
Contribution of Arbuscular Mycorrhizal Fungal Communities to Soil Carbon Accumulation during the Development of Cunninghamia lanceolata Plantations
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sample Collection
2.3. Soil Chemical Analyses
2.4. Mycorrhizal Colonization, Spore Density, Phospholipid Lipid Fatty Acids and Glomalin-Related Soil Proteins Analyses
2.5. Soil DNA Extraction and High-Throughput Sequencing
2.6. Bioinformatics and Statistics
3. Results
3.1. Soil Physicochemical Properties and AM Fungal Biomass
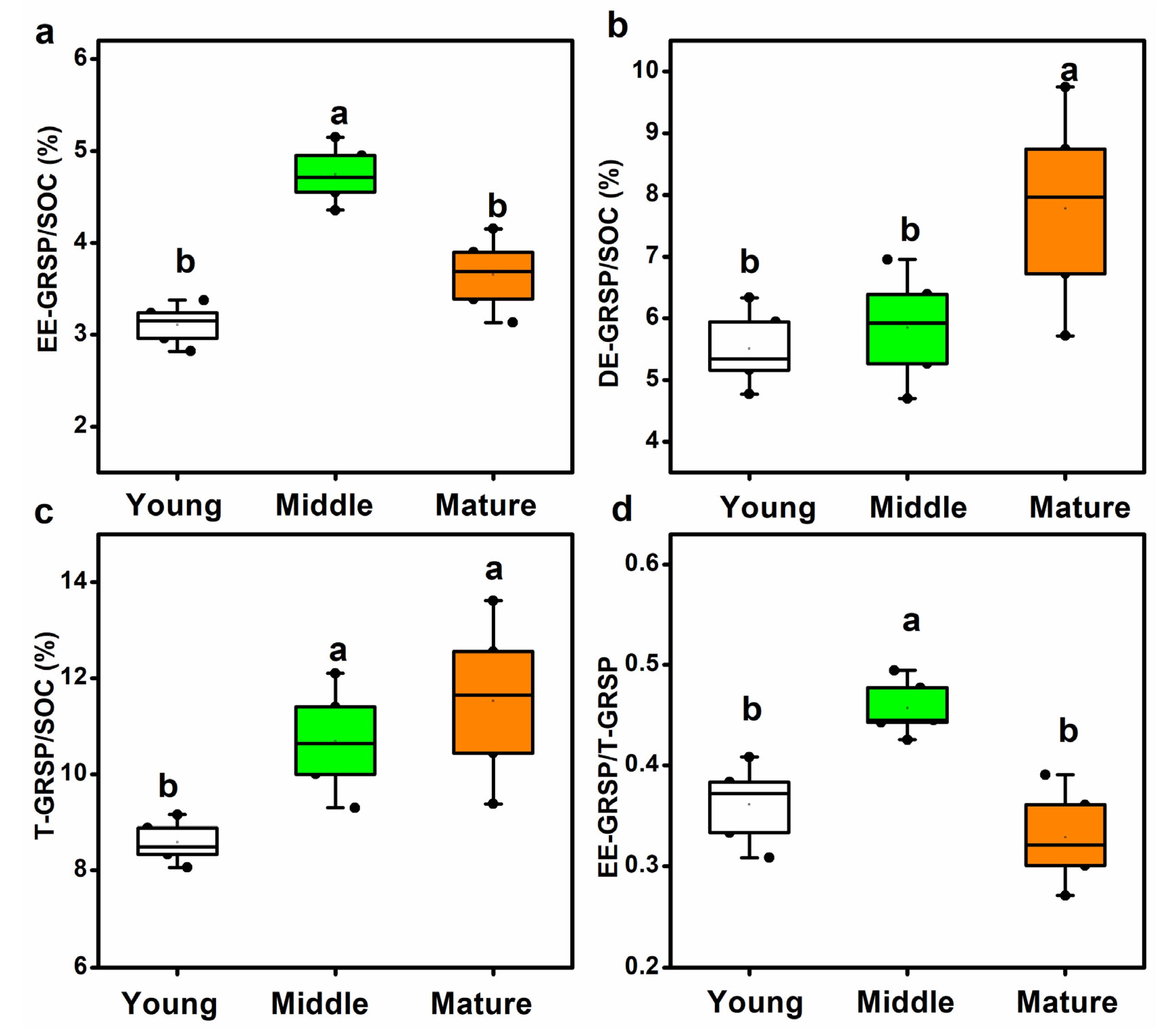
3.2. Mycorrhizal Colonization, Spore Density, AM Fungal Biomass and Glomalin-Related Soil Protein Content
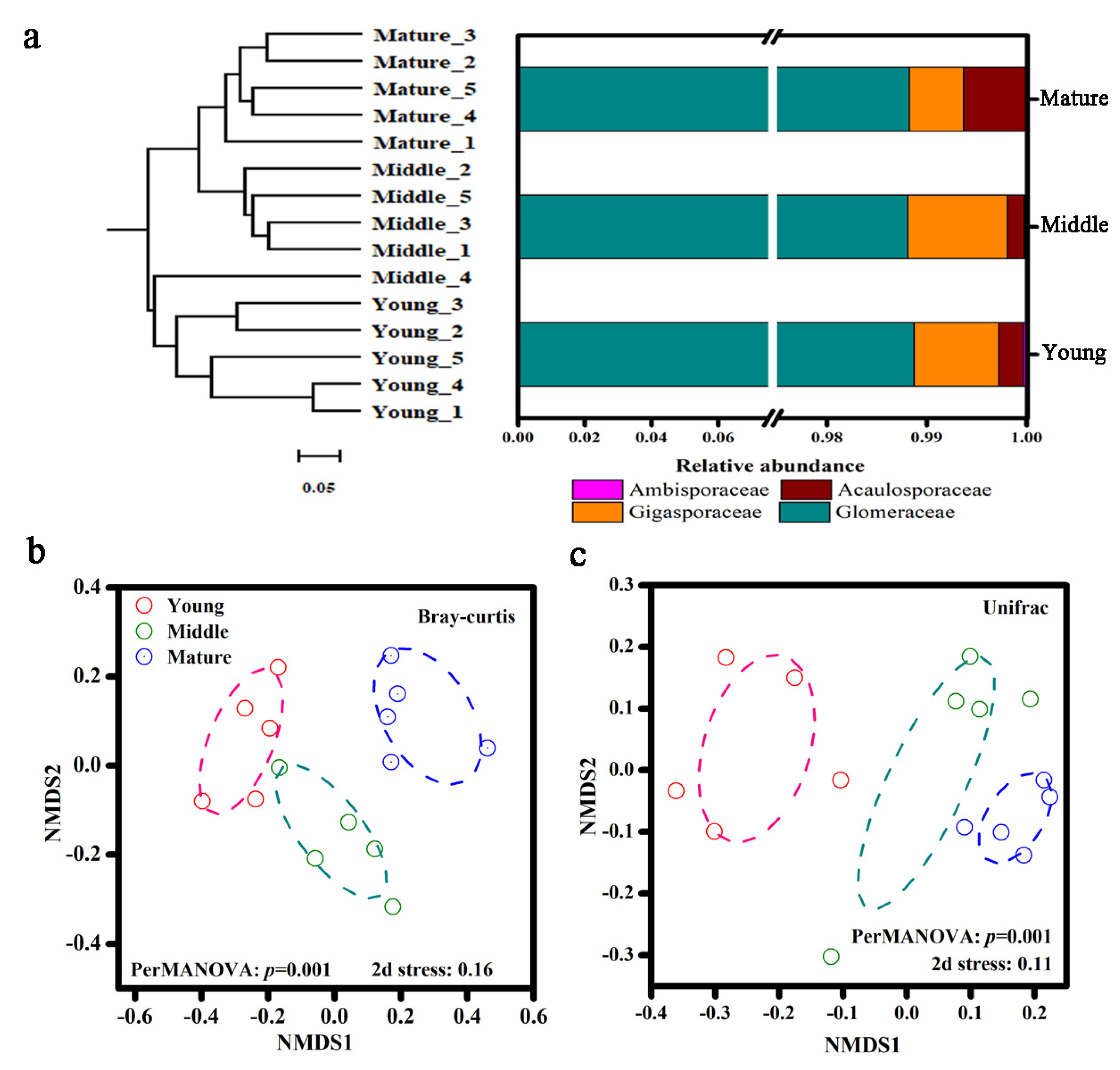
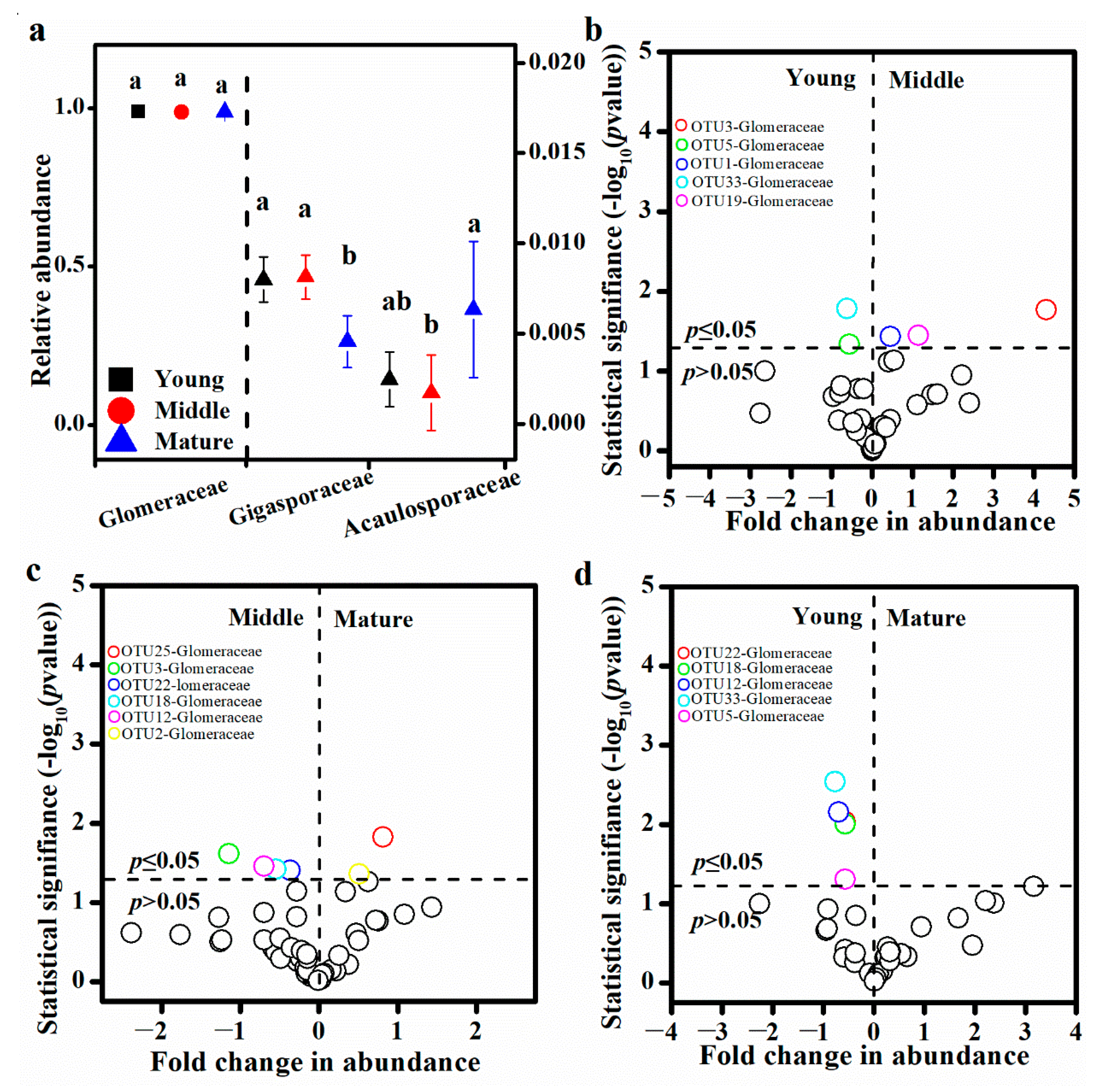
3.3. Soil AM Fungal Diversity and Community Composition
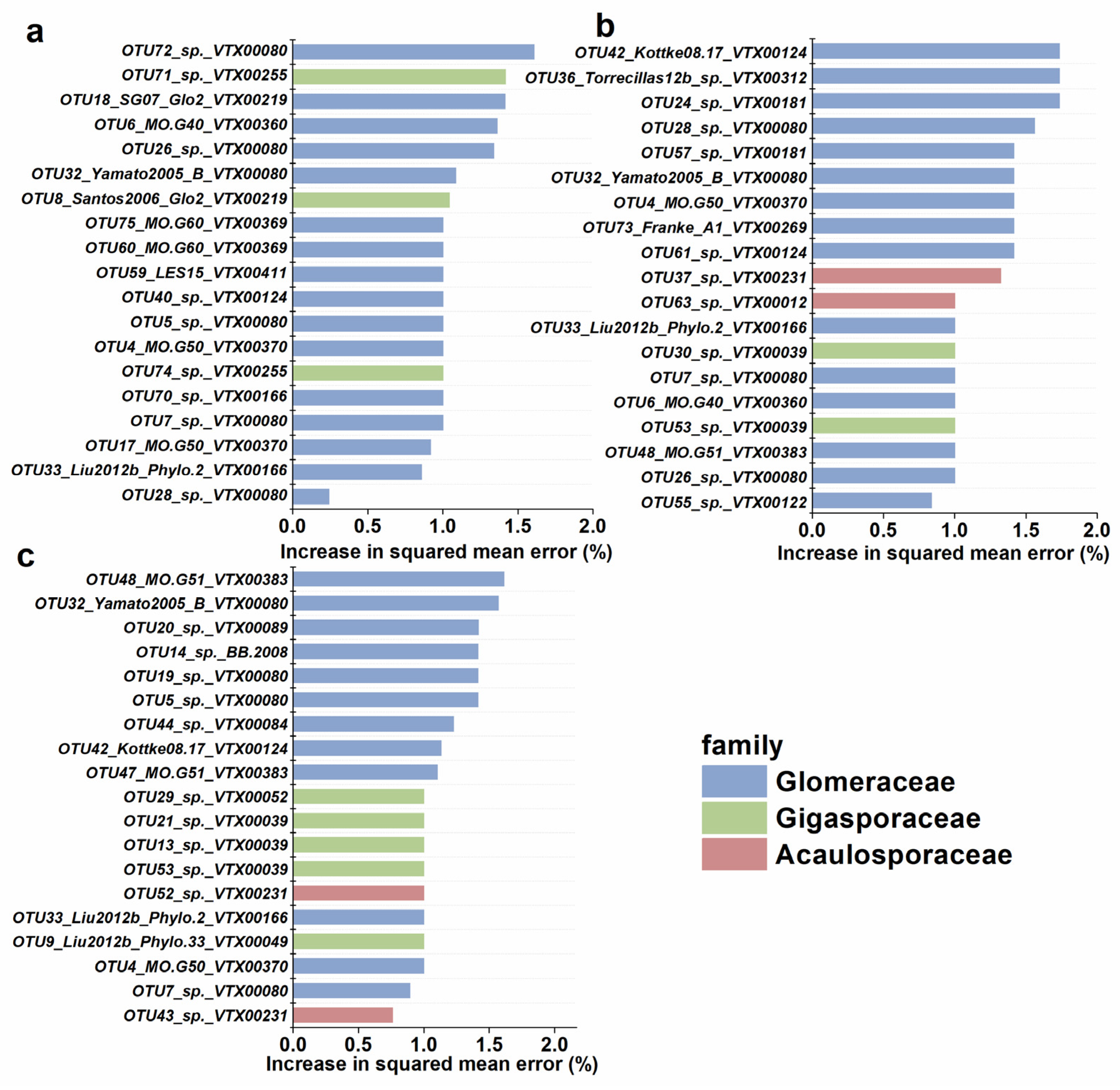
3.4. Potential Key AM Fungal Taxa
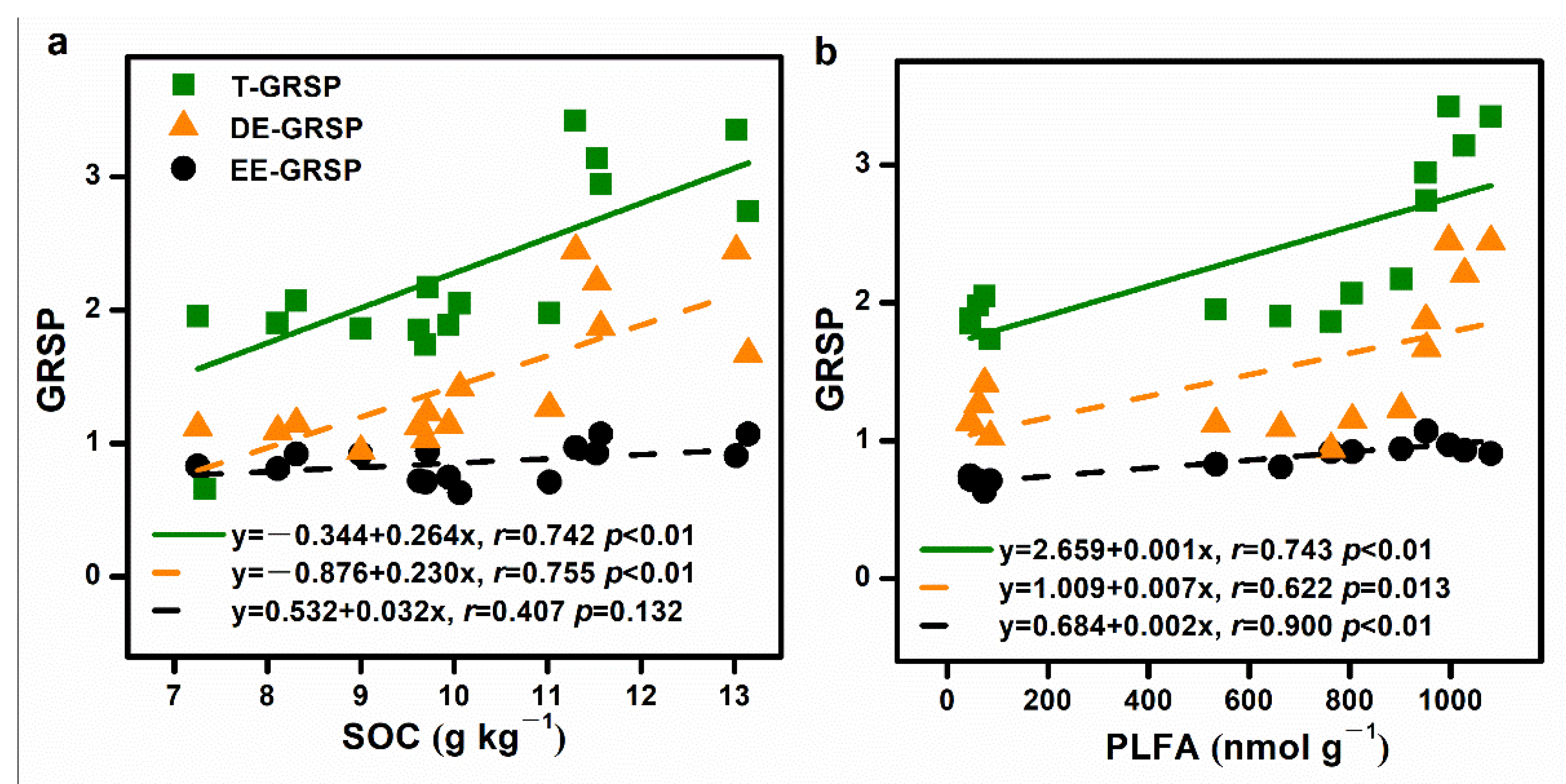
3.5. Linkage between AM Fungal Community and Soil Environmental Factors
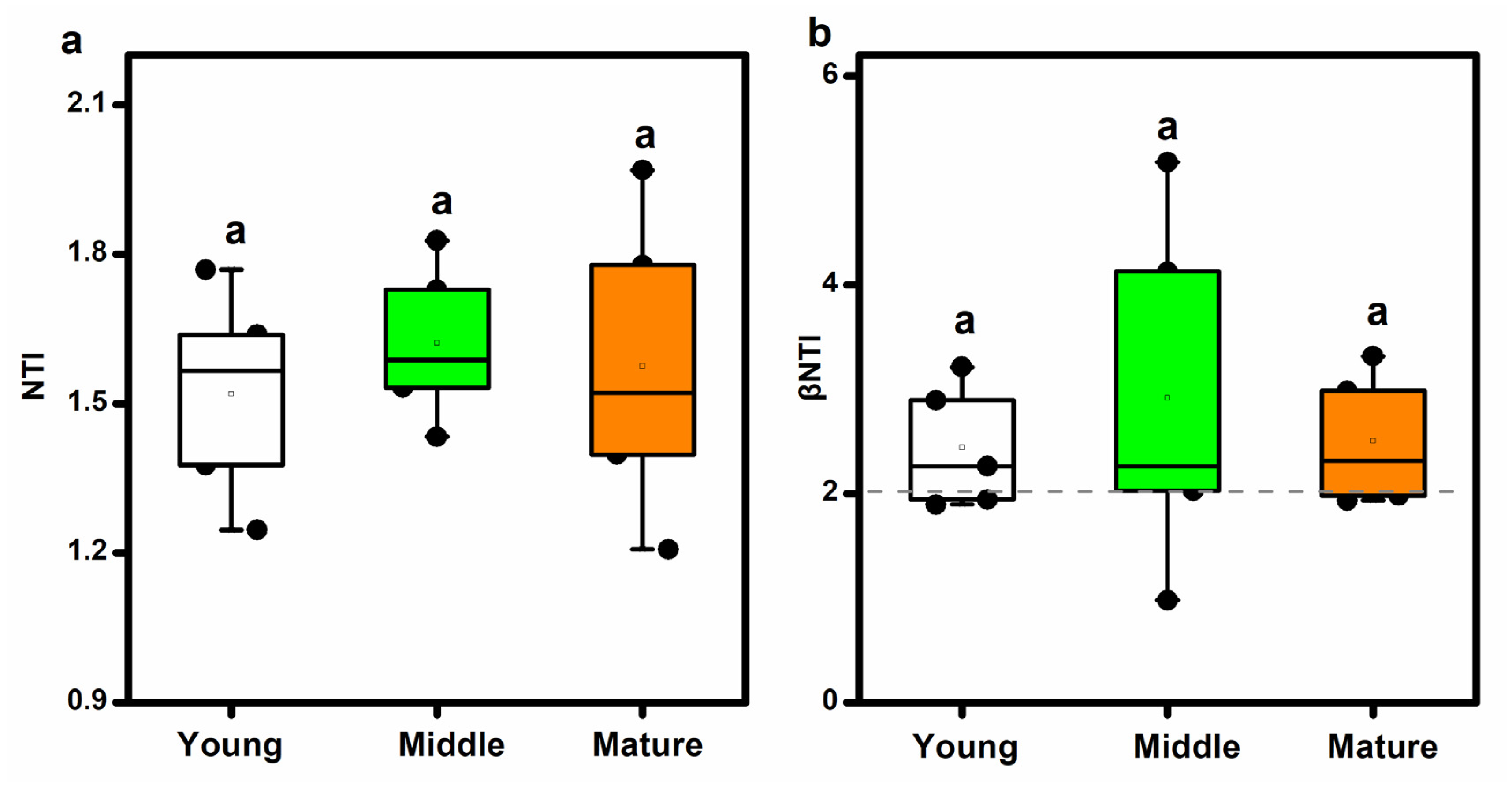
3.6. Phylogenetic Structure of Soil AM Fungal Community
4. Discussion
4.1. Increased AM Fungal Diversity and Altered Community Composition during the Development of C. lanceolata Plantations
4.2. Stimulated Contribution of AM Fungi-Associated Soil C to Soil C Pools during the Development of C. lanceolata Plantations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- State Forestry Administration of China. Forest Resources Report in China (2009–2013); China Forestry Publishing House: Beijing, China, 2014. (In Chinese)
- Jones, I.L.; DeWalt, S.J.; Lopez, O.R.; Bunnefeld, L.; Pattison, Z.; Dent, D.H. Above- and belowground carbon stocks are decoupled in secondary tropical forests and are positively related to forest age and soil nutrients respectively. Sci. Total Environ. 2019, 697, 133987. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Hedo, J.; Cerdá, A.; Candel-Pérez, D.; Viñegla, B. Unravelling the importance of forest age stand and forest structure driving microbiological soil properties, enzymatic activities and soil nutrients content in Mediterranean Spanish black pine (Pinus nigra Ar. ssp. salzmannii) forest. Sci. Total Environ. 2016, 562, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.Y.; Zou, B.Z.; Huang, Z.Q.; Wang, S.R.; Su, X.P.; Ding, X.X.; Zheng, G.C.; Chen, H.Y.H. Fine root biomass and necromass dynamics of Chinese fir plantations and natural secondary forests in subtropical China. For. Ecol. Manag. 2021, 496, 119413. [Google Scholar] [CrossRef]
- Yu, S.B.; Wang, D.; Dai, W.; Li, P. Soil carbon budget in different-aged Chinese fir plantations in south China. J. For. Res. 2014, 25, 621–626. [Google Scholar] [CrossRef]
- Wang, C.Q.; Lin, X.; Jiao, R.Z. Soil organic carbon fractions, C-cycling associated hydrolytic enzymes, and microbial carbon metabolism vary with stand age in Cunninghamia lanceolate (Lamb.) Hook plantations. For. Ecol. Manag. 2021, 482, 118887. [Google Scholar] [CrossRef]
- Barber, N.A.; Chantos-Davidson, K.M.; Peralta, R.A.; Sherwood, J.P.; Swingley, W.D. Soil microbial community composition in tallgrass prairie restorations converge with remnants across a 27-year chronosequence. Environ. Microbiol. 2017, 19, 3118–3131. [Google Scholar] [CrossRef]
- Cao, J.L.; Zheng, Y.X.; Yang, Y.S. Phylogenetic structure of soil bacterial communities along age sequence of subtropical cunninghamia lanceolata plantations. Sustainability 2020, 12, 1864. [Google Scholar] [CrossRef] [Green Version]
- Berg, G. Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef]
- Rillig, M.C.; Wright, S.F.; Nichols, K.A.; Schmidt, W.F.; Torn, M.S. Large contribution of arbuscular mycorrhizal fungi to soil carbon pools in tropical forest soils. Plant Soil 2001, 233, 167–177. [Google Scholar] [CrossRef]
- Treseder, K.K.; Holden, S.R. Fungal carbon sequestration. Science 2013, 339, 1528–1529. [Google Scholar] [CrossRef]
- Wilson, G.W.T.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, E.; Veresoglou, S.D.; Anderson, I.C.; Caruso, T.; Hammer, E.C.; Kohler, J.; Rillig, M.C. Arbuscular mycorrhizal fungi—Short-term liability but long-term benefits for soil carbon storage? New Phytol. 2013, 197, 366–368. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Booker, F.L.; Tu, C.; Burkey, K.O.; Zhou, L.S.; Shew, H.D.; Rufty, T.W.; Hu, S.J. Arbuscular mycorrhizal fungi increase organic carbon decomposition under elevated CO2. Science 2012, 337, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Treseder, K.K.; Turner, K.M.; Mack, M.C. Mycorrhizal responses to nitrogen fertilization in boreal ecosystems: Potential consequences for soil carbon storage. Glob. Chang. Biol. 2007, 13, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Carrington, M.E.; Bardahl, J.G.; Dillenburg, L.M. Glomalin-related soil protein (GRSP) across a chronosequence of tallgrass prairie restorations in northeastern Illinois. In Proceedings of the 95th ESA Annual Meeting, Pittsburgh, PA, USA, 1–6 August 2010. [Google Scholar]
- Zhu, R.H.; Zheng, Z.C.; Li, T.X.; He, S.Q.; Zhang, S.Z.; Wang, Y.D.; Liu, T. Effect of tea plantation age on the distribution of glomalin-related soil protein in soil water-stable aggregates in southwestern China. Environ. Sci. Pollut. Res. 2019, 26, 1973–1982. [Google Scholar] [CrossRef]
- Qin, H.; Chen, J.H.; Wu, Q.F.; Niu, L.M.; Li, Y.C.; Liang, C.F.; Shen, Y.; Xu, Q.F. Intensive management decreases soil aggregation and changes the abundance and community compositions of arbuscular mycorrhizal fungi in Moso bamboo (Phyllostachys pubescens) forests. For. Ecol. Manag. 2017, 400, 246–255. [Google Scholar] [CrossRef]
- Wang, T.; Shen, Y.K.; Wang, Y.X.; Gan, B.P.; Li, Q.; Liu, M.H.; Song, X.Z. Effects of nitrogen and phosphorus addition on arbuscular mycorrhizal fungi and easily extracted glomalin-related soil protein in the Chinese Fir plantation. Terr. Ecosyst. Conserv. 2021, 1, 1–10. [Google Scholar]
- Tang, L.L.; Wang, S.Q. Dynamics of soil aggregate-related C-N-P stoichiometric characteristics with stand age and soil depth in Chinese fir plantations. Land Degrad. Dev. 2022, 33, 1290–1306. [Google Scholar] [CrossRef]
- Angelard, C.; Tanner, C.J.; Fontanillas, P.; Niculita-Hirzel, H.; Masclaux, F.; Sanders, I.R. Rapid genotypic change and plasticity in arbuscular mycorrhizal fungi is caused by a host shift and enhanced by segregation. ISME J. 2014, 8, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Lu, N.; Xu, X.F.; Wang, P.; Zhang, P.; Ji, B.M.; Wang, X.J. Succession in arbuscular mycorrhizal fungi can be attributed to a chronosequence of Cunninghamia lanceolata. Sci. Rep. 2019, 9, 18057. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.N.; Guo, H.T.; Wang, J.P.; Huang, Z.H.; Zhang, J.C.; Liu, G.Z.; Wang, L.Y.; Jiang, J. Study on the characteristics of mycorrhizal colonization in Chinese fir plantations at different ages. Acta Ecol. Sin. 2019, 39, 1926–1934. (In Chinese) [Google Scholar]
- Yu, X.T. The Chinese Fir Silviculture; Fujian Science and Technology Press: Fuzhou, China, 1997. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. Methods Soil Anal. Part 2 Chem. Microbiol. Prop. 1982, 9, 539–580. [Google Scholar]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–45. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular-arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Olsson, P.A. Signature fatty acids provide tools for determination of the distribution and interactions of mycorrhizal fungi in soil. FEMS Microbiol. Ecol. 1999, 29, 303–310. [Google Scholar] [CrossRef]
- Wright, S.F.; Upadhyaya, A. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Sci. 1996, 161, 575–586. [Google Scholar] [CrossRef]
- Schindler, F.V.; Mercer, E.J.; Rice, J.A. Chemical characteristics of glomalinrelated soil protein (GRSP) extracted from soils of varying organic matter content. Soil Biol. Biochem. 2007, 39, 320–329. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, X.L.; He, X.H.; Liu, J.X. Glomalin-related soil protein responses to elevated CO2 and nitrogen addition in a subtropical forest: Potential consequences for soil carbon accumulation. Soil Biol. Biochem. 2015, 83, 142–149. [Google Scholar] [CrossRef]
- Biddle, J.F.; Fitz-Gibbon, S.; Schuster, S.C.; Brenchley, J.E.; House, C.H. Metagenomic signatures of the Peru Margin subseafloor biosphere show a genetically distinct environment. Proc. Natl. Acad. Sci. USA 2008, 105, 10583–10588. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of highthroughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Opik, M.; Vanatoa, A.; Vanatoa, E.; Moora, M.; Davison, J.; Kalwij, J.M.; Reier, U.; Zobel, M. The online database MaarjAM reveals global and ecosystemic distribution patterns in arbuscular mycorrhizal fungi (Glomeromycota). New Phytol. 2010, 188, 223–241. [Google Scholar] [CrossRef]
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. mvabund-An R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed]
- Stegen, J.C.; Lin, X.J.; Konopka, A.E.; Fredrickson, J.K. Stochastic and deterministic assembly processes in subsurface microbial communities. ISME J. 2012, 6, 1653–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleveland, C.C.; Townsend, A.R.; Taylor, P.; Alvarez-Clare, S.; Bustamante, M.M.C.; Chuyong, G.; Dobrowski, S.Z.; Grierson, P.; Harms, K.E.; Houlton, B.Z. Relationships among net primary productivity, nutrients and climate in tropical rain forest: A pan-tropical analysis. Ecol. Lett. 2011, 14, 939. [Google Scholar] [CrossRef]
- Hart, M.M.; Gorzelak, M.; Ragone, D.; Murch, S. Arbuscular mycorrhizal fungal succession in a long-lived perennia. Botany 2014, 92, 313–320. [Google Scholar] [CrossRef]
- Beauchamp, V.B.; Stromberg, J.C.; Stutz, J.C. Arbuscular mycorrhizal fungi associated with Populus–Salix stands in a semiarid riparian ecosystem. New Phytol. 2006, 170, 369–380. [Google Scholar] [CrossRef]
- Sheng, M.; Chen, X.D.; Zhang, X.L.; Hamel, C.; Cui, X.W.; Chen, J.; Chen, H.; Tang, M. Changes in arbuscular mycorrhizal fungal attributes along a chronosequence of black locust (Robinia pseudoacacia) plantations can be attributed to the plantation-induced variation in soil properties. Sci. Total Environ. 2017, 599, 273–283. [Google Scholar] [CrossRef]
- Johnson, N.C. Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales. New Phytol. 2010, 185, 631–647. [Google Scholar] [CrossRef]
- van Diepen, L.T.A.; Lilleskov, E.A.; Pregitzer, K.S. Simulated nitrogen deposition affects community structure of arbuscular mycorrhizal fungi in northern hardwood forests. Mol. Ecol. 2011, 20, 799–811. [Google Scholar] [CrossRef]
- Hart, M.M.; Reader, R.J. Taxonomic basis for variation in the colonization strategy of arbuscular mycorrhizal fungi. New Phytol. 2002, 153, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Chagnon, P.L.; Bradley, R.L.; Maherali, H. A trait-based framework to understand life history of mycorrhizal fungi. Trends Plant Sci. 2013, 18, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R.; Parrent, J.L.; Hart, M.M.; Klironomos, J.N.; Rillig, M.C.; Maherali, H. Phylogenetic trait conservatism and the evolution of functional trade-offs in arbuscular mycorrhizal fungi. Proc. R. Soc. B Biol. Sci. 2009, 276, 4237–4245. [Google Scholar] [CrossRef] [PubMed]
- Egan, G.; Crawley, M.J.; Fornara, D.A. Effects of long-term grassland management on the carbon and nitrogen pools of different soil aggregate fractions. Sci. Total Environ. 2018, 613, 810–819. [Google Scholar] [CrossRef]
- Horn, S.; Caruso, T.; Verbruggen, E.; Rillig, M.C.; Hempel, S. Arbuscular mycorrhizal fungal communities are phylogenetically clustered at small scales. ISME J. 2014, 8, 2231–2242. [Google Scholar] [CrossRef] [Green Version]
- Johnson, N.C.; Rowland, D.L.; Corkidi, L. Nitrogen enrichment alters mycorrhizal allocation at five mesic to semiarid grasslands. Ecology 2003, 84, 1895–1908. [Google Scholar] [CrossRef]
- Liu, Y.J.; Johnson, N.C.; Mao, L.; Shi, G.X.; Jiang, S.J.; Ma, X.J.; Du, G.Z.; An, L.Z.; Feng, H.Y. Phylogenetic structure of arbuscular mycorrhizal community shifts in response to increasing soil fertility. Soil Biol. Biochem. 2015, 89, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Voets, L.; De La Providencia, I.; Declerck, S. Glomeraceae and Gigasporaceae differ in their ability to form hyphal networks. New Phytol. 2006, 172, 185–188. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef]
- Qin, H.; Niu, L.; Wu, Q.; Chen, J.; Li, Y.; Liang, C.; Xu, Q.; Fuhrmann, J.J.; Shen, Y. Bamboo forest expansion increases soil organic carbon through its effect on soil arbuscular mycorrhizal fungal community and abundance. Plant Soil 2017, 420, 407–421. [Google Scholar] [CrossRef]
- Lovelok, C.E.; Wright, S.F.; Nichols, K.A. Using glomalin as an indicator for arbuscular mycorrhizal hyphal growth: An example from a tropical rain forest soil. Soil Biol. Biochem. 2004, 36, 1009–1012. [Google Scholar] [CrossRef]
- Chi, G.G.; Wu, Q.S. Effects of mycorrhizal fungi on plant and growth soil properties in trifoliate orange seedlings grown in a root–box. Philipp. Agric. Sci. 2017, 3, 271–277. [Google Scholar]
- Zhang, J.; Tang, X.L.; Zhong, S.Y.; Yin, G.C.; Gao, Y.F.; He, X.H. Recalcitrant carbon components in glomalin-related soil protein facilitate soil organic carbon preservation in tropical forests. Sci. Rep. 2017, 7, 2391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Li, L.W.; Chen, J.Y.; Hong, H.L.; Lu, H.L.; Liu, J.C.; Dong, Y.W.; Yan, C.L. Glomalin-related soil protein deposition and carbon sequestration in the old Yellow River delta. Sci. Total Environ. 2018, 625, 619–626. [Google Scholar] [CrossRef]
- He, J.D.; Chi, G.G.; Zou, Y.N.; Shu, B.; Wu, Q.S.; Srivastava, A.K.; Kuča, K. Contribution of glomalin-related soil proteins to soil organic carbon in trifoliate orange. Appl. Soil Ecol. 2020, 154, 103592. [Google Scholar] [CrossRef]
- Zhang, H.Q.; Liu, T.; Wang, Y.Y.; Tang, M. Exogenous arbuscular mycorrhizal fungi increase soil organic carbon and change microbial community in poplar rhizosphere. Plant Soil Environ. 2019, 65, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Bedini, S.; Pellegrino, E.; Avio, L.; Pellegrini, S.; Bazzoffi, P.; Argese, E.; Giovanetti, M. Changes in soil aggregation and glomalin related soil protein content as affected by the arbuscular mycorrhizal fungal species Glomus mosseae and Glomus intraradices. Soil Biol. Biochem. 2009, 41, 1491–1496. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observed OTUs | Chao1 | ACE | Shannon | |
|---|---|---|---|---|
| Young | 20.4 ± 3.36 b | 21.8 ± 2.28 b | 23.2 ± 2.09 b | 2.06 ± 0.17 b |
| Middle | 36.25 ± 1.26 a | 34.9 ± 7.39 a | 36.0 ± 8.11 a | 2.66 ± 0.20 a |
| Mature | 37.6 ± 3.78 a | 38.7 ± 3.45 a | 39.40 ± 3.82 a | 2.53 ± 0.36 a |
| Variable | r | p |
|---|---|---|
| Stand age | 0.5965 | 0.001 |
| Soil moisture | 0.0842 | 0.173 |
| pH | −0.008 | 0.487 |
| DOC | 0.0286 | 0.364 |
| SOC | 0.2889 | 0.009 |
| TN | 0.2500 | 0.037 |
| AN | −0.086 | 0.767 |
| AP | 0.2534 | 0.041 |
| AK | 0.1071 | 0.184 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Han, Y.; Lai, F.; Zhao, H.; Cao, J. Contribution of Arbuscular Mycorrhizal Fungal Communities to Soil Carbon Accumulation during the Development of Cunninghamia lanceolata Plantations. Forests 2022, 13, 2099. https://doi.org/10.3390/f13122099
Liu Z, Han Y, Lai F, Zhao H, Cao J. Contribution of Arbuscular Mycorrhizal Fungal Communities to Soil Carbon Accumulation during the Development of Cunninghamia lanceolata Plantations. Forests. 2022; 13(12):2099. https://doi.org/10.3390/f13122099
Chicago/Turabian StyleLiu, Zhiyuan, Yu Han, Faying Lai, Haiying Zhao, and Jiling Cao. 2022. "Contribution of Arbuscular Mycorrhizal Fungal Communities to Soil Carbon Accumulation during the Development of Cunninghamia lanceolata Plantations" Forests 13, no. 12: 2099. https://doi.org/10.3390/f13122099