
Daily Dynamics of Soil Heat Flux and Its Relationship with Net Radiation in Different Urban Riparian Woodlands

Abstract
:1. Introduction
2. Methods
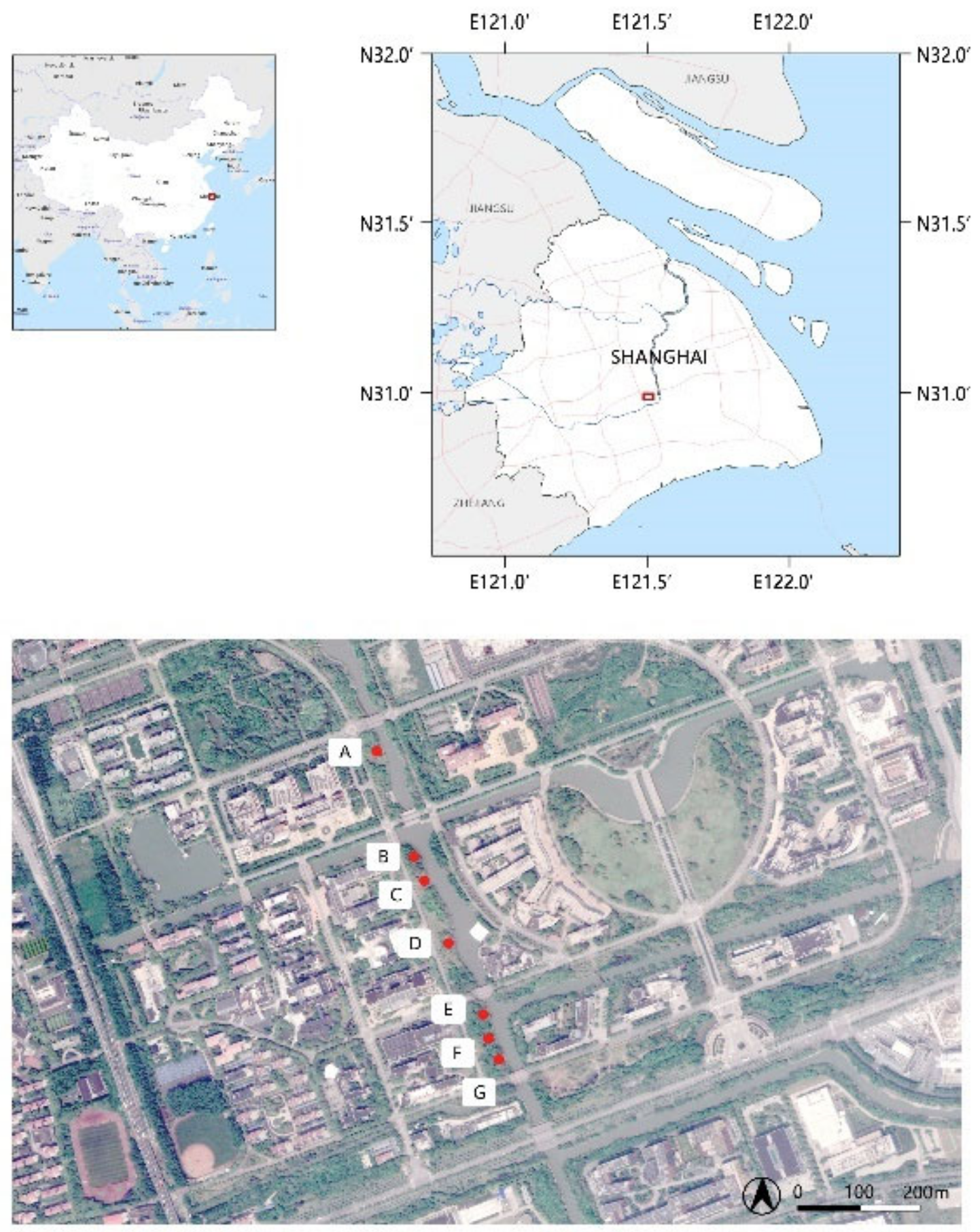
2.1. Site Description
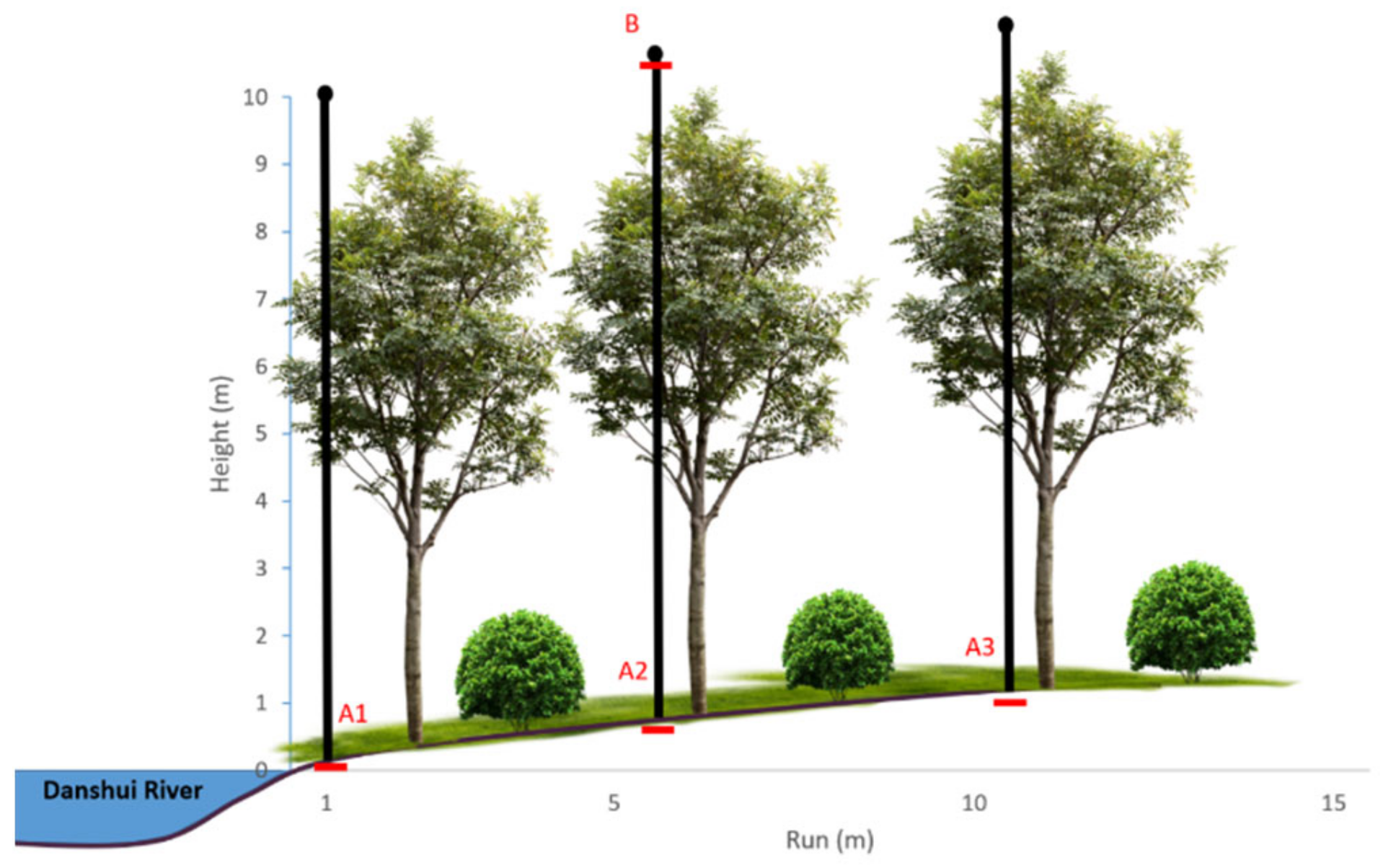
2.2. Experimental Design
2.3. Data Analysis
2.3.1. Data Control
2.3.2. Data Analysis
3. Results
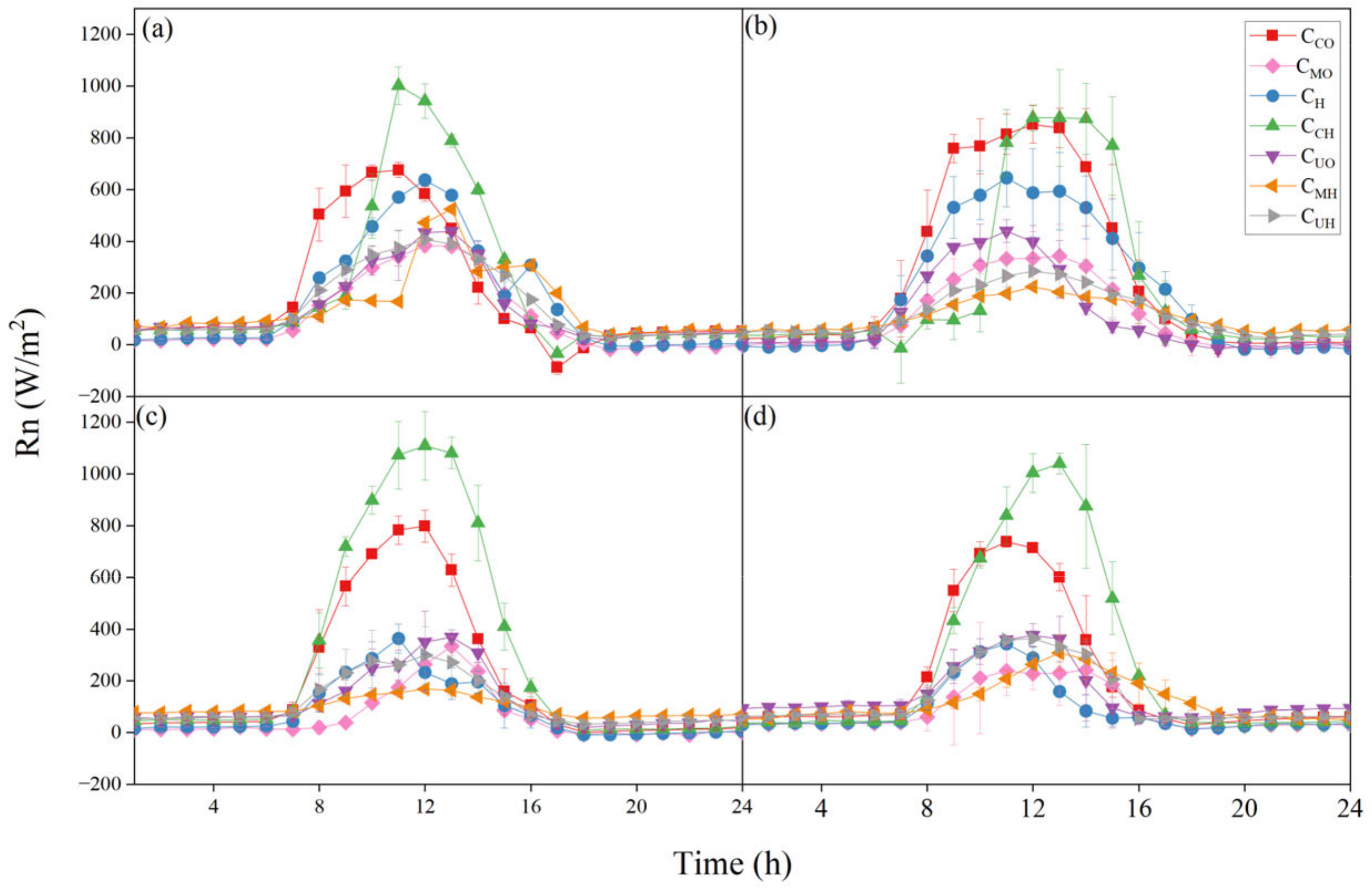
3.1. Diurnal and Seasonal Variations in Rn
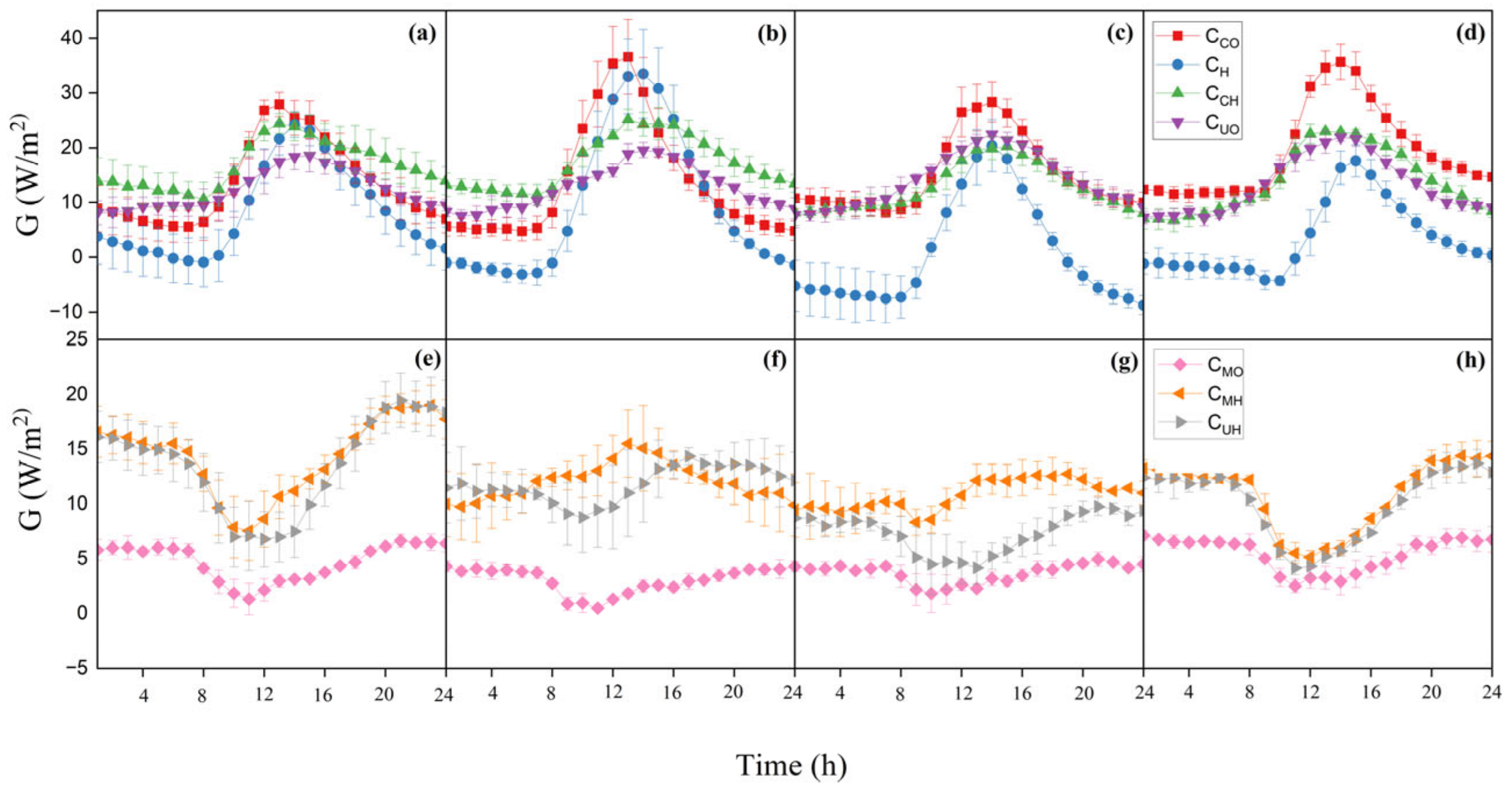
3.2. Diurnal and Seasonal Variations in G
3.3. Relationships between G and Rn
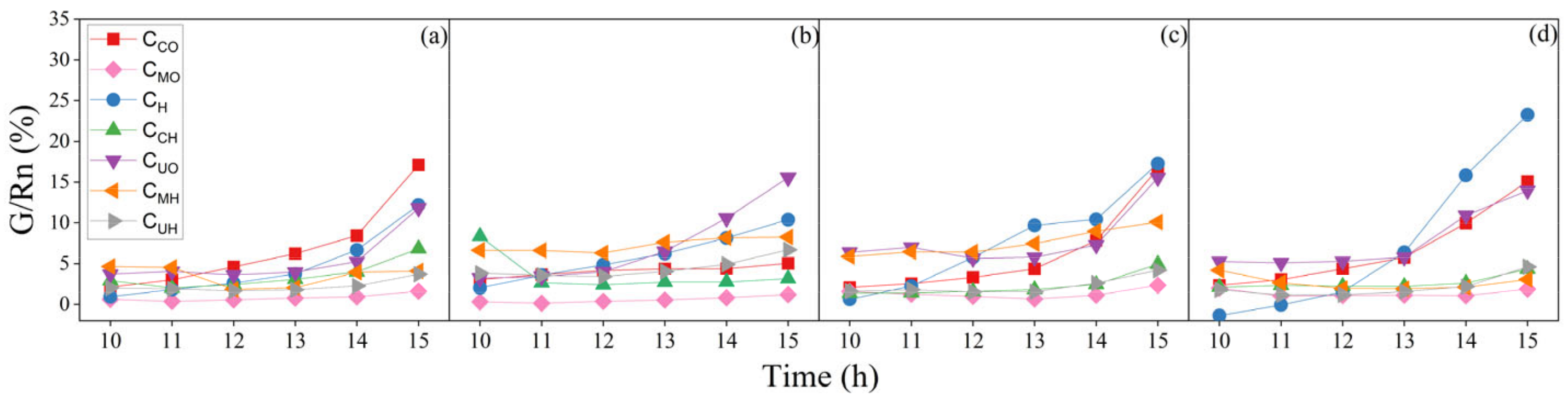
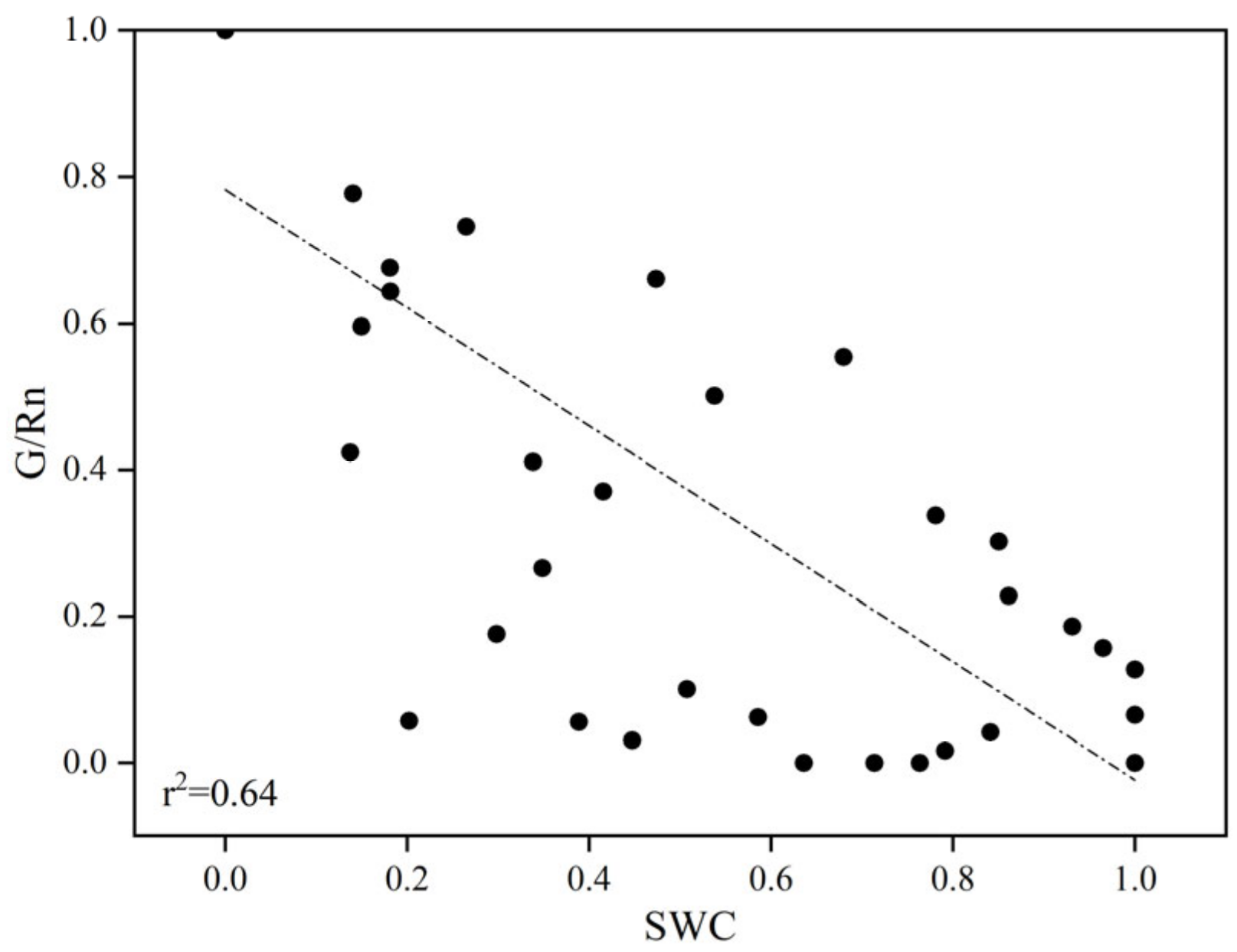
3.3.1. Midday Variations in G/Rn Ratios
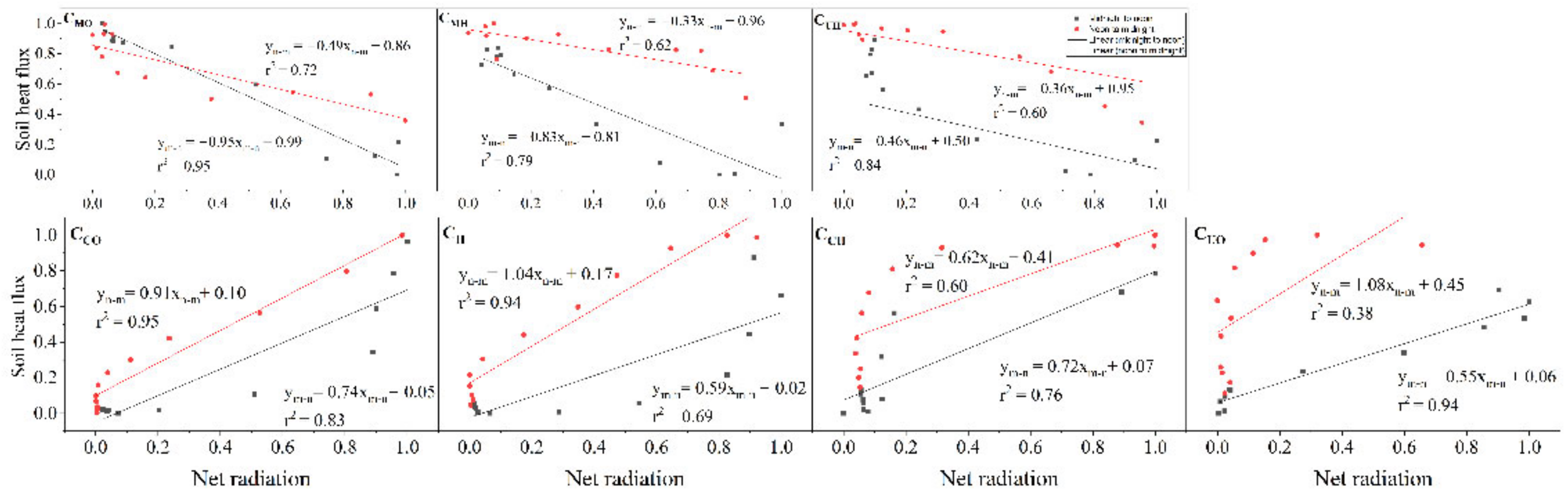
3.3.2. Phase Shift of G to Rn
4. Discussion
4.1. Factors Influencing Rn
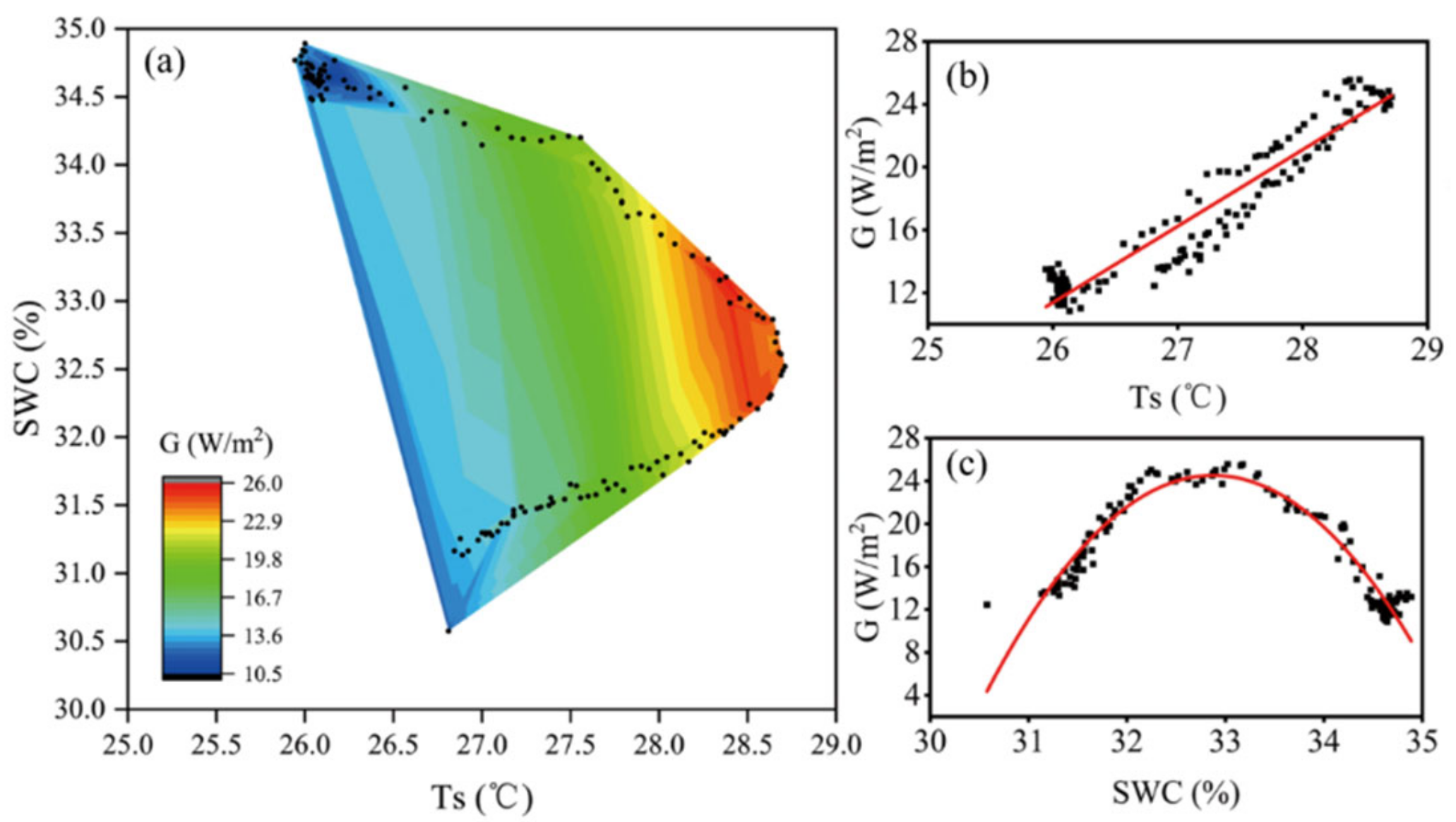
4.2. Factors Influencing G and Its Relationships with Rn
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sun, R.; Chen, L. How can urban water bodies be designed for climate adaptation? Landsc. Urban Plan. 2012, 105, 27–33. [Google Scholar] [CrossRef]
- Zhu, W.; Wu, B.; Yan, N.; Feng, X.; Xing, Q. A method to estimate diurnal surface soil heat flux from MODIS data for a sparse vegetation and bare soil. J. Hydrol. 2014, 511, 139–150. [Google Scholar] [CrossRef]
- Liu, X.; Yang, S.; Xu, J.; Zhang, J.; Liu, J. Effects of soil heat storage and phase shift correction on energy balance closure of paddy fields. Atmósfera 2017, 30, 39–52. [Google Scholar] [CrossRef]
- Bianca, N.I.E.; Madsen, L.; Hagar, J.C.; Temesgen, H. Estimating Riparian Understory Vegetation Cover with Beta Regression and Copula Models. For. Sci. 2011, 57, 212. [Google Scholar]
- Yue, P.; Zhang, Q.; Zhang, L.; Li, H.; Yang, Y.; Zeng, J.; Wang, S. Long-term variations in energy partitioning and evapotranspiration in a semiarid grassland in the Loess Plateau of China. Agric. For. Meteorol. 2019, 278, 107671. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Baker, J.M. Micrometeorology in Agricultural Systems; American Society of Agronomy: Madison, WI, USA, 2005; pp. 131–143. [Google Scholar]
- Tanguy, M.; Baille, A.; González-Real, M.M.; Lloyd, C.; Cappelaere, B.; Kergoat, L.; Cohard, J.M. A new parameterisation scheme of ground heat flux for land surface flux retrieval from remote sensing information. J. Hydrol. 2012, 454–455, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Payero, J.O.; Neale, C.M.U.; Wright, J.L. Estimating soil heat flux for alfalfa and clipped tall fescue grass. Appl. Eng. Agric. 2005, 3, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, P.; Meyers, T.P.; Scott, R.L.; Kennedy, L.; Heuer, M. Energy exchange and evapotranspiration over two temperate semi-arid grasslands in North America. Agric. For. Meteorol. 2012, 153, 31–44. [Google Scholar] [CrossRef]
- Bryś, K.; Bryś, T.; Ojrzyńska, H.; Sayegh, M.A.; Głogowski, A. Variability and role of long-wave radiation fluxes in the formation of net radiation and thermal features of grassy and bare soil active surfaces in Wrocław. Sci. Total Environ. 2020, 747, 141192. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, W.; He, J.; Zhang, K. Energy exchange and evapotranspiration over irrigated seed maize agroecosystems in a desert-oasis region, northwest China. Agric. For. Meteorol. 2016, 223, 48–59. [Google Scholar] [CrossRef]
- Jason Beringer, A.H.L.F. The Representation of Arctic Soils in the Land Surface Model: The Importance of Mosses. J. Clim. 2001, 14, 3324–3335. [Google Scholar] [CrossRef]
- Hernandez-Ramirez, G.; Hatfield, J.L.; Prueger, J.H.; Sauer, T.J. Energy balance and turbulent flux partitioning in a corn–soybean rotation in the Midwestern US. Theor. Appl. Climatol. 2010, 100, 79–92. [Google Scholar] [CrossRef]
- Tianjiao, F.; Dong, W.; Ruoshui, W.; Yixin, W.; Zhiming, X.; Fengmin, L.; Yuan, M.; Xing, L.; Huijie, X.; Caballero-Calvo, A.; et al. Spatial-temporal heterogeneity of environmental factors and ecosystem functions in farmland shelterbelt systems in desert oasis ecotones. Agric. Water Manag. 2022, 271, 107790. [Google Scholar] [CrossRef]
- Drexler, J.Z.; Anderson, F.E.; Snyder, R.L. Evapotranspiration rates and crop coefficients for a restored marsh in the Sacramento–San Joaquin Delta, California, USA. Hydrol. Process. 2008, 22, 725–735. [Google Scholar] [CrossRef]
- Welsh, J.H.H.; Hodgson, G.R.; Karraker, N.E. Influences of the vegetation mosaic on riparian and stream environments in a mixed forest-grassland landscape in “Mediterranean” northwestern California. Ecography 2005, 28, 537–551. [Google Scholar] [CrossRef]
- Acharya, R.H.; Sigdel, M.; Ma, Y.; Wang, B. Diurnal and seasonal variation of heat fluxes over an agricultural field in southeastern Nepal. Theor. Appl. Clim. 2019, 137, 2949–2960. [Google Scholar] [CrossRef]
- Masseroni, D.; Facchi, A.; Romani, M.; Chiaradia, E.A.; Gharsallah, O.; Gandolfi, C. Surface energy flux measurements in a flooded and an aerobic rice field using a single eddy-covariance system. Paddy Water Environ. 2015, 13, 405–424. [Google Scholar] [CrossRef]
- Ma, J.; Zha, T.; Jia, X.; Tian, Y.; Bourque, C.P.A.; Liu, P.; Bai, Y.; Wu, Y.; Ren, C.; Yu, H.; et al. Energy and water vapor exchange over a young plantation in northern China. Agric. For. Meteorol. 2018, 263, 334–345. [Google Scholar] [CrossRef]
- Wilson, K.B.; Baldocchi, D.D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Dolman, H.; Falge, E.; Field, C.; Goldstein, A.; Granier, A.; et al. Energy partitioning between latent and sensible heat flux during the warm season at FLUXNET sites. Water Resour. Res. 2002, 38, 30–31. [Google Scholar] [CrossRef] [Green Version]
- Dupont, S.; Patton, E.G. Influence of stability and seasonal canopy changes on micrometeorology within and above an orchard canopy: The CHATS experiment. Agric. For. Meteorol. 2012, 157, 11–29. [Google Scholar] [CrossRef]
- Leuning, R.; Van Gorsel, E.; Massman, W.J.; Isaac, P.R. Reflections on the surface energy imbalance problem. Agric. For. Meteorol. 2012, 156, 65–74. [Google Scholar] [CrossRef]
- Nelli, N.R.; Temimi, M.; Fonseca, R.M.; Weston, M.J.; Thota, M.S.; Valappil, V.K.; Branch, O.; Wizemann, H.; Wulfmeyer, V.; Wehbe, Y. Micrometeorological measurements in an arid environment: Diurnal characteristics and surface energy balance closure. Atmos. Res. 2020, 234, 104745. [Google Scholar] [CrossRef]
- Scott, R.L.; Edwards, E.A.; Shuttleworth, W.J.; Huxman, T.E.; Watts, C.; Goodrich, D.C. Interannual and seasonal variation in fluxes of water and carbon dioxide from a riparian woodland ecosystem. Agric. For. Meteorol. 2004, 122, 65–84. [Google Scholar] [CrossRef]
- Wang, X.; Zhou, G.; Zhang, D.; Wang, C. Soil heat fluxes of mixed coniferous and broad-leaf forest in the south subtropics in China. Ecol. Environ. 2005, 14, 260–265. (In Chinese) [Google Scholar]
- Wang, X.; Liu, D.; Li, Y.; Wang, Z.; Ma, B. Soil Heat Fluxes of Larix gmelinii in Daxing’anling of Heilongjiang Province. Prot. For. Sci. Technol. 2020, 52, 88–93. (In Chinese) [Google Scholar]
- Hossen, M.S.; Mano, M.; Miyata, A.; Baten, M.A.; Hiyama, T. Surface energy partitioning and evapotranspiration over a double-cropping paddy field in Bangladesh. Hydrol. Process. 2012, 26, 1311–1320. [Google Scholar] [CrossRef]
- Kustas, W.P.; Prueger, J.H.; Hatfield, J.L.; Ramalingam, K.; Hipps, L.E. Variability in soil heat flux from a mesquite dune site. Agric. For. Meteorol. 2000, 103, 249–264. [Google Scholar] [CrossRef]
- Ogée, J.; Lamaud, E.; Brunet, Y.; Berbigier, P.; Bonnefond, J.M. A long-term study of soil heat flux under a forest canopy. Agric. For. Meteorol. 2001, 106, 173–186. [Google Scholar] [CrossRef]
- Shao, C.; Chen, J.; Li, L.; Xu, W.; Chen, S.; Gwen, T.; Xu, J.; Zhang, W. Spatial variability in soil heat flux at three Inner Mongolia steppe ecosystems. Agric. For. Meteorol. 2008, 148, 1433–1443. [Google Scholar] [CrossRef]
- Beringer, J.; Chapin, F.S.; Thompson, C.C.; McGuire, A.D. Surface energy exchanges along a tundra-forest transition and feedbacks to climate. Agric. For. Meteorol. 2005, 131, 143–161. [Google Scholar] [CrossRef]
- Meyers, T.P. A comparison of summertime water and CO2 fluxes over rangeland for well watered and drought conditions. Agric. For. Meteorol. 2001, 106, 205–214. [Google Scholar] [CrossRef]
- Santanello, J.A., Jr.; Friedl, M.A. Diurnal covariation in soil heat flux and net radiation. Am. Meteorol. Soc. 2003, 42, 851–862. [Google Scholar] [CrossRef]
- Guo, D.; Yang, M.; Wang, H. Sensible and latent heat flux response to diurnal variation in soil surface temperature and moisture under different freeze/thaw soil conditions in the seasonal frozen soil region of the central Tibetan Plateau. Environ. Earth Sci. 2011, 63, 97–107. [Google Scholar] [CrossRef]
- Lenters, J.D.; Cutrell, G.J.; Istanbullupglu, E.; Scott, D.T.; Herman, K.S.; Irmak, A.; Eisenhauer, D.E. Water and Energy Balance in Response to the Removal of Invasive Phragmites Australis in a Riparian Wetland. J. Hydrol. 2011, 2011, 19–34. [Google Scholar] [CrossRef] [Green Version]
- Behera, S.K.; Mishra, A.K.; Sahu, N.; Kumar, A.; Singh, N.; Kumar, A.; Bajpai, O.; Chaudhary, L.B.; Khare, P.B.; Tuli, R. The study of microclimate in response to different plant community association in tropical moist deciduous forest from northern India. Biodivers. Conserv. 2012, 21, 1159–1176. [Google Scholar] [CrossRef]
- Zheng, L.; Zhao, G.; Dong, J.; Ge, Q.; Tao, J.; Zhang, X.; Qi, Y.; Doughty, R.B.; Xiao, X. Spatial, temporal, and spectral variations in albedo due to vegetation changes in China’s grasslands. ISPRS J. Photogramm. Remote Sens. 2019, 152, 1–12. [Google Scholar] [CrossRef]
- Kovács, B.; Tinya, F.; Ódor, P. Stand structural drivers of microclimate in mature temperate mixed forests. Agric. For. Meteorol. 2017, 234–235, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Friedl, M.A. Forward and inverse modeling of land surface energy balance using surface temperature measurements. Remote Sens. Environ. 2002, 79, 344–354. [Google Scholar] [CrossRef]
- Gao, Z.; Horton, R.; Liu, H.P. Impact of wave phase difference between soil surface heat flux and soil surface temperature on soil surface energy balance closure. J. Geophys. Res. 2010, 115, D16112. [Google Scholar] [CrossRef]
- Sun, T.; Wang, Z.; Ni, G. Revisiting the hysteresis effect in surface energy budgets. Geophys. Res. Lett. 2013, 40, 1741–1747. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| Plant type | Evergreen broadleaf woodland +shrubland | Coniferous woodland +shrubland | Grassland | Evergreen broadleaf woodland +grassland | Deciduous woodland +shrubland | Coniferous woodland +grassland | Deciduous woodland +grassland |
| Plant type | CCO | CMO | CH | CCH | CUO | CMH | CUH |
| DBH (cm) | 26 ± 2.43 | 18.80 ± 2.95 | / | 26 ± 4.79 | 21 ± 7.23 | 17.50 ± 4.34 | 19 ± 1.87 |
| Height (m) | 9.76 | 12.80 | 0.20 | 9.80 | 9.60 | 13.00 | 9.04 |
| Density (/hm2) | 375 | 1050 | / | 325 | 400 | 850 | 400 |
| Crown ratio | 59.6% | 72.68% | / | 49.45% | 52.20% | 74.91% | 64.85% |
| Summer LAI | 1.80 | 2.99 | / | 2.10 | 2.45 | 3.95 | 3.22 |
| Site photos |  |  |  |  |  |  |  |
| Parameter | Instrument | Model | Accuracy | Measuring Height 1 |
|---|---|---|---|---|
| Net radiation | Net radiometer | QT-1 | ±5% | 10/12 m |
| Soil temperature | Soil temperature and humidity probes | JXBS-3001 | ±0.5 °C | −0.2 m |
| Soil humidity | ±3% | −0.2 m | ||
| Soil heat flux | Soil heat flux plate | HFP01 | <0.1%/°C | −0.2 m |
| ΔRn (W/m2) | CCH | CCO | CH | CUO | CMO | CUH | CMH |
|---|---|---|---|---|---|---|---|
| Spring | 1035.72 | 767.45 | 642.6 | 420.08 | 400.82 | 377.25 | 484.88 |
| Summer | 898.40 | 881.83 | 662.93 | 462.3 | 359.05 | 257.38 | 181.17 |
| Autumn | 1099.93 | 796.17 | 372.32 | 348.17 | 342.85 | 266.17 | 111.77 |
| Winter | 981.98 | 708.63 | 328.73 | 318.54 | 228.68 | 312.75 | 258.84 |
| G0 | G1h | G2h | G3h | G4h | G5h | G6h | G7h | G8h | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| r2 | p | r2 | p | r2 | p | r2 | p | r2 | p | r2 | p | r2 | p | r2 | p | r2 | p | |
| CCO | 0.888 | 0.01 | 0.967 | 0.01 | 0.935 | 0.01 | 0.797 | 0.01 | 0.563 | 0.01 | / | / | / | / | ||||
| CH | 0.759 | 0.01 | 0.913 | 0.01 | 0.986 | 0.01 | 0.957 | 0.01 | 0.817 | 0.01 | 0.567 | 0.05 | / | / | / | |||
| CCH | 0.75 | 0.01 | 0.823 | 0.01 | 0.817 | 0.01 | 0.705 | 0.01 | 0.552 | 0.01 | / | / | / | / | ||||
| CUO | 0.393 | 0.01 | 0.58 | 0.01 | 0.747 | 0.01 | 0.874 | 0.01 | 0.923 | 0.01 | 0.86 | 0.01 | 0.674 | 0.01 | / | / | ||
| CMO | −0.908 | 0.01 | −0.846 | 0.01 | −0.695 | 0.01 | −0.476 | 0.05 | / | / | / | 0.604 | 0.05 | 0.797 | 0.01 | |||
| CMH | −0.671 | 0.01 | −0.448 | 0.05 | / | / | / | 0.685 | 0.01 | 0.831 | 0.01 | 0.876 | 0.01 | 0.834 | 0.01 | |||
| CUH | −0.787 | 0.01 | −0.564 | 0.01 | / | / | / | 0.6 | 0.01 | 0.796 | 0.01 | 0.894 | 0.01 | 0.906 | 0.01 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, A.; Xie, C.; Wang, J.; Che, S. Daily Dynamics of Soil Heat Flux and Its Relationship with Net Radiation in Different Urban Riparian Woodlands. Forests 2022, 13, 2062. https://doi.org/10.3390/f13122062
Liang A, Xie C, Wang J, Che S. Daily Dynamics of Soil Heat Flux and Its Relationship with Net Radiation in Different Urban Riparian Woodlands. Forests. 2022; 13(12):2062. https://doi.org/10.3390/f13122062
Chicago/Turabian StyleLiang, Anze, Changkun Xie, Jing Wang, and Shengquan Che. 2022. "Daily Dynamics of Soil Heat Flux and Its Relationship with Net Radiation in Different Urban Riparian Woodlands" Forests 13, no. 12: 2062. https://doi.org/10.3390/f13122062