1. Introduction
Tree growth models describe the growth and development of forest ecosystems by considering how the dimensions of each simulated tree change through time, i.e., periodic increment of a tree in response to life processes [
1,
2] and can be divided into single-tree models (dendrometric variables) and whole stand models that include stand characteristics such as age, size, density, site index and species affiliation and composition [
3,
4]. These models are used to answer ecological questions, for example, determining the impact of the interdependence between tree species and, their environment on forest development and assessing the forest yields under certain prescribed conditions [
1]. Tree growth models are therefore important tools for understanding the dynamics of forest ecosystems and for effective and sustainable forest management [
5].
Among others, the Hossfeld [
6], Weibull [
7], Korf [
8] and Richards [
9] models are well-known flexible sigmoid growth functions [
10,
11]. These models have commonly used three growth parameters that describe various biological processes and behaviours: (i) the upper asymptote (
), which is the maximal yield indicated by a final dimension, as the diameter at breast height (dbh) at the end of the growth stage, (ii) the maximum specific growth rate (
), which is defined as the slope of the tangent at the inflexion point and (iii) the time elapsed (
), which is defined by the intercept of this tangent with the
t-axis [
12]. To the best of our knowledge, however, associations between the three parameters have not been documented for tree species.
Tree growth models are based on height-age or diameter-age relationships for evaluating tree growth, yield potential and site productivity [
13]. To avoid the problems inherent to base-age-specific (BAS) site models, Bailey and Clutter [
14] simultaneously estimated the site-specific or site-index parameters and the common or global model parameters, using Algebraic Difference Approach (ADA) models. Later, Cieszewski [
15,
16] proposed the Generalized Algebraic Difference Approach (GADA) models, which allow (and need) more than one parameter to be site specific using at least three growth parameters. GADA produces dynamic site curves with “concurrent” variable asymptotes and polymorphism, and it uses a variety of growth characteristics found across different site qualities. This is why GADA has been widely used for modelling tree growth and site quality in forest yield science [
17]. However, GADA also has some weak points, e.g. there is no closed-form solution for the growth intensity factor
, when
is assumed to be site-specific in a three-parameter Chapman–Richards model, and therefore neither
nor
can be site specific along with
[
15,
16,
18].
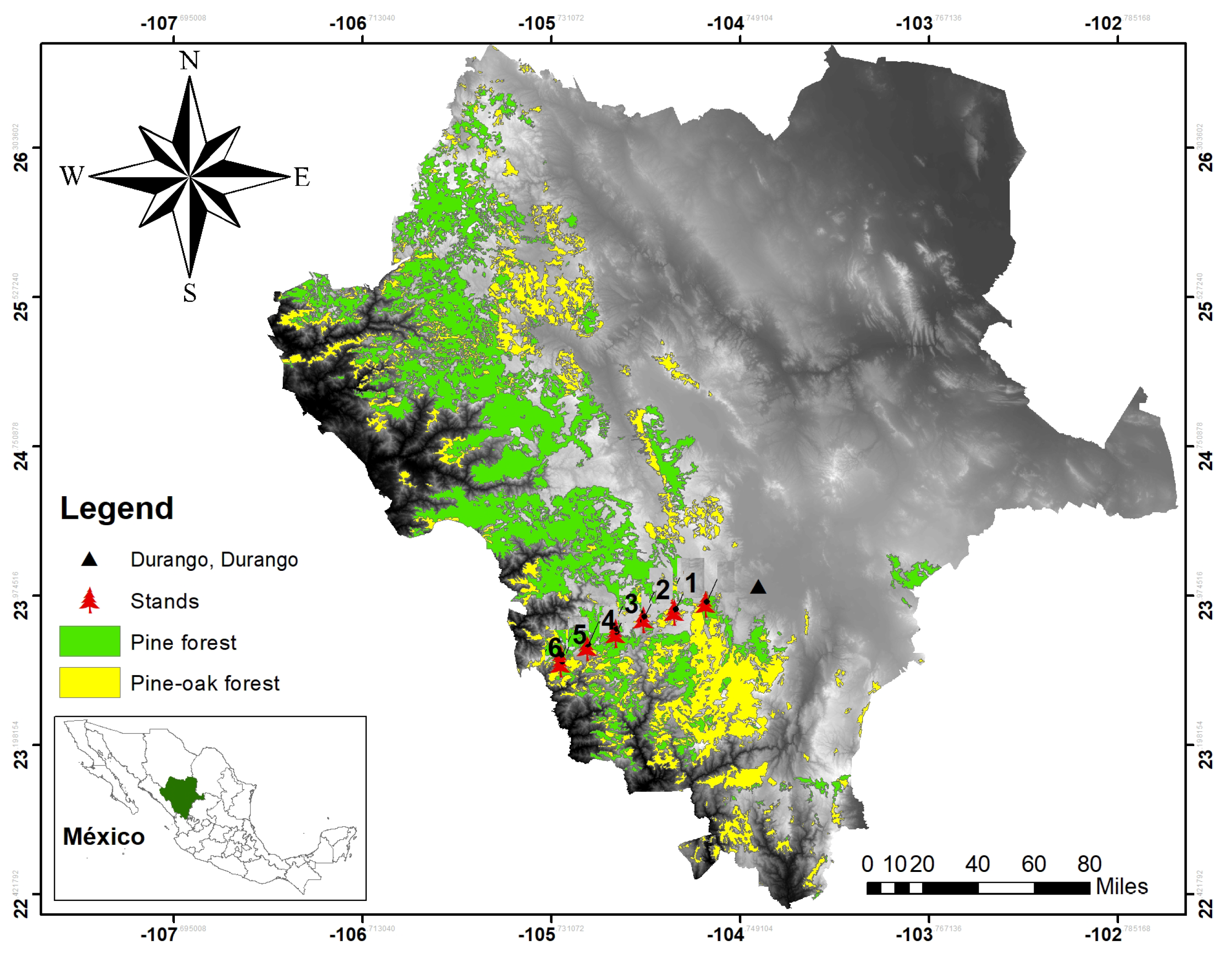
Using diameter growth data from pine trees located in typical mixed and uneven-aged pine-oak forests in the Sierra Madre Occidental, Mexico [
19,
20], our study aims were: [
21] to quantify the putative associations between the growth model parameters and, to test the accuracy of a novel Chapman-Richards growth model, in relation to Hossfeld, Lundqvist and Chapman-Richards GADA models, all with three parameters [
22]. Our research results are important for forestry practice, as the new model can be used to predict future yields in a simpler way and with the same accuracy.
3. Results
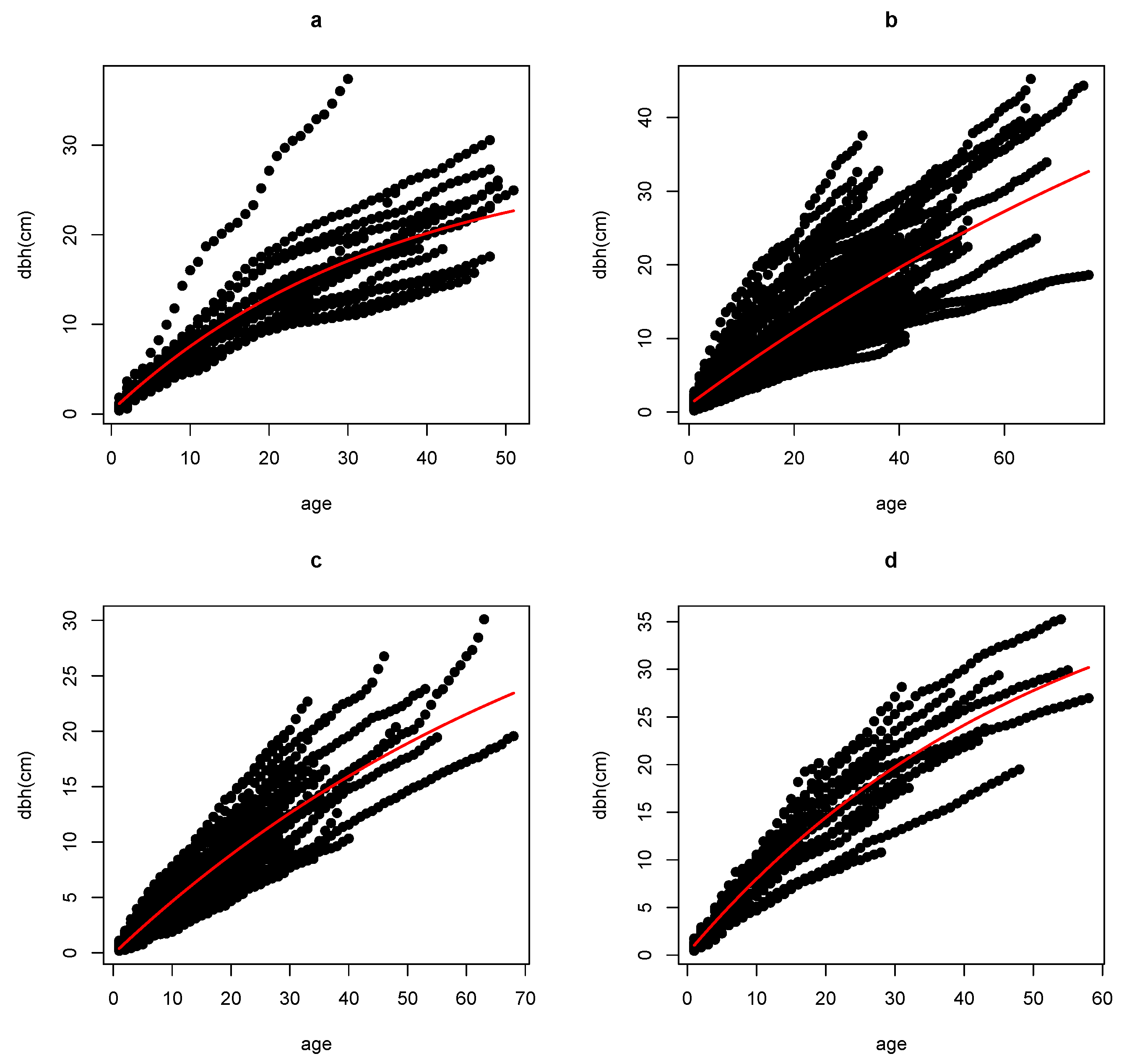
As we can see in
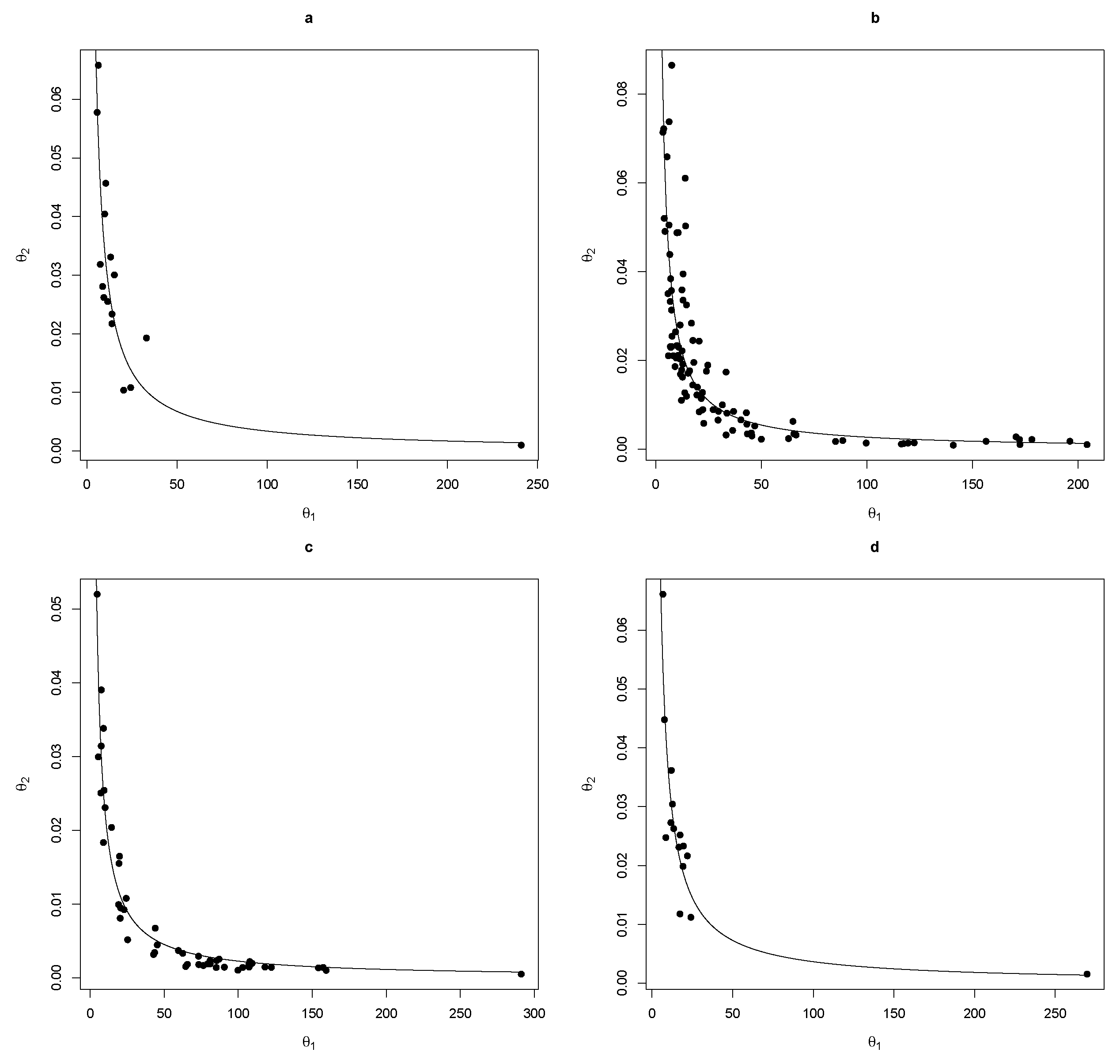
Figure 2, the Chapman-Richards BAS model fits very well with the growth history of the species involved in the study. The results show a strong negative relationship between the growth parameters
and
(
Figure 3). The scale factor
a varied from 0.23–0.34 depending on the tree species. From the adjusted models, it was thus possible to write
as a function of
of the form
.
Table 3 shows the results of fitting the parameters for the models presented in
Table 2 and for the hybrid model (CR-H) proposed for each of the species included in the study.
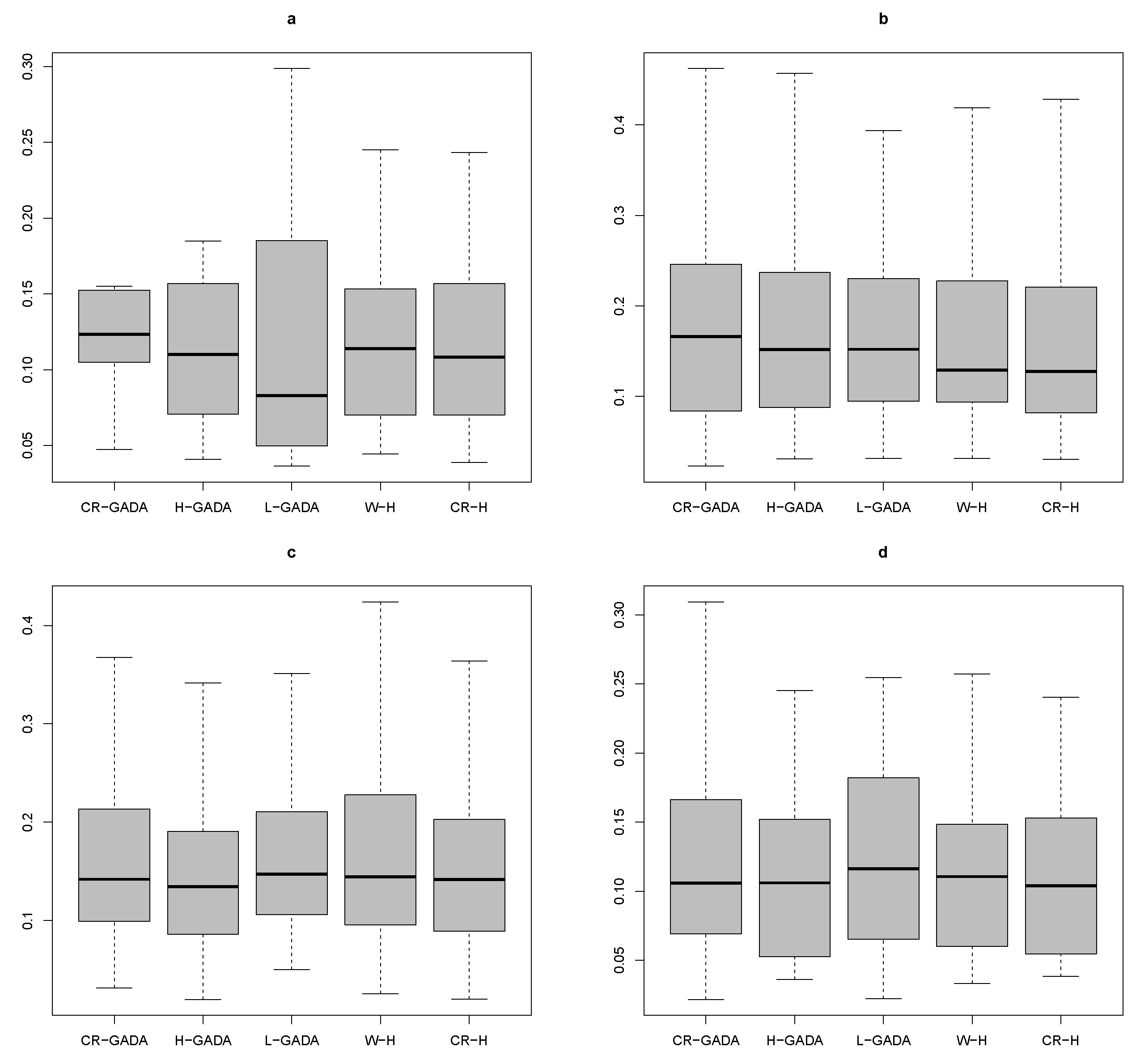
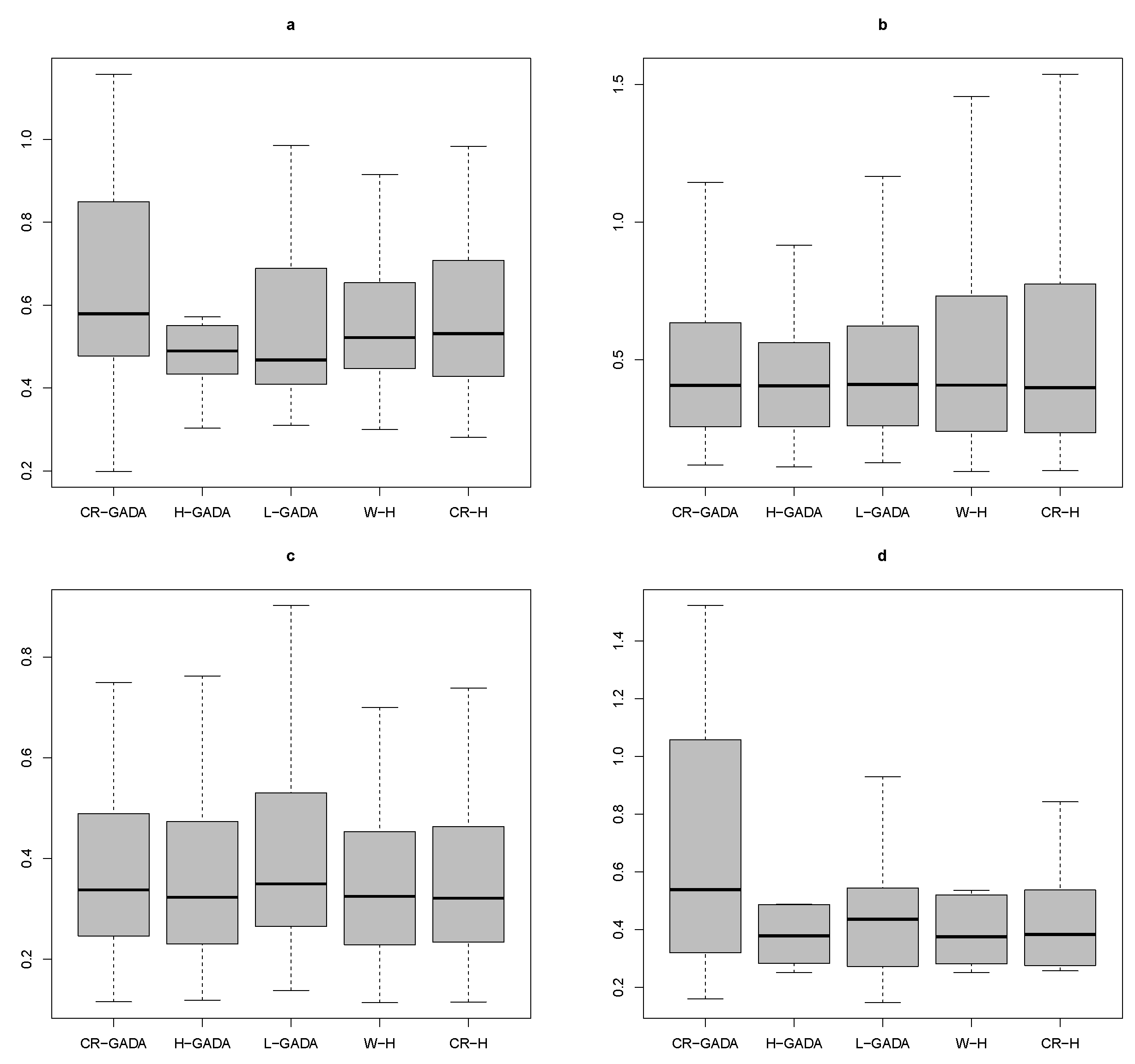
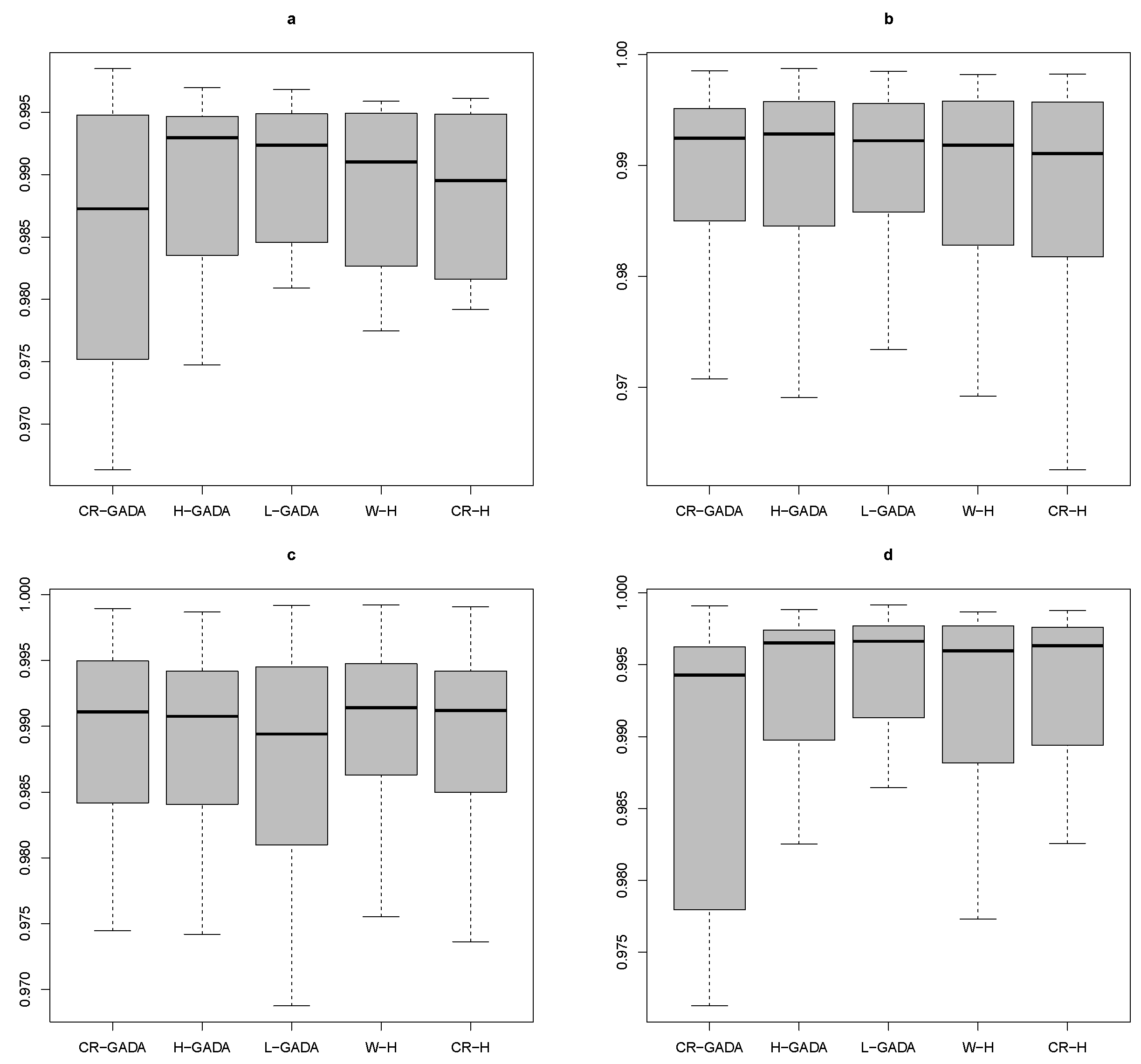
Applying the Kruskal-Wallis test to the four species studied (
Pinus arizonica,
Pinus engelmannii,
Pinus strobiformis and
Pinus teocote), we searched for possible differences between the CR-GADA, H-GADA, L-GADA and W-H models with respect to our proposed model (CR-H). Our results show that there are no significant differences in any of the parameters (MAPE, RMSE and
R2) tested to compare the models, as the
p-values for the K-W test were above 0.65 in all cases. In addition, the AIC values were also similar for each model (see
Table 4,
Table 5,
Table 6 and
Table 7 and
Figure 4,
Figure 5 and
Figure 6).
4. Discussion and Conclusions
The results show a strong negative association between the maximum yield of the growth parameters, given at the final dimension reached at the end of their respective growth period (
) and the maximum specific growth rate (
). In other words, the less value has the slope (current annual increment) in the inflexion point of the growth curve of the tree, the greater the final maximum yield. Additionally,
depends on the actual environment and initial physiological state of the population/trees [
28]. The accuracy of our GADA and H models (W-H and CR-H) was similar to the GADA models of other studies (e.g., [
17,
29,
30,
31]).
The main advantage of the widely used GADA site index models over ADA is that they can be polymorphic and have multiple asymptotes caused by more than one site-specific parameter to create functions of
(i.e., one unobservable independent variable which describes site productivity, as a summary of management regimes, soil, climatic and ecological factors) [
29,
30]. This leads to more flexible dynamic models [
31]. However, if these site-specific parameters are highly correlated to each other, as we have shown (
Figure 2), these several parameters could be replaced by the single parameter easiest to estimate in the forest practice, making the model mathematically and economically more attractive due to the smaller number of parameters, which makes a complex multiparametric
function obsolete, without compromising the fitting capability of the model
Table 4,
Table 5,
Table 6 and
Table 7 [
22,
26]. I.e., the main advantage of this novel CR-H model over GADA is the simplification described above by reducing the number of site-specific parameters to the only one most easily measurable site-specific parameter, while statistically maintaining the predictive quality of the model. Moreover, in contrast to the CR-GADA factor
, when
is assumed to be site-specific [
18], the CR-H has always a closed-form solution.
The study findings show that the model proposed in this study is accurate and feasible, at least for the population analyzed. This model can be classified as a hybrid, as we initially applied a variable substitution of the type , to then assume that the parameter a is site-specific (ADA methodology). If the relationship shown by parameters and can also be reproduced in data sets for other organisms, this new model will greatly reduce the economical and computational effort invested in obtaining the model parameters.
Although the quality of the five growth models applied in the experiment was very similar from a statistical point of view, the proposed CR-H, like the W-H, is easy to apply, as it has only as a parameter, which is the maximum tree diameter or the final diameter of the tree at the end of its respective growth stage. This parameter can be easily obtained, by measuring just the dominant trees, especially in coniferous forests with irregular ages.
It would be interesting for a future study to test the same hypothesis with other species and other growth models, in order to determine whether the CR-H model performance is replicated. Furthermore, sufficiently high correlations of the site-specific parameters , and , and therefore, the accuracy of these simplified dynamic H models should be tested, when environmental changes and competition parameters are incorporated into the models that effectively perturb the growth of individuals.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}