
Sooty Bark Disease in Sycamore: Seasonal and Vertical Variation in Spore Release of Cryptostroma corticale
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
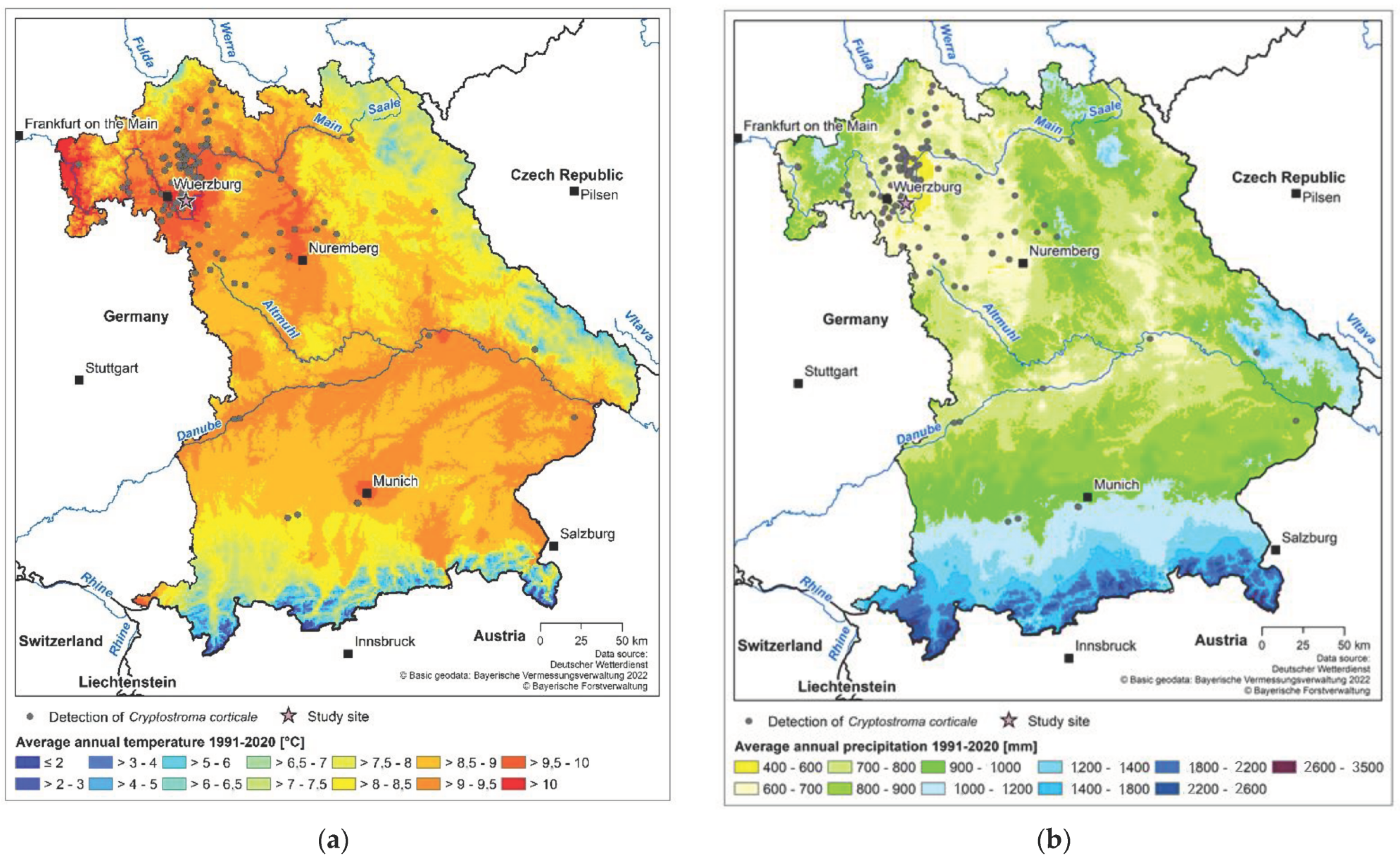
2.1. Study Site
2.2. Detection of C. corticale
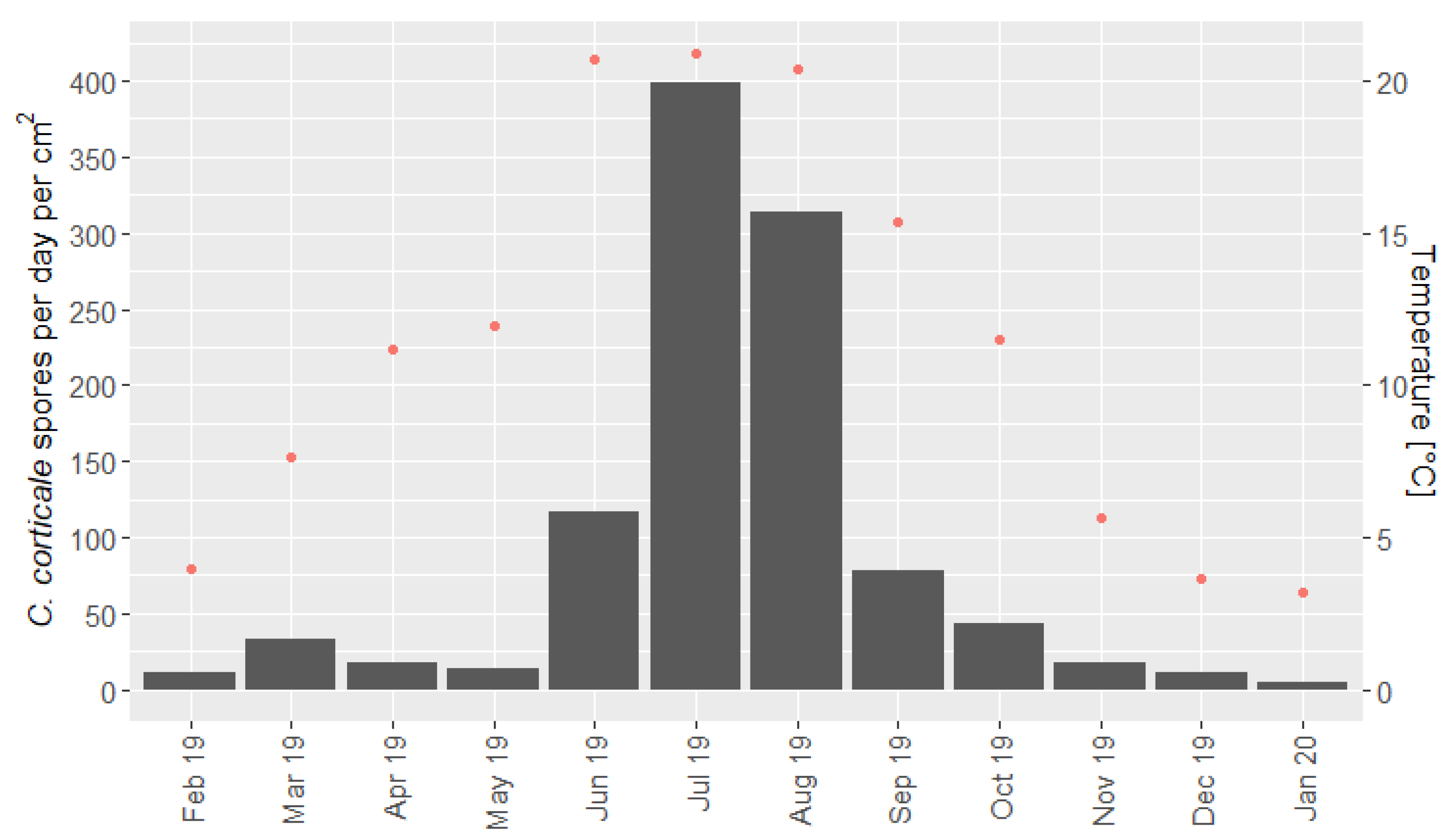
2.3. Spore Release According to Season
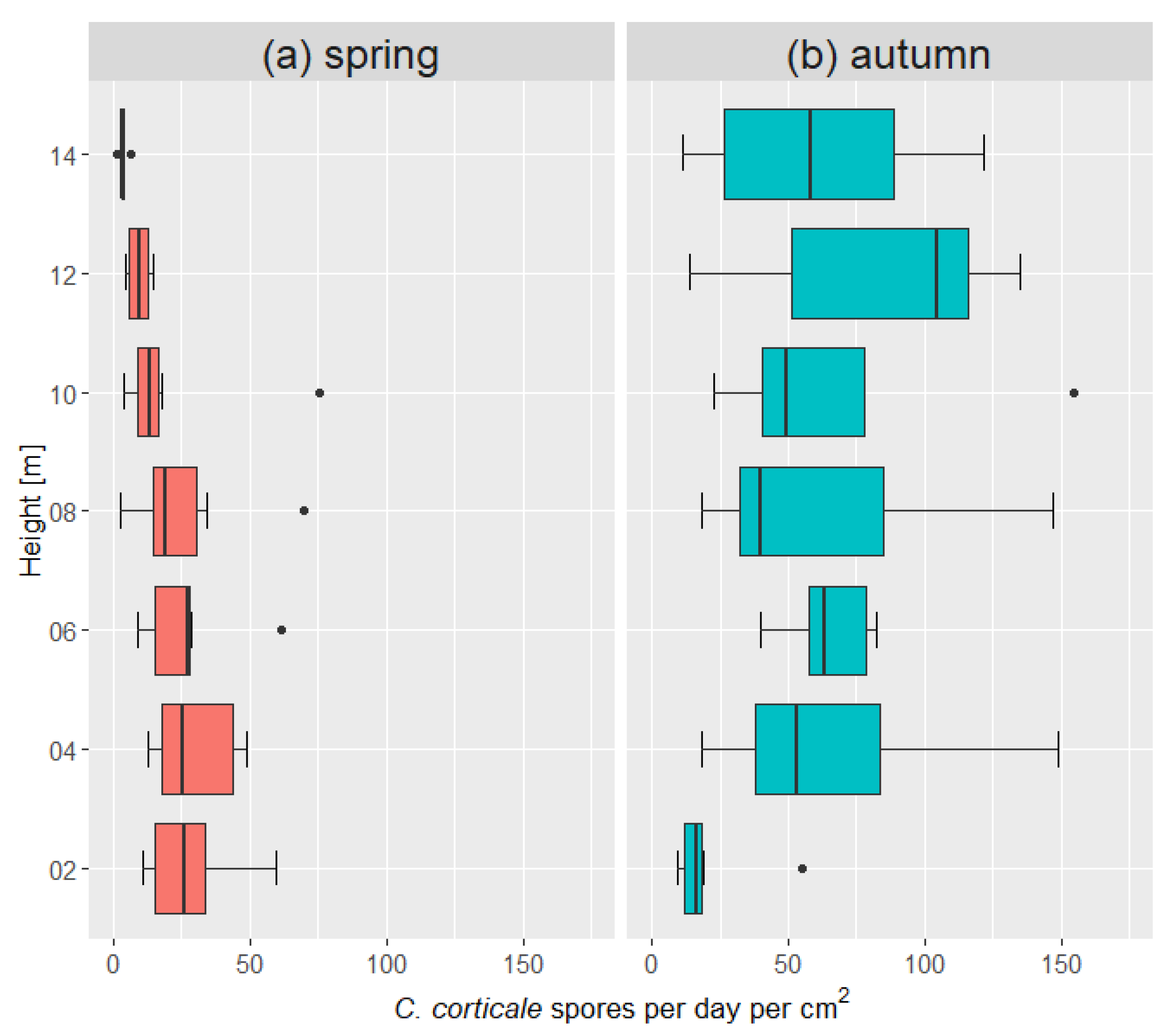
2.4. Spore Release along Vertical Gradient
2.5. Counting Spores by Microscopy
3. Results
3.1. Detection of C. corticale
3.2. Spore Release According to Season
3.3. Spore Release along Vertical Gradient
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gregory, P.H.; Waller, S. Cryptostroma corticale and sooty bark disease of sycamore (Acer pseudoplatanus). Mycol. Res. 1951, 34, 579–597. [Google Scholar] [CrossRef]
- Ellis, J.B.; Everhart, B.M. New species of hyphomycetes fungi. J. Mycol. 1889, 5, 68–72. [Google Scholar] [CrossRef]
- Gregory, P.H.; Peace, T.R.; Waller, S. Death of sycamore trees associated with an unidentified fungus. Nature 1949, 164, 275. [Google Scholar] [CrossRef] [Green Version]
- Peace, T.R. Sooty bark disease of sycamore. A disease in eclipse. Q. J. For. 1955, 49, 197–204. [Google Scholar]
- Young, C.W.T. Sooty Bark Disease of Sycamore. Arboric. Leafl. 1978, 3, 8. [Google Scholar]
- Gibbs, J.N. Fifty years of sooty bark disease of sycamore. Q. J. For. 1997, 91, 215–221. [Google Scholar]
- Dickenson, S.; Wheeler, B.E.J. Effects of temperature, and water stress in sycamore, on growth of Cryptostroma corticale. Trans. Br. Mycol. Soc. 1981, 76, 181–185. [Google Scholar] [CrossRef]
- Moreau, C.; Moreau, M. La suie des Sycamores a Paris. Bull. La Société Mycol. Fr. 1951, 67, 404–418. [Google Scholar]
- Plate, H.-P.; Schneider, R. Ein Fall von asthmaartiger Allergie, verursacht von Cryptostroma corticale. Nachr. Des Dtsch. Pflanzenschutzd. 1965, 17, 100–101. [Google Scholar]
- Metzler, B. Cryptostroma corticale an Berg-Ahorn nach dem Trockenjahr 2003. Mitt. Biol. Bundesanst. Land Forstwirtsch. 2006, 400, 161–162. [Google Scholar]
- Kehr, R. Neue Krankheiten an Platane, Linde und Ahorn. Jahrb. Baumpflege 2007, 2007, 144–156. [Google Scholar]
- Robeck, P.; Heinrich, R.; Schumacher, J.; Feindt, R.; Kehr, R. Status der Rußrindenkrankheit des Ahorns in Deutschland. Jahrb. Baumpflege 2008, 2008, 238–245. [Google Scholar]
- Cech, T.L. Bemerkenswerte Krankheiten in 2004. Forstsch. Aktuell 2004, 32, 31–34. [Google Scholar]
- Cochard, B.; Crovadore, J.; Bovigny, P.Y.; Chablais, R.; Lefort, F. First reports of Cryptostroma corticale causing sooty bark disease in Acer sp. in Canton Geneva, Switzerland. New Dis. Rep. 2015, 31, 8. [Google Scholar] [CrossRef] [Green Version]
- Verkooijen, R.; Willems, J. Roetschorsziekte nu ook in Nederland. Tijdschr Bedr. Verzek. 2016, 24, 166–167. [Google Scholar] [CrossRef]
- Bencheva, S. First report of Cryptostroma corticale (Ellis & Everh.) P.H. Greg. & S. Waller on Acer platanoides L. in Bulgaria. Silva Balc. 2014, 15, 101–104. [Google Scholar]
- Koukol, O.; Kelnarová, I.; Černý, K. Recent observations of sooty bark disease of sycamore maple in Prague (Czech Republic) and the phylogenetic placement of Cryptostroma corticale. For. Pathol. 2015, 45, 21–27. [Google Scholar] [CrossRef]
- Longa, C.M.O.; Vai, N.; Maresi, G. Cryptostroma corticale in the northern Apennines (Italy). Phytopathol. Mediterr. 2016, 55, 136–138. [Google Scholar]
- Ogris, N.; Brglez, A.; Piškur, B. Drought Stress Can Induce the Pathogenicity of Cryptostroma corticale, the Causal Agent of Sooty Bark Disease of Sycamore Maple. Forests 2021, 12, 377. [Google Scholar] [CrossRef]
- Forest Health Watch (USA). Available online: https://foresthealth.org/sooty-bark-disease-on-maples-in-seattle/ (accessed on 14 October 2022).
- Kelnarová, I.; Černý, K.; Zahradník, D.; Koukol, O. Widespread latent infection of Cryptostroma corticale in asymptomatic Acer pseudoplatanus as a risk for urban plantations. For. Pathol. 2017, 47, e12344. [Google Scholar] [CrossRef]
- Burgdorf, N.; Härtl, L.; Straßer, L. Ahorn-Rußrindenkrankheit: Untersuchungen zur Verbreitung von Cryptostroma corticale. Jahrb. Baumpflege 2020, 2020, 277–284. [Google Scholar]
- Burgdorf, N.; Straßer, L. Schadverlauf und Mortalität durch die Rußrindenkrankheit in Bergahornbeständen Bayerns. Forstsch. Aktuell 2021, 66, 30–35. [Google Scholar]
- Braun, M.; Klingelhöfer, D.; Groneberg, D.A. Sooty bark disease of maples: The risk for hypersensitivity pneumonitis by fungal spores not only for woodman. J. Occup. Med. Toxicol. 2021, 16, 2. [Google Scholar] [CrossRef] [PubMed]
- Towey, J.W.; Sweany, H.C.; Huron, W.H. Severe bronchial asthma apparently due to fungus spores found in maple bark. J. Am. Med. Assoc. 1932, 99, 453–459. [Google Scholar] [CrossRef]
- German Meteorological Service (Deutscher Wetterdienst). Available online: https://www.dwd.de/DE/leistungen/klimadatendeutschland/klarchivtagmonat.html?nn=16102 (accessed on 30 August 2022).
- Ellis, M.B.; Ellis, J.P. Microfungi on Land Plants: An Identification Handbook; Croom Helm: London, UK, 1985. [Google Scholar]
- Härtl, L. Untersuchungen zur Biologie und Verbreitung von Cryptostroma corticale, Erreger der Ahorn-Rußrindenkrankheit. Bachelor’s Thesis, Department of Forestry, University of Applied Sciences Weihenstephan-Triesdorf, Freising, Germany, 2020. [Google Scholar]
- Bork, K. Rußrindenkrankheit an Ahorn—Erstfund in Bayern. AFZ Wald 2018, 20, 34–35. [Google Scholar]
- Grüner, J.; Berens, A.; Delb, H. Die Ahorn-Rußrindenkrankheit in Südwestdeutschland: Gefahren, Prognose und Empfehlungen. Waldschutz Info 2020, 2, 1–8. [Google Scholar]
- Schlösser, R.; Langer, G. Verbreitung der Rußrindenkrankheit des Ahorns in Deutschland. AFZ Wald 2021, 24, 28–32. [Google Scholar]
- Garbelotto, M.; Smith, T.; Schweigkofler, W. Variation in rates of spore deposition of Fusarium circinatum, the causal agent of pine pitch canker, over a 12-month-period at two locations in Northern California. Phytopathology 2008, 98, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Mesanza, N.; García-García, D.; Raposo, E.R.; Raposo, R.; Iturbide, M.; Pascual, M.T.; Barrena, I.; Urkola, A.; Berano, N.; Sáez de Zerain, A.; et al. Weather Variables Associated with Spore Dispersal of Lecanosticta acicola Causing Pine Needle Blight in Northern Spain. Plants 2021, 10, 2788. [Google Scholar] [CrossRef]
- Chandelier, A.; Helson, M.; Dvorak, M.; Gischer, F. Detection and quantification of airborne inoculum of Hymenoscyphus pseudoalbidus using real-time PCR assays. Plant Pathol. 2014, 63, 1296–1305. [Google Scholar] [CrossRef]
- Dvorak, M.; Rotkova, G.; Botella, L. Detection of airborne inoculum of Hymenoscyphus fraxineus and H. albidus during seasonal fluctuations associated with absence of apothecia. Forests 2015, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Wyka, S.A.; McIntire, C.D.; Smith, C.; Munck, I.A.; Rock, B.N.; Asbjornsen, H.; Broders, K.D. Effect of climatic variables on abundance and dispersal of Lecanosticta acicola spores and their impact on defoliation on eastern white pine. Phytopathology 2018, 108, 374–383. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burgdorf, N.; Härtl, L.; Hahn, W.A. Sooty Bark Disease in Sycamore: Seasonal and Vertical Variation in Spore Release of Cryptostroma corticale. Forests 2022, 13, 1956. https://doi.org/10.3390/f13111956
Burgdorf N, Härtl L, Hahn WA. Sooty Bark Disease in Sycamore: Seasonal and Vertical Variation in Spore Release of Cryptostroma corticale. Forests. 2022; 13(11):1956. https://doi.org/10.3390/f13111956
Chicago/Turabian StyleBurgdorf, Nicole, Lisa Härtl, and W. Andreas Hahn. 2022. "Sooty Bark Disease in Sycamore: Seasonal and Vertical Variation in Spore Release of Cryptostroma corticale" Forests 13, no. 11: 1956. https://doi.org/10.3390/f13111956