

Chances and Limitations of Mixed Oak Regeneration under Continuous Canopy Cover—Evidence from Long-Term Observations

Abstract
:1. Introduction
- i.
- What are the survival probabilities of oak in small-scale and long-term regenerated stands and do they differ between different forest types?
- ii.
- What is the long-term development of the regenerated tree species’ density and the effect of canopy cover on regeneration biomass?
- iii.
- How does canopy cover and interspecific competition modify the heights of regenerated oaks?
- iv.
- Is there a feedback effect of advanced regeneration on the productivity of the mature stand?
2. Materials and Methods
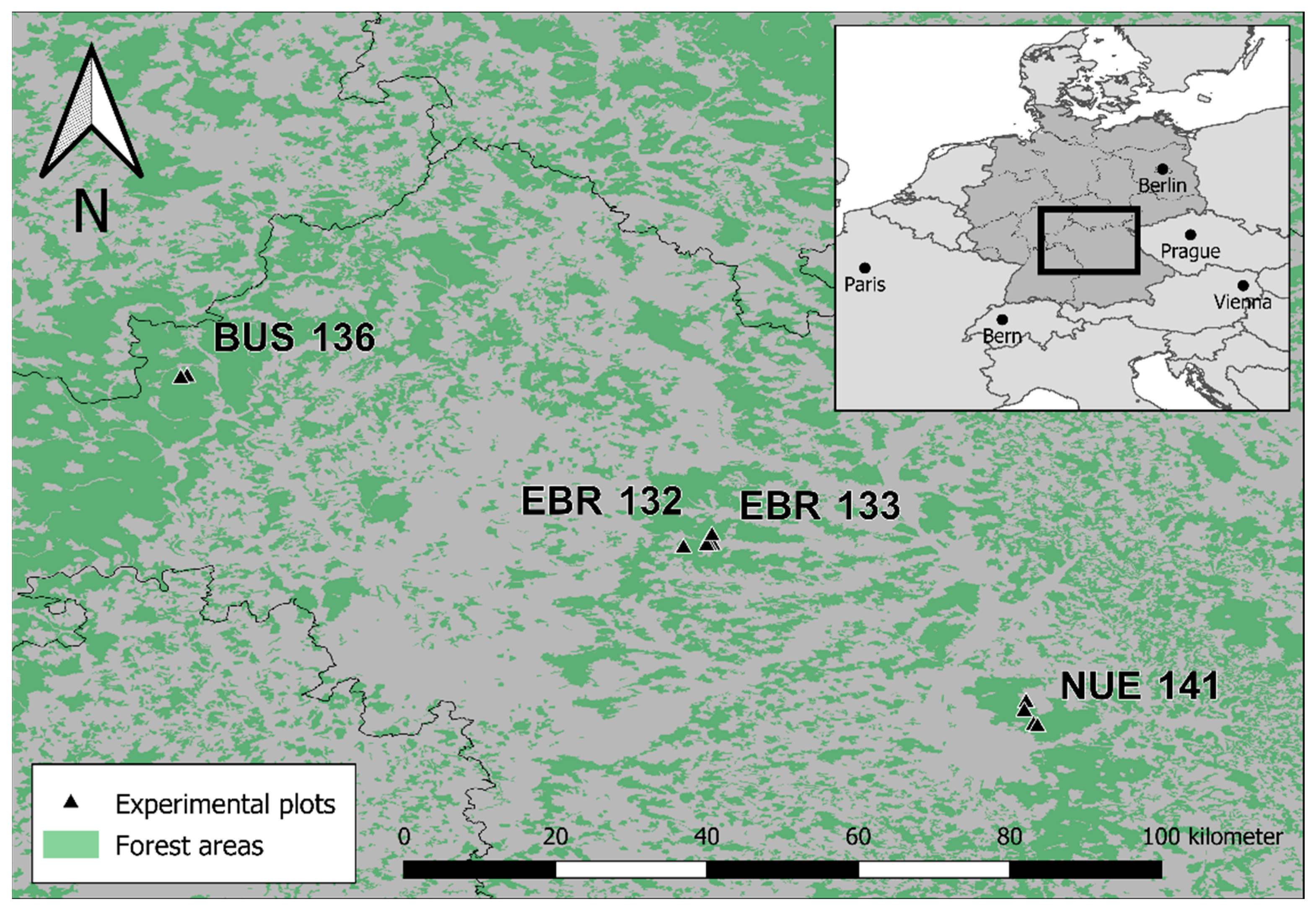
2.1. Long-Term Experiments
2.2. Yield Data of the Mature Stand
2.3. Regeneration Data
2.4. Statistical Analyses
pijk/1 − pijk = a0 + a1 durik + a2 sitei + bi + bij + εijk
3. Results
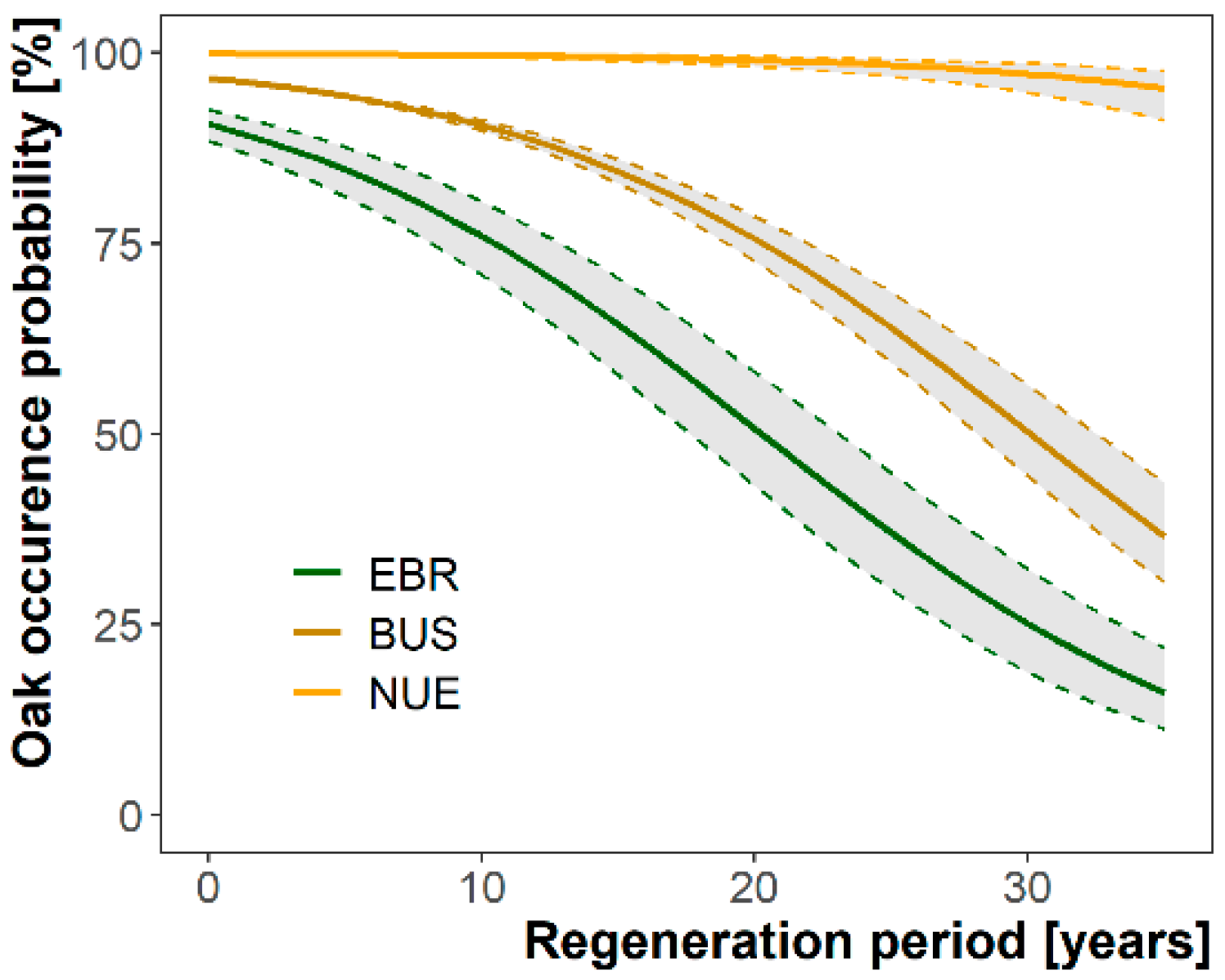
3.1. Oak Occurrence Probabilities
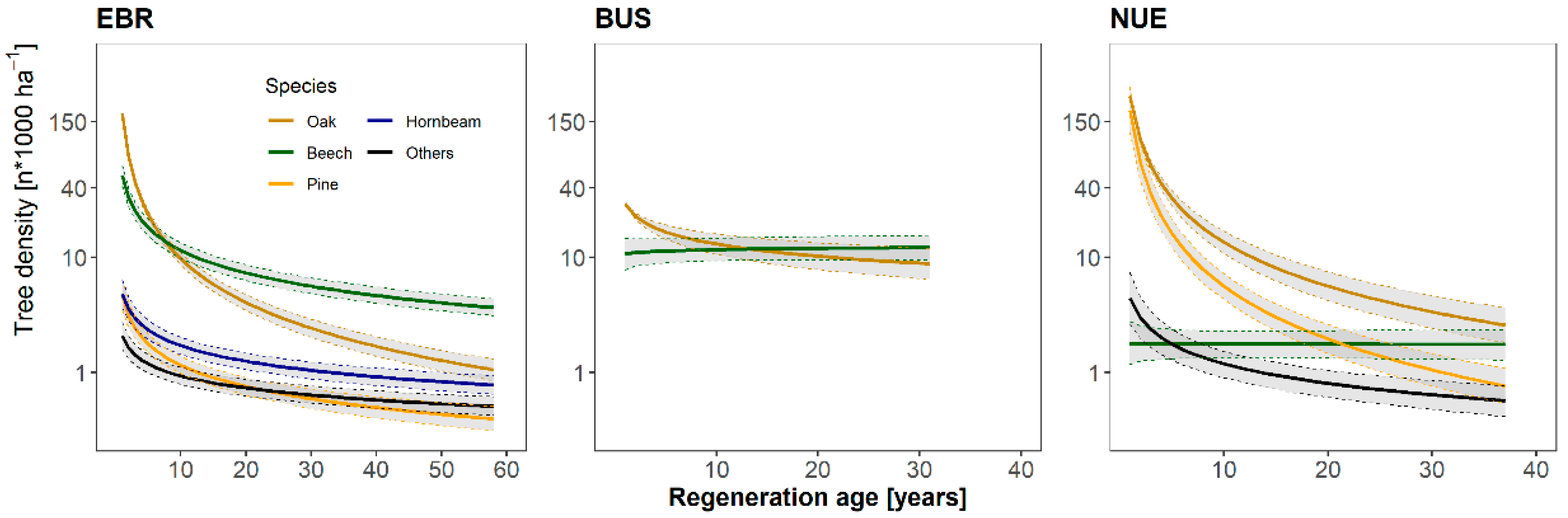
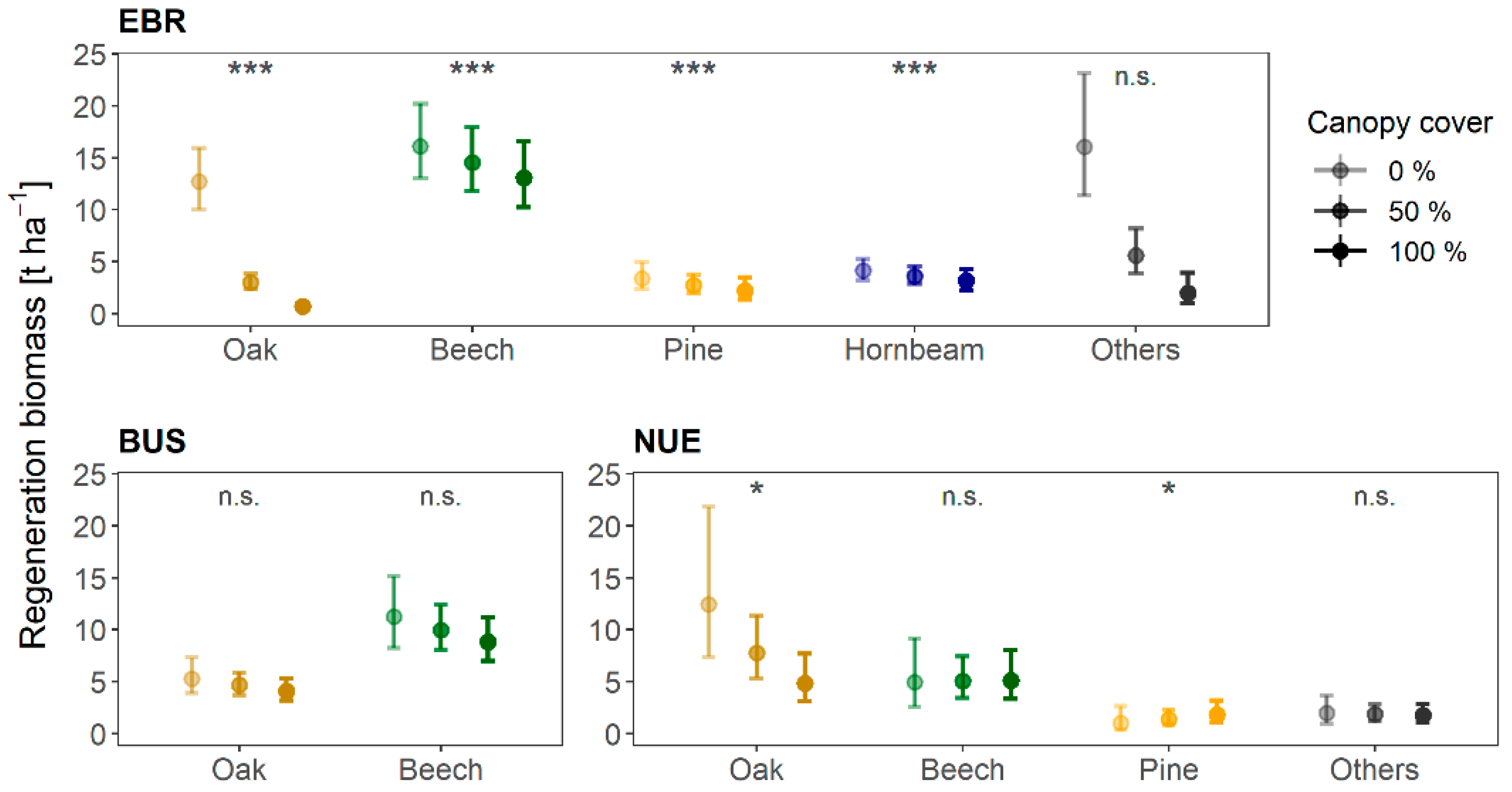
3.2. Regeneration Density and the Effect of Canopy Cover on Regeneration Biomass
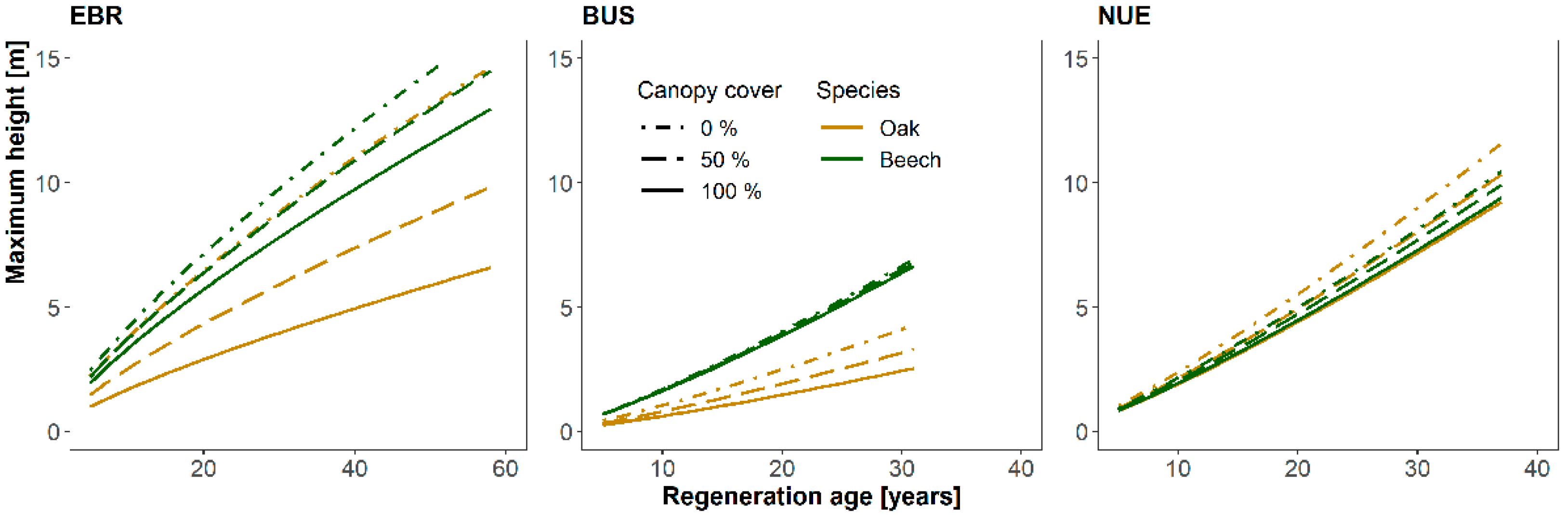
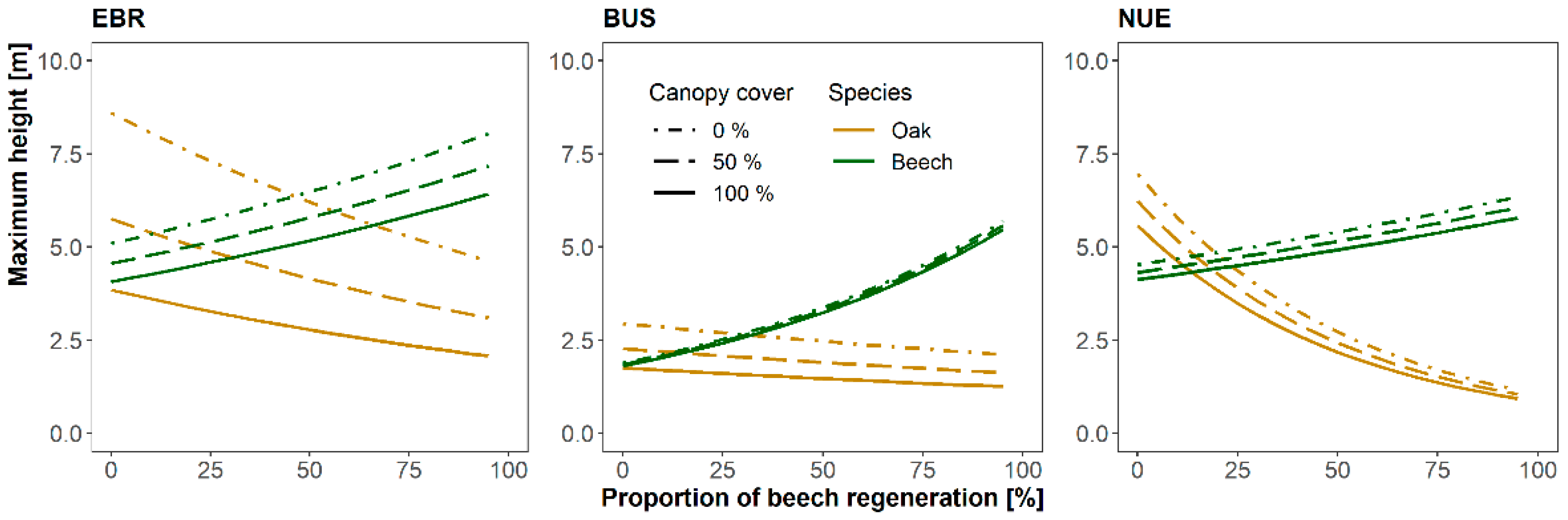
3.3. Influence of Canopy Cover and Interspecific Competition on Oaks’ Maximum Heights
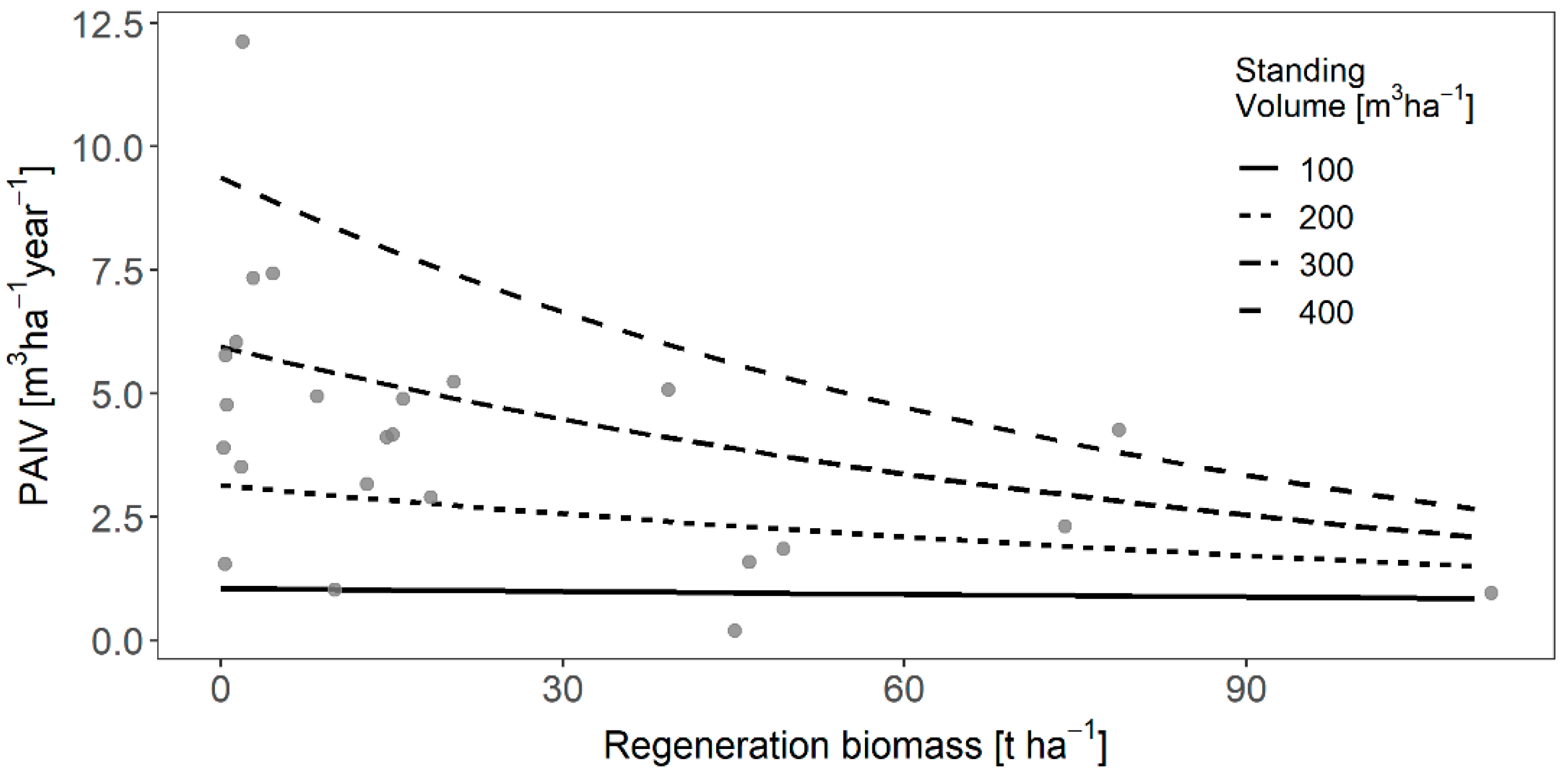
3.4. Feedback of Regeneration on Main Stand Productivity
4. Discussion
4.1. Valuable Insights from Long-Term Observations as Research Basis
4.2. Long-Term Development and Survival of the Regeneration
4.3. Influence of Canopy Cover and Interspecific Competition
4.4. Influence of Regeneration on Old-Growth Productivity
4.5. Silvicultural Consequences
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eaton, E.; Caudullo, G.; Oliveira, S.; de Rigo, D. Quercus robur and Quercus petraea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; pp. 160–163. ISBN 978-92-79-52833-0. [Google Scholar]
- Mette, T.; Dolos, K.; Meinardus, C.; Bräuning, A.; Reineking, B.; Blaschke, M.; Pretzsch, H.; Beierkuhnlein, C.; Gohlke, A.; Wellstein, C. Climatic turning point for beech and oak under climate change in Central Europe. Ecosphere 2013, 4, 1–19. [Google Scholar] [CrossRef]
- Mette, T.; Brandl, S.; Kölling, C. Climate Analogues for Temperate European Forests to Raise Silvicultural Evidence Using Twin Regions. Sustainability 2021, 13, 6522. [Google Scholar] [CrossRef]
- Schroeder, H.; Nosenko, T.; Ghirardo, A.; Fladung, M.; Schnitzler, J.-P.; Kersten, B. Oaks as Beacons of Hope for Threatened Mixed Forests in Central Europe. Front. For. Glob. Chang. 2021, 4, 78. [Google Scholar] [CrossRef]
- Albert, M.; Nagel, R.-V.; Nuske, R.; Sutmöller, J.; Spellmann, H. Tree Species Selection in the Face of Drought Risk—Uncertainty in Forest Planning. Forests 2017, 8, 363. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Bielak, K.; Block, J.; Bruchwald, A.; Dieler, J.; Ehrhart, H.-P.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zasada, M.; et al. Productivity of mixed versus pure stands of oak (Quercus petraea (Matt.) Liebl. and Quercus robur L.) and European beech (Fagus sylvatica L.) along an ecological gradient. Eur. J. For. Res. 2013, 132, 263–280. [Google Scholar] [CrossRef]
- Ammer, C. Unraveling the Importance of Inter- and Intraspecific Competition for the Adaptation of Forests to Climate Change. In Progress in Botany Vol. 78; Cánovas, F.M., Lüttge, U., Matyssek, R., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 345–367. ISBN 978-3-319-49489-0. [Google Scholar]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.-J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive forest management in central Europe: Climate change impacts, strategies and integrative concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Puettmann, K.J.; Messier, C. Simple Guidelines to Prepare Forests for Global Change: The Dog and the Frisbee. NWSC 2019, 93, 209. [Google Scholar] [CrossRef]
- Brändle, M.; Brandl, R. Species richness of insects and mites on trees: Expanding Southwood. J. Anim. Ecol. 2001, 70, 491–504. [Google Scholar] [CrossRef]
- Manos, P.S.; Stanford, A.M. The Historical Biogeography of Fagaceae: Tracking the Tertiary History of Temperate and Subtropical Forests of the Northern Hemisphere. Int. J. Plant Sci. 2001, 162, S77–S93. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef] [Green Version]
- Löf, M.; Brunet, J.; Filyushkina, A.; Lindbladh, M.; Skovsgaard, J.P.; Felton, A. Management of oak forests: Striking a balance between timber production, biodiversity and cultural services. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2016, 12, 59–73. [Google Scholar] [CrossRef] [Green Version]
- van der Plas, F.; Ratcliffe, S.; Ruiz-Benito, P.; Scherer-Lorenzen, M.; Verheyen, K.; Wirth, C.; Zavala, M.A.; Ampoorter, E.; Baeten, L.; Barbaro, L.; et al. Continental mapping of forest ecosystem functions reveals a high but unrealised potential for forest multifunctionality. Ecol. Lett. 2018, 21, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, G.T.; Spiecker, H. Silviculture of high-quality oaks: Questions and future research needs. Ann. Sci. 1993, 50, 531–534. [Google Scholar] [CrossRef] [Green Version]
- Attocchi, G. Silviculture of Oak for High-Quality Wood Production: Effects of Thinning on Crown size, Volume Growth and Stem Quality in Even-Aged Stands of Pedunculate oak (Quercus robur L.) in Northern Europe; Southern Swedish Forest Research Centre, Swedish University of Agricultural Sciences: Alnarp, Sweden, 2015; ISBN 978-91-576-8277-2. [Google Scholar]
- von Lüpke, B. Silvicultural methods of oak regeneration with special respect to shade tolerant mixed species. For. Ecol. Manag. 1998, 106, 19–26. [Google Scholar] [CrossRef]
- Mölder, A.; Sennhenn-Reulen, H.; Fischer, C.; Rumpf, H.; Schönfelder, E.; Stockmann, J.; Nagel, R.-V. Success factors for high-quality oak forest (Quercus robur, Q. petraea) regeneration. For. Ecosyst. 2019, 6, 262. [Google Scholar] [CrossRef] [Green Version]
- Ellenberg, H.H.; Strutt, G.K. Vegetation Ecology of Central Europe; Cambridge University Press: Cambridge, UK, 1988; ISBN 9780521236423. [Google Scholar]
- Manthey, M.; Leuschner, C.; Härdtle, W. Buchenwälder und Klimawandel. Nat. Landsch. 2007, 82, 441–445. [Google Scholar]
- Aas, G. Quercus petraea (Matt.) Liebl., Traubeneiche. In Enzyklopädie der Holzgewächse; Roloff, A., Weisgerber, H., Lang, U., Stimm, B., Eds.; Wiley-VCH: Weinheim, Germany, 2000. [Google Scholar]
- Krahl-Urban, J. Die Eichen: Forstliche Monographie der Traubeneiche und der Stieleiche; P. Parey: Hamburg, Germany, 1959. [Google Scholar]
- Mölder, A.; Meyer, P.; Nagel, R.-V. Integrative management to sustain biodiversity and ecological continuity in Central European temperate oak (Quercus robur, Q. petraea) forests: An overview. For. Ecol. Manag. 2019, 437, 324–339. [Google Scholar] [CrossRef]
- Fleder, W. Vom unterfränkischen Verjüngungsbetrieb. Allg. Forstz. 1983, 38, 1013–1014. [Google Scholar]
- Jedicke, E.; Hakes, W. Management von Eichenwäldern im Rahmen der FFH-Richtlinie. Nat. Landsch. 2005, 37, 37–45. [Google Scholar]
- Meyer, P. Forstwirtschaft und Naturschutz—Konfliktpotenzial und Synergien am Beispiel von Natura 2000. In Natura 2000 im Wald—Lebensraumtypen, Erhaltungszustand, Management, Naturschutz und Biologische Vielfalt; Lehrke, S., Ellwanger, G., Buschmann, A., Frederking, W., Paulsch, C., Schröder, E., Ssymank, A., Eds.; Landwirtschaftsverlag: Münster, Germany, 2013; pp. 177–197. [Google Scholar]
- Brang, P.; Spathelf, P.; Larsen, J.B.; Bauhus, J.; Boncina, A.; Chauvin, C.; Drossler, L.; Garcia-Guemes, C.; Heiri, C.; Kerr, G.; et al. Suitability of close-to-nature silviculture for adapting temperate European forests to climate change. Forestry 2014, 87, 492–503. [Google Scholar] [CrossRef] [Green Version]
- Puettmann, K.J.; Wilson, S.M.; Baker, S.C.; Donoso, P.J.; Drössler, L.; Amente, G.; Harvey, B.D.; Knoke, T.; Lu, Y.; Nocentini, S.; et al. Silvicultural alternatives to conventional even-aged forest management—What limits global adoption? For. Ecosyst. 2015, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Spathelf, P.; Bolte, A.; van der Maaten, E.C.D. Is Close-to-Nature Silviculture (CNS) an adequate concept to adapt forests to climate change? Landbauforsch.-Appl. Agric. For. Res. 2015, 65, 161–170. [Google Scholar] [CrossRef]
- Maleki, K.; Zeller, L.; Pretzsch, H. Oak often needs to be promoted in mixed beech-oak stands—The structural processes behind competition and silvicultural management in mixed stands of European beech and sessile oak. iForest 2020, 13, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Mosandl, R.; Abt, A. Waldbauverfahren in Eichenwäldern gestern und heute. AFZ-Der Wald 2016, 20, 28–32. [Google Scholar]
- Březina, I.; Dobrovolný, L. Natural regeneration of sessile oak under different light conditions. J. For. Sci. 2011, 57, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Ligot, G.; Balandier, P.; Fayolle, A.; Lejeune, P.; Claessens, H. Height competition between Quercus petraea and Fagus sylvatica natural regeneration in mixed and uneven-aged stands. For. Ecol. Manag. 2013, 304, 391–398. [Google Scholar] [CrossRef]
- Modrow, T.; Kuehne, C.; Saha, S.; Bauhus, J.; Pyttel, P.L. Photosynthetic performance, height growth, and dominance of naturally regenerated sessile oak (Quercus petraea [Mattuschka] Liebl.) seedlings in small-scale canopy openings of varying sizes. Eur. J. For. Res. 2019, 116, 346. [Google Scholar] [CrossRef]
- Annighöfer, P.; Beckschäfer, P.; Vor, T.; Ammer, C. Regeneration patterns of European oak species (Quercus petraea (Matt.) Liebl., Quercus robur L.) in dependence of environment and neighborhood. PLoS ONE 2015, 10, e0134935. [Google Scholar] [CrossRef]
- Kanjevac, B.; Krstić, M.; Babić, V.; Govedar, Z. Regeneration Dynamics and Development of Seedlings in Sessile Oak Forests in Relation to the Light Availability and Competing Vegetation. Forests 2021, 12, 384. [Google Scholar] [CrossRef]
- Kohler, M.; Pyttel, P.; Kuehne, C.; Modrow, T.; Bauhus, J. On the knowns and unknowns of natural regeneration of silviculturally managed sessile oak (Quercus petraea (Matt.) Liebl.) forests—A literature review. Ann. For. Sci. 2020, 77, 101. [Google Scholar] [CrossRef]
- Kuehne, C.; Pyttel, P.; Modrow, T.; Kohnle, U.; Bauhus, J. Seedling development and regeneration success after 10 years following group selection harvesting in a sessile oak (Quercus petraea [Mattuschka] Liebl.) stand. Ann. For. Sci. 2020, 77, 71. [Google Scholar] [CrossRef]
- Löf, M. Establishment and growth in seedlings of Fagus sylvatica and Quercus robur: Influence of interference from herbaceous vegetation. Can. J. For. Res. 2000, 30, 855–864. [Google Scholar] [CrossRef]
- Löf, M.; Barrere, J.; Engman, M.; Petersson, L.K.; Villalobos, A. The influence of fencing on seedling establishment during reforestation of oak stands: A comparison of artificial and natural regeneration techniques including costs. Eur. J. For. Res. 2021, 140, 807–817. [Google Scholar] [CrossRef]
- Johann, K. DESER-Norm 1993. Normen der Sektion Ertragskunde im Deutschen Verband Forstlicher Forschungsanstalten zur Aufbereitung von waldwirtschaftlichen Dauerversuchen. Ber. Jahrestag. Dtsch. Verb. Forstl. Sekt. Ertragskunde 1993, 96–104. [Google Scholar]
- Forrester, D.I.; Tachauer, I.; Annighoefer, P.; Barbeito, I.; Pretzsch, H.; Ruiz-Peinado, R.; Stark, H.; Vacchiano, G.; Zlatanov, T.; Chakraborty, T.; et al. Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate. For. Ecol. Manag. 2017, 396, 160–175. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Soft. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Petritan, A.M.; Bouriaud, O.; Frank, D.C.; Petritan, I.C. Dendroecological reconstruction of disturbance history of an old-growth mixed sessile oak-beech forest. J. Veg. Sci. 2017, 28, 117–127. [Google Scholar] [CrossRef]
- Dietz, L.; Gégout, J.-C.; Dupouey, J.-L.; Lacombe, E.; Laurent, L.; Collet, C. Beech and hornbeam dominate oak 20 years after the creation of storm-induced gaps. For. Ecol. Manag. 2022, 503, 119758. [Google Scholar] [CrossRef]
- Manso, R.; Ligot, G.; Fortin, M. A recruitment model for beech–oak pure and mixed stands in Belgium. For. Int. J. For. Res. 2020, 93, 124–132. [Google Scholar] [CrossRef]
- Vanselow, K. Die Waldbautechnik der Eiche im bayerischen Spessart in geschichtlicher Betrachtung. Forstwiss. Cent. 1960, 79, 270–286. [Google Scholar] [CrossRef]
- von Lüpke, B. Einfluss unterschiedlicher Hiebsformen auf die Naturverjüngung eines Traubeneichen-Buchen-Mischbestandes. Forstarchiv 2008, 79, 4–15. [Google Scholar]
- Leuschner, C.; Förster, A.; Diers, M.; Culmsee, H. Are northern German Scots pine plantations climate smart? The impact of large-scale conifer planting on climate, soil and the water cycle. For. Ecol. Manag. 2022, 507, 120013. [Google Scholar] [CrossRef]
- von Lüpke, B.; Hauskeller-Bullerjahn, K. Beitrag zur Modellierung der Jungwuchsentwicklung am Beispiel von Traubeneichen-Buchen-Mischverjüngungen. Allg. Forst. U J. Ztg 2004, 175, 61–69. [Google Scholar]
- Mölder, A.; Nagel, R.-V.; Meyer, P.; Schmidt, M.; Rumpf, H.; Spellmann, H. Historischer Rückblick auf die Verjüngung von Eichen im Spessart des 19. Jahrhunderts–Bedeutung der angewandten Verfahren für die heutige Eichenwirtschaft. Forstarchiv 2017, 88, 67–78. [Google Scholar]
- Hauskeller-Bullerjahn, K. Wachstum Junger Eichen Unter Schirm. Ph.D. Dissertation, Universität Göttingen, Göttingen, Germany, 1997. [Google Scholar]
- Skrzyszewski, J.; Pach, M. Crookedness of pedunculate oak (Quercus robur L.) growing under a canopy of Scots pine (Pinus sylvestris L.). Scand. J. For. Res. 2015, 30, 688–698. [Google Scholar] [CrossRef]
- Leuschner, C.; Hertel, D.; Coners, H.; Büttner, V. Root competition between beech and oak: A hypothesis. Oecologia 2001, 126, 276–284. [Google Scholar] [CrossRef]
- Meesenburg, H.; Schmidt, M.; Sutmöller, J.; Albert, M. Klimaanpassung ist Vorsorge für den Wald. proWald 2015, 11, 4–10. [Google Scholar]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dauber, E. Long-term stand dynamics of managed spruce–fir–beech mountain forests in Central Europe: Structure, productivity and regeneration success. Forestry 2015, 88, 407–428. [Google Scholar] [CrossRef]
- Knapp, E. Zur Wuchsleistung der Unterbaubuche im ungleichaltrigen Kiefern-Buchen-Mischbestand vor und nach ihrer Übernahme als Hauptbestand auf Standorten des nordostdeutschen Tieflandes. Ber. Jahrestag. 1991, 96–110. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp | Nr | n | Size | Comp 1 | RT 2 | Surveys | Per | Lat | Lon | Soil | P | T | Alt |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BUS | 136 | 2 | 0.5 | Oa-Be | nat | 1986, 1995, 2012 | 26 | 50.129 | 9.593 | Cambisol | 796 | 7.8 | 445 |
| EBR | 132 | 1 | 0.5 | Oa-Be-Pi | nat | 1982, 1993, 2019 | 36 | 49.836 | 10.547 | Cambisol | 683 | 8.1 | 338 |
| EBR | 133 | 5 | 1.0 | Oa-Be-(Pi) | nat/art | 1983, 1999, 2019 | 36 | 49.853 | 10.547 | Cambisol—(Pseudogley) | 675 | 7.9 | 385 |
| NUE | 141 | 4 | 0.1 | Pi | nat/art | 1991, 1998, 2019 | 28 | 49.499 | 11.144 | Cambisol/Pseudogley | 759 | 8.7 | 333 |
| First Survey | Last Survey | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exp (Nr) | Plot | S | Age | N | H100 | D100 | BA | V | CC | Age | N | H100 | D100 | BA | V | PAIBA | PAIV | CC | Per |
| EBR (132) | 1 | Pi | 147 | 21 | 30.9 | 53.3 | 4.65 | 66.14 | 37 | 184 | 12 | 33.27 | 65.01 | 4.15 | 63.15 | 0.05 | 0.76 | 30 | 36 |
| Be | 147 | 42 | 31.34 | 45.12 | 6.66 | 110.2 | 184 | 21 | 33.05 | 69.6 | 7.93 | 138.81 | 0.1 | 1.76 | |||||
| Oa | 147 | 4 | 31.59 | 46.9 | 0.72 | 12.26 | 184 | 4 | 32.78 | 67.4 | 1.49 | 26.99 | 0.02 | 0.38 | |||||
| total | 67 | 12.03 | 188.61 | 37 | 13.56 | 228.94 | 0.17 | 2.9 | |||||||||||
| EBR (133) | 2 | Pi | 143 | 8 | 27.03 | 51.64 | 1.66 | 20.98 | 60 | 178 | 2 | 32.07 | 52.2 | 0.39 | 5.79 | 0.01 | 0.11 | 43 | 36 |
| Be | 143 | 77 | 32.44 | 47.15 | 12.24 | 209.08 | 178 | 53 | 34.78 | 64.37 | 15.44 | 282.88 | 0.19 | 4.09 | |||||
| Oa | 143 | 14 | 29.02 | 46 | 2.33 | 36.71 | 178 | 13 | 32.65 | 62.1 | 3.94 | 71.18 | 0.05 | 1.05 | |||||
| total | 99 | 16.23 | 266.78 | 68 | 19.77 | 359.84 | 0.24 | 5.24 | |||||||||||
| 4 | Be | 164 | 44 | 34.88 | 49.35 | 8.42 | 159.51 | 22 | 199 | 33 | 35.1 | 70.11 | 12.74 | 237.99 | 0.16 | 3.35 | 43 | 36 | |
| Oa | 164 | 12 | 33.55 | 46.74 | 2.06 | 37.18 | 199 | 12 | 33.33 | 61.5 | 3.56 | 65.38 | 0.04 | 0.87 | |||||
| Hb | 164 | 1 | 24.38 | 30.7 | 0.07 | 0.92 | 199 | 1 | 28.44 | 42.5 | 0.14 | 2.11 | 0 | 0.04 | |||||
| total | 57 | 10.55 | 197.61 | 46 | 16.44 | 305.48 | 0.2 | 4.27 | |||||||||||
| 6 | Pi | 162 | 11 | 27.09 | 53.14 | 2.4 | 30.21 | 62 | 197 | 9 | 28.68 | 60.38 | 2.44 | 32.49 | 0.02 | 0.18 | 54 | 36 | |
| Be | 162 | 44 | 28.72 | 46.56 | 4.92 | 71.82 | 197 | 34 | 31.29 | 60.43 | 5.64 | 107.13 | 0.07 | 1.51 | |||||
| Oa | 162 | 67 | 28.67 | 43.7 | 9.46 | 145.15 | 197 | 62 | 30.91 | 54.69 | 14.58 | 241.67 | 0.15 | 3.33 | |||||
| Hb | 162 | 10 | 23.53 | 22.54 | 0.24 | 1.81 | 197 | 10 | 20.37 | 28.98 | 0.49 | 4.73 | 0 | 0.07 | |||||
| total | 132 | 17.02 | 249 | 115 | 23.98 | 386.03 | 0.24 | 5.08 | |||||||||||
| 7 | Sp | 146 | 1 | 27.55 | 47 | 0.17 | 2.18 | 22 | 181 | 16 | 28.5 | 50.85 | 3.25 | 42.9 | 0.03 | 0.55 | 30 | 36 | |
| Pi | 146 | 24 | 27.02 | 41.36 | 3.22 | 40.31 | 0 | - | - | - | - | - | - | ||||||
| Be | 146 | 25 | 30.12 | 44.64 | 3.91 | 62.53 | 181 | 19 | 27.76 | 58.43 | 5.09 | 74.79 | 0.06 | 0.72 | |||||
| Oa | 146 | 18 | 25.97 | 40.38 | 2.3 | 32.22 | 181 | 17 | 26.44 | 57.23 | 4.37 | 64.48 | 0.06 | 1.04 | |||||
| total | 68 | 9.61 | 137.25 | 52 | 12.72 | 182.17 | 0.15 | 2.31 | |||||||||||
| 8 | Be | 153 | 13 | 32.81 | 48.84 | 2.44 | 42.33 | 25 | 188 | 4 | 26.21 | 64.18 | 1.29 | 18.11 | 0.01 | 0.11 | 9 | 36 | |
| Oa | 153 | 19 | 29.34 | 41.73 | 2.6 | 40.89 | 188 | 15 | 27.67 | 55.3 | 3.6 | 55.22 | 0.05 | 0.85 | |||||
| Hb | 153 | 1 | 21.72 | 18 | 0.03 | 0.26 | 0 | - | - | - | - | - | - | ||||||
| total | 33 | 5.06 | 83.48 | 19 | 4.9 | 73.33 | 0.06 | 0.96 | |||||||||||
| BUS (136) | 1 | Be | 192 | 40 | 24.97 | 43.52 | 2.52 | 30.58 | 67 | 218 | 4 | 27.01 | 39.6 | 0.42 | 5.52 | 0.02 | 0.3 | 38 | 26 |
| Oa | 192 | 66 | 29.31 | 60.12 | 18.74 | 305.77 | 218 | 52 | 31.34 | 70.58 | 20.34 | 356.57 | 0.19 | 3.82 | |||||
| total | 106 | 21.26 | 336.35 | 56 | 20.76 | 362.09 | 0.21 | 4.11 | |||||||||||
| 2 | Be | 202 | 106 | 28.08 | 44.76 | 7.03 | 88.29 | 67 | 228 | 26 | 27.78 | 49.97 | 3.78 | 53.8 | 0.08 | 0.94 | 41 | 26 | |
| Oa | 202 | 74 | 28.18 | 61 | 21.63 | 340.19 | 228 | 48 | 30.92 | 68.54 | 17.71 | 305.72 | 0.16 | 3.24 | |||||
| total | 180 | 28.66 | 428.48 | 74 | 21.48 | 359.52 | 0.24 | 4.17 | |||||||||||
| NUE (141) | 1 | Pi | 88 | 400 | 24.1 | 35.76 | 27.07 | 287.45 | 66 | 116 | 144 | 26.77 | 39.99 | 16.69 | 203.99 | 0.23 | 3.52 | 35 | 28 |
| 2 | Pi | 97 | 400 | 25.52 | 35.95 | 29.53 | 332.25 | 77 | 125 | 233 | 30 | 43.83 | 29.25 | 389.24 | 0.45 | 7.44 | 57 | 28 | |
| 3 | Pi | 125 | 189 | 27.57 | 47.56 | 27.62 | 346.41 | 63 | 153 | 111 | 30.19 | 52.67 | 23.59 | 327.32 | 0.29 | 4.95 | 42 | 28 | |
| 4 | Pi | 130 | 133 | 31.65 | 48.8 | 22.46 | 324.85 | 57 | 158 | 44 | 29.79 | 54.33 | 10.3 | 141.56 | 0.1 | 1.03 | 23 | 28 | |
| First Survey | Last Survey | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Exp (Nr) | Plot | S | A | d | bm | h | A | d | bm | h |
| EBR (132) | 1 | Pi | 4 | 3504 | 182 | 0.67 | 40 | 4 | 202 | 16.07 |
| Be | 18,308 | 2033 | 1.05 | 3492 | 55,745 | 7.41 | ||||
| Oa | 22,104 | 562 | 1.07 | 271 | 11,015 | 13.45 | ||||
| Hb | 3383 | 605 | 1.21 | 354 | 9235 | 7.99 | ||||
| Others | 350 | 33 | 1.55 | 4 | 26 | 8.19 | ||||
| EBR (133) | 2 | Pi | 6 | 366 | 22 | 0.89 | 42 | 0 | - | - |
| Be | 15,464 | 1767 | 1.11 | 3709 | 27,696 | 7.1 | ||||
| Oa | 8892 | 165 | 0.87 | 5 | 68 | 11.48 | ||||
| Hb | 130 | 16 | 1.74 | 6 | 716 | 15.48 | ||||
| Others | 697 | 118 | 2.85 | 59 | 3305 | 12.06 | ||||
| 4 | Pi | 22 | 6 | 0 | 0.62 | 58 | 0 | - | - | |
| Be | 30,554 | 6100 | 3.19 | 2394 | 91,381 | 11.4 | ||||
| Oa | 1 140 | 15 | 1.44 | 6 | 279 | 13.97 | ||||
| Hb | 197 | 28 | 1.64 | 6 | 1038 | 18 | ||||
| Others | 284 | 60 | 4.71 | 98 | 32,299 | 19.89 | ||||
| 6 | Pi | 6 | 100 | 4 | 0.36 | 42 | 5 | 125 | 9.27 | |
| Be | 15,791 | 1056 | 0.65 | 5280 | 49,307 | 5.72 | ||||
| Oa | 119,009 | 833 | 0.32 | 98 | 921 | 7.12 | ||||
| Hb | 69 | 9 | 0.87 | 95 | 2033 | 9.87 | ||||
| Others | 153 | 15 | 1.07 | 77 | 8714 | 10.21 | ||||
| 7 | Pi | 11 | 2198 | 161 | 1.44 | 47 | 59 | 4672 | 13.24 | |
| Be | 19,284 | 2863 | 1.71 | 3972 | 49,136 | 7.13 | ||||
| Oa | 17,402 | 479 | 1.84 | 613 | 43,182 | 14.41 | ||||
| Hb | 2925 | 594 | 1.67 | 453 | 7973 | 9.51 | ||||
| Others | 3609 | 477 | 2.03 | 184 | 9655 | 9.54 | ||||
| 8 | Pi | 13 | 70 | 5 | 1.36 | 49 | 16 | 2340 | 16.55 | |
| Be | 7634 | 891 | 1.53 | 3203 | 29,103 | 5.72 | ||||
| Oa | 41,570 | 1483 | 1.89 | 2500 | 102,447 | 10.07 | ||||
| Hb | 3642 | 897 | 2.14 | 1553 | 33,505 | 7.81 | ||||
| Others | 219 | 27 | 2.37 | 22 | 6699 | 14.86 | ||||
| BUS (136) | 1 | Be | 5 | 15,878 | 1052 | 0.67 | 31 | 11,628 | 12,644 | 2.8 |
| Oa | 54,286 | 1686 | 0.3 | 6678 | 10,142 | 3.52 | ||||
| 2 | Be | 3 | 12,245 | 812 | 0.64 | 29 | 8175 | 23,505 | 3.9 | |
| Oa | 13,302 | 411 | 0.25 | 5 | 26 | 7.13 | ||||
| NUE (141) | 1 | Pi | 3 | 31,911 | 849 | 0.3 | 31 | 2133 | 2753 | 2.77 |
| Be | 1278 | 130 | 0.32 | 1478 | 8653 | 6.35 | ||||
| Oa | 18,889 | 785 | 0.45 | 2444 | 6696 | 3.1 | ||||
| Others | 544 | 162 | 0.47 | 356 | 1714 | 4.67 | ||||
| 2 | Pi | 5 | 86,967 | 2429 | 0.32 | 33 | 122 | 92 | 2.81 | |
| Be | 1122 | 129 | 0.43 | 3567 | 46,948 | 8.17 | ||||
| Oa | 42,911 | 1742 | 0.53 | 822 | 3120 | 5.09 | ||||
| Others | 156 | 47 | 0.38 | 222 | 950 | 6.14 | ||||
| 3 | Pi | 9 | 2300 | 157 | 0.88 | 37 | 0 | - | - | |
| Be | 5011 | 1095 | 1.75 | 2622 | 76,353 | 10.96 | ||||
| Oa | 40,211 | 4230 | 1.56 | 89 | 3900 | 14.34 | ||||
| Others | 7089 | 3060 | 1.22 | 322 | 13,313 | 10.41 | ||||
| 4 | Pi | 9 | 1800 | 128 | 0.89 | 37 | 0 | - | - | |
| Be | 322 | 54 | 0.94 | 500 | 3888 | 3.58 | ||||
| Oa | 23,911 | 2517 | 1.54 | 1733 | 100,861 | 11.58 | ||||
| Others | 1633 | 670 | 1.22 | 256 | 6497 | 6.86 | ||||
| EBR | BUS | NUE | ||||
|---|---|---|---|---|---|---|
| Fixed Effects | Est (SE) | p | Est (SE) | p | Est (SE) | p |
| (Intercept) | 12.09 (0.16) | <0.001 | 10.28 (0.28) | <0.001 | 12.43 (0.25) | <0.001 |
| ln (A) | −1.26 (0.03) | <0.001 | −0.35 (0.05) | <0.001 | −1.27 (0.07) | <0.001 |
| Be | −1.24 (0.12) | <0.001 | −1.00 (0.17) | <0.001 | −4.94 (0.26) | <0.001 |
| Be × ln (A) | 0.61 (0.04) | <0.001 | 0.39 (0.10) | <0.001 | 1.26 (0.10) | <0.001 |
| Pi | −3.63 (0.17) | <0.001 | - | - | −0.28 (0.2) | 0.316 |
| Pi × ln (A) | 0.65 (0.06) | <0.001 | - | - | −0.26 (0.11) | 0.020 |
| Hb | −3.61 (0.15) | <0.001 | - | - | - | - |
| Hb × ln (A) | 0.81 (0.05) | <0.001 | - | - | - | - |
| Others | −4.45 (0.14) | <0.001 | - | - | −4.04 (0.30) | <0.001 |
| Others × ln (A) | 0.92 (0.05) | <0.001 | - | - | 0.70 (0.11) | <0.001 |
| Random Effects | ||||||
| σ2 | 1.16 | 1.55 | 1.00 | |||
| τ00 Squ:P | 0.03 | 0.33 | 0.04 | |||
| τ00 P | 0.11 | 0.12 | 0.11 | |||
| N Squ | 256 | 200 | 36 | |||
| N P | 6 | 2 | 4 | |||
| N measurements | 10,599 | 2101 | 1405 | |||
| AIC | 31,971.7 | 7215.1 | 4084.5 | |||
| EBR | BUS | NUE | ||||
|---|---|---|---|---|---|---|
| Fixed Effects | Est (SE) | p | Est (SE) | p | Est (SE) | p |
| (Intercept) | −3.71 (0.29) | <0.001 | −3.99 (0.34) | <0.001 | −3.04 (0.47) | <0.001 |
| ln (A) | 2.11 (0.08) | <0.001 | 1.93 (0.10) | <0.001 | 1.90 (0.12) | <0.001 |
| CC | −2.90 (0.13) | <0.001 | −0.26 (0.19) | 0.168 | −0.95 (0.38) | 0.013 |
| Be | 0.24 (0.08) | 0.002 | 0.76 (0.17) | <0.001 | −0.93 (0.38) | 0.015 |
| Be × CC | 2.69 (0.15) | <0.001 | 0.01 (0.24) | 0.955 | 0.99 (0.54) | 0.065 |
| Pi | −1.32 (0.17) | <0.001 | - | - | −2.51 (0.55) | <0.001 |
| Pi × CC | 2.46 (0.32) | <0.001 | - | - | 1.53 (0.75) | 0.041 |
| Hb | −1.13 (0.09) | <0.001 | - | - | - | - |
| Hb × CC | 2.64 (0.21) | <0.001 | - | - | - | - |
| Others | 0.22 (0.17) | 0.194 | - | - | −1.85 (0.40) | <0.001 |
| Others × CC | 0.81 (0.42) | 0.056 | - | - | 0.84 (0.58) | 0.144 |
| Random Effects | ||||||
| σ2 | 2.61 | 2.15 | 2.50 | |||
| τ00 Squ:P | 0.10 | 0.18 | 0.00 | |||
| τ00 P | 0.07 | 0.01 | 0.09 | |||
| N Squ | 256 | 200 | 36 | |||
| N P | 6 | 2 | 4 | |||
| N measurements | 5195 | 1307 | 772 | |||
| AIC | 19,983.3 | 4829.5 | 2931.8 | |||
| EBR | BUS | NUE | ||||
|---|---|---|---|---|---|---|
| Fixed Effects | Est (SE) | p | Est (SE) | p | Est (SE) | P |
| (Intercept) | −0.20 (0.10) | 0.251 | −2.60 (0.30) | <0.001 | −1.70 (0.40) | <0.001 |
| ln (A) | 0.77 (0.00) | <0.001 | 1.24 (0.00) | <0.001 | 1.20 (0.10) | <0.001 |
| CC | −0.80 (0.10) | <0.001 | −0.50 (0.10) | <0.001 | −0.20 (0.30) | 0.387 |
| Be_perc | −0.70 (0.10) | <0.001 | −0.40 (0.10) | 0.001 | −1.90 (0.70) | 0.008 |
| Be | −0.50 (0.00) | <0.001 | −0.50 (0.10) | <0.001 | −0.40 (0.30) | 0.077 |
| Be × Be_perc | 1.13 (0.10) | <0.001 | 1.52 (0.10) | <0.001 | 2.25 (0.70) | 0.001 |
| Be × CC | 0.58 (0.10) | <0.001 | 0.48 (0.10) | <0.001 | 0.12 (0.30) | 0.703 |
| Pi | 0.01 (0.10) | 0.932 | - | - | −0.80 (0.40) | 0.052 |
| Pi × Be_perc | −0.20 (0.20) | 0.546 | - | - | 0.15 (1.00) | 0.880 |
| Pi × CC | 0.69 (0.20) | <0.001 | - | - | −0.20 (0.50) | 0.631 |
| Hb | −0.50 (0.10) | <0.001 | - | - | - | - |
| Hb × Be_perc | 0.66 (0.10) | <0.001 | - | - | - | - |
| Hb × CC | 0.66 (0.10) | <0.001 | - | - | - | - |
| Others | −0.10 (0.10) | 0.147 | - | - | −1.10 (0.30) | <0.001 |
| Others × Be_perc | 0.43 (0.20) | 0.010 | - | - | 0.90 (0.80) | 0.242 |
| Others × CC | 0.12 (0.20) | 0.425 | - | - | 0.33 (0.40) | 0.354 |
| Random Effects | ||||||
| σ2 | 0.27 | 0.33 | 0.53 | |||
| τ00 Squ:P | 0.03 | 0.07 | 0.06 | |||
| τ00 P | 0.03 | 0.18 | 0.06 | |||
| N Squ | 256 | 200 | 36 | |||
| N P | 6 | 2 | 4 | |||
| N measurements | 3519 | 1266 | 464 | |||
| AIC | 5752.0 | 2423.0 | 1103.3 | |||
| Fixed Effects | Est (SE) | p |
|---|---|---|
| (Intercept) | −7.23 (1.79) | <0.001 |
| bm_reg | 0.03 (0.03) | 0.358 |
| ln (V) | 1.58 (0.33) | <0.001 |
| ln (V) × bm_reg | −0.01 (0.01) | 0.279 |
| Random Effects | ||
| σ2 | 0.23 | |
| τ00 P | 0 | |
| N P | 7 | |
| N measurements | 24 | |
| AIC | 62.5 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stimm, K.; Uhl, E.; Pretzsch, H. Chances and Limitations of Mixed Oak Regeneration under Continuous Canopy Cover—Evidence from Long-Term Observations. Forests 2022, 13, 2052. https://doi.org/10.3390/f13122052
Stimm K, Uhl E, Pretzsch H. Chances and Limitations of Mixed Oak Regeneration under Continuous Canopy Cover—Evidence from Long-Term Observations. Forests. 2022; 13(12):2052. https://doi.org/10.3390/f13122052
Chicago/Turabian StyleStimm, Kilian, Enno Uhl, and Hans Pretzsch. 2022. "Chances and Limitations of Mixed Oak Regeneration under Continuous Canopy Cover—Evidence from Long-Term Observations" Forests 13, no. 12: 2052. https://doi.org/10.3390/f13122052