
Promoting Effect of Choline-Phosphate Cytidylyltransferase Gene (pcyt-1) on Departure of Pinewood Nematode from Monochamus alternatus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. PWN Collection and RNA Extraction
2.2. Transcriptome Sequencing (Biomarker Technologies, Beijing, China)
2.2.1. RNA Quantification and Qualification
2.2.2. Clustering and Sequencing
2.2.3. Quality Control
2.2.4. Comparative Analysis and Gene Functional Annotation
2.2.5. Differential Expression Analysis
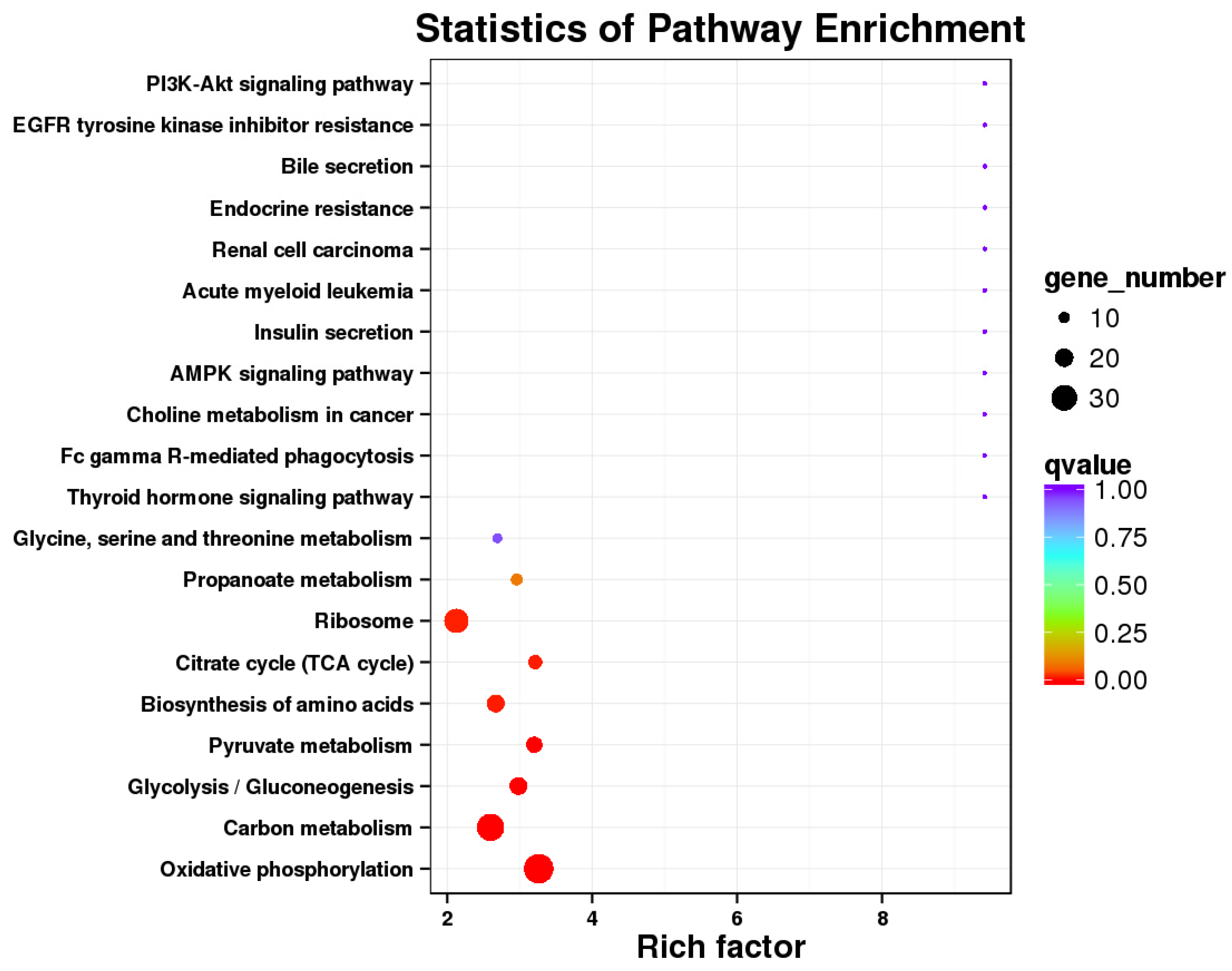
2.2.6. Pathway Enrichment Analysis
2.3. RNA Interference (RNAi) and Quantitative Real-Time PCR (qPCR)
2.4. Comparison of Difference in Motility and Lipid Content of PWN after pcyt-1 RNAi
3. Results
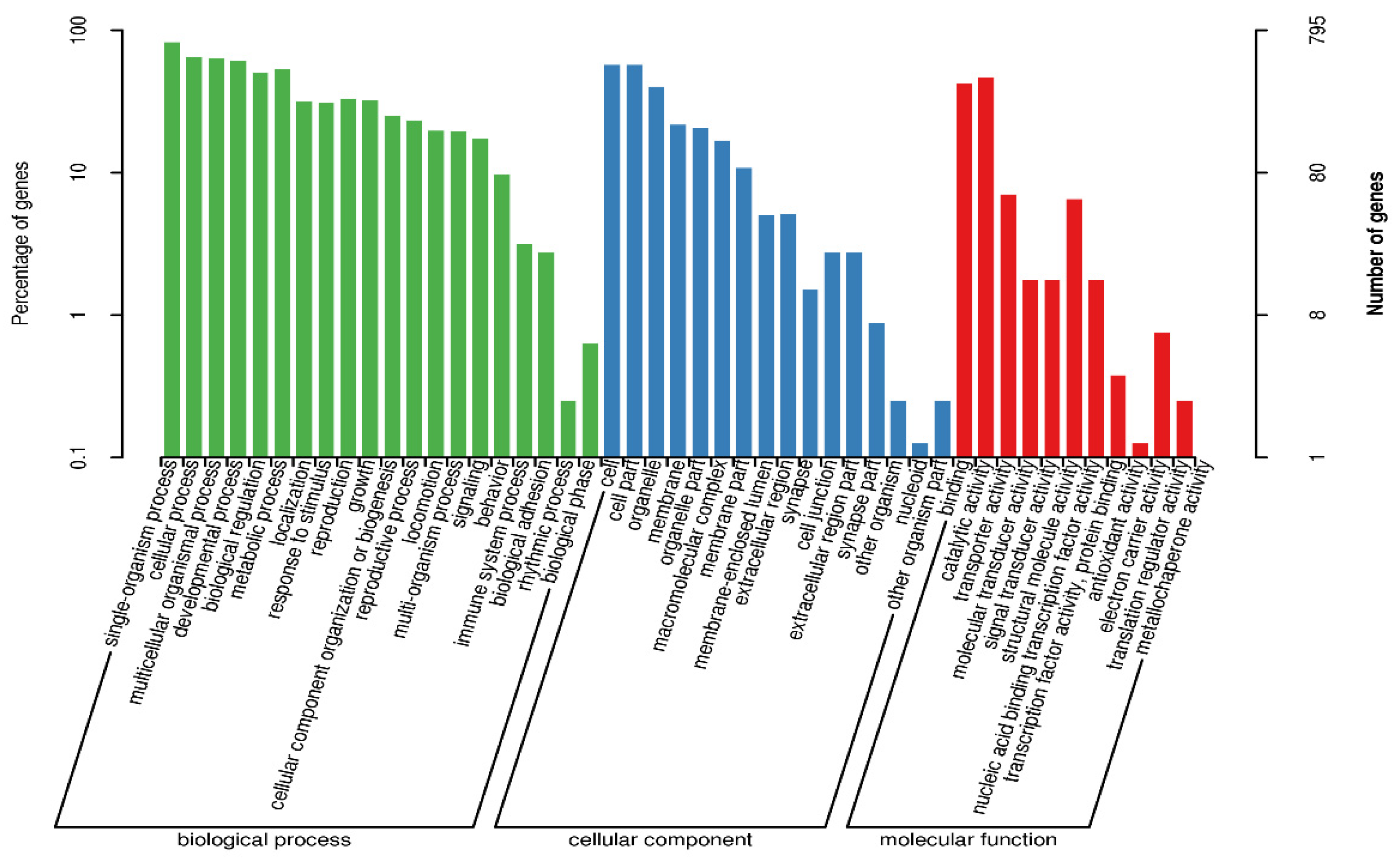
3.1. Comparison of Transcriptome Results
3.2. qPCR Gene Experession Studies
3.3. Motility and Lipid Content of PWN
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yi, C.K.; Byun, B.H.; Park, J.D.; Yang, S.I.; Chang, K.H. First finding of the pine wood nematode, Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle and its insect vector in Korea. Res. Rep. For. Res. Inst. 1989, 38, 141–149. [Google Scholar]
- Cheng, H.; Lin, M.; Li, W.; Fang, Z. The occurrence of a pine wilting disease caused by a nematode found in Nanjing. For. Pest Dis. 1983, 4, 1–5. [Google Scholar]
- Fonseca, L.; Cardoso, J.; Lopes, A.; Pestana, M.; Abreu, F.; Nunes, N.; Mota, M.; Abrantes, I. The pine wood nematode, Bursaphelenchus xylophilus, in Madeira Island. Helminthologia 2012, 49, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Robertson, L.; Arcos, S.C.; Escuer, M.; Merino, R.S.; Esparrago, G.; Abelleira, A.; Navas, A. Incidence of the pine wood nematode Bursaphelenchus xylophlius Steiner & Buhrer, 1934 (Nickle, 1970) in Spain. Nematology 2011, 13, 755–757. [Google Scholar]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus Xylophilus, Causal Agent of Pine Wilt Disease on Pinus pinaster in Northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef] [PubMed]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Bergdahl, D.R. Impact of pinewood nematode in north america: Present and future. J. Nematol. 1988, 20, 260–265. [Google Scholar]
- Morimoto, K.; Iwasaki, A. Role of Monochamus alternatus (Coleoptera: Cerambycidae) as a vector of Bursaphelenchus lignicolus (Nematoda: Aphelenchoididae). J. Jpn. For. Soc. 1972, 54, 177–183. [Google Scholar]
- Linit, M.J. Temporal Pattern of Pinewood Nematode Exit from the Insect VectorMonochamus carolinensis. J. Nematol. 1989, 21, 105–107. [Google Scholar] [PubMed]
- Yoshimura, A.; Kawasaki, K.; Takasu, F.; Togashi, K.; Futai, K.; Shigesada, N. Modeling the spread of pine wilt disease caused by nematodes with pine sawyer as vector. Ecology 1999, 80, 1691–1702. [Google Scholar] [CrossRef]
- Togashi, K.; Shigesada, N. Spread of the pinewood nematode vectored by the Japanese pine sawyer: Modeling and analytical approaches. Popul. Ecol. 2006, 48, 271–283. [Google Scholar] [CrossRef]
- Wingfield, M.J. Transmission of pine wood nematode to cut timber and girdled trees. Plant Dis. 1983, 67, 35–37. [Google Scholar] [CrossRef]
- Mamiya, Y. The pine wood nematode. Helminthologia 1984, 49, 96–103. [Google Scholar]
- Linit, M.J. Nematode-vector relationships in the pine wilt disease system. J. Nematol. 1988, 20, 227–235. [Google Scholar]
- Zhao, L.; Mota, M.; Vieira, P.; Rebecca, A.B.; Sun, J. Interspecific communication between pinewood nematode, its insect vector, and associated microbes. Trends Parasitol. 2014, 30, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Linit, M.J. Transmission of pinewood nematode through feeding wounds of Monochamus carolinensis (Coleoptera: Cerambycidae). J. Nematol. 1990, 22, 231–236. [Google Scholar] [PubMed]
- Necibi, S.; Linit, M.J. Effect of Monochamus carolinensis on Bursaphelenchus xylophilus Dispersal Stage Formation. J. Nematol. 1998, 30, 246–254. [Google Scholar] [PubMed]
- Zhao, L.L.; Wei, W.; Kang, L.; Sun, J.H. Chemotaxis of the pinewood nematode, Bursaphelenchus xylophilus, to volatiles associated with host pine, Pinus massoniana, and its vector Monochamus alternatus. J. Chem. Ecol. 2007, 33, 1207–1216. [Google Scholar] [CrossRef]
- Stamps, W.T.; Linit, M.J. Interaction of intrinsic and extrinsic chemical cues in the behaviour of Bursaphelenchus xylophilus (Aphelenchida: Aphelenchoididae) in relation to its beetle vectors. Nematology 2001, 3, 295–301. [Google Scholar]
- Niu, H.; Zhao, L.; Lu, M.; Zhang, S.; Sun, J. The ratio and concentration of two monoterpenes mediate fecundity of the pine wood nematode and growth of its associated fungi. PLoS ONE 2012, 7, e31716. [Google Scholar] [CrossRef]
- Stamps, W.T.; Linit, M.J. Neutral Storage Lipid and Exit Behavior of Bursaphelenchus xylophilus Fourth- stage dispersal juveniles from their beetle vectors. J. Nematol. 1988, 30, 255–261. [Google Scholar]
- Tominaga, Y.; Yamamoto, M.; Kuwahara, Y.; Sugawara, R. Behavioral responses of the pine wood nematode to terpenes. J. Agric. Chem. Soc. Jpn. 1984, 48, 519–520. [Google Scholar] [CrossRef]
- Watanabe, H.; Ishikawa, M.; Shuto, Y. Attractive activity of β-myrcene in pine species to pine wood nematode Bursaphelenchus lignicolus. Kagaku Seibutsu 1982, 20, 123. [Google Scholar]
- Ishkawa, M.; Shuto, Y.; Watanabe, H. β-Myrcene, a potent attractant component of pine wood for the pine wood nematode, Bursaphelenchus xylophilus. J. Agric. Chem. Soc. Jpn. 1986, 50, 1863–1866. [Google Scholar] [CrossRef] [Green Version]
- Enda, N.; Ikeda, T. Role of volatiles of a pine tree as emerging stimulants for attracting the pine wood nematode from the pine sawyer. Trans. 94th Ann. Meet. Jpn. For. Soc. 1983, 94, 479–480. [Google Scholar]
- Stamps, W.T.; Linit, M.J. Chemotactic response of propagative and dispersal forms of the pine wood nematode Bursaphelenchus xylophilus to beetle and pine derived compounds. Fundam. Appl. Nematol. 1998, 21, 243–250. [Google Scholar]
- Aikawa, T.; Togashi, K. An effect of pine volatiles on departure of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) from Monochamus alternatus (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1998, 33, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, F.; Wang, L.; Zhou, L.; Song, J. Study of the departure of pine wood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) from Monochamus alternatus (Coleoptera: Cerambycidae). J. Asia-Pac. Entomol. 2020, 23, 981–987. [Google Scholar] [CrossRef]
- Futai, K. Pine Wood Nematode, Bursaphalenchus xylopholus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aikawa, T.; Togashi, K. Movement of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) in tracheal system of adult Monochamus alternatus (Coleoptera: Cerambycidae). Nematology 2000, 2, 495–500. [Google Scholar] [CrossRef]
- Jingyan, W. Biochemistry, 3rd ed.; Higher Education Press: Beijing, China, 2002; pp. 230–231. [Google Scholar]
- Viglierchio, D.R.; Schmitt, R.V. On the methodology of nematode extraction from field samples: Baermann funnel modifications. J. Nematol. 1983, 15, 438–444. [Google Scholar] [PubMed]
- Han-song, Y.; Shuai, P.; Yuan-hong, X.; Yao-hui, H. Study on improvement of RNA isolating reagent Kit Trizol. Food Sci. Technol. 2005, 26, 39–42. [Google Scholar]
- Mao, X.; Tao, C.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Davanloo, P.; Rosenberg, A.H.; Dunn, J.J.; Studier, F.W. Cloning and expression of the gene for bacteriophage T7 RNA polymerase. Proc. Natl. Acad. Sci. USA 1984, 81, 2035–2039. [Google Scholar] [CrossRef] [Green Version]
- Ai-ling, B.; Zheng-min, H.; Xu, H.; Ping, S. A study of the acquisition of sterilized nematode and growth of the pine wood nematode. J. Nanjing For. Univ. 2008, 32, 99–102. [Google Scholar]
- Yun-tao, J.; Yi-shu, W.; Xing-chen, L.; Chang-qing, Q. Visualization of fat droplets of Caenorhabditis elegans. Chin. J. Biochem. Pharm. 2011, 32, 141–142. [Google Scholar]
- Wang, Y.; Chen, J.; Chen, F.; Zhou, Q.; Zhou, L.; Sun, S. Transmission of the pine wood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), through feeding activity of Monochamus alternatus (Coleoptera: Cerambycidae). J. Nanjing For. Univ. 2019, 43, 1–10. [Google Scholar]
- HosodaR, K.K. Drop off procedures of the pine wood nematode from the pine sawyer. Trans. Ann. Meet. Jpn. For. Soc. 1977, 28, 255–258. [Google Scholar]
- Aikawa, T. Transmission Biology of Bursaphelenchus xylophilus in relation to its insect vector. In Pine Wilt Disease; Springer: Tokyo, Japan, 2008; pp. 123–138. [Google Scholar]
- Jikumaru, S.; Togashi, K. Temperature effects on the transmission of Bursaphelenchus xylophilus (Nemata: Aphelenchoididae) by Monochamus alternatus (Coleoptera: Cerambycidae). J. Nematol. 2000, 32, 110–116. [Google Scholar]
- Togashi, K. Transmission curves of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) from its vector, Monochamus alternatus (Coleoptera: Cerambycidae), to pine trees with reference to population performance. Appl. Entomol. Zool. 1985, 20, 246–251. [Google Scholar] [CrossRef] [Green Version]
- Hasengschwandtner, F. Phosphatidylcholine treatment to induce lipolysis. J. Cosmet. Dermtol. 2006, 4, 308–313. [Google Scholar] [CrossRef]
- Salles, A.G.; Valler, C.S.; Ferreira, M.C. Histologic response to injected phosphatidylcholine in fat tissue: Experimental study in a New Rabbit Model. Aesthet. Plast. Surg. 2006, 30, 479–484. [Google Scholar] [CrossRef]
- Young, V.L. Lipostabil the effect of phosphatidylcholine on subcutaneous fat. Aesthet. Surg. J. 2003, 23, 413–417. [Google Scholar] [CrossRef] [Green Version]
- Morton, C.C.; Aitchison, A.J.; Gehrig, K.; Ridgway, N.D. A mechanism for suppression of the CDP-choline pathway during apoptosis. J. Lipid Res. 2013, 54, 3373–3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, E.P. The metabolism and function of complex lipids. Harvey Lect. 1962, 57, 143–171. [Google Scholar]
- Choy, P.C.; Lim, P.H.; Vance, D.E. Purification and characterization of CTP: Cholinephosphate cytidylytransferase from rat liver cytosol. J. Biol. Chem. 1977, 252, 7673–7677. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Jin, M.; Wang, L.; Yu, A.; Xie, G.; Chen, F. Promoting Effect of Choline-Phosphate Cytidylyltransferase Gene (pcyt-1) on Departure of Pinewood Nematode from Monochamus alternatus. Forests 2022, 13, 114. https://doi.org/10.3390/f13010114
Wang Y, Jin M, Wang L, Yu A, Xie G, Chen F. Promoting Effect of Choline-Phosphate Cytidylyltransferase Gene (pcyt-1) on Departure of Pinewood Nematode from Monochamus alternatus. Forests. 2022; 13(1):114. https://doi.org/10.3390/f13010114
Chicago/Turabian StyleWang, Yang, Mingxia Jin, Lichao Wang, Ailin Yu, Guai Xie, and Fengmao Chen. 2022. "Promoting Effect of Choline-Phosphate Cytidylyltransferase Gene (pcyt-1) on Departure of Pinewood Nematode from Monochamus alternatus" Forests 13, no. 1: 114. https://doi.org/10.3390/f13010114