
Morpho-Physiological Strategies of Shorea leprosula Miq. and Shorea acuminata Dyer in Response to Light Intensity and Nutrient Amendments

Abstract
:1. Introduction
2. Materials and Methods
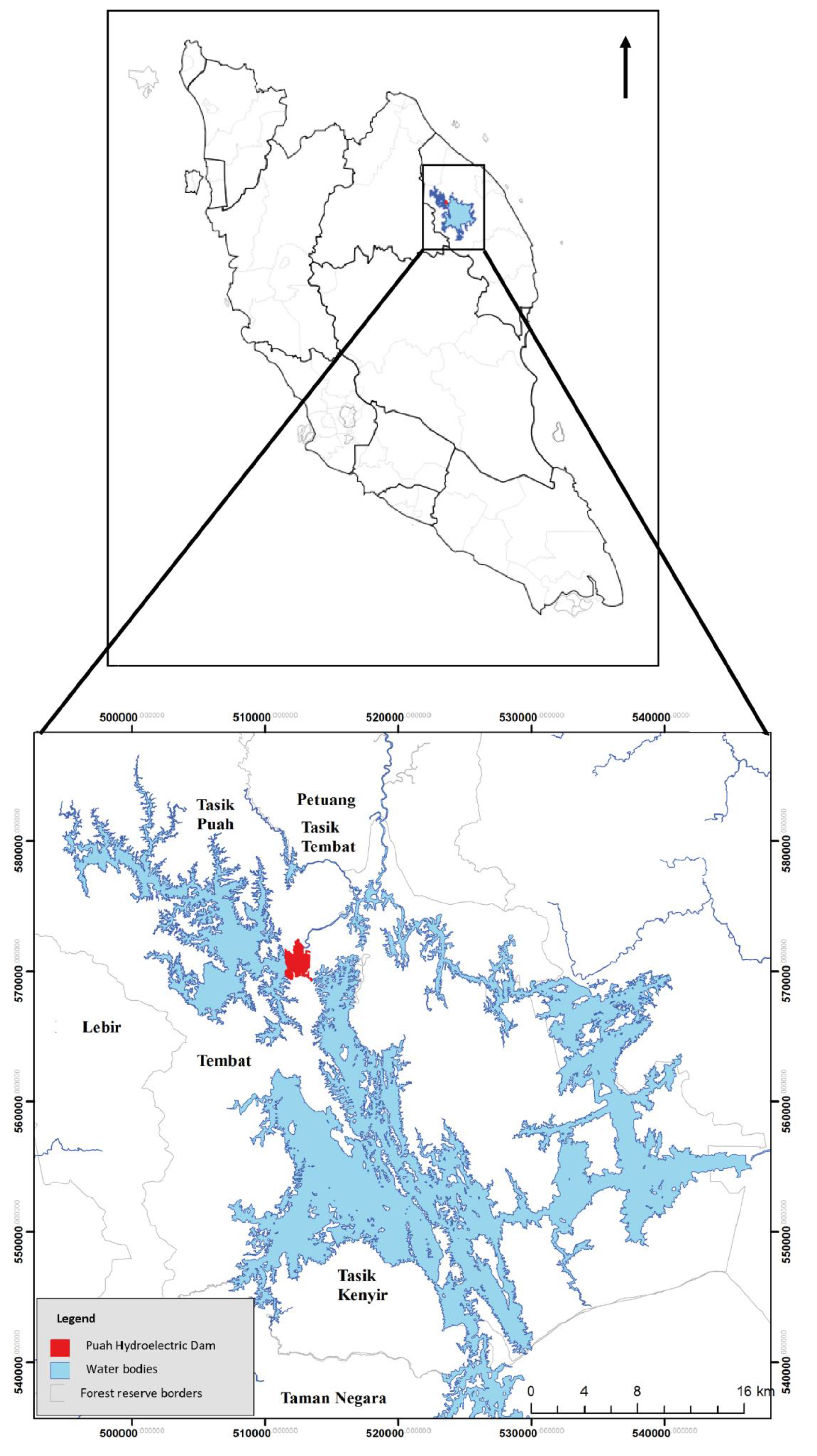
2.1. Study Area
2.2. Plant Materials and Experimental Design
2.3. Data Collection
2.3.1. Growth Parameters
2.3.2. Photosynthetic Parameters
2.4. Data Analysis
3. Results
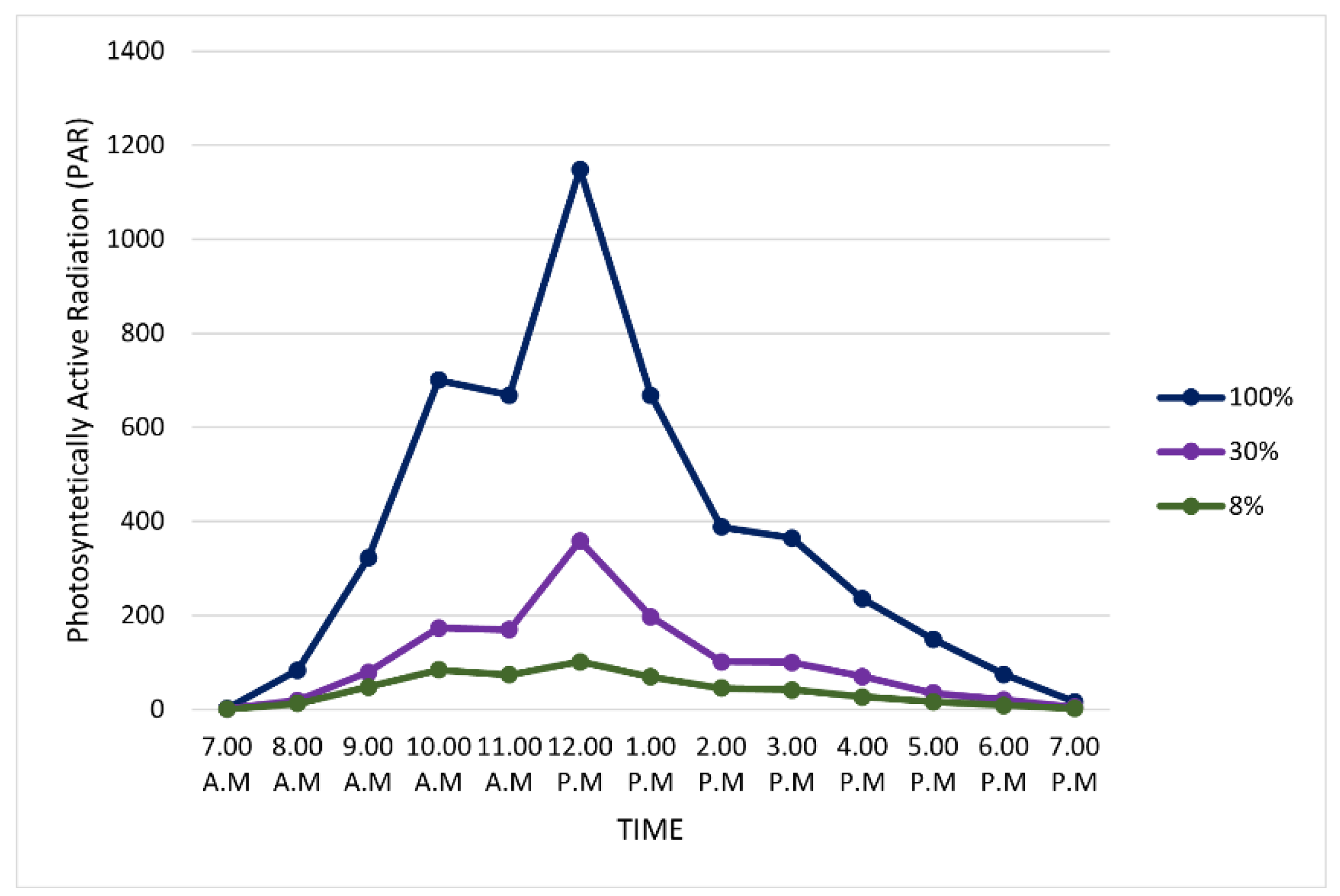
3.1. Growth and Photosynthetic Responses of S. leprosula and S. acuminata Grown under Different Light Irradiance
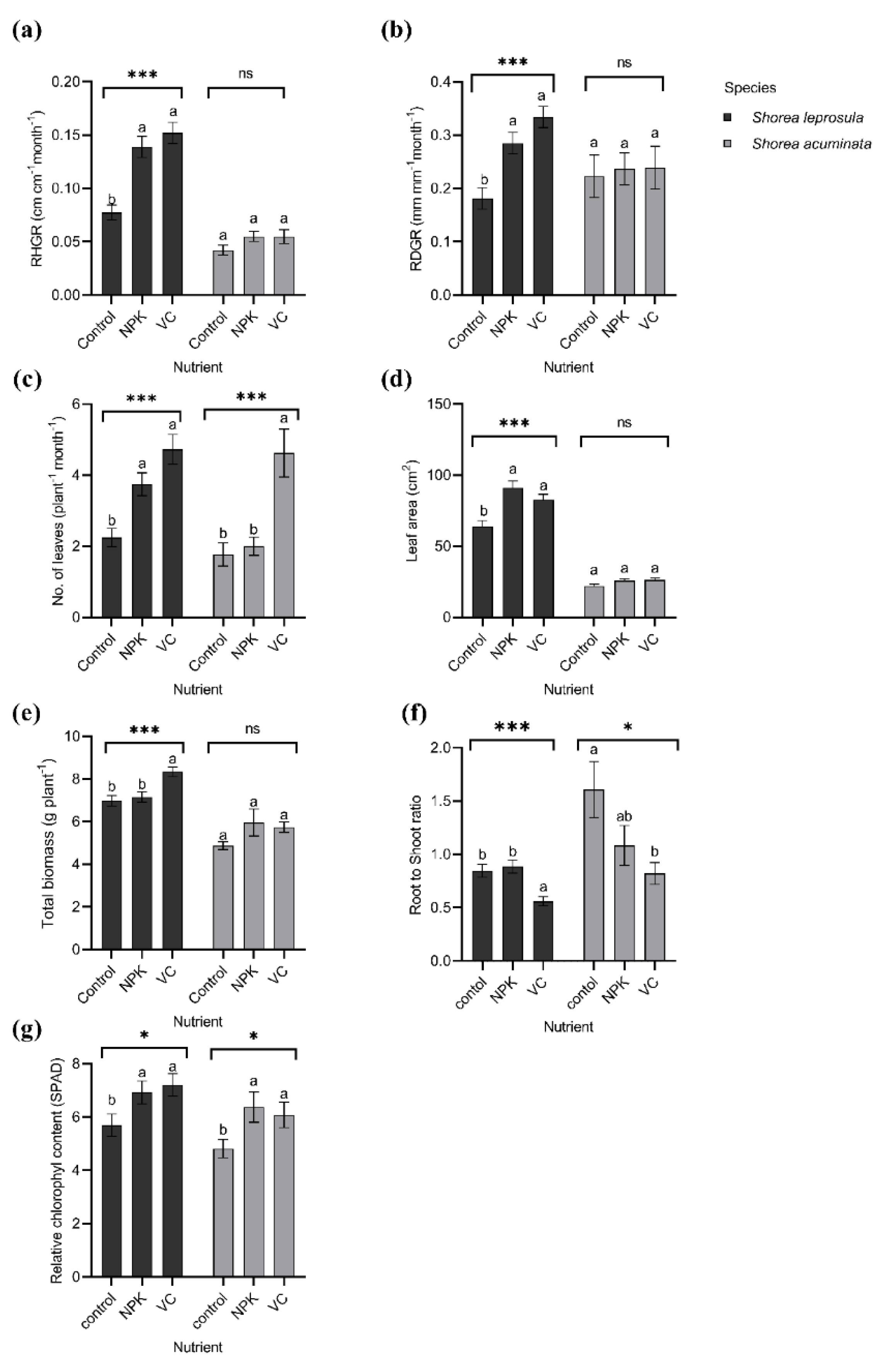
3.2. Growth and Photosynthetic Responses of S. leprosula and S. acuminata Grown in Different Nutrient Amendments
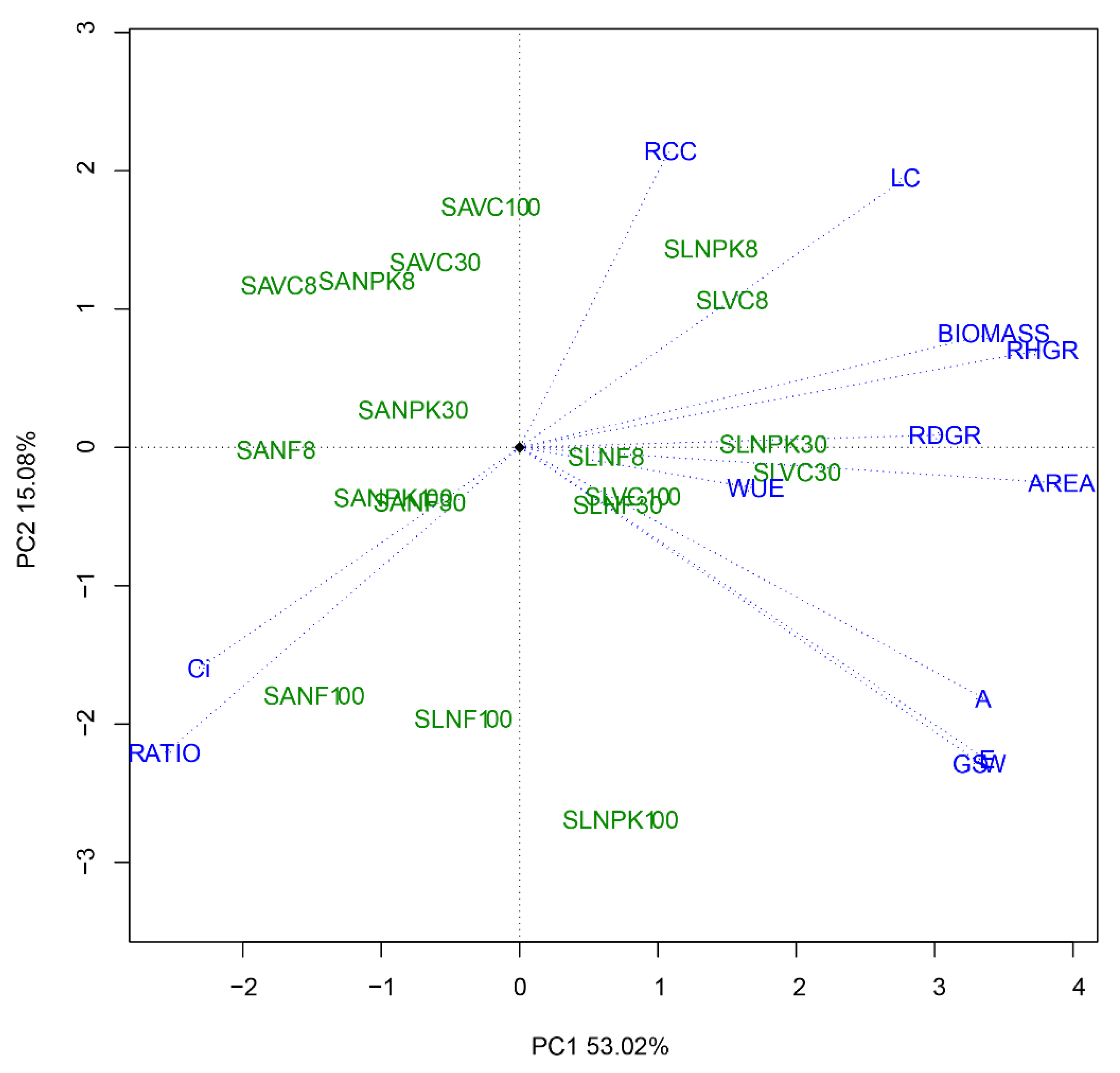
3.3. Establishing the Optimum Level of Light Irradiance and Nutrient Amendments
4. Discussion
4.1. Growth and Photosynthetic Responses of S. leprosula and S. acuminata Grown in Different Light Irradiances
4.2. Growth and Photosynthetic Responses of S. leprosula and S. acuminata Grown in Different Nutrient Amendments
4.3. Interaction between Species or Factors
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slik, J.; Poulsen, A.; Ashton, P.; Cannon, C.; Eichhorn, K.; Kartawinata, K.; Lanniari, I.; Nagamasu, H.; Nakagawa, M.; van Nieuwstadt, M.; et al. A Floristic Analysis of the Lowland Dipterocarp Forests of Borneo. J. Biogeogr. 2003, 30, 1517–1531. [Google Scholar] [CrossRef]
- Brearley, F.Q.; Banin, L.F.; Saner, P. The Ecology of the Asian Dipterocarps. Plant Ecol. Divers. 2016, 9, 429–436. [Google Scholar] [CrossRef]
- Ghazoul, J. A Short and Sweet (but Not Too Sweet), Introduction to Our Forests Forests: A Very Short Introduction. Biotopica 2016, 48, 927. [Google Scholar]
- Appanah, S.; Ashton, M.; Bawa, K.; Curtet, L.; Elouard, C.; Jantan, I.; Krishnapillay, B.; Lee, S.; Maury-Lechon, G.; Shiva, M.; et al. A Review of Dipetrocarps: Taxonomy, Ecology and Silviculture; Center for International Forestry Research: Bogor, Indonesia, 1998. [Google Scholar]
- Fatma, N.; Wan Juliana, W.A.; Shaharuddin, M.I.; Wickneswari, R. Stand Structure of Shorea and Spatial Distribution of Shorea Acuminata in a Rehabilitated Area of Kenaboi Forest Reserve. J. Trop. For. Sci. 2020, 32, 70–80. [Google Scholar] [CrossRef]
- Ashton, P. Dipterocarpaceae. In Flowering Plants. Dicotyledons. The Families and Genera of Vascular Plants; Kubitzki, K., Bayer, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; Volume 5, pp. 182–197. [Google Scholar] [CrossRef]
- FAO. Assessing Forest Degradation: Towards the Development of Globally Applicable Guidelines; FAO: Rome, Italy, 2011; p. 177. [Google Scholar]
- Mongabay. Deforestation Statistics for Malaysia. Available online: Rainforest.mongabay.com (accessed on 12 December 2020).
- Kira, T. Forest Ecosystems of East and Southeast Asia in a Global Perspective. Ecol. Res. 1991, 6, 185–200. [Google Scholar] [CrossRef]
- International Union for Conservation Nature. Bonn Challenge. Available online: https://www.bonnchallenge.org/ (accessed on 21 August 2021).
- United Nations. The Sustainable Development Goals Report 2021; UN: New York, NY, USA, 2021. [Google Scholar]
- United Nations. Quick guide to the aichi biodiversity targets. In Convention on Biological Divresity; UN: New York, NY, USA, 2013; pp. 1–42. [Google Scholar]
- UNFCCC. Key Decisions Relevant for Reducing Emissions from Deforestation and Forest Degradation in Developing Countries (REDD+); UNFCCC: Bonn, Germany, 2014. [Google Scholar]
- Kementerian Tenaga dan Sumber Asli. Penghijauan Malaysia: Kempen Penanaman 100 Juta Pokok. Available online: https://100jutapokok.gov.my/ (accessed on 11 September 2021).
- Kobayashi, S. Landscape Rehabilitation of Degraded Tropical Forest Ecosystems: Case Study of CIFOR/Japan Project in Indonesia and Peru. For. Ecol. Manag. 2004, 201, 13–22. [Google Scholar] [CrossRef]
- Kettel, J. Ecological Considerations for Using Dipterocarps for Restoration of Lowland Rainforest in Southeast Asia. Biodivers. Conserv. 2010, 19, 1137–1151. [Google Scholar] [CrossRef]
- Daisuke, H.; Tanaka, K.; Joseph Jawa, K.; Ikuo, N.; Katsutoshi, S. Rehabilitation of Degraded Tropical Rainforest Using Dipterocarp Trees in Sarawak, Malaysia. Int. J. For. Res. 2013, 2013, 683017. [Google Scholar] [CrossRef] [Green Version]
- Widiyatno, W.; Soekotjo, S.; Naiem, M.; Purnomo, S.; Setiyanto, P.E. Early Performance of 23 Dipterocarp Species Planted in Logged-over Rainforest. J. Trop. For. Sci. 2014, 26, 259–266. [Google Scholar]
- Crouzeilles, R.; Ferreira, M.S.; Chazdon, R.L.; Lindenmayer, D.B.; Sansevero, J.B.B.; Monteiro, L.; Iribarrem, A.; Latawiec, A.E.; Strassburg, B.B.N. Ecological Restoration Success Is Higher for Natural Regeneration than for Active Restoration in Tropical Forests. Sci. Adv. 2017, 3, e1701345. [Google Scholar] [CrossRef] [Green Version]
- Gallegos, S.C.; Hensen, I.; Schleuning, M. Secondary Dispersal by Ants Promotes Forest Regeneration after Deforestation. J. Ecol. 2014, 102, 659–666. [Google Scholar] [CrossRef]
- Charles, L.S.; Dwyer, J.M.; Smith, T.J.; Connors, S.; Marschner, P.; Mayfield, M.M. Species Wood Density and the Location of Planted Seedlings Drive Early-Stage Seedling Survival during Tropical Forest Restoration. J. Appl. Ecol. 2018, 55, 1009–1018. [Google Scholar] [CrossRef]
- Blakesley, D.; Anusarnsunthorn, V.; Kerby, J.; Navakitbumrung, P.; Kuarak, C.; Zangkum, S.; Hadwick, K.; Elliott, S. Nursery technology and tree species selection for restoring forest biodiversity in Northern Thailand. In Forest Restoration for Wildlife Conservation; Elliott, S., Kerby, J., Blakesley, D., Hardwick, K., Woods, K., Anusarnsunthorn, V., Eds.; International Tropical Timber Organization dan Forest: Chiang Mai, Thailand, 2000; pp. 207–222. [Google Scholar]
- Rhie, Y.H.; Lee, S.Y.; Jung, H.H.; Kim, K.S. Light Intensity Influences Photosynthesis and Crop Characteristics of Jeffersonia Dubia. Korean J. Hortic. Sci. Technol. 2014, 32, 584–589. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, Ü.; Sun, Z.; Talts, E. Controls of the Quantum Yield and Saturation Light of Isoprene Emission in Different-Aged Aspen Leaves. Plant Cell Environ. 2015, 38, 2707–2720. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.; Chiykunova, O.; Zhigalova, T.; Solovchenko, A. In Situ Optical Properties of Foliar Flavonoids: Implication for Non-Destructive Estimation of Flavonoid Content. J. Plant Physiol. 2017, 218, 258–264. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Photosynthesis: Physiological and ecological considerations. In Plant Physiology; Sinauer Associates: Sunderland, UK, 2006; pp. 249–264. [Google Scholar]
- Yong, J.W.H.; Ng, Y.F.; Tan, S.N.; Chew, A.Y.L. Effect of Fertilizer Application on Photosynthesis and Oil Yield of Jatropha curcas L. Photosynthetica 2010, 48, 208–218. [Google Scholar] [CrossRef]
- Lüttge, U. Plant physiology. In Encyclopedia of Ecology; Fath, B., Ed.; Elsevier Inc.: Darmstadt, Germany, 2019; pp. 549–557. [Google Scholar] [CrossRef]
- Sasaki, S. Ecology and physiology of Dipterocarpaceae. In Plantation Technology in Tropical Forest Science; Springer: Berlin/Heidelberg, Germany, 2006; pp. 3–22. [Google Scholar] [CrossRef]
- Turner, I.M. The Ecology of Trees in the Tropical Rain Forest, 1st ed.; Ashton, P., Hubbell, S., Janzen, D., Raven, P., Tomlinson, P., Eds.; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Kenzo, T.; Yoneda, R.; Azani, M.A. Artificial Shade Shelters Mitigate Harsh Microclimate Conditions and Enhance Growth in Tropical Tree Seedlings Planted in Degraded Land. Tropics 2020, 29, 121–132. [Google Scholar] [CrossRef]
- Kenzo, T.; Yoneda, R.; Matsumoto, Y.; Mohamad Azani, A.; Nik Majid, M. Growth and Photosynthetic Response of Four Malaysian Indigenous Tree Species under Different Light Conditions. J. Trop. For. Sci. 2011, 23, 271–281. [Google Scholar]
- Pan, J.; Guo, B. Effects of Light Intensity on the Growth, Photosynthetic Characteristics, and Flavonoid Content of Epimedium Pseudowushanense B.L.Guo. Molecules 2016, 21, 1475. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Shen, Z.; Liu, Y.; Wang, L.; Hannaway, D.; Lu, H. Effects of Shade Treatments on the Photosynthetic Capacity, Chlorophyll Fluorescence, and Chlorophyll Content of Tetrastigma Hemsleyanum Diels et Gilg. Environ. Exp. Bot. 2009, 65, 177–182. [Google Scholar] [CrossRef]
- Idris, A.; Tun, U.; Onn, H.; Linatoc, A.; Tun, U.; Onn, H.; Fadzelly, M.; Bakar, A. Effect of Light Intensity on the Photosynthesis and Stomatal Density of Selected Plant Species of Gunung Ledang, Johor. Malays. Appl. Biol. 2019, 48, 133–140. [Google Scholar]
- Liu, M.; Gong, J.; Yang, B.; Ding, Y.; Zhang, Z.; Wang, B.; Zhu, C.; Hou, X. Differences in the Photosynthetic and Physiological Responses of Leymus Chinensis to Different Levels of Grazing Intensity. BMC Plant Biol. 2019, 19, 558. [Google Scholar] [CrossRef]
- Formisano, L.; Ciriello, M.; Cirillo, V.; Pannico, A.; El-nakhel, C.; Cristofano, F.; Duri, L.G.; Giordano, M.; Rouphael, Y.; De Pascale, S. Divergent Leaf Morpho-Physiological and Anatomical Adaptations of Four Lettuce Cultivars in Response to Different Greenhouse Irradiance Levels in Early Summer Season. Plants 2021, 10, 1179. [Google Scholar] [CrossRef]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmülling, T.; Cortleven, A. Light Acts as a Stressor and Influences Abiotic and Biotic Stress Responses in Plants. Plant Cell Environ. 2021, 44, 645–664. [Google Scholar] [CrossRef]
- Usuda, H. Evaluation of the Effect of Photosynthesis on Biomass Production with Simultaneous Analysis of Growth and Continuous Monitoring of CO2 Exchange in the Whole Plants of Radish, Cv Kosena under Ambient and Elevated CO2. Plant Prod. Sci. 2004, 7, 386–396. [Google Scholar] [CrossRef] [Green Version]
- Bhatla, S.C.; Lal, M.A. Plant Physiology, Development and Metabolism; Springer: Singapore, 2018. [Google Scholar] [CrossRef]
- Farquhar, G.; von Caemmerer, S.; Berry, J. A Biochemical Model of Photosynthetic CO2 Assimilation in Leaves of C3 Species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Driesen, E.; Van den Ende, W.; De Proft, M.; Saeys, W. Influence of Environmental Factors Light, Co2, Temperature, and Relative Humidity on Stomatal Opening and Development: A Review. Agronomy 2020, 10, 1975. [Google Scholar] [CrossRef]
- Lynch, J.; Marschner, P.; Rengel, Z. Effect of internal and external factors on root growth and development. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 331–346. [Google Scholar] [CrossRef]
- Blouin, M.; Barrere, J.; Meyer, N.; Lartigue, S.; Barot, S.; Mathieu, J. Vermicompost Significantly Affects Plant Growth. A Meta-Analysis. Agron. Sustain. Dev. 2019, 39, 34–50. [Google Scholar] [CrossRef]
- Wong, W.S.; Zhong, H.T.; Cross, A.T.; Yong, J.W.H. Plant biostimulants in vermicomposts: Characteristics and plausible mechanisms. In The Chemical Biology of Plant Biostimulants; Gelen, D., Xu, L., Eds.; John Wiley & Sons Ltd.: Singapore, 2020; pp. 156–180. [Google Scholar]
- Devi, C.; Khwairakpam, M. Vermicompost for sustainable agriculture and bioconversion of terrestrial weed biomass into vermicompost. In New Generation of Organic Fertilizers; Turan, M., Yildiri, E., Eds.; IntechOpen: London, UK, 2022. [Google Scholar]
- Lim, S.L.; Wu, T.Y.; Lim, P.N.; Shak, K.P.Y. The Use of Vermicompost in Organic Farming: Overview, Effects on Soil and Economics. J. Sci. Food Agric. 2015, 95, 1143–1156. [Google Scholar] [CrossRef]
- Arancon, N.Q.; Solarte, Z.; Arancon, N.Q.; Solarte, Z. Vermiculture in Greenhouse Plants, Field Crop Production, and Hydroponics; Oxford University Press: Oxford, UK, 2019. [Google Scholar] [CrossRef]
- Xu, C.; Mou, B. Vermicompost Affects Soil Properties and Spinach Growth, Physiology, and Nutritional Value. HortScience 2016, 51, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Lal, R. Soil Carbon Sequestration Impacts on Global Climate Change and Food Security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [Green Version]
- Dignac, M.F.; Derrien, D.; Barré, P.; Barot, S.; Cécillon, L.; Chenu, C.; Chevallier, T.; Freschet, G.T.; Garnier, P.; Guenet, B.; et al. Increasing Soil Carbon Storage: Mechanisms, Effects of Agricultural Practices and Proxies. A Review. Agron. Sustain. Dev. 2017, 37, 14. [Google Scholar] [CrossRef] [Green Version]
- Kirschbaum, M.U.F.F. Does Enhanced Photosynthesis Enhance Growth? Lessons Learned from CO2 Enrichment Studies. Plant Physiol. 2011, 155, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Longstreth, D.J.; Nobel, P.S. Nutrient Influences on Leaf Photosynthesis. Plant Physiol. 1980, 65, 541–543. [Google Scholar] [CrossRef] [Green Version]
- Harpole, W.S.; Ngai, J.T.; Cleland, E.E.; Seabloom, E.W.; Borer, E.T.; Bracken, M.E.S.; Elser, J.J.; Gruner, D.S.; Hillebrand, H.; Shurin, J.B.; et al. Nutrient Co-Limitation of Primary Producer Communities. Ecol. Lett. 2011, 14, 852–862. [Google Scholar] [CrossRef]
- Ingestad, T.; Lund, A. Nitrogen Stress in Birch Seedlings: II. N, K, P, Ca, and Mg Nutrition. Physiol. Plant. 1979, 45, 149–157. [Google Scholar] [CrossRef]
- Knecht, M.F.; Göransson, A. Terrestrial Plants Require Nutrients in Similar Proportions. Tree Physiol. 2004, 24, 447–460. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J.; Yavitt, J.B.; Wurzburger, N.; Turner, B.I.; Tanner, E.V.J.; Sayer, E.J.; Santiago, L.S.; Kaspari, M.; Hedin, L.O.; Harms, K.E.; et al. Potassium, Phosphorus, or Nitrogen Limit Root Allocation, Tree Growth, or Litter Production in a Lowland Tropical Forest. Ecology 2011, 92, 1616–1625. [Google Scholar] [CrossRef]
- Pamoengkas, P.; Zamzam, A.; Dwisutono, A. Vegetation Recovery of Logged-over Dipterocarp Forests In Central Kalimantan, Indonesia. Floresta Ambient. 2019, 26, e20171239. [Google Scholar] [CrossRef]
- Widiyatno; Hidayati, F.; Hardiwinoto, S.; Indrioko, S.; Purnomo, S.; Jatmoko; Tani, N.; Naiem, M. Selection of Dipterocarp Species for Enrichment Planting in a Secondary Tropical Rainforest. For. Sci. Technol. 2020, 16, 206–215. [Google Scholar] [CrossRef]
- Ghazoul, J. Dipterocarp Biology, Ecology, and Conservation, 1st ed.; Oxford University Press: New York, NY, USA, 2016. [Google Scholar]
- Schmitt, J.; Wulff, R.D. Light Spectral Quality, Phytochrome and Plant Competition. Trends Ecol. Evol. 1993, 8, 47–51. [Google Scholar] [CrossRef]
- Newman, M.; Burgess, P.; Whitmore, T. Manuals of Dipterocarps for Foresters, Borneo Island Light Hardwoods: Anisoptera, Parashorea, Shorea (Red, White and Yellow Meranti); Royal Botanic Garden: Edinburgh, UK, 1996. [Google Scholar]
- Hirasawa, Y. Pasoh Ecology of a Lowland Rain Forest in Southeast Asia, 1st ed.; Okuda, T., Manokaran, N., Matsumoto, Y., Niiyama, K., Thomas, S., Ashton, P., Eds.; Springer: Tokyo, Japan, 2003. [Google Scholar]
- Suzuki, R.O.; Numata, S.; Okuda, T.; Nur Supardi, M.N.; Kachi, N. Growth Strategies Differentiate the Spatial Patterns of 11 Dipterocarp Species Coexisting in a Malaysian Tropical Rain Forest. J. Plant Res. 2009, 122, 81–93. [Google Scholar] [CrossRef]
- Van Volkenburgh, E. Leaf Expansion—An Integrating Plant Behaviour. Plant Cell Environ. 1999, 22, 1463–1473. [Google Scholar] [CrossRef]
- Mathur, S.; Jain, L.; Jajoo, A. Photosynthetic Efficiency in Sun and Shade Plants. Photosynthetica 2018, 56, 354–365. [Google Scholar] [CrossRef]
- Terashima, I.; Hanba, Y.T.; Tholen, D.; Niinemets, Ü. Leaf Functional Anatomy in Relation to Photosynthesis. Plant Physiol. 2011, 155, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Pasquini, S.C.; Santiago, L.S. Nutrients Limit Photosynthesis in Seedlings of a Lowland Tropical Forest Tree Species. Oecologia 2012, 168, 311–319. [Google Scholar] [CrossRef]
- Chapin, F.S.; Schulze, E.D.; Mooney, H.A. The Ecology and Economics of Storage in Plants. Annu. Rev. Ecol. Syst. 1990, 21, 423–447. [Google Scholar] [CrossRef]
- Craine, J.M.; Dybzinski, R. Mechanisms of Plant Competition for Nutrients, Water and Light. Funct. Ecol. 2013, 27, 833–840. [Google Scholar] [CrossRef]
- Connolly, J. On the Use of Response Models in Mixture Experiments. Oecologia 1987, 72, 95–103. [Google Scholar] [CrossRef]
- Connolly, J.; Wayne, P.; Bazzaz, F.A. Interspecific Competition in Plants: How Well Do Current Methods Answer Fundamental Questions? Am. Nat. 2001, 157, 107–125. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Significance | Means | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Light Irradiance (LI) | Nutrient | Species × LI | Species × Nutrient | LI × Nutrient | 100% | 30% | 8% | NF | NPK | VC | |
| RHGR (cm cm−1 month−1) | *** | *** | *** | ** | ns | ns | 0.056 | 0.079 | 0.064 | 0.051 | 0.073 | 0.075 |
| RDGR (mm mm−1 month−1) | *** | *** | *** | *** | ns | ns | 0.13 | 0.13 | 0.11 | 0.10 | 0.13 | 0.14 |
| Mean number of leaves (plant−1 month−1) | ** | ns | *** | *** | ns | ns | 2.94 | 3.65 | 2.90 | 2.02 | 2.96 | 4.67 |
| Leaf area (cm2) | *** | *** | *** | *** | *** | ns | 43.8 | 57.6 | 55.4 | 43.3 | 58.5 | 54.9 |
| Biomass (g plant−1) | *** | ns | ** | *** | ns | * | 6.42 | 6.28 | 6.96 | 5.98 | 6.59 | 7.12 |
| Root-to-shoot ratio | *** | *** | *** | * | ns | ** | 1.26 | 0.75 | 0.88 | 1.21 | 0.98 | 0.69 |
| Traits | Significance | Means | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Light Irradiance (LI) | Nutrient | Species × LI | Species × Nutrient | LI × Nutrient | 100% | 30% | 8% | NF | NPK | VC | |
| RCC (SPAD) | ** | *** | *** | ns | ns | ns | 4.65 | 6.04 | 8.00 | 5.27 | 6.65 | 6.69 |
| A (µmol m−2 s−1) | *** | ns | * | ns | ns | ns | 7.84 | 8.56 | 7.07 | 7.65 | 8.78 | 7.04 |
| E (mmol−2 s−1) | *** | ** | ns | ns | ns | ns | 0.0027 | 0.0028 | 0.0022 | 0.0027 | 0.0026 | 0.0024 |
| Ci (µmol mol−1) | ns | ns | ns | ns | ns | ns | 282 | 217 | 216 | 247 | 221 | 249 |
| gsw (mol m−2 s−1) | *** | ** | ns | ns | ns | ns | 0.16 | 0.14 | 0.13 | 0.16 | 0.16 | 0.14 |
| WUE (µmol CO2 mmol−1 H2O) | ns | ns | ns | ns | ns | ns | 0.00048 | 0.00049 | 0.00039 | 0.00042 | 0.00037 | 0.00058 |
| Abbreviation | PC1 | PC2 | |
|---|---|---|---|
| Relative height growth rate (cm cm−1 month−1) | RHGR | 0.362 | 0.127 |
| Relative diameter growth rate (mm mm−1 month−1) | RDGR | 0.294 | 0.016 |
| Mean number of leaves (plant−1 month−1) | LC | 0.269 | 0.351 |
| Leaf area (cm2) | AREA | 0.377 | −0.047 |
| Biomass (g plant−1) | BIOMASS | 0.329 | 0.149 |
| Root-to-shoot ratio | RATIO | −0.246 | −0.398 |
| Relative chlorophyll concentration (SPAD) | RCC | 0.104 | 0.386 |
| E (mmol−2 s−1) | E | 0.324 | −0.406 |
| A (µmol m−2 s−1) | A | 0.323 | −0.327 |
| Ci (µmol mol−1) | Ci | −0.223 | −0.288 |
| gsw (mol m−2 s−1) | GSW | 0.321 | −0.413 |
| WUE (µmol CO2 mmol−1 H2O) | WUE | 0.164 | −0.053 |
| Variability (%) | 53.0 | 15.1 | |
| Cumulative variability (%) | 53.0 | 68.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siti Nurfaeiza, A.R.; Wan Juliana, W.A.; Shamsul, K.; Shukor, M.N. Morpho-Physiological Strategies of Shorea leprosula Miq. and Shorea acuminata Dyer in Response to Light Intensity and Nutrient Amendments. Forests 2022, 13, 1768. https://doi.org/10.3390/f13111768
Siti Nurfaeiza AR, Wan Juliana WA, Shamsul K, Shukor MN. Morpho-Physiological Strategies of Shorea leprosula Miq. and Shorea acuminata Dyer in Response to Light Intensity and Nutrient Amendments. Forests. 2022; 13(11):1768. https://doi.org/10.3390/f13111768
Chicago/Turabian StyleSiti Nurfaeiza, Abd Razak, Wan Ahmad Wan Juliana, Khamis Shamsul, and Md. Nor Shukor. 2022. "Morpho-Physiological Strategies of Shorea leprosula Miq. and Shorea acuminata Dyer in Response to Light Intensity and Nutrient Amendments" Forests 13, no. 11: 1768. https://doi.org/10.3390/f13111768