
The Effect of Intercropping Mulberry (Morus alba L.) with Peanut (Arachis hypogaea L.), on the Soil Rhizosphere Microbial Community

Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Varieties and Planting Patterns
2.2. Collection and Chemical Property Analysis of Soil Samples
2.3. DNA Extraction and Sequencing of Soil Microorganisms
2.4. Statistical Analysis
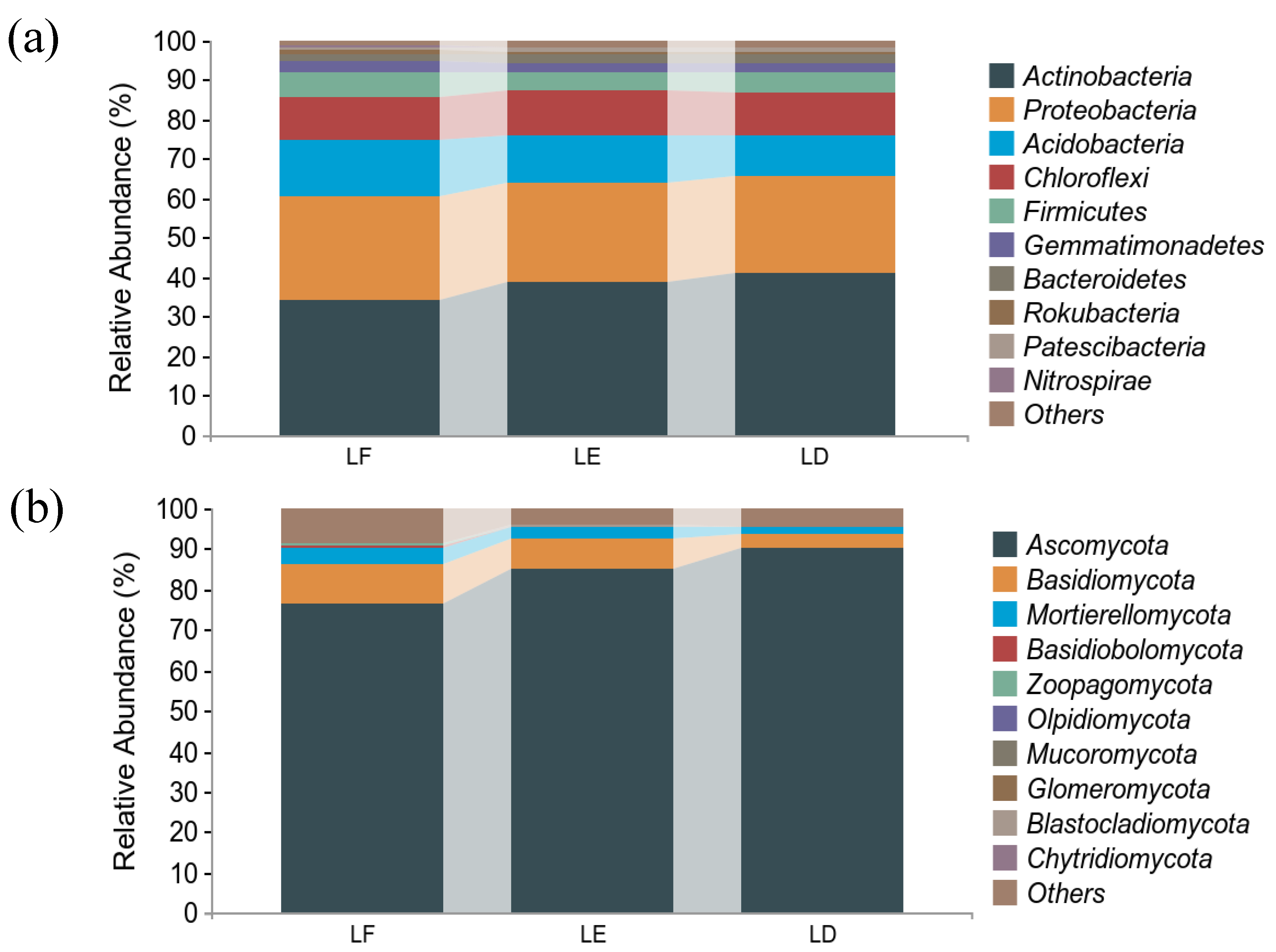
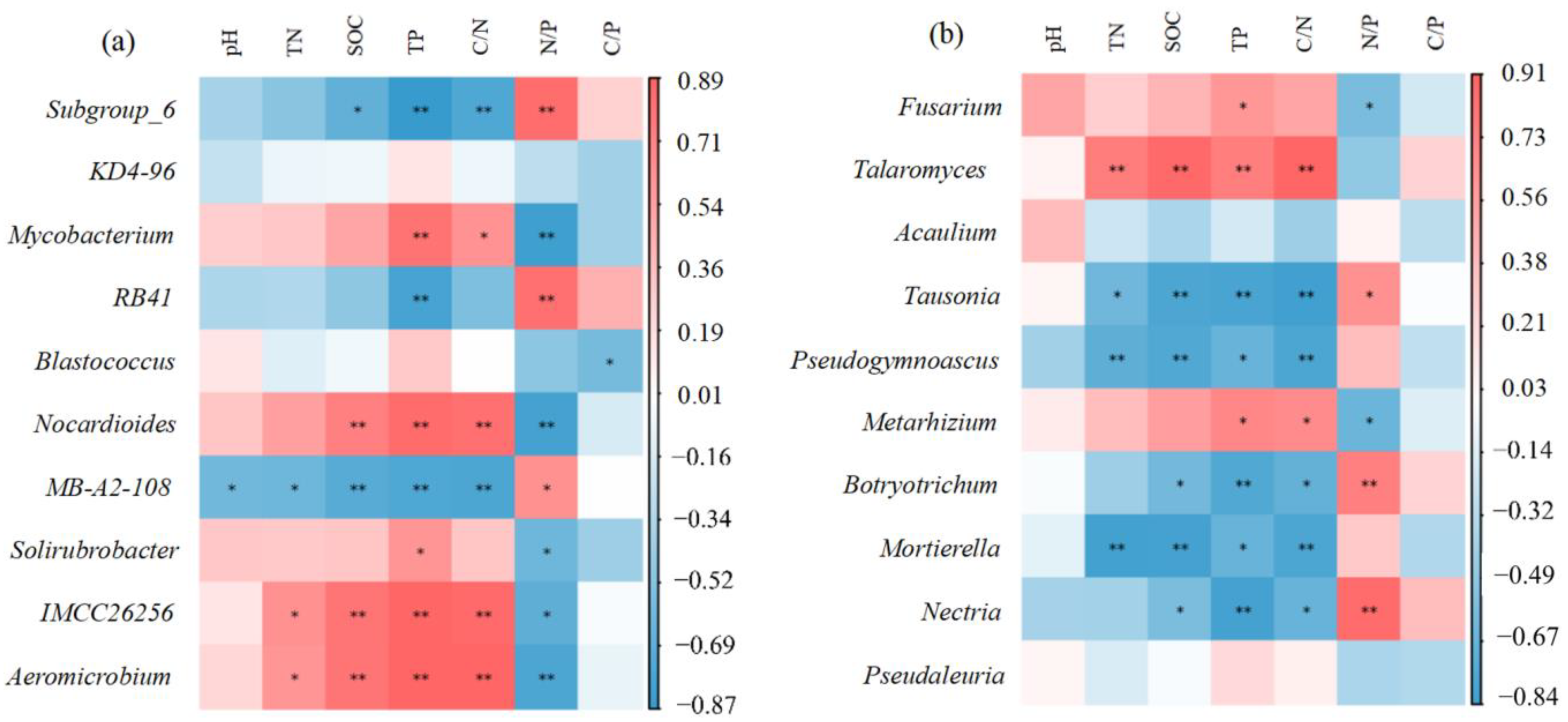
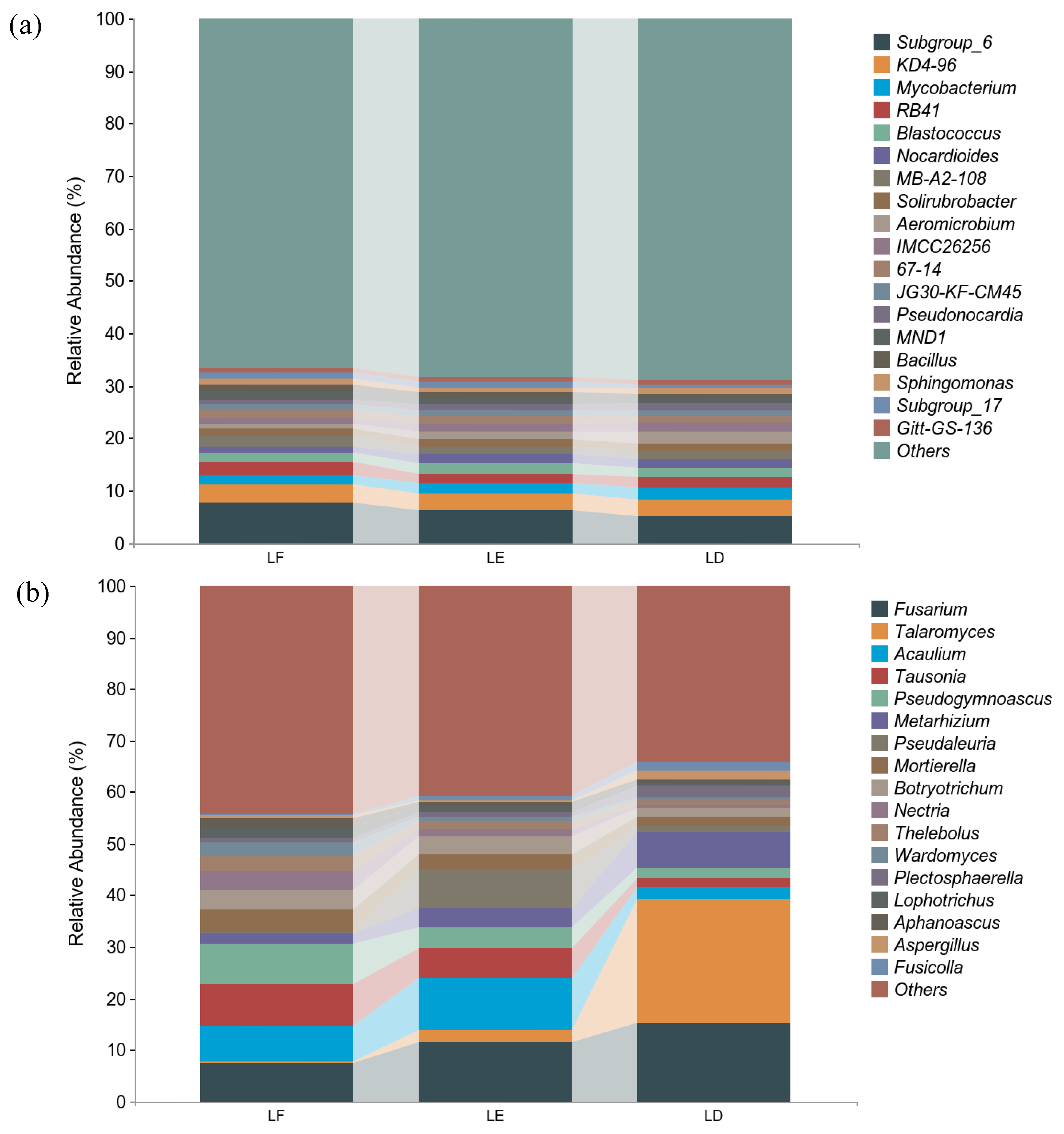
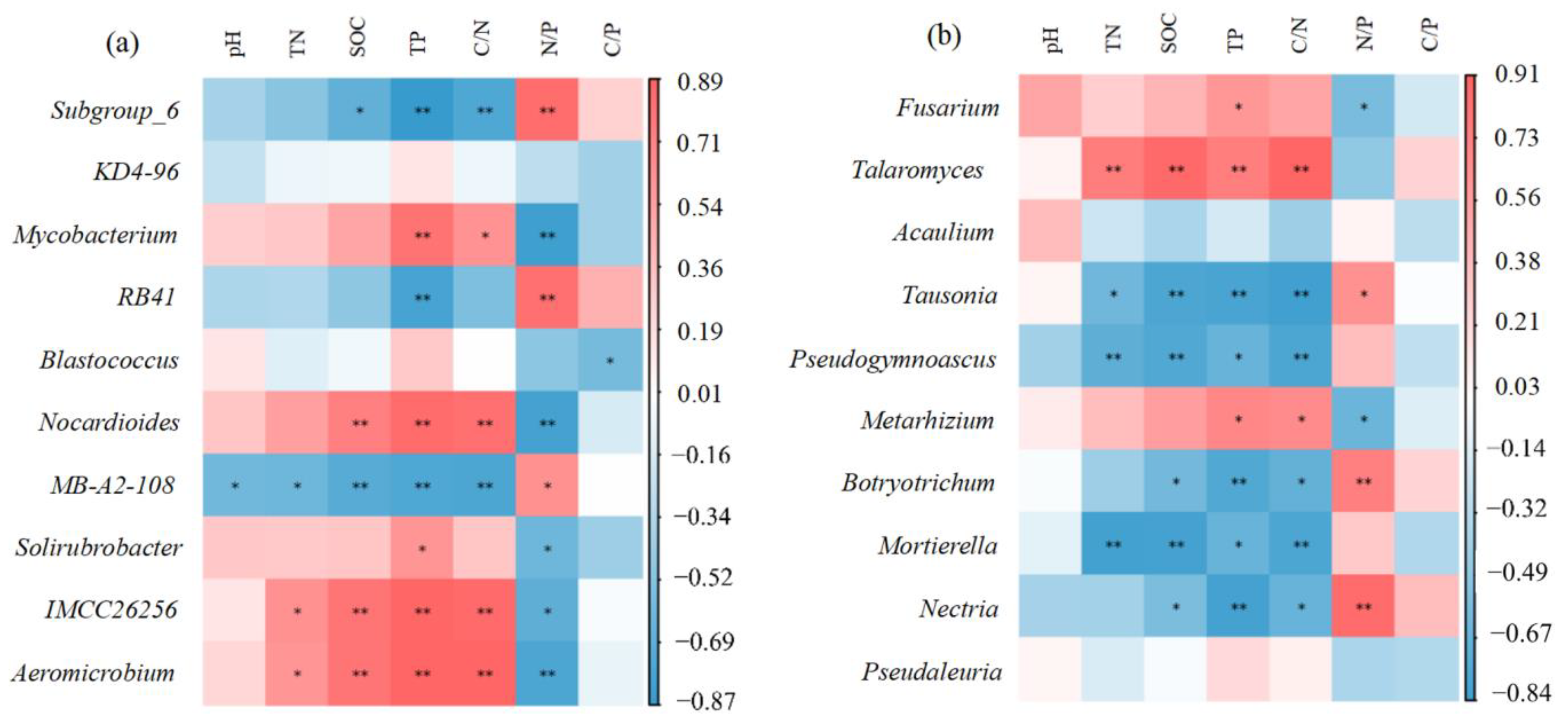
3. Results
3.1. Effects of Intercropping on Soil Carbon, Nitrogen and Phosphorus Stoichiometry
3.2. Correlation Analysis of Relative Abundances of Soil Microbial Communities and Soil Properties
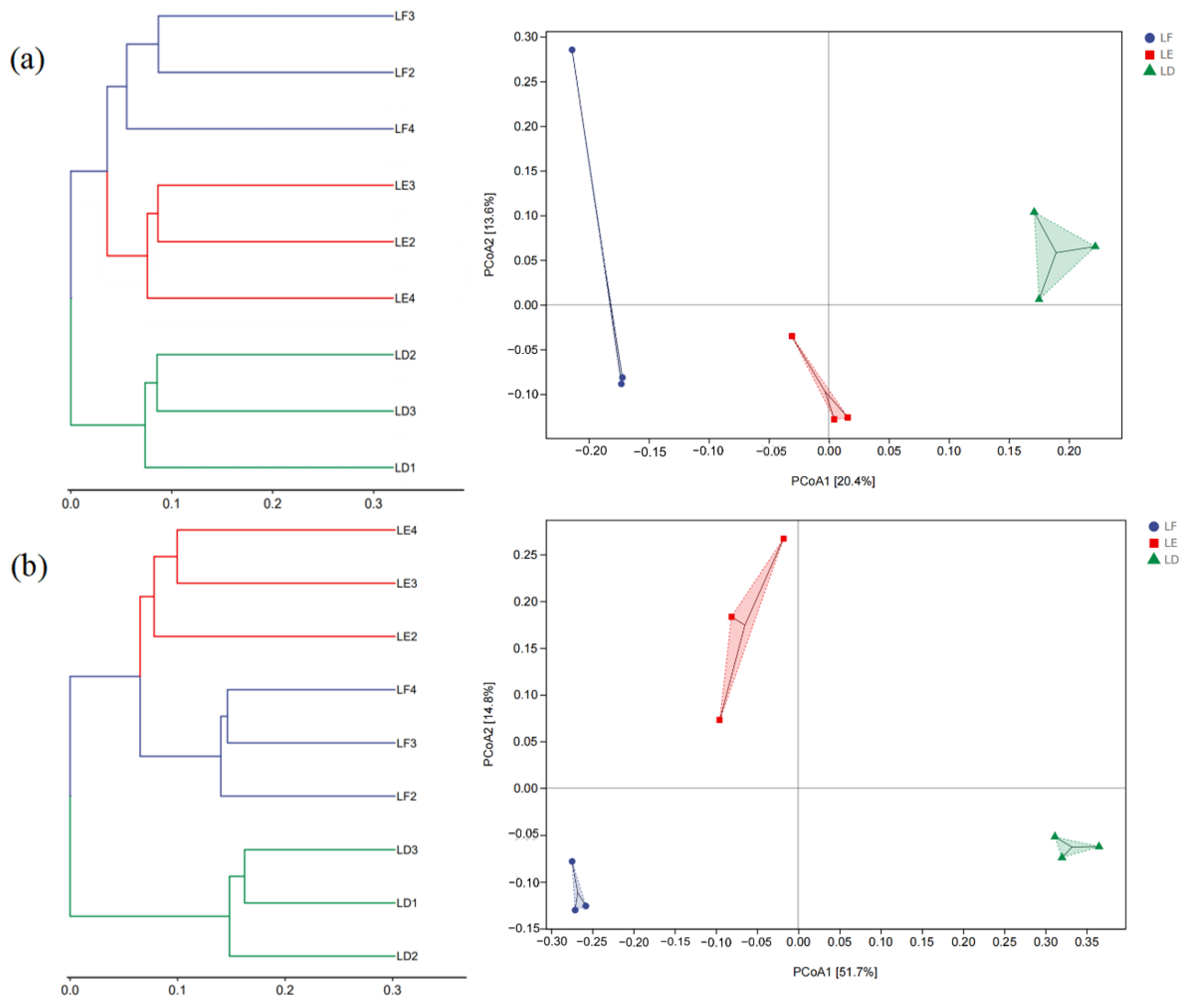
3.3. Structural Diversity of Soil Microbial Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jovino, R.S.; Rosa, S.T.; Terezinha, R.R.; Jackson, S.C.R.; Barboza, C.J.A.; Maria, L.L.; Cavalcanti, S.R.; Rosália, S.C.E.; Rose, R.P.; Santiago, F.A.D.; et al. Elite Bradyrhizobium strains boost biological nitrogen fixation and peanut yield in tropical drylands. Braz. J. Microbiol. 2022, 53, 1623–1632. [Google Scholar] [CrossRef] [PubMed]
- Toomer, O.T. Nutritional chemistry of the peanut (Arachis hypogaea). Crit. Rev. Food 2017, 58(17), 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, X.G.; Kevin, P.B.; Yao, X.D.; Devin, C.D.; Ding, C.F.; Wang, X.X.; Ruan, H.H. Peanut plant growth was altered by monocropping-associated microbial enrichment of rhizosphere microbiome. Plant Soil 2020, 446, 655–669. [Google Scholar] [CrossRef]
- Wang, G.Z.; Li, X.G.; Xi, X.Q.; Cong, W.F. Crop diversification reinforces soil microbiome functions and soil health. Plant Soil 2022, 476, 375–383. [Google Scholar] [CrossRef]
- Bamboriya, S.D.; Bana, R.S.; Kuri, B.R.; Kumar, V.; Bamboriya, S.D.; Meena, R.P. Achieving higher production from low inputs using synergistic crop interactions under maize-based polyculture systems. Environ. Sustain. 2022, 5, 145–159. [Google Scholar] [CrossRef]
- Namazi, Y.; Rezaei, C.E.; Siavash, M.S.; Leonardo, B.M. The effects of microbial inoculation and intercropping on yield and active ingredients of savory (Satureja hortensis L.) intercropped with common bean (Phaseolus vulgaris L.). Int. J. Environ. Sci. 2022, 19, 8273–8288. [Google Scholar] [CrossRef]
- Feng, C.; Sun, Z.X.; Zhang, L.Z.; Feng, L.S.; Zheng, J.M.; Bai, W.; Gu, C.F.; Wang, Q.; Xu, Z.; Wopke, W. Maize/peanut intercropping increases land productivity: A meta-analysis. Field Crop Res. 2021, 270, 108208. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Khan, M.U.; Lin, X.Q.; Lin, Z.M.; Lin, S.S.; Lin, W.X. Evaluation of maize/peanut intercropping effects on microbial assembly, root exudates and peanut nitrogen uptake. Plant Physiol. Bioch. 2022, 171, 75–83. [Google Scholar] [CrossRef]
- Surki, A.A.; Nazari, M.; Fallah, S.; Iranipour, R. Improvement of the soil properties, nutrients, and carbon stocks in different cereal–legume agroforestry systems. Int. J. Environ. Sci. 2020, 18, 1–8. [Google Scholar]
- Tang, X.M.; Zhong, R.C.; Jiang, J.; He, L.Q.; Huang, Z.P.; Shi, G.Y.; Wu, H.N.; Liu, J.; Xiong, F.Q.; Han, Z.Q.; et al. Cassava/peanut intercropping improves soil quality via rhizospheric microbes increased available nitrogen contents. BMC Biotechnol. 2020, 20, 13. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.H.; Dong, Q.Q.; Han, Y.; Zhang, K.Z.; Shi, X.L.; Yang, X.; Yuan, Y.; Zhou, D.Y.; Wang, K.; Wang, X.G.; et al. Maize/peanut intercropping improves nutrient uptake of side-row maize and system microbial community diversity. BMC Microbiol. 2022, 22, 14. [Google Scholar] [CrossRef]
- Phoenix, G.K.; Johnson, D.A.; Muddimer, S.P.; Leake, J.R.; Cameron, D.D. Niche differentiation and plasticity in soil phosphorus acquisition among co-occurring plants. Nat. Plants 2020, 6, 349–354. [Google Scholar] [CrossRef]
- Dai, C.C.; Chen, Y.; Wang, X.X.; Li, P.D. Effects of intercropping of peanut with the medicinal plant Atractylodes lancea on soil microecology and peanut yield in subtropical China. Agroforest Syst. 2013, 87, 417–426. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Putten, W.H.V.D. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Sivojiene, D.; Kacergius, A.; Baksiene, E.; Maseviciene, A.; Zickiene, L. The influence of organic fertilizers on the abundance of soil microorganism communities, agrochemical indicators, and yield in east lithuanian lightsoils. Plants 2021, 10, 2648. [Google Scholar] [CrossRef]
- He, X.J.; Xie, H.; Gao, D.M.; Khashi, U.; Rahman, M.; Zhou, X.G.; Wu, F.Z. Biochar and intercropping with potato–onion enhanced the growth and yield advantages of tomato by regulating the soil properties, nutrientuptake, and soil microbial community. Front. Microbiol. 2021, 12, 695447. [Google Scholar] [CrossRef]
- Ke, P.J.; Zee, P.C.; Fukami, T. Dynamic plant-soil microbe interactions: The neglected effect of soil conditioning time. New Phytol. 2021, 231, 1546–1558. [Google Scholar] [CrossRef]
- Cui, Y.X.; Fang, L.C.; Guo, X.B.; Wang, X.; Zhang, Y.J.; Li, P.F.; Zhang, X.C. Ecoenzymatic stoichiometry and microbial nutrient limitation in rhizosphere soil in the arid area of the northern Loess Plateau, China. Soil Biol. Biochem. 2018, 116, 11–21. [Google Scholar] [CrossRef]
- Wang, M.; Chen, S.B.; Chen, L.; Wang, D. Responses of soil microbial communities and their network interactions to saline-alkaline stress in Cd-contaminated soils. Environ. Pollut. 2019, 252, 1609–1621. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Yuan, M.S.; Tang, L.; Shen, Y.F.; Yu, Q.; Li, S.Q. Integrated microbiology and metabolomics analysis reveal responses of soil microorganisms and metabolic functions to phosphorus fertilizer on semiarid farm. Sci. Total Environ. 2022, 817, 152878. [Google Scholar] [CrossRef]
- Liu, Y.; Willison, J.H.M. Prospects for cultivating white mulberry (Morus alba) in the drawdown zone of the Three Gorges Reservoir, China. Environ Sci Pollut Res. 2013, 20, 7142–7151. [Google Scholar] [CrossRef] [PubMed]
- Durgadevi, R.; Vijayalakshmi, D. Mulberry with increased stomatal frequency regulates gas exchange traits for improved drought tolerance. Plant Physiol. Rep. 2020, 25, 24–32. [Google Scholar] [CrossRef]
- Li, R.X.; Su, X.Q.; Zhang, Y.P.; Wang, T.C. Molecular mechanism of mulberry response to drought stress revealed by complementary transcriptomic and iTRAQ analyses. BMC Plant Biol. 2022, 22, 36. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, N.; Ji, G. Responses of biomass allocation and photosynthesis in mulberry to Pb-contaminated soil. Acta Physiol. Plant 2022, 44, 43–52. [Google Scholar] [CrossRef]
- Wang, L.; Ji, G. Glutathione and calcium biomineralization of mulberry (Morus alba L.) involved in the heavy metal detoxification of lead-contaminated soil. J. Soil. Sci. Plant Nut. 2021, 21, 1182–1190. [Google Scholar] [CrossRef]
- Zhang, X.L.; Teng, Z.T.; Zhang, H.H.; Cai, D.J.; Zhang, J.Y.; Meng, F.J.; Sun, G.Y. Nitrogen application and intercropping change microbial community diversity and physicochemical characteristics in mulberry and alfalfa rhizosphere soil. J. For. Res. 2021, 32, 2121–2133. [Google Scholar] [CrossRef]
- Benjamin, J.C.; Paul, J.M.M.; Michael, J.R.; Andrew, W.H.; Amy, J.A.J.; Susan, P.H. Dada2: High-resolution sample inference from illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar]
- Zhong, H.; Liu, Y.J.; Wang, B.H.; He, M.J.; Wu, L. Effect of analysis method on 16S rRNA gene amplification and sequencing analysis results of bacterial community. Biotechnol. Bull. 2022, 38, 81–92. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern wisconsin. Ecol. Monogr. 1957, 27, 327–349. [Google Scholar] [CrossRef]
- Ramette, A. Multivariate analyses in microbial ecology. Fems. Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Luo, H.; Tang, B.; Guo, H.; Cao, Z.; Zeng, Q.; Chen, S.; Chen, Z. Dynamic changes of rhizosphere soil bacterial community and nutrients in cadmium polluted soils with soybean-corn intercropping. BMC Microbiol. 2022, 22, 57. [Google Scholar] [CrossRef]
- Bainard, L.D.; Koch, A.M.; Gordon, A.M.; Klironomos, J.N. Growth response of crops to soil microbial communities from conventional monocropping and tree-based intercropping systems. Plant Soil 2013, 363, 345–356. [Google Scholar] [CrossRef]
- Wang, J.; Liao, L.R.; Ye, Z.C.; Liu, H.F.; Zhang, C.; Zhang, L.; Liu, G.B.; Wang, G.L. Different bacterial co-occurrence patterns and community assembly between rhizosphere and bulk soils under N addition in the plant-soil system. Plant Soil 2022, 471, 697–713. [Google Scholar] [CrossRef]
- Tomohiro, R.; Arizono, I.; Takemoto, Y. Economic design of double sampling Cpm control chart for monitoring process capability. Int. J. Prod. Econ. 2020, 221, 107468. [Google Scholar] [CrossRef]
- Kuramae, E.E.; Yergeau, E.; Wong, L.C.; Pijl, A.S.; Johannes, A.V.V.; Kowalchuk, G.A. Soil characteristics more strongly influence soil bacterial communities than land-use type. FEMS Microbiol. Ecol. 2012, 79, 12–24. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.; Bhatia, A.; Yadav, S.; Kumar, V.; Singh, B. The effects of elevated CO2 and elevated O3 exposure on plant growth, yield and quality of grains of two wheat cultivars grown in North India. Heliyon 2019, 5, e02317. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.Y.; Tian, H.; Zhang, H.S.; Xiong, J.B.; Yang, H.M.; Liu, Y. Shoot-soil ecological stoichiometry of alfalfa under nitrogen and phosphorus fertilization in the Loess Plateau. Sci. Rep. 2021, 11, 15049. [Google Scholar] [CrossRef]
- Qiao, X.; Chen, X.W.; Lei, J.J.; Sai, L.H.; Xue, L.H. Apricot-based agroforestry system in Southern Xinjiang Province of China: Inflfluence on yield and quality of intercropping wheat. Agroforest Syst. 2020, 94, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.L.; Chu, X.B.; Zhu, H.Y.; Zou, D.S.; Li, L.C.; Du, L.S. The response of soil nutrients and microbial community structures in long-term tea plantations and diverse agroforestry intercropping systems. Sustainability 2021, 13, 7799. [Google Scholar] [CrossRef]
- Zhou, G.X.; Zhang, J.B.; Zhang, C.Z.; Feng, Y.Z.; Chen, L.; Yu, Z.H.; Xin, X.L.; Zhao, B.Z. Effects of changes in straw chemical properties and alkaline soils on bacterial communities engaged in straw decomposition at different temperatures. Sci. Rep. 2016, 6, 22186. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.L.; Xu, M.; Zou, X.; Chen, J.; Zhang, J.; Zhang, J. Effects of different vegetation types on soil bacterial community characteristics in mountainous and hilly areas of central Guizhou. J. Ecol. Rural Environ. 2021, 37, 518–525. [Google Scholar]
- Joel, P.D.; Angel, V.; Joachim, M.S.; Don, A.C.; Jacqueline, E.V.D.W. Differences in bacterial diversity, composition and function due to long-term agriculture in soils in the eastern free state of South Africa. Diversity 2019, 11, 61. [Google Scholar]
- Liu, H.Y.; Xu, W.; Li, J.Y.; Yu, Z.Y.; Zeng, Q.C.; Tan, W.F.; Mi, W.T. Short-term effect of manure and straw application on bacterial and fungal community compositions and abundances in an acidic paddy soil. J. Soil Sediment 2021, 21, 3057–3071. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, P.; Yoza, B.; Liu, W.; Liang, H. Phytoremediation of metal-contaminated rare-earth mining sites using Paspalum conjugatum. Chemosphere 2020, 259, 127280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Song, G.; Gelardi, D.L.; Huang, L.B.; Khan, E.; Mašek, O.; Parikh, S.J.; Sik, O.Y. Evaluating biochar and its modifications for the removal of ammonium, nitrate, and phosphate in water. Water Res. 2020, 186, 116303. [Google Scholar] [CrossRef]
- Kazerooni, E.A.; Rethinasamy, V.; Al-Sadi, A.M. Talaromyces pinophilus inhibits Pythium and Rhizoctonia-induced damping-off of cucumber. J. Plant Pathol. 2019, 101, 377–383. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, X.; Zhou, B.K.; Zhao, B.S.; Ma, M.C.; Guan, D.W.; Li, J.; Chen, S.F.; Cao, F.M.; Shen, D.L.; et al. Thirty four years of nitrogen fertilization decreases fungal diversity and alters fungal community composition in black soil in northeast China. Soil Biol. Biochem. 2016, 95, 135–143. [Google Scholar] [CrossRef]
- Bärbel, U.F.; Manfred, R.; Jörg, O. Blastocatella fastidiosa gen. nov., sp. nov., isolated from semiarid savanna soil-The first described species of Acidobacteria subdivision 4. Syst. Appl. Microbiol. 2013, 36, 82–89. [Google Scholar]
- Wang, Z.T.; Li, T.; Li, Y.Z.; Zhao, D.Q.; Han, J.; Liu, Y.; Liao, Y.C. Relationship between the microbial community and catabolic diversity in response to conservation tillage. Soil Till. Res. 2019, 196, 104431. [Google Scholar] [CrossRef]
- Shi, Y.X.; Meng, S.S.; Xie, X.W.; Chai, A.L.; Li, B.J. Dry Heat Treatment Reduces the Occurrence of Cladosporium cucumerinum, Ascochyta citrullina, and Colletotrichum orbiculare on the Surface and Interior of Cucumber Seeds. Horticult. Plant J. 2016, 2, 35–40. [Google Scholar] [CrossRef]
- Tan, Z.T.; Li, A.D.; Yang, R.; Cai, G.J.; Wu, F.C.; Guo, C.Y. Effects on community structure of soil fungi in Passiflora edulis continuous cropping. Mol. Plant Breed 2022, 20, 1373–1382. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Chemistry | LF | LE | LD |
|---|---|---|---|
| pH Value | 7.91 ± 0.03 b | 8.02 ± 0.03 a | 7.96 ± 0.01 ab |
| TN (g kg−1) | 0.47 ± 0.01 b | 0.49 ± 0.01 b | 0.54 ± 0.02 a |
| SOC (g kg−1) | 4.54 ± 0.18 c | 5.34 ± 0.15 b | 6.79 ± 0.30 a |
| TP (g kg−1) | 0.44 ± 0.01 c | 0.58 ± 0.01 b | 0.65 ± 0.02 a |
| C:N | 9.70 ± 0.19 c | 10.90 ± 0.11 b | 12.56 ± 0.18 a |
| N:P | 1.07 ± 0.05 a | 0.84 ± 0.05 b | 0.85 ± 0.02 b |
| C:P | 10.42 ± 0.62 a | 9.23 ± 0.22 a | 10.59 ± 0.78 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Wei, Y.; Yin, Y.; Ding, H.; Zhu, W.; Zhou, Y. The Effect of Intercropping Mulberry (Morus alba L.) with Peanut (Arachis hypogaea L.), on the Soil Rhizosphere Microbial Community. Forests 2022, 13, 1757. https://doi.org/10.3390/f13111757
Li M, Wei Y, Yin Y, Ding H, Zhu W, Zhou Y. The Effect of Intercropping Mulberry (Morus alba L.) with Peanut (Arachis hypogaea L.), on the Soil Rhizosphere Microbial Community. Forests. 2022; 13(11):1757. https://doi.org/10.3390/f13111757
Chicago/Turabian StyleLi, Muzi, Yawei Wei, You Yin, Hui Ding, Wenxu Zhu, and Yongbin Zhou. 2022. "The Effect of Intercropping Mulberry (Morus alba L.) with Peanut (Arachis hypogaea L.), on the Soil Rhizosphere Microbial Community" Forests 13, no. 11: 1757. https://doi.org/10.3390/f13111757