
Linking Bacterial Rhizosphere Communities of Two Pioneer Species, Brachystegia boehmii and B. spiciformis, to the Ecological Processes of Miombo Woodlands
 , , , ,
, , , ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Design
2.2. Genomic DNA Extraction, Library Construction, and Illumina Sequencing
2.3. Assembly of Reads and Taxonomical Assignment
2.4. Functional Prediction of Miombo Soils
2.5. Isotopic Composition and Elemental Analysis
3. Results
3.1. Microbial Community Composition and Diversity
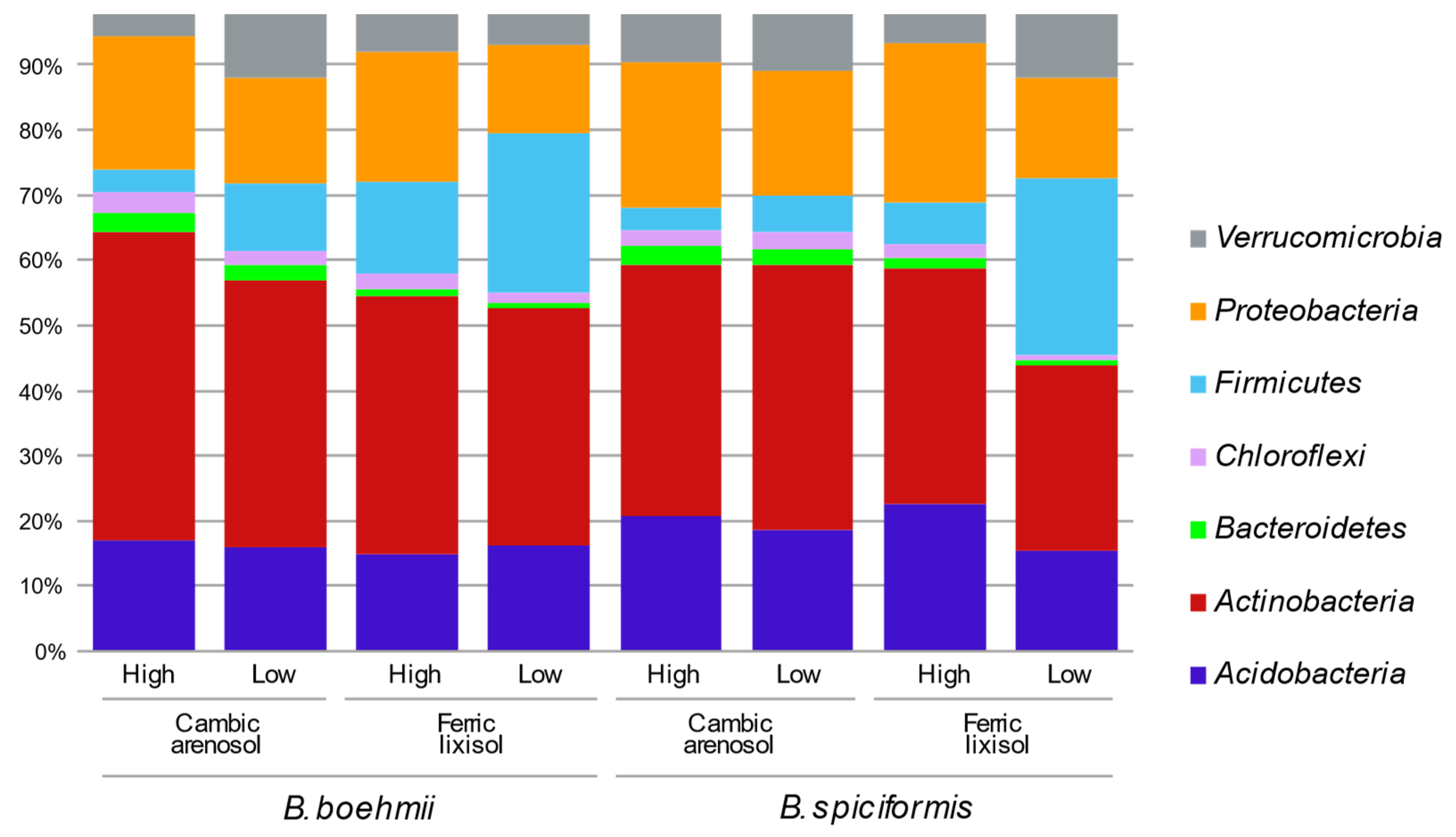
3.2. Composition of Soil Bacterial Communities
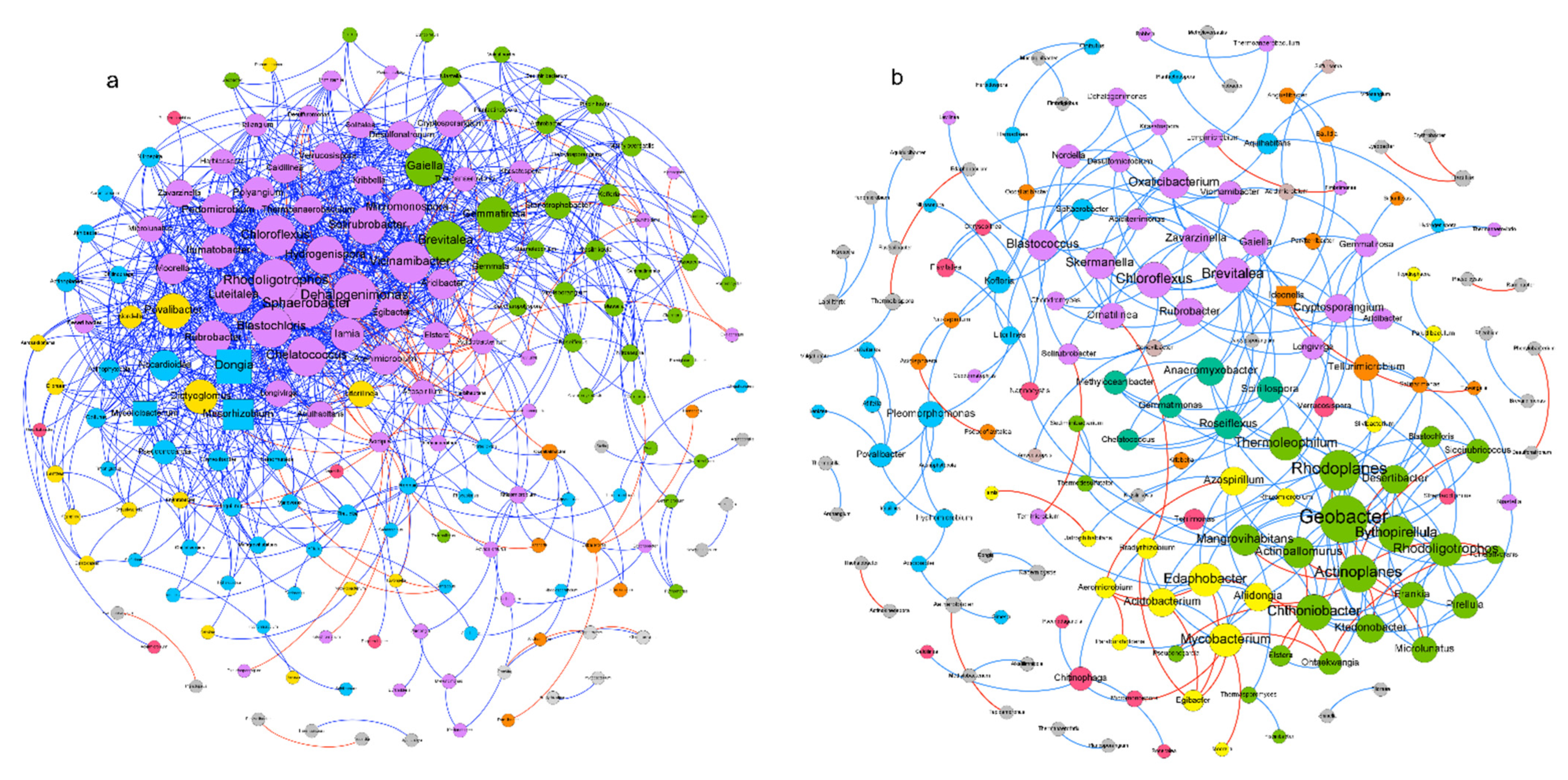
3.3. Co-Occurrence Patterns and Community Complexity
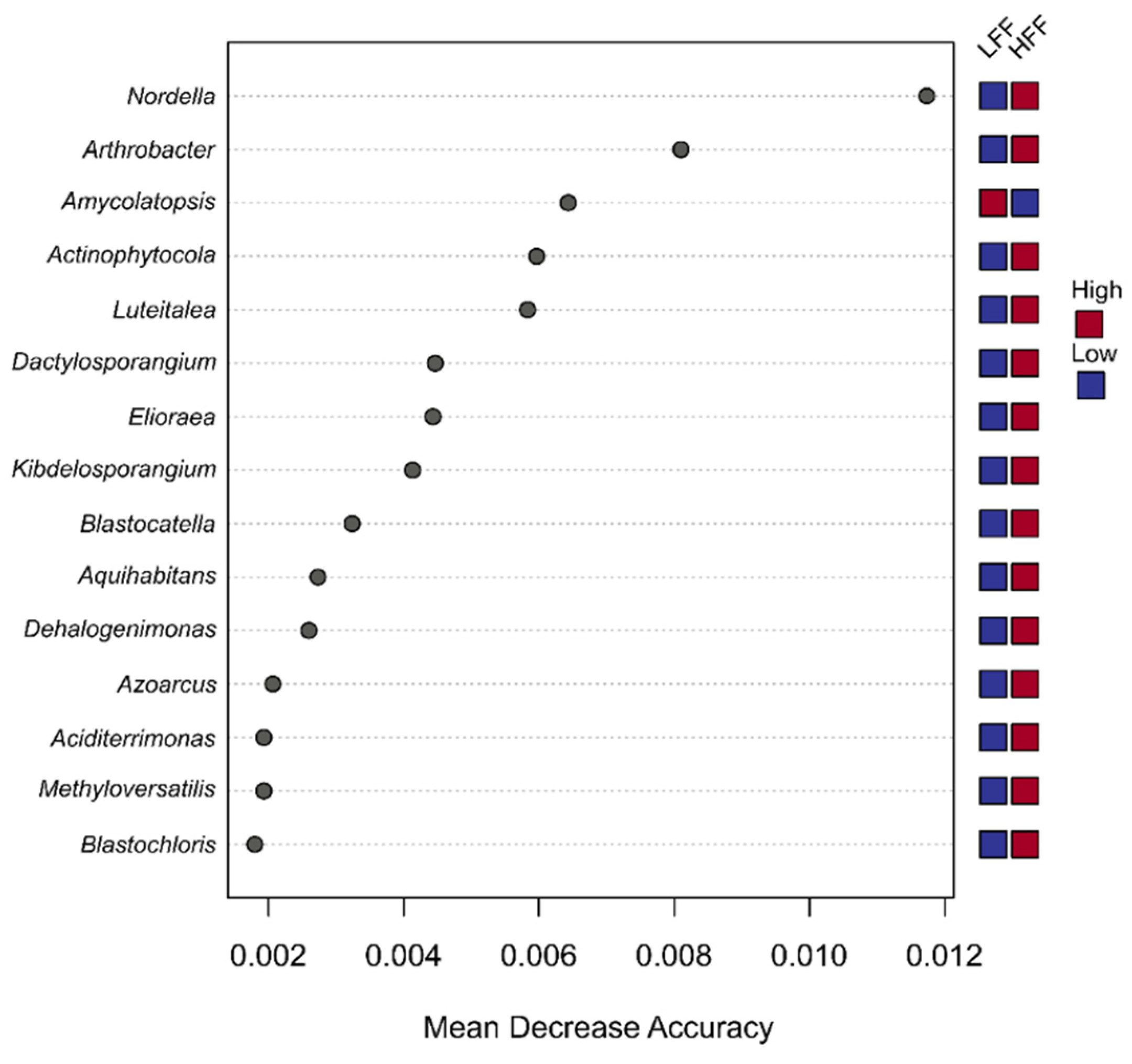
3.4. Functional Analysis of Bacterial Communities
3.5. Carbon and Nitrogen Isotopic Composition
4. Discussion
4.1. Effects of Fire and Soil Types in Microbial Rhizosphere Diversity
4.2. Disentangling Changes in the Rhizosphere Community Structure
4.3. New Insights from Miombo Bacterial Ecological Networks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WWF Miombo Eco-Region. “Home of the Zambezi”. Conservation Strategy: 2011–2020; WWF: Harare, Zimbabwe, 2012. [Google Scholar]
- Macave, O.A.; Ribeiro, N.S.; Ribeiro, A.I.; Chaúque, A.; Bandeira, R.; Branquinho, C.; Washington-Allen, R. Modelling Aboveground Biomass of Miombo Woodlands in Niassa Special Reserve, Northern Mozambique. Forests 2022, 13, 311. [Google Scholar] [CrossRef]
- Jew, E.K.K.; Dougill, A.J.; Sallu, S.M.; O’Connell, J.; Benton, T.G. Miombo woodland under threat: Consequences for tree diversity and carbon storage. For. Ecol. Manag. 2016, 361, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Lupala, Z.J.; Lusambo, L.P.; Ngaga, Y.M. Management, Growth, and Carbon Storage in Miombo Woodlands of Tanzania. Int. J. For. Res. 2014, 2014, 629317. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro-Barros, A.I.; Silva, M.J.; Moura, I.; Ramalho, J.C.; Máguas-Hanson, C.; Ribeiro, N.S. The Potential of Tree and Shrub Legumes in Agroforestry Systems. In Nitrogen in Agriculture-Updates; IntechOpen: London, UK, 2018; ISBN 978-953-51-3769-6. [Google Scholar]
- Ribeiro, N.S.; Saatchi, S.S.; Shugart, H.H.; Washington-Allen, R.A. Aboveground biomass and leaf area index (LAI) mapping for Niassa Reserve, northern Mozambique. J. Geophys. Res. Biogeosciences 2008, 113. [Google Scholar] [CrossRef] [Green Version]
- Chinder, G.B.; Hattas, D.; Massad, T.J. Growth and functional traits of Julbernardia globiflora (Benth) resprouts and seedlings in response to fire frequency and herbivory in miombo woodlands. S. Afr. J. Bot. 2020, 135, 476–483. [Google Scholar] [CrossRef]
- Ribeiro, N.S.; Armstrong, A.H.; Fischer, R.; Kim, Y.S.; Shugart, H.H.; Ribeiro-Barros, A.I.; Chauque, A.; Tear, T.; Washington-Allen, R.; Bandeira, R.R. Prediction of forest parameters and carbon accounting under different fire regimes in Miombo woodlands, Niassa Special Reserve, Northern Mozambique. For. Policy Econ. 2021, 133, 102625. [Google Scholar] [CrossRef]
- Maquia, I.S.A.; Fareleira, P.; Castro, I.V.E.; Soares, R.; Brito, D.R.A.; Mbanze, A.A.; Chaúque, A.; Máguas, C.; Ezeokoli, O.T.; Ribeiro, N.S.; et al. The nexus between fire and soil bacterial diversity in the african miombo woodlands of niassa special reserve, mozambique. Microorganisms 2021, 9, 1562. [Google Scholar] [CrossRef]
- Cardenas, E.; Kranabetter, J.M.; Hope, G.; Maas, K.R.; Hallam, S.; Mohn, W.W. Forest harvesting reduces the soil metagenomic potential for biomass decomposition. ISME J. 2015, 9, 2465–2476. [Google Scholar] [CrossRef] [Green Version]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ.-Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- de Souza, L.C.; Procópio, L. The profile of the soil microbiota in the Cerrado is influenced by land use. Appl. Microbiol. Biotechnol. 2021, 105, 4791–4803. [Google Scholar] [CrossRef]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire effects on belowground sustainability: A review and synthesis. In Forest Ecology and Management; Elsevier: Amsterdam, The Netherlands, 1999; Volume 122, pp. 51–71. [Google Scholar]
- Maquia, I.S.; Fareleira, P.; Castro, I.V.E.; Brito, D.R.A.; Soares, R.; Chaúque, A.; Ferreira-Pinto, M.M.; Lumini, E.; Berruti, A.; Ribeiro, N.S.; et al. Mining the microbiome of key species from african savanna woodlands: Potential for soil health improvement anwd plant growth promotion. Microorganisms 2020, 8, 1291. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, H.; Rodríguez-Echeverría, S. Identification of symbiotic nitrogen-fixing bacteria from three African leguminous trees in Gorongosa National Park. Syst. Appl. Microbiol. 2016, 39, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Kurdali, F.; Al-Shamma’a, M. Natural abundances of 15N and 13C in leaves of some N2-fixing and non-N2-fixing trees and shrubs in Syria. Isotopes Environ. Health Stud. 2009, 45, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Craine, J.M.; Brookshire, E.N.J.; Cramer, M.D.; Hasselquist, N.J.; Koba, K.; Marin-Spiotta, E.; Wang, L.; Craine, J.M.; Ventures, J.; Cramer, M.D.; et al. Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils. Plant Soil 2015, 396, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Ulm, F.; Hellmann, C.; Cruz, C.; Máguas, C. N/P imbalance as a key driver for the invasion of oligotrophic dune systems by a woody legume. Oikos 2017, 126, gc735. [Google Scholar] [CrossRef]
- del Amor, F.M.; Cuadra-Crespo, P. Alleviation of salinity stress in broccoli using foliar urea or methyl-jasmonate: Analysis of growth, gas exchange, and isotope composition. Plant Growth Regul. 2011, 63, 55–62. [Google Scholar] [CrossRef]
- Ribeiro, N.S.; Grundy, I.M.; Gonçalves, F.M.P.; Moura, I.; Santos, M.J.; Kamoto, J.; Ribeiro-Barros, A.I.; Gandiwa, E. People in the Miombo Woodlands: Socio-Ecological Dynamics. In Miombo Woodlands in a Changing Environment: Securing the Resilience and Sustainability of People and Woodlands; Springer International Publishing: Manhattan, NY, USA, 2020; pp. 55–100. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Campbell, B.M. The Miombo in Transition: Woodlands and Welfare in Africa; Center for International Forestry Research: Bogor Barat, Indonesia, 1996; ISBN 9798764072. [Google Scholar]
- Ryan, C.M.; Williams, M. How does fire intensity and frequency affect miombo woodland tree populations and biomass? Ecol. Appl. 2011, 21, 48–60. [Google Scholar] [CrossRef]
- Fernandes, J.F. Os solos do Parque Nacional da Gorongosa. Inst. Investig. Agronómica Moçambique Commun. 1968, 19, 1–78. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 2016, e2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greengenes. Available online: https://greengenes.lbl.gov/Download/ (accessed on 12 October 2022).
- Alpha_Diversity.Py—Calculate Alpha Diversity on Each Sample in an Otu Table, Using a Variety of Alpha Diversity Metrics—Homepage. Available online: http://qiime.org/scripts/alpha_diversity.html (accessed on 12 October 2022).
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Jiang, Y.H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [Green Version]
- Maslov, S.; Sneppen, K. Specificity and stability in topology of protein networks. Science 2002, 296, 910–913. [Google Scholar] [CrossRef] [Green Version]
- Guimerà, R.; Amaral, L.A.N. Functional cartography of complex metabolic networks. Nature 2005, 433, 895–900. [Google Scholar] [CrossRef] [Green Version]
- Aßhauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2: An improved and extensible approach for metagenome inference. bioRxiv 2019, 672295. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Doebeli, M. Efficient comparative phylogenetics on large trees. Bioinformatics 2018, 34, 1053–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Doak, T.G. A parsimony approach to biological pathway reconstruction/inference for genomes and metagenomes. PLoS Comput. Biol. 2009, 5, e1000465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspi, R.; Billington, R.; Fulcher, C.A.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Latendresse, M.; Midford, P.E.; Ong, Q.; Ong, W.K.; et al. The MetaCyc database of metabolic pathways and enzymes. Nucleic Acids Res. 2018, 46, D633–D639. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Duro, N.; Batista-Santos, P.; da Costa, M.; Maia, R.; Castro, I.V.; Ramos, M.; Ramalho, J.C.; Pawlowski, K.; Máguas, C.; Ribeiro-Barros, A. The impact of salinity on the symbiosis between Casuarina glauca Sieb. ex Spreng. and N2-fixing Frankia bacteria based on the analysis of Nitrogen and Carbon metabolism. Plant Soil 2016, 398, 327–337. [Google Scholar] [CrossRef]
- Sampaio, L.; Freitas, R.; Máguas, C.; Rodrigues, A.; Quintino, V. Coastal sediments under the influence of multiple organic enrichment sources: An evaluation using carbon and nitrogen stable isotopes. Mar. Pollut. Bull. 2010, 60, 272–282. [Google Scholar] [CrossRef]
- Coleman, M.; Meier-Augenstein, W. Ignoring IUPAC guidelines for measurement and reporting of stable isotope abundance values affects us all. Rapid Commun. Mass Spectrom. 2014, 28, 1953–1955. [Google Scholar] [CrossRef]
- Zhou, L.; Ng, H.K.; Drautz-Moses, D.I.; Schuster, S.C.; Beck, S.; Kim, C.; Chambers, J.C.; Loh, M. Systematic evaluation of library preparation methods and sequencing platforms for high-throughput whole genome bisulfite sequencing. Sci. Rep. 2019, 9, 10383. [Google Scholar] [CrossRef] [PubMed]
- Kara, O.; Bolat, I. Short-term effects of wildfire on microbial biomass and abundance in black pine plantation soils in Turkey. Ecol. Indic. 2009, 9, 1151–1155. [Google Scholar] [CrossRef]
- Docherty, K.M.; Balser, T.C.; Bohannan, B.J.M.; Gutknecht, J.L.M. Soil microbial responses to fire and interacting global change factors in a California annual grassland. Biogeochemistry 2012, 109, 63–83. [Google Scholar] [CrossRef]
- Whitman, T.; Whitman, E.; Woolet, J.; Flannigan, M.D.; Thompson, D.K.; Parisien, M.A. Soil bacterial and fungal response to wildfires in the Canadian boreal forest across a burn severity gradient. Soil Biol. Biochem. 2019, 138, 107571. [Google Scholar] [CrossRef]
- Fernández-González, A.J.; Martínez-Hidalgo, P.; Cobo-Díaz, J.F.; Villadas, P.J.; Martínez-Molina, E.; Toro, N.; Tringe, S.G.; Fernández-López, M. The rhizosphere microbiome of burned holm-oak: Potential role of the genus Arthrobacter in the recovery of burned soils. Sci. Rep. 2017, 7, 6008. [Google Scholar] [CrossRef]
- Villadas, P.J.; Díaz-Díaz, S.; Rodríguez-Rodríguez, A.; del Arco-Aguilar, M.; Fernández-González, A.J.; Pérez-Yépez, J.; Arbelo, C.; González-Mancebo, J.M.; Fernández-López, M.; León-Barrios, M. The soil microbiome of the laurel forest in garajonay national park (La Gomera, Canary Islands): Comparing unburned and burned habitats after a wildfire. Forests 2019, 10, 1051. [Google Scholar] [CrossRef] [Green Version]
- Castañeda, L.E.; Barbosa, O. Metagenomic analysis exploring taxonomic and functional diversity of soil microbial communities in Chilean vineyards and surrounding native forests. PeerJ 2017, 2017, e3098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Araujo, A.S.F.; Mendes, L.W.; Lemos, L.N.; Antunes, J.E.L.; Beserra, J.E.A.; de Lyra, M.D.C.C.P.; Figueiredo, M.D.V.B.; Lopes, C.D.A.; Gomes, R.L.F.; Bezerra, W.M.; et al. Protist species richness and soil microbiome complexity increase towards climax vegetation in the Brazilian Cerrado. Commun. Biol. 2018, 1, 135. [Google Scholar] [CrossRef] [Green Version]
- de Souza, L.C.; Procópio, L. The adaptations of the microbial communities of the savanna soil over a period of wildfire, after the first rains, and during the rainy season. Environ. Sci. Pollut. Res. 2022, 29, 14070–14082. [Google Scholar] [CrossRef]
- Ganz, H.H.; Karaoz, U.; Getz, W.M.; Versfeld, W.; Brodie, E.L. Diversity and structure of soil bacterial communities associated with vultures in an African savanna. Ecosphere 2012, 3, art47. [Google Scholar] [CrossRef]
- Lan, G.; Li, Y.; Jatoi, M.T.; Tan, Z.; Wu, Z.; Xie, G. Change in Soil Microbial Community Compositions and Diversity Following the Conversion of Tropical Forest to Rubber Plantations in Xishuangbanan, Southwest China. Trop. Conserv. Sci. 2017, 10, 1–14. [Google Scholar] [CrossRef]
- Dube, J.P.; Valverde, A.; Steyn, J.M.; Cowan, D.A.; van der Waals, J.E. Differences in bacterial diversity, Composition and function due to long-term agriculture in soils in the Eastern Free State of South Africa. Diversity 2019, 11, 61. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.P.; You, L.X.; Yang, X.J.; Yu, Y.S.; Zhang, H.T.; Yang, B.; Chorover, J.; Feng, R.W.; Rensing, C. Extrapolymeric substances (EPS) in Mucilaginibacter rubeus P2 displayed efficient metal(loid) bio-adsorption and production was induced by copper and zinc. Chemosphere 2022, 291, 132712. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kong, H.G.; Song, G.C.; Ryu, C.M. Disruption of Firmicutes and Actinobacteria abundance in tomato rhizosphere causes the incidence of bacterial wilt disease. ISME J. 2021, 15, 330–347. [Google Scholar] [CrossRef]
- Philippot, L.; Andersson, S.G.E.; Battin, T.J.; Prosser, J.I.; Schimel, J.P.; Whitman, W.B.; Hallin, S. The ecological coherence of high bacterial taxonomic ranks. Nat. Rev. Microbiol. 2010, 8, 523–529. [Google Scholar] [CrossRef]
- Beier, S.; Bertilsson, S. Bacterial chitin degradation-mechanisms and ecophysiological strategies. Front. Microbiol. 2013, 4, 149. [Google Scholar] [CrossRef] [Green Version]
- Pointing, S.B.; Chan, Y.; Lacap, D.C.; Lau, M.C.Y.; Jurgens, J.A.; Farrell, R.L. Highly specialized microbial diversity in hyper-arid polar desert. Proc. Natl. Acad. Sci. USA 2009, 106, 19964–19969. [Google Scholar] [CrossRef] [Green Version]
- Black, M.; Moolhuijzen, P.; Chapman, B.; Barrero, R.; Howieson, J.; Hungria, M.; Bellgard, M. The genetics of symbiotic nitrogen fixation: Comparative genomics of 14 rhizobia strains by resolution of protein clusters. Genes 2012, 3, 138–166. [Google Scholar] [CrossRef] [Green Version]
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Corrigendum: Bacillus velezensis FZB42 in 2018: The gram-positive model strain for plant growth promotion and biocontrol. Front. Microbiol. 2018, 9, 2491, Erratum in: Front. Microbiol. 2019, 10, 1279. [Google Scholar] [CrossRef] [Green Version]
- Osama, N.; Bakeer, W.; Raslan, M.; Soliman, H.A.; Abdelmohsen, U.R.; Sebak, M. Anti-cancer and antimicrobial potential of five soil Streptomycetes: A metabolomics-based study. R. Soc. Open Sci. 2022, 9, 211509. [Google Scholar] [CrossRef]
- Vergnes, S.; Gayrard, D.; Veyssière, M.; Toulotte, J.; Martinez, Y.; Dumont, V.; Bouchez, O.; Rey, T.; Dumas, B. Phyllosphere colonization by a soil Streptomyces sp. promotes plant defense responses against fungal infection. Mol. Plant-Microbe Interact. 2020, 33, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Severino, R.; Froufe, H.J.C.; Barroso, C.; Albuquerque, L.; Lobo-da-Cunha, A.; da Costa, M.S.; Egas, C. High-quality draft genome sequence of Gaiella occulta isolated from a 150 meter deep mineral water borehole and comparison with the genome sequences of other deep-branching lineages of the phylum Actinobacteria. Microbiologyopen 2019, 8, e00840. [Google Scholar] [CrossRef] [Green Version]
- Parente, C.E.T.; Brito, E.M.S.; Caretta, C.A.; Cervantes-Rodríguez, E.A.; Fábila-Canto, A.P.; Vollú, R.E.; Seldin, L.; Malm, O. Bacterial diversity changes in agricultural soils influenced by poultry litter fertilization. Braz. J. Microbiol. 2021, 52, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Pertile, M.; Sousa, R.M.S.; Mendes, L.W.; Antunes, J.E.L.; Oliveira, L.M.D.S.; de Araujo, F.F.; Melo, V.M.M.; Araujo, A.S.F. Response of soil bacterial communities to the application of the herbicides imazethapyr and flumyzin. Eur. J. Soil Biol. 2021, 102, 103252. [Google Scholar] [CrossRef]
- Corrêa, L.D.R.; Paim, D.C.; Schwambach, J.; Fett-Neto, A.G. Carbohydrates as regulatory factors on the rooting of Eucalyptus saligna Smith and Eucalyptus globulus Labill. Plant Growth Regul. 2005, 45, 63–73. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Hashem, A.; Abd Allah, E.F. Bacillus: A biological tool for crop improvement through bio-molecular changes in adverse environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, S.; Jaggi, V.; Gangola, S.; Singh, A.; Sah, V.K.; Sahgal, M. Contrasting rhizosphere bacterial communities of healthy and wilted Dalbergia sissoo Roxb. forests. Rhizosphere 2021, 17, 100295. [Google Scholar] [CrossRef]
- Meier, M.A.; Lopez-Guerrero, M.G.; Guo, M.; Schmer, M.R.; Herr, J.R.; Schnable, J.C.; Alfano, J.R.; Yang, J. Rhizosphere Microbiomes in a Historical Maize-Soybean Rotation System Respond to Host Species and Nitrogen Fertilization at the Genus and Subgenus Levels. Appl. Environ. Microbiol. 2021, 87, e0313220. [Google Scholar] [CrossRef]
- Moe, L.A. Amino acids in the rhizosphere: From plants to microbes. Am. J. Bot. 2013, 100, 1692–1705. [Google Scholar] [CrossRef]
- Ribeiro, N.S.; Cangela, A.; Chauque, A.; Bandeira, R.R.; Ribeiro-Barros, A.I. Characterisation of spatial and temporal distribution of the fire regime in Niassa National Reserve, northern Mozambique. Int. J. Wildl. Fire 2017, 26, 1021–1029. [Google Scholar] [CrossRef]
- Ezeokoli, O.T.; Nuaila, V.N.A.; Obieze, C.C.; Muetanene, B.A.; Fraga, I.; Tesinde, M.N.; Ndayiragije, A.; Coutinho, J.; Melo, A.M.P.; Adeleke, R.A.; et al. Assessing the impact of rice cultivation and off-season period on dynamics of soil enzyme activities and bacterial communities in two agro-ecological regions of mozambique. Agronomy 2021, 11, 694. [Google Scholar] [CrossRef]
- Santolini, M.; Barabási, A.L. Predicting perturbation patterns from the topology of biological networks. Proc. Natl. Acad. Sci. USA 2018, 115, E6375–E6383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Ning, D.; Yang, Y.; Van Nostrand, J.D.; Zhou, J.; Wen, X. Biodegradability of wastewater determines microbial assembly mechanisms in full-scale wastewater treatment plants. Water Res. 2020, 169, 115276. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.P.; Srivastava, A.K.; Gupta, V.K.; O’Donovan, A.; Ramteke, P.W. Enhanced yield of diverse varieties of chickpea (Cicer arietinum L.) by different isolates of Mesorhizobium ciceri. Environ. Sustain. 2018, 1, 425–435. [Google Scholar] [CrossRef]
- del Barrio-Duque, A.; Ley, J.; Samad, A.; Antonielli, L.; Sessitsch, A.; Compant, S. Beneficial Endophytic Bacteria-Serendipita indica Interaction for Crop Enhancement and Resistance to Phytopathogens. Front. Microbiol. 2019, 10, 2888. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Turner, S.J.; Silvester, W.B.; Park, D.C.; Young, J.M. Unexpectedly diverse Mesorhizobium strains and Rhizobium leguminosarum nodulate native legume genera of New Zealand, while introduced legume weeds are nodulated by Bradyrhizobium species. Appl. Environ. Microbiol. 2004, 70, 5980–5987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Li, J.; Liu, X.; Li, X. Variations in carbon isotope ratios of plants across a temperature gradient along the 400 mm isoline of mean annual precipitation in north China and their relevance to paleovegetation reconstruction. Quat. Sci. Rev. 2013, 63, 83–90. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Soil Type | Fire Frequency | Fire Interval | Latitude | Longitude |
|---|---|---|---|---|
| Cambic arenosol | High | 1 year | −19.00125 | 34.20163 |
| High | 1 year | −19.00135 | 34.20174 | |
| Low | 6 years | −19.00009 | 34.20236 | |
| Low | 6 years | −18.99999 | 34.20720 | |
| Ferric lixisol | High | 1 year | −18.95165 | 34.17806 |
| High | 1 year | −18.95198 | 34.17942 | |
| Low | 6 years | −18.96186 | 34.16772 | |
| Low | 6 years | −18.96351 | 34.16873 |
| Soil Type | Species | Fire Regime | OTUs | Shannon Index |
|---|---|---|---|---|
| Cambic arenosol | B. spiciformis | HFF | 1144 ± 38 | 7.88 ± 0.34 |
| LFF | 1083 ± 40 | 7.62 ± 0.29 | ||
| B. boehmii | HFF | 1076 ± 224 | 7.87 ± 0.27 | |
| LFF | 1086 ± 82 | 7.53 ± 0.45 | ||
| Ferric lixisol | B. spiciformis | HFF | 1072 ± 150 | 7.71 ± 0.57 |
| LFF | 895 ± 219 | 6.09 ± 0.58 | ||
| B. boehmii | HFF | 1083 ± 160 | 7.44 ± 0.64 | |
| LFF | 855 ± 241 | 6.28 ± 0.32 |
| Soil Type | Species | Fire Regime | δ15N (‰) | δ13C (‰) | N (%) | C (%) | C/N |
|---|---|---|---|---|---|---|---|
| Cambic arenosol | B. spiciformis | HFF | 1.8 ± 0.2 | −28.2 ± 0.2 | 1.6 ± 0.2 | 43.9 ± 0.9 | 27.9 ± 2.3 |
| LFF | 1.9 ± 0.2 | −27.4 ± 0.3 | 1.8 ± 0.1 | 45.7 ± 0.8 | 26.1 ± 2.5 | ||
| B. boehmii | HFF | 2.1 ± 0.1 | −27.0 ± 0.3 | 1.5 ± 0.2 | 47.1 ± 1.1 | 31.1 ± 2.9 | |
| LFF | 0.7 ± 0.2 | −28.2 ± 0.4 | 1.5 ± 0.2 | 47.5 ± 1.2 | 31.6 ± 3.0 | ||
| Ferric lixisol | B. spiciformis | HFF | 2.2 ± 0.2 | −27.2 ± 0.2 | 1.7 ± 0.1 | 44.2 ± 1.4 | 26.9 ± 2.3 |
| LFF | 2.0 ± 0.2 | −27.8 ± 0.3 | 1.6 ± 0.1 | 44.8 ± 1.1 | 27.7 ± 2.5 | ||
| B. boehmii | HFF | 1.6 ± 0.1 | −27.6 ± 0.3 | 1.5 ± 0.2 | 48.0 ± 1.1 | 31.8 ± 2.9 | |
| LFF | 1.7 ± 0.2 | −27.8 ± 0.4 | 1.3 ± 0.1 | 48.2 ± 0.2 | 36.7 ± 3.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
António, C.B.S.; Obieze, C.; Jacinto, J.; Maquia, I.S.A.; Massad, T.; Ramalho, J.C.; Ribeiro, N.S.; Máguas, C.; Marques, I.; Ribeiro-Barros, A.I. Linking Bacterial Rhizosphere Communities of Two Pioneer Species, Brachystegia boehmii and B. spiciformis, to the Ecological Processes of Miombo Woodlands. Forests 2022, 13, 1840. https://doi.org/10.3390/f13111840
António CBS, Obieze C, Jacinto J, Maquia ISA, Massad T, Ramalho JC, Ribeiro NS, Máguas C, Marques I, Ribeiro-Barros AI. Linking Bacterial Rhizosphere Communities of Two Pioneer Species, Brachystegia boehmii and B. spiciformis, to the Ecological Processes of Miombo Woodlands. Forests. 2022; 13(11):1840. https://doi.org/10.3390/f13111840
Chicago/Turabian StyleAntónio, Camilo B. S., Chinedu Obieze, João Jacinto, Ivete S. A. Maquia, Tara Massad, José C. Ramalho, Natasha S. Ribeiro, Cristina Máguas, Isabel Marques, and Ana I. Ribeiro-Barros. 2022. "Linking Bacterial Rhizosphere Communities of Two Pioneer Species, Brachystegia boehmii and B. spiciformis, to the Ecological Processes of Miombo Woodlands" Forests 13, no. 11: 1840. https://doi.org/10.3390/f13111840