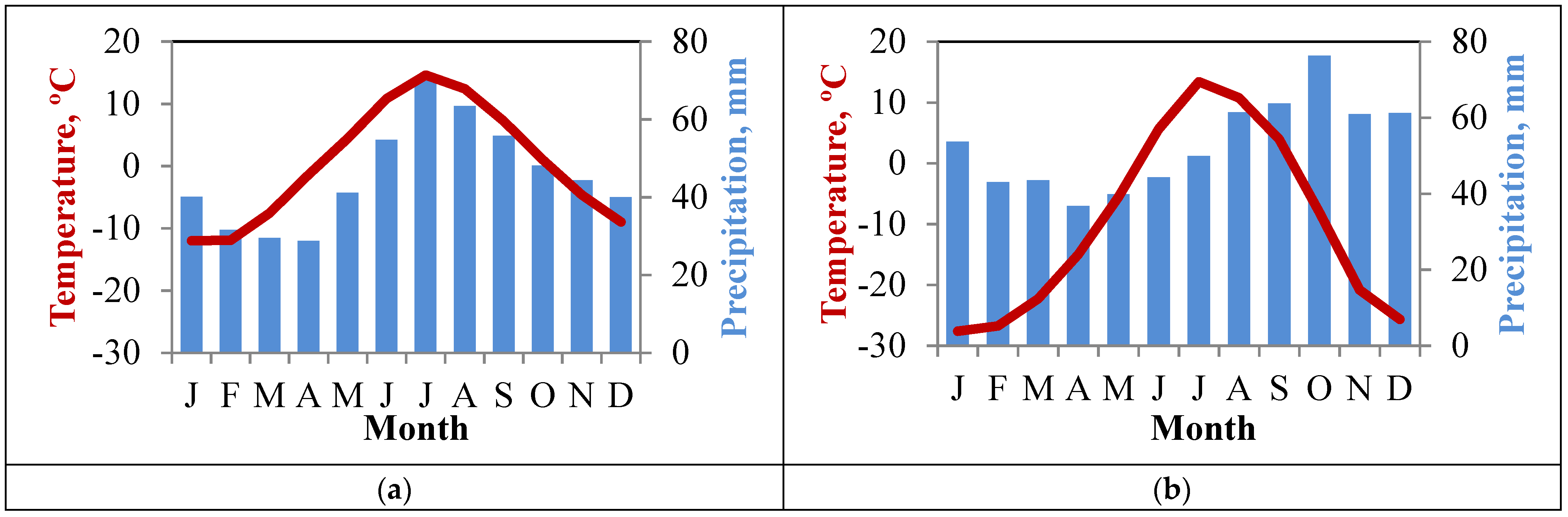
| Mean June air temperature, °C 1 | 10.2 ± 5.0 | 7.0 ± 2.4 |
| Average summer sunshine duration (h) | 659 ± 131 | 786 ± 136 |
| Mean January air temperature (°C) | −12.6 ± 3.9 | −27.6 ± 5.0 |
| Annual precipitation (mm) | 928 ± 185 | 635 ± 131 |
| Duration of the growing season (days) | 104 | 73 |
| Variables | Khibiny Massif | Putorana Plateau | ||
|---|---|---|---|---|
| Imandra | Maly Vudayvr | Kitchepakh | Sukhiye Gory Massif | |
| Latitude N | 67.83 | 67.67 | 67.59 | 69.40 |
| Longitude E | 33.34 | 33.58 | 34.07 | 90.75 |
| Altitudinal range of the treeline ecotone (m a.s.l.) | 345–420 | 442–667 | 264–448 | 225–730 |
| Transect exposition | S, N | NE, E, SE, SW | E, SE, S | N, E, S, W |
| Study tree species 1 | BP, PS | BP | BP, PO | LG |
| No. sampled cores | 1176 | 1412 | 2241 | 1963 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grigoriev, A.A.; Shalaumova, Y.V.; Vyukhin, S.O.; Balakin, D.S.; Kukarskikh, V.V.; Vyukhina, A.A.; Camarero, J.J.; Moiseev, P.A. Upward Treeline Shifts in Two Regions of Subarctic Russia Are Governed by Summer Thermal and Winter Snow Conditions. Forests 2022, 13, 174. https://doi.org/10.3390/f13020174
Grigoriev AA, Shalaumova YV, Vyukhin SO, Balakin DS, Kukarskikh VV, Vyukhina AA, Camarero JJ, Moiseev PA. Upward Treeline Shifts in Two Regions of Subarctic Russia Are Governed by Summer Thermal and Winter Snow Conditions. Forests. 2022; 13(2):174. https://doi.org/10.3390/f13020174
Chicago/Turabian StyleGrigoriev, Andrey A., Yulia V. Shalaumova, Sergey O. Vyukhin, Dmitriy S. Balakin, Vladimir V. Kukarskikh, Arina A. Vyukhina, Jesús Julio Camarero, and Pavel A. Moiseev. 2022. "Upward Treeline Shifts in Two Regions of Subarctic Russia Are Governed by Summer Thermal and Winter Snow Conditions" Forests 13, no. 2: 174. https://doi.org/10.3390/f13020174