
Higher Soil Aggregate Stability in Subtropical Coniferous Plantations Than Natural Forests Due to Microbial and Aggregate Factors
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
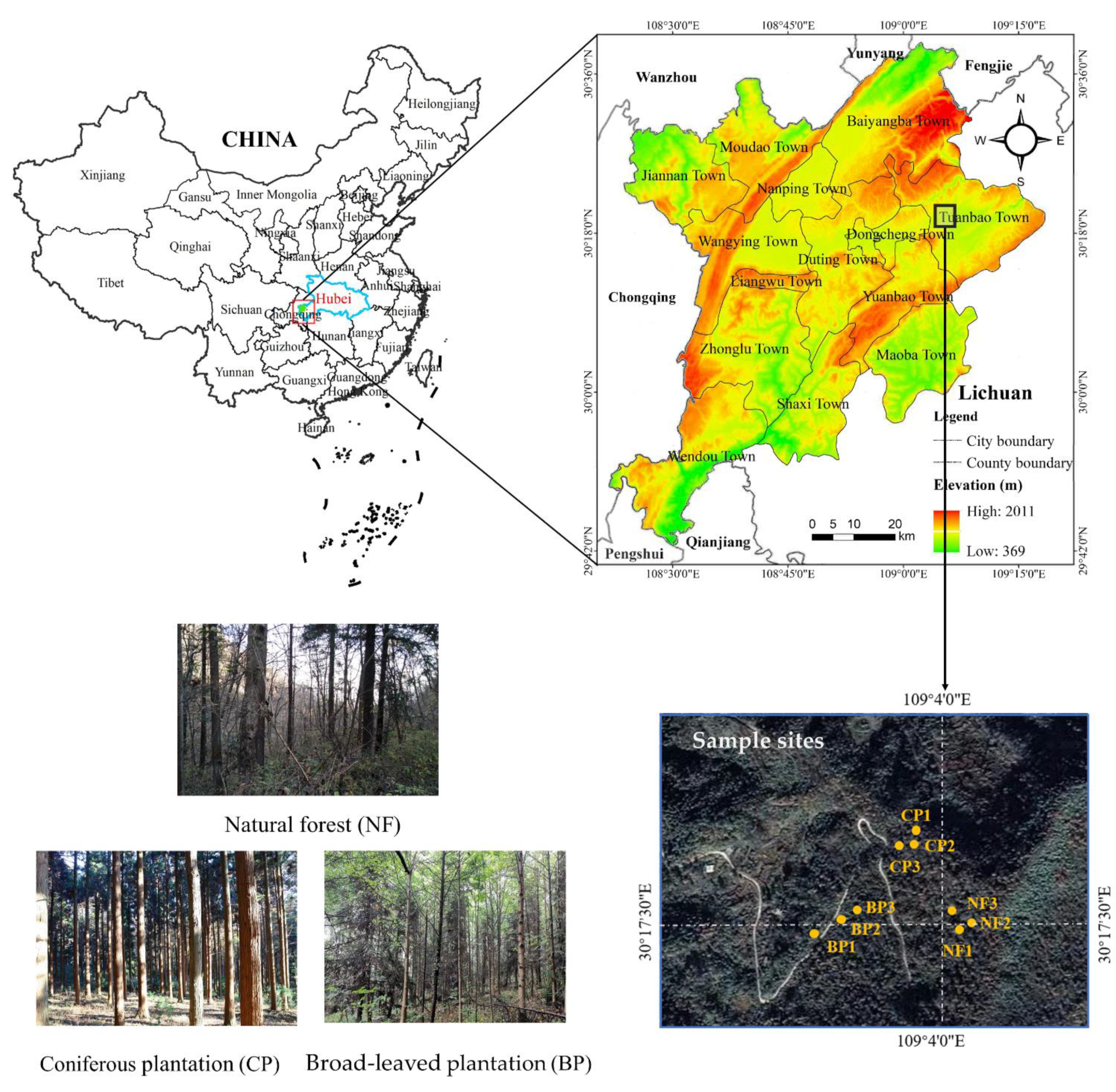
2.1. Study Site
2.2. Experimental Design and Sampling
2.3. Measurements
2.3.1. Soil Physicochemical Properties
2.3.2. Soil Microbial Analysis
2.3.3. Classification of Soil Aggregate and Possible Influencing Factors
2.4. Statistical Analysis
3. Results
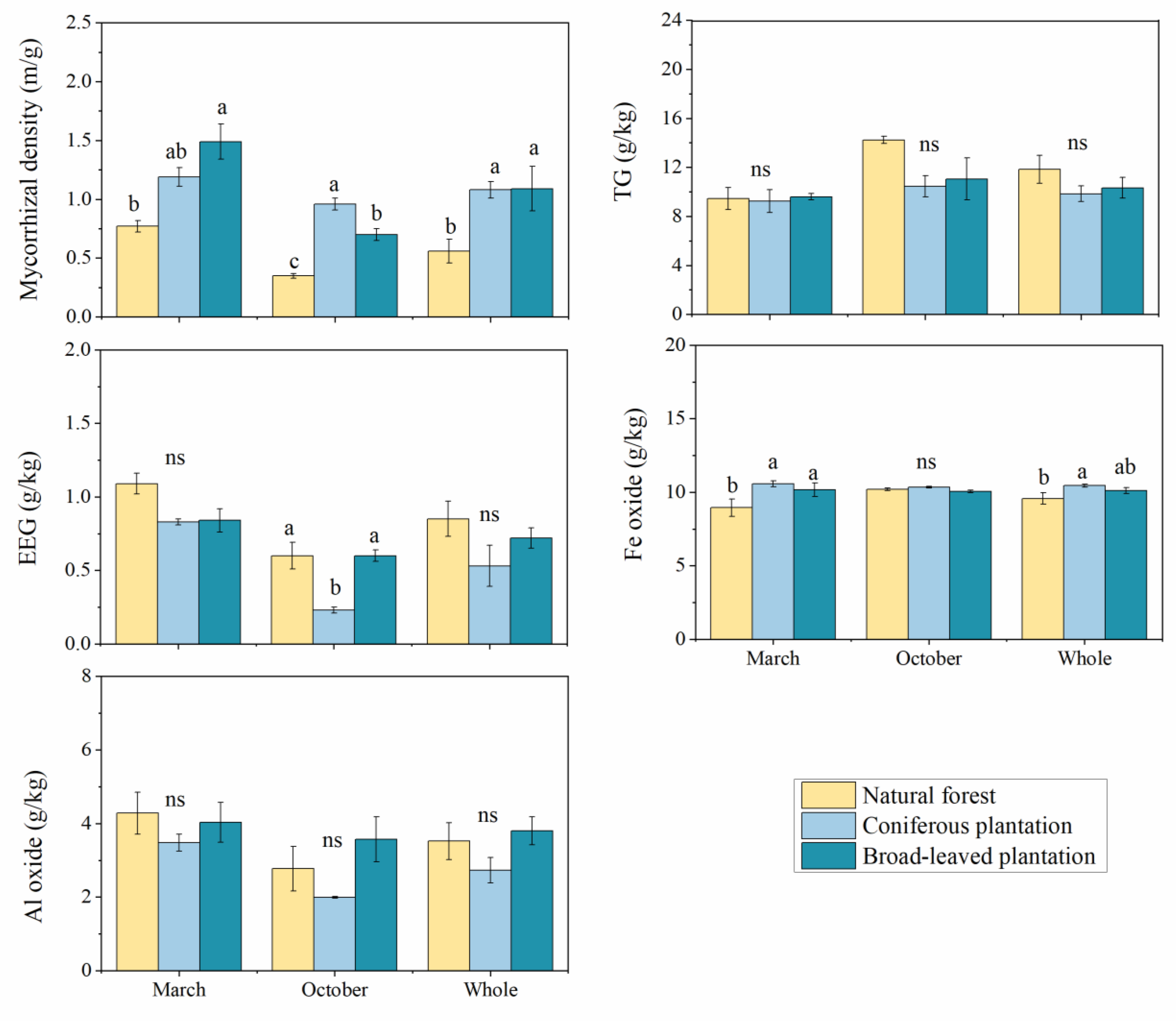
3.1. Soil Physicochemical Properties
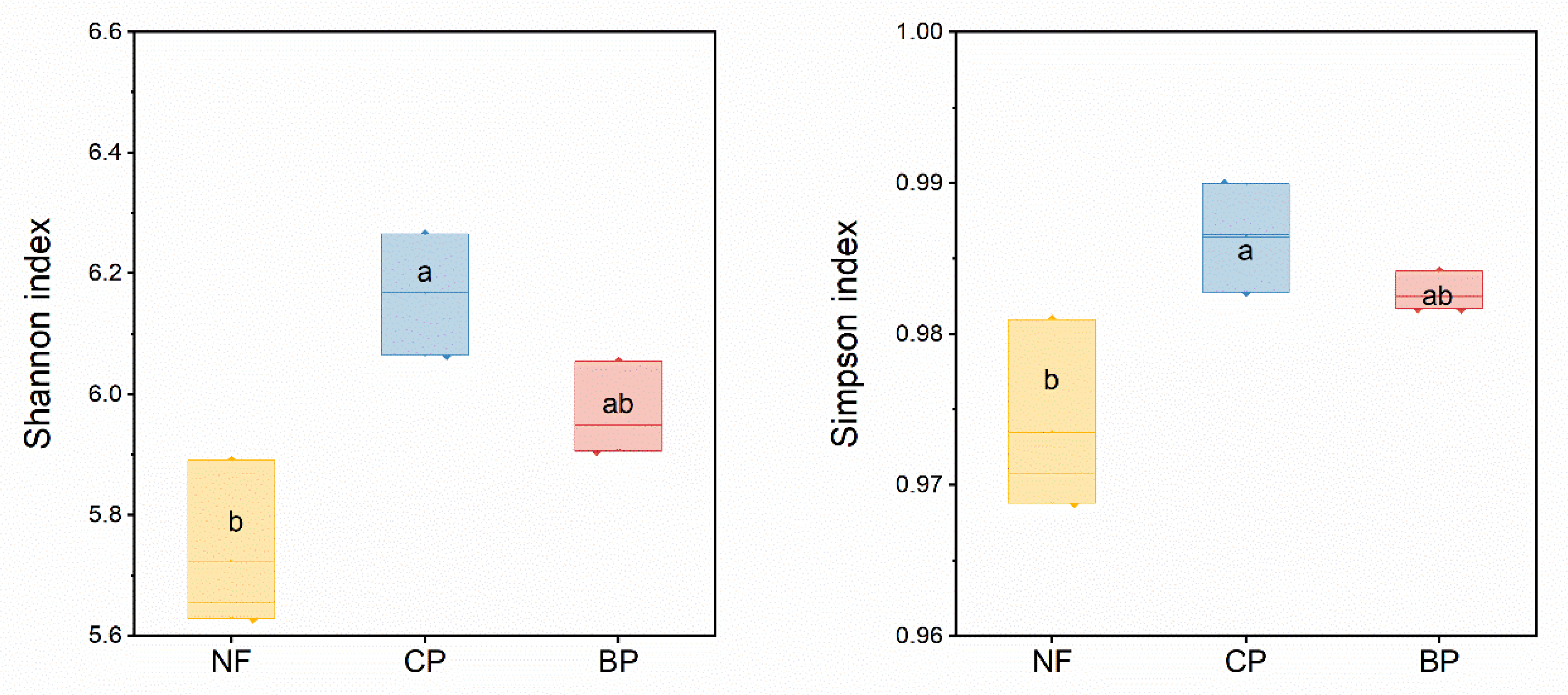
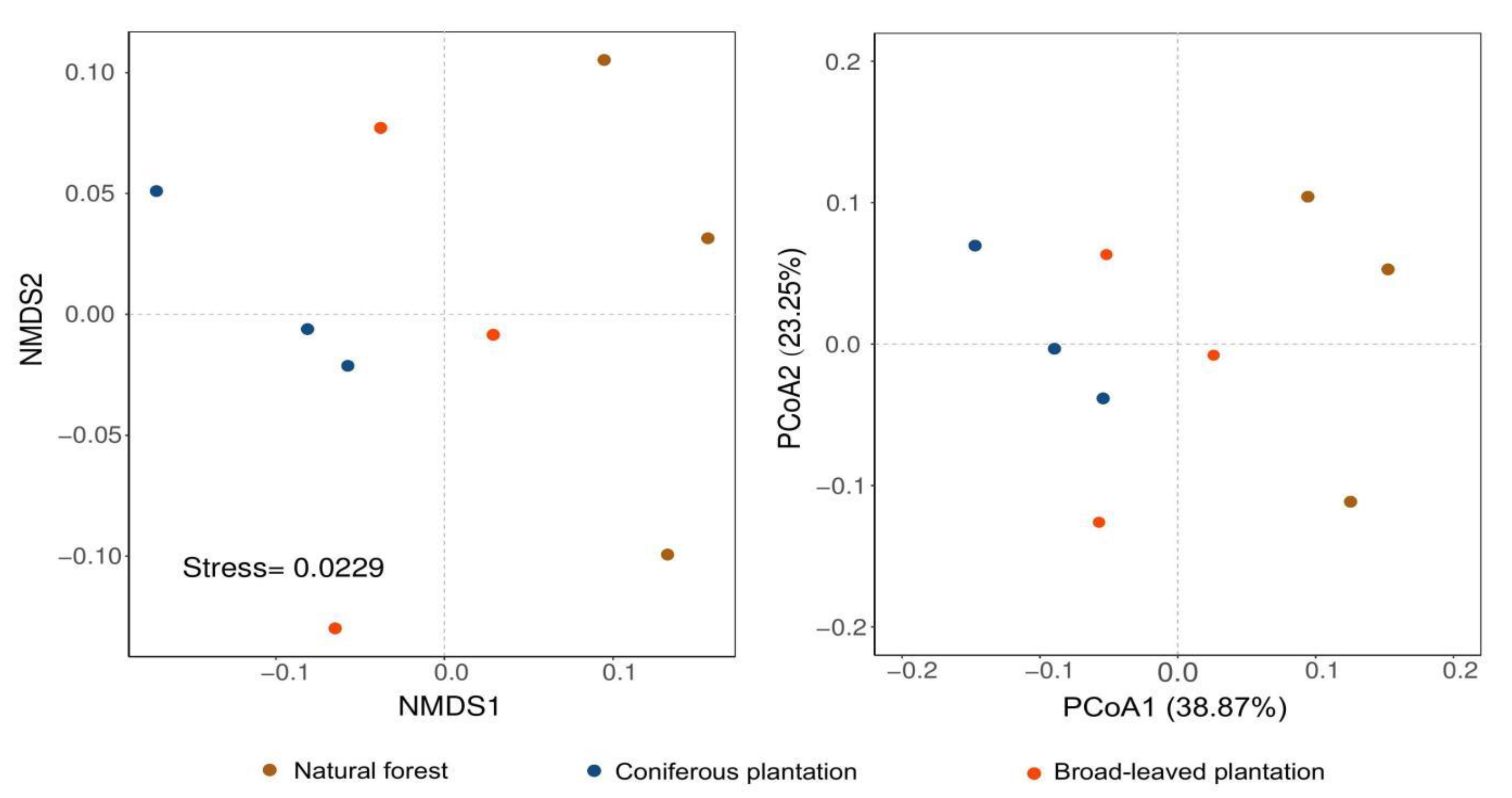
3.2. Composition and Diversity of Soil Microbial Communities
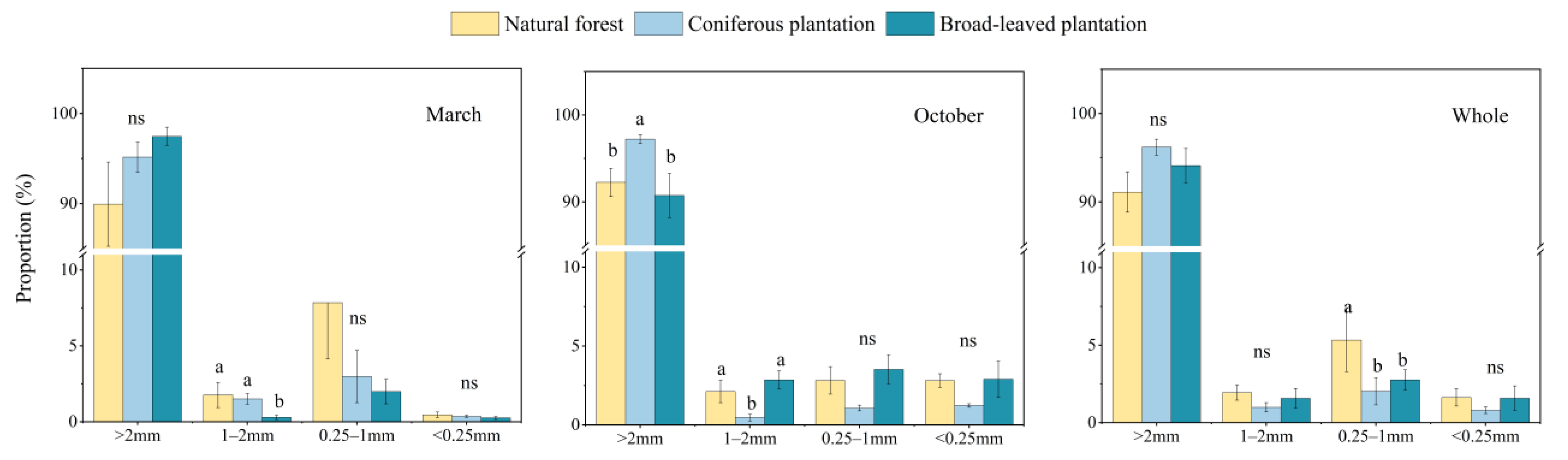
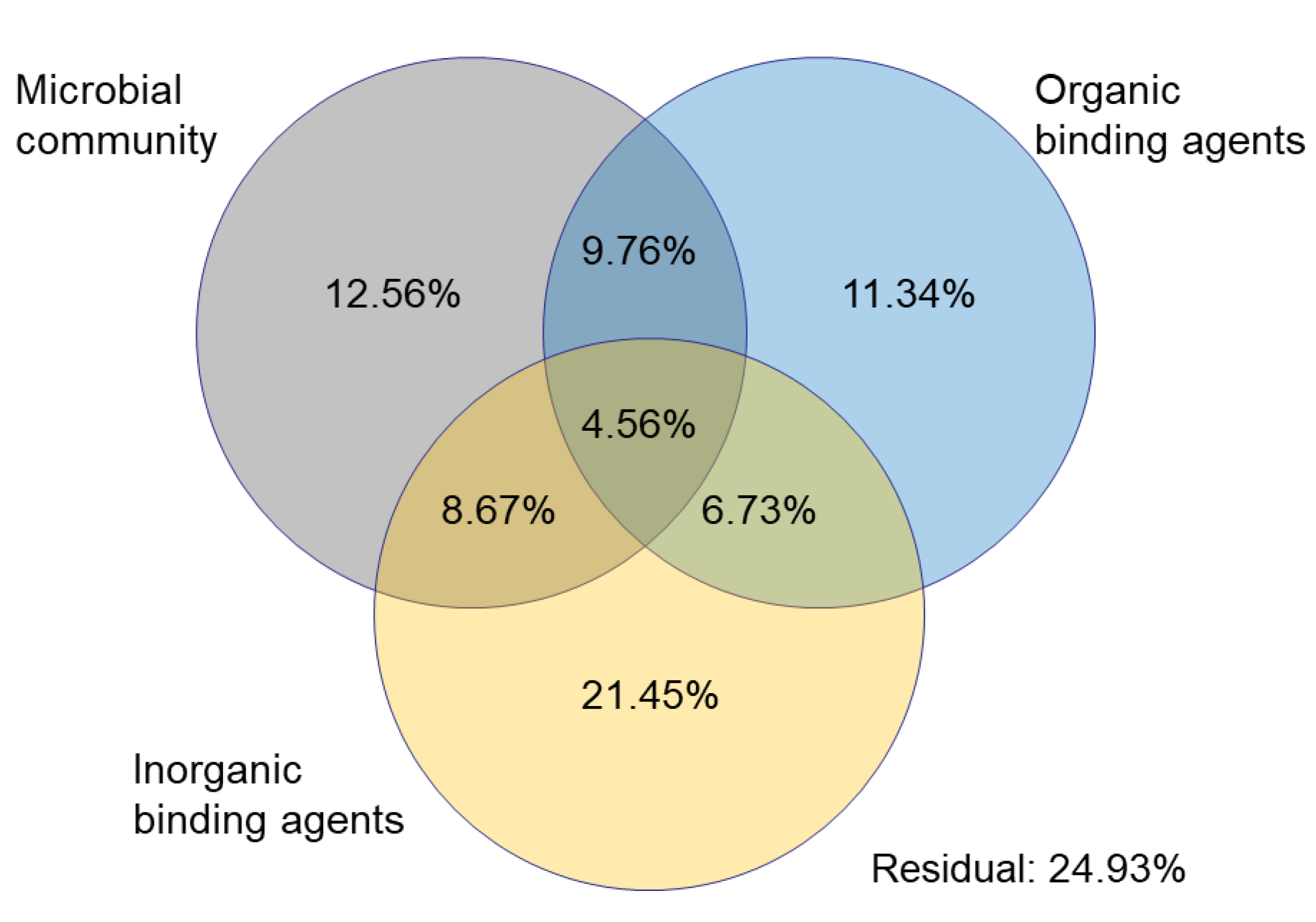
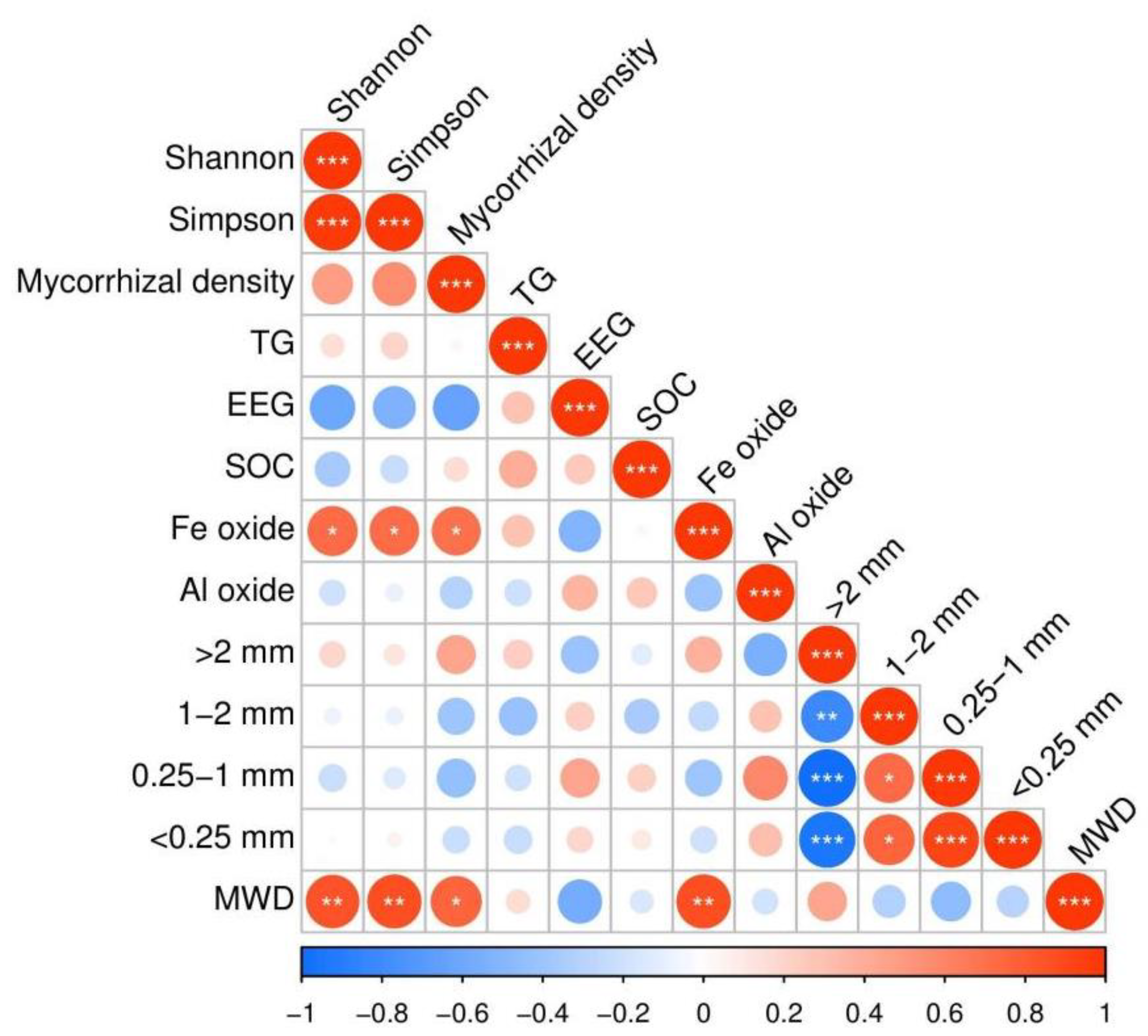
3.3. Soil Aggregate Properties and Influencing Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Acharya, R.P.; Maraseni, T.; Cockfield, G. Global trend of forest ecosystem services valuation—An analysis of publications. Ecosyst. Serv. 2019, 39, 100979. [Google Scholar] [CrossRef]
- Global Forest Resources Assessment. 2020. Available online: https://www.fao.org/forest-resources-assessment/2020/en (accessed on 1 November 2021).
- Camarero, J.J.; Gazol, A.; Linares, J.C.; Fajardo, A.; Colangelo, M.; Valeriano, C.; Sánchez-Salguero, R.; Sangüesa-Barreda, G.; Granda, E.; Gimeno, T.E. Differences in temperature sensitivity and drought recovery between natural stands and plantations of conifers are species-specific. Sci. Total Environ. 2021, 796, 148930. [Google Scholar] [CrossRef] [PubMed]
- Mansourian, S.; Parrotta, J. From addressing symptoms to tackling the illness: Reversing forest loss and degradation. Environ. Sci. Policy 2019, 101, 262–265. [Google Scholar] [CrossRef]
- Getaneh, S.; Honnay, O.; Desie, E.; Helsen, K.; Couck, L.; Shibru, S.; Muys, B. Impact of tree litter identity, litter diversity and habitat quality on litter decomposition rates in tropical moist evergreen forest. For. Ecosyst. 2022, 9, 100023. [Google Scholar] [CrossRef]
- Romanelli, J.P.; Meli, P.; Santos, J.P.B.; Jacob, I.N.; Souza, L.R.; Rodrigues, A.V.; Trevisan, D.P.; Huang, C.; Almeida, D.R.A.; Silva, L.G.M.; et al. Biodiversity responses to restoration across the Brazilian Atlantic Forest. Sci. Total Environ. 2022, 821, 153403. [Google Scholar] [CrossRef]
- Yang, Y.; Dou, Y.; Cheng, H.; An, S. Plant functional diversity drives carbon storage following vegetation restoration in Loess Plateau, China. J. Environ. Manag. 2019, 246, 668–678. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Mentler, A.; Mayer, H.; Blum, W.E.H. Soil aggregation, aggregate stability, organic carbon and nitrogen in different soil aggregate fractions under forest and shrub vegetation on the Loess Plateau, China. Catena 2010, 81, 226–233. [Google Scholar] [CrossRef]
- Le Bissonnais, Y. Aggregate stability and assessment of soil crustability and erodibility: I. Theory and methodology. Eur. J. Soil Sci. 2016, 67, 11–21. [Google Scholar] [CrossRef]
- Sanji, R.; Kooch, Y.; Rey, A. Impact of forest degradation and reforestation with Alnus and Quercus species on soil quality and function in northern Iran. Ecol. Indic. 2020, 112, 106132. [Google Scholar] [CrossRef]
- Caravaca, F.; Lax, A.; Albaladejo, J. Aggregate stability and carbon characteristics of particle-size fractions in cultivated and forested soils of semiarid Spain. Soil Tillage Res. 2004, 78, 83–90. [Google Scholar] [CrossRef]
- Gupta, V.V.S.R.; Germida, J.J. Soil aggregation: Influence on microbial biomass and implications for biological processes. Soil Biol. Biochem. 2015, 80, A3–A9. [Google Scholar] [CrossRef]
- Liu, T.; Jiang, K.; Tan, Z.; He, Q.; Zhang, H.; Wang, C. A Method for Performing Reforestation to Effectively Recover Soil Water Content in Extremely Degraded Tropical Rain Forests. Front. Ecol. Evol. 2021, 9, 643994. [Google Scholar] [CrossRef]
- Koryś, K.A.; Latawiec, A.E.; Mendes, M.S.; Sansevero, J.B.B.; Rodrigues, A.F.; Iribarrem, A.S.; Dib, V.; Jakovac, C.C.; Allek, A.; Pena, I.A.B.; et al. Early Response of Soil Properties under Different Restoration Strategies in Tropical Hotspot. Land 2021, 10, 768. [Google Scholar] [CrossRef]
- Parhizkar, M.; Shabanpour, M.; Lucas-Borja, M.E.; Zema, D.A.; Li, S.; Tanaka, N.; Cerdà, A. Effects of length and application rate of rice straw mulch on surface runoff and soil loss under laboratory simulated rainfall. Int. J. Sediment Res. 2021, 36, 468–478. [Google Scholar] [CrossRef]
- Huang, B.; Yuan, Z.; Zheng, M.; Liao, Y.; Nguyen, K.L.; Nguyen, T.H.; Sombatpanit, S.; Li, D. Soil and Water Conservation Techniques in Tropical and Subtropical Asia: A Review. Sustainability 2022, 14, 5035. [Google Scholar] [CrossRef]
- Zhao, R.; He, M.; Jiang, C.; Li, C.; Liu, F. Microbial community structure in rhizosphere soil rather than that in bulk soil characterizes aggregate-associated organic carbon under long-term forest conversion in subtropical region. Rhizosphere 2021, 20, 100438. [Google Scholar] [CrossRef]
- Collis-george, N.; Lal, R. Infiltration into columns of swelling soil as studied by high speed photography. Soil Res. 1970, 8, 195–207. [Google Scholar] [CrossRef]
- Kimura, S.D.; Melling, L.; Goh, K.J. Influence of soil aggregate size on greenhouse gas emission and uptake rate from tropical peat soil in forest and different oil palm development years. Geoderma 2012, 185, 1–5. [Google Scholar] [CrossRef]
- Peng, J.; Wu, X.; Ni, S.; Wang, J.; Song, Y.; Cai, C. Investigating intra-aggregate microstructure characteristics and influencing factors of six soil types along a climatic gradient. Catena 2022, 210, 105867. [Google Scholar] [CrossRef]
- Wilson, G.W.T.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef]
- Peng, X.; Yan, X.; Zhou, H.; Zhang, Y.Z.; Sun, H. Assessing the contributions of sesquioxides and soil organic matter to aggregation in an Ultisol under long-term fertilization. Soil Tillage Res. 2015, 146, 89–98. [Google Scholar] [CrossRef]
- Wu, Q.S.; He, X.H.; Zou, Y.N.; He, K.P.; Sun, Y.H.; Cao, M.Q. Spatial distribution of glomalin-related soil protein and its relationships with root mycorrhization, soil aggregates, carbohydrates, activity of protease and β-glucosidase in the rhizosphere of Citrus unshiu. Soil Biol. Biochem. 2012, 45, 181–183. [Google Scholar] [CrossRef]
- Xiao, L.; Zhang, Y.; Li, P.; Xu, G.; Shi, P.; Zhang, Y. Effects of freeze-thaw cycles on aggregate-associated organic carbon and glomalin-related soil protein in natural-succession grassland and Chinese pine forest on the Loess Plateau. Geoderma 2019, 334, 1–8. [Google Scholar] [CrossRef]
- Jeewani, P.H.; Luo, Y.; Yu, G.; Fu, Y.; He, X.; Van Zwieten, L.; Liang, C.; Kumar, A.; He, Y.; Kuzyakov, Y.; et al. Arbuscular mycorrhizal fungi and goethite promote carbon sequestration via hyphal-aggregate mineral interactions. Soil Biol. Biochem. 2021, 162, 108417. [Google Scholar] [CrossRef]
- Bedini, S.; Pellegrino, E.; Avio, L.; Pellegrini, S.; Bazzoffi, P.; Argese, E.; Giovannetti, M. Changes in soil aggregation and glomalin-related soil protein content as affected by the arbuscular mycorrhizal fungal species Glomus mosseae and Glomus intraradices. Soil Biol. Biochem. 2009, 41, 1491–1496. [Google Scholar] [CrossRef]
- Martin, S.L.; Mooney, S.J.; Dickinson, M.J.; West, H.M. The effects of simultaneous root colonisation by three Glomus species on soil pore characteristics. Soil Biol. Biochem. 2012, 49, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Lu, H.; Chen, J.; Jiang, Y.; Williams, M.A.; Wu, S.; Li, J.; Liu, J.; Yang, G.; Yan, C. Interactions of soil metals with glomalin-related soil protein as soil pollution bioindicators in mangrove wetland ecosystems. Sci. Total Environ. 2020, 709, 136051. [Google Scholar] [CrossRef]
- Staunton, S.; Saby, N.P.A.; Arrouays, D.; Quiquampoix, H. Can soil properties and land use explain glomalin-related soil protein (GRSP) accumulation? A nationwide survey in France. Catena 2020, 193, 104620. [Google Scholar] [CrossRef]
- Zhu, R.; Zheng, Z.; Li, T.; He, S.; Zhang, X.; Wang, Y.; Liu, T. Effect of tea plantation age on the distribution of glomalin-related soil protein in soil water-stable aggregates in southwestern China. Environ. Sci. Pollut. Res. 2019, 26, 1973–1982. [Google Scholar] [CrossRef]
- Huang, J.; Kogbara, R.B.; Hariharan, N.; Masad, E.A.; Little, D.N. A state-of-the-art review of polymers used in soil stabilization. Constr. Build. Mater. 2021, 305, 124685. [Google Scholar] [CrossRef]
- Xue, B.; Huang, L.; Huang, Y.; Zhou, F.; Li, F.; Kubar, K.A.; Li, X.; Lu, J.; Zhu, J. Roles of soil organic carbon and iron oxides on aggregate formation and stability in two paddy soils. Soil Tillage Res. 2019, 187, 161–171. [Google Scholar] [CrossRef]
- Yang, C.; Liu, N.; Zhang, Y. Soil aggregates regulate the impact of soil bacterial and fungal communities on soil respiration. Geoderma 2019, 337, 444–452. [Google Scholar] [CrossRef]
- Ngo Bieng, M.A.; Finegan, B.; Sist, P. Active restoration of secondary and degraded forests in the context of the UN Decade on Ecosystem Restoration. For. Ecol. Manag. 2022, 503, 119770. [Google Scholar] [CrossRef]
- Zhang, Y.; E, S.; Wang, Y.; Su, S.; Bai, L.; Wu, C.; Zeng, X. Long-term manure application enhances the stability of aggregates and aggregate-associated carbon by regulating soil physicochemical characteristics. Catena 2021, 203, 105342. [Google Scholar] [CrossRef]
- Carugati, L.; Gatto, B.; Rastelli, E.; Lo Martire, M.; Coral, C.; Greco, S.; Danovaro, R. Impact of mangrove forests degradation on biodiversity and ecosystem functioning. Sci. Rep. 2018, 8, 13298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyvindson, K.; Repo, A.; Mönkkönen, M. Mitigating forest biodiversity and ecosystem service losses in the era of bio-based economy. For. Policy. Econ. 2018, 92, 119–127. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, P. Soil bacterial community varies but fungal community stabilizes along five vertical climate zones. Catena 2020, 195, 104841. [Google Scholar] [CrossRef]
- Wang, Q.; Han, Y.; Lan, S.; Hu, C. Metagenomic Insight into Patterns and Mechanism of Nitrogen Cycle During Biocrust Succession. Front. Microbiol. 2021, 12, 633428. [Google Scholar] [CrossRef]
- Yang, Y.J.; Liu, S.R.; Wang, H.; Chen, L.; Lu, L.H.; Cai, D.X. Reduction in throughfall reduces soil aggregate stability in two subtropical plantations. Eur. J. Soil Sci. 2019, 70, 301–310. [Google Scholar] [CrossRef]
- Muleta, D.; Assefa, F.; Nemomissa, S.; Granhall, U. Composition of coffee shade tree species and density of indigenous arbuscular mycorrhizal fungi (AMF) spores in Bonga natural coffee forest, southwestern Ethiopia. For. Ecol. Manag. 2007, 241, 145–154. [Google Scholar] [CrossRef]
- Wright, S.F.; Upadhyaya, A. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of Arbuscular Mycorrhizal Fungi. Soil Sci. 1996, 161, 575–586. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, X.; Zhang, S.; Xing, Y.; Wang, R.; Liang, W. Organic amendment effects on aggregate-associated organic C, microbial biomass C and glomalin in agricultural soils. Catena 2014, 123, 188–194. [Google Scholar] [CrossRef]
- Bai, Y.; Zhou, Y.; He, H. Effects of rehabilitation through afforestation on soil aggregate stability and aggregate-associated carbon after forest fires in subtropical China. Geoderma 2020, 376, 114548. [Google Scholar] [CrossRef]
- Kooch, Y.; Rostayee, F.; Hosseini, S.M. Effects of tree species on topsoil properties and nitrogen cycling in natural forest and tree plantations of northern Iran. Catena 2016, 144, 65–73. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, W.; Entemake, W.; Wang, J.; Liu, H.; Zhao, Z.; Li, Y.; Qiao, L.; Yang, B.; Liu, G.; et al. Long-term vegetation restoration promotes the stability of the soil micro-food web in the Loess Plateau in North-west China. Catena 2021, 202, 105293. [Google Scholar] [CrossRef]
- Parhizkar, M.; Shabanpour, M.; Miralles, I.; Zema, D.A.; Lucas-Borja, M.E. Effects of plant species on soil quality in natural and planted areas of a forest park in northern Iran. Sci. Total Environ. 2021, 778, 146310. [Google Scholar] [CrossRef]
- Cai, H.; Li, F.; Jin, G. Fine root biomass, production and turnover rates in plantations versus natural forests: Effects of stand characteristics and soil properties. Plant Soil 2019, 436, 463–474. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, Y.; Brunel, C.; van Kleunen, M. Soil-microorganism-mediated invasional meltdown in plants. Nat. Ecol. Evol. 2020, 4, 1612–1621. [Google Scholar] [CrossRef]
- Wang, M.; Guo, X.; Zhang, S.; Xiao, L.; Mishra, U.; Yang, Y.; Zhu, B.; Wang, G.; Mao, X.; Qian, T.; et al. Global soil profiles indicate depth-dependent soil carbon losses under a warmer climate. Nat. Commun. 2022, 13, 5514. [Google Scholar] [CrossRef]
- Baumann, K.; Dignac, M.-F.; Rumpel, C.; Bardoux, G.; Sarr, A.; Steffens, M.; Maron, P.-A. Soil microbial diversity affects soil organic matter decomposition in a silty grassland soil. Biogeochemistry 2013, 114, 201–212. [Google Scholar] [CrossRef]
- Fokom, R.; Adamou, S.; Teugwa, M.C.; Begoude Boyogueno, A.D.; Nana, W.L.; Ngonkeu, M.E.L.; Tchameni, N.S.; Nwaga, D.; Tsala Ndzomo, G.; Amvam Zollo, P.H. Glomalin related soil protein, carbon, nitrogen and soil aggregate stability as affected by land use variation in the humid forest zone of south Cameroon. Soil Tillage Res. 2012, 120, 69–75. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef]
- Leifheit, E.F.; Veresoglou, S.D.; Lehmann, A.; Morris, E.K.; Rillig, M.C. Multiple factors influence the role of arbuscular mycorrhizal fungi in soil aggregation—A meta-analysis. Plant Soil 2014, 374, 523–537. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, S.; Hu, R.; Li, Y. Aggregate stability and size distribution of red soils under different land uses integrally regulated by soil organic matter, and iron and aluminum oxides. Soil Tillage Res. 2017, 167, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Wang, L.; Liang, C.; Xi, F.; Pei, Z.; Du, L. Soil aggregate stability and iron and aluminium oxide contents under different fertiliser treatments in a long-term solar greenhouse experiment. Pedosphere 2016, 26, 760–767. [Google Scholar] [CrossRef]
- Wang, J.; Liu, L.; Qiu, X.; Wei, Y.; Li, Y.; Shi, Z. Contents of soil organic carbon and nitrogen in water-stable aggregates in abandoned agricultural lands in an arid ecosystem of Northwest China. J. Arid Land 2016, 8, 350–363. [Google Scholar] [CrossRef] [Green Version]
- Swanepoel, P.A.; Habig, J.; du Preez, C.C.; Botha, P.R.; Snyman, H.A. Biological quality of a podzolic soil after 19 years of irrigated minimum-till kikuyuryegrass pasture. Soil Res. 2014, 52, 64–75. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest | Forest Type | Longitude (E) | Latitude (N) | Elevation (m) | Slope Aspect | Stand Age |
|---|---|---|---|---|---|---|
| NF1 | Natural forest | 109°4′3.41″ | 30°17′30.31″ | 1485 | 19° E | Mature forest |
| NF2 | Natural forest | 109°4′4.03″ | 30°17′31.44″ | 1482 | 18° NE | Mature forest |
| NF3 | Natural forest | 109°4′2.33″ | 30°17′32.29″ | 1468 | 23° NE | Mature forest |
| CP1 | Coniferous plantation | 109°3′53.48″ | 30°17′44.08″ | 1382 | 17° N | Near-mature forest |
| CP2 | Coniferous plantation | 109°3′54.15″ | 30°17′43.75″ | 1394 | 16° N | Near-mature forest |
| CP3 | Coniferous plantation | 109°3′54.78″ | 30°17′43.35″ | 1393 | 20° NE | Near-mature forest |
| BP1 | Broad-leaved plantation | 109°3′42.83″ | 30°17′27.07″ | 1560 | 10° NE | Near-mature forest |
| BP2 | Broad-leaved plantation | 109°3′51.12″ | 30°17′29.82″ | 1534 | 17° NW | Near-mature forest |
| BP3 | Broad-leaved plantation | 109°3′52.56″ | 30°17′30.55″ | 1541 | 10° N | Near-mature forest |
| Property | SM (%) | ST (°C) | pH | BD (g/cm3) | SOC (g/kg) | TN (g/kg) | C/N | TP (g/kg) | AP (mg/kg) | |
|---|---|---|---|---|---|---|---|---|---|---|
| March | NF | 41.96 ± 3.24 a | 10.00 ± 0.30 a | 5.21 ± 0.22 b | 1.12 ± 0.13 a | 17.64 ± 0.97 ab | 2.60 ± 0.24 b | 6.87 ± 0.64 a | 0.28 ± 0.04 b | 1.23 ± 0.26 b |
| CP | 60.10 ± 2.68 a | 9.61 ± 0.07 a | 6.39 ± 0.31 a | 0.73 ± 0.02 a | 14.78 ± 0.49 b | 2.49 ± 0.03 b | 5.95 ± 0.25 a | 0.89 ± 0.09 a | 1.25 ± 0.21 b | |
| BP | 36.06 ± 5.49 a | 9.22 ± 0.17 a | 5.67 ± 0.11 ab | 1.12 ± 0.12 a | 18.32 ± 0.89 a | 3.63 ± 0.15 a | 5.05 ± 0.14 a | 0.86 ± 0.10 a | 3.01 ± 0.38 a | |
| October | NF | 63.95 ± 6.42 a | 20.23 ± 0.30 a | 4.12 ± 0.01 a | 1.20 ± 0.24 a | 14.88 ± 0.33 a | 4.23 ± 0.22 a | 3.52 ± 0.10 a | 0.43 ± 0.04 b | 1.41 ± 0.14 b |
| CP | 60.52 ± 8.08 a | 19.37 ± 0.25 a | 4.19 ± 0.04 a | 0.83 ± 0.06 a | 12.70 ± 0.41 a | 3.43 ± 0.22 a | 3.72 ± 0.16 a | 0.64 ± 0.08 ab | 1.66 ± 0.12 b | |
| BP | 71.24 ± 5.70 a | 18.13 ± 0.17 b | 4.15 ± 0.02 a | 0.96 ± 0.04 a | 16.43 ± 1.73 a | 4.16 ± 0.38 a | 3.94 ± 0.16 a | 0.90 ± 0.11 a | 3.22 ± 0.36 a | |
| Whole | NF | 52.96 ± 5.88 a | 15.12 ± 2.30 a | 4.67 ± 0.26 a | 1.16 ± 0.12 a | 16.26 ± 0.77 ab | 3.42 ± 0.39 a | 5.20 ± 0.80 a | 0.36 ± 0.04 b | 1.32 ± 0.14 b |
| CP | 60.31 ± 3.81 a | 14.49 ± 2.19 a | 5.29 ± 0.51 a | 0.78 ± 0.04 b | 13.74 ± 0.54 b | 2.96 ± 0.23 a | 4.83 ± 0.51 a | 0.76 ± 0.08 a | 1.45 ± 0.14 a | |
| BP | 53.65 ± 8.63 a | 13.68 ± 2.00 a | 4.91 ± 0.34 a | 1.04 ± 0.07 b | 17.37 ± 0.97 a | 3.90 ± 0.22 a | 4.49 ± 0.27 a | 0.88 ± 0.07 a | 3.12 ± 0.24 a | |
| p values in ANOVA | Forest | 0.213 | 0.004 | 0.027 | 0.012 | 0.020 | 0.039 | 0.190 | 0.008 | 0.003 |
| Time | 0.013 | <0.001 | <0.001 | 0.952 | 0.027 | <0.001 | <0.001 | 0.176 | 0.117 | |
| F × T | 0.106 | 0.034 | 0.033 | 0.639 | 0.890 | 0.056 | 0.018 | <0.001 | 0.799 |
| Kingdom | Phylum | Natural Forest | Coniferous Plantation | Broad-Leaved Plantation |
|---|---|---|---|---|
| Bacteria | Proteobacteria | 80.75 ± 1.00 a | 82.06 ± 0.73 a | 82.25 ± 0.59 a |
| Bacteria | Actinobacteria | 14.32 ± 0.58 a | 12.70 ± 0.83 a | 12.88 ± 0.84 a |
| Bacteria | Firmicutes | 1.90 ± 0.55 a | 1.68 ± 0.51 a | 2.14 ± 0.64 a |
| Bacteria | Bacteroidetes | 0.17 ± 0.05 a | 1.21 ± 0.80 a | 0.20 ± 0.02 a |
| Bacteria | Acidobacteria | 1.68 ± 0.28 a | 1.23 ± 0.11 a | 1.42 ± 0.24 a |
| Fungi | Ascomycota | 0.48 ± 0.32 a | 0.11 ± 0.02 a | 0.11 ± 0.05 a |
| Bacteria | Verrucomicrobia | 0.24 ± 0.02 b | 0.35 ± 0.03 a | 0.32 ± 0.03 ab |
| Bacteria | Nitrospirae | 0.02 ± 0.01 b | 0.13 ± 0.03 a | 0.07 ± 0.01 ab |
| Bacteria | Chloroflexi | 0.11 ± 0.01 a | 0.14 ± 0.04 a | 0.21 ± 0.03 a |
| Bacteria | Planctomycetes | 0.15 ± 0.02 a | 0.17 ± 0.01 a | 0.19 ± 0.03 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, L.; Yang, Y.; Chong, Y.; Xiong, J.; Wu, J.; Ai, X.; Guo, Q.; Yuan, Y.; Li, Z. Higher Soil Aggregate Stability in Subtropical Coniferous Plantations Than Natural Forests Due to Microbial and Aggregate Factors. Forests 2022, 13, 2110. https://doi.org/10.3390/f13122110
Cai L, Yang Y, Chong Y, Xiong J, Wu J, Ai X, Guo Q, Yuan Y, Li Z. Higher Soil Aggregate Stability in Subtropical Coniferous Plantations Than Natural Forests Due to Microbial and Aggregate Factors. Forests. 2022; 13(12):2110. https://doi.org/10.3390/f13122110
Chicago/Turabian StyleCai, Lin, Yujing Yang, Yujie Chong, Jiatai Xiong, Juyang Wu, Xunru Ai, Qiuju Guo, Yiping Yuan, and Zhongqiang Li. 2022. "Higher Soil Aggregate Stability in Subtropical Coniferous Plantations Than Natural Forests Due to Microbial and Aggregate Factors" Forests 13, no. 12: 2110. https://doi.org/10.3390/f13122110