
Attempt to Identify Sex Hormones in the Bodies of Selected Norway Spruce Bark Beetles


Abstract
:1. Introduction
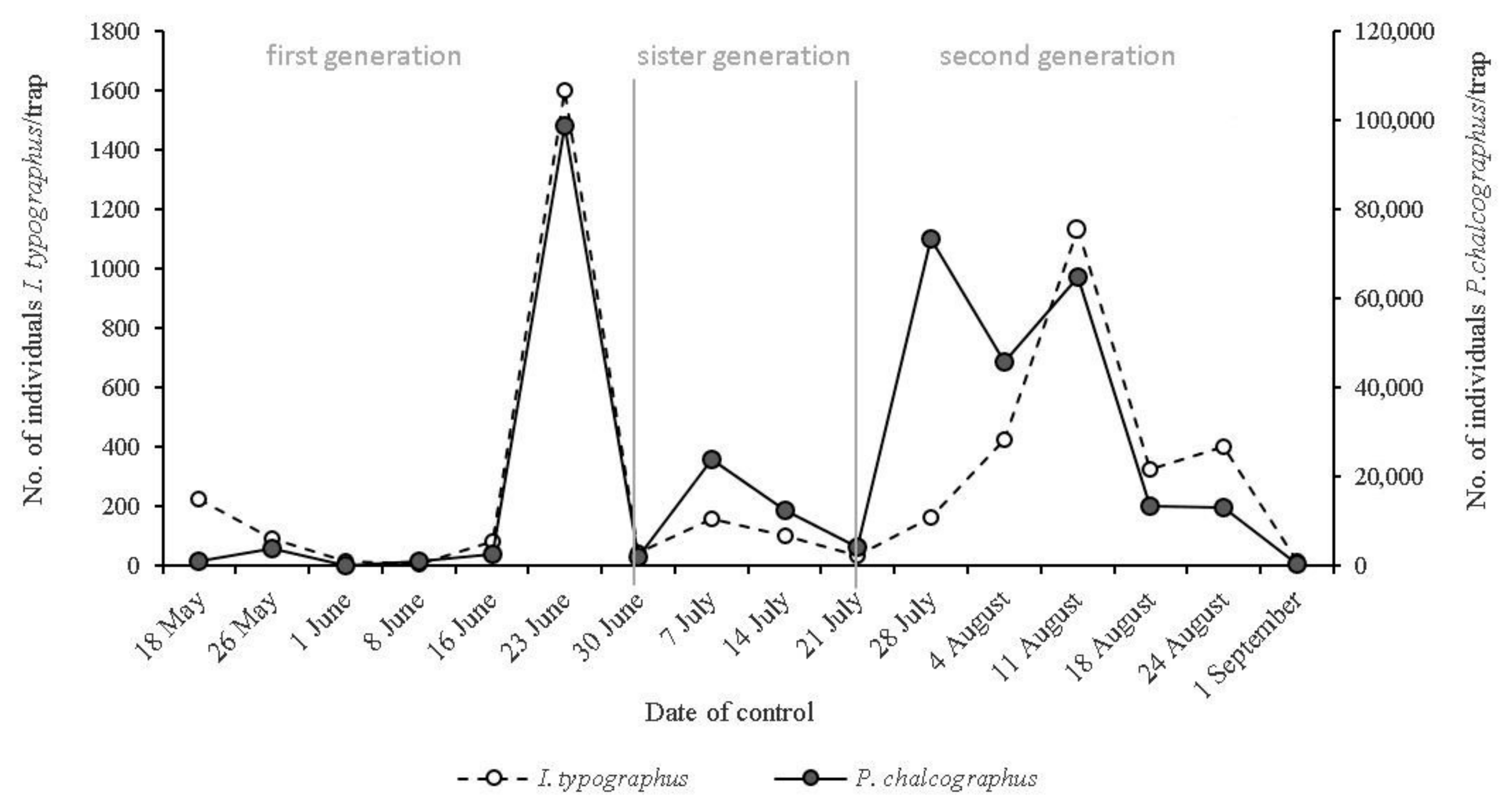
2. Materials and Methods
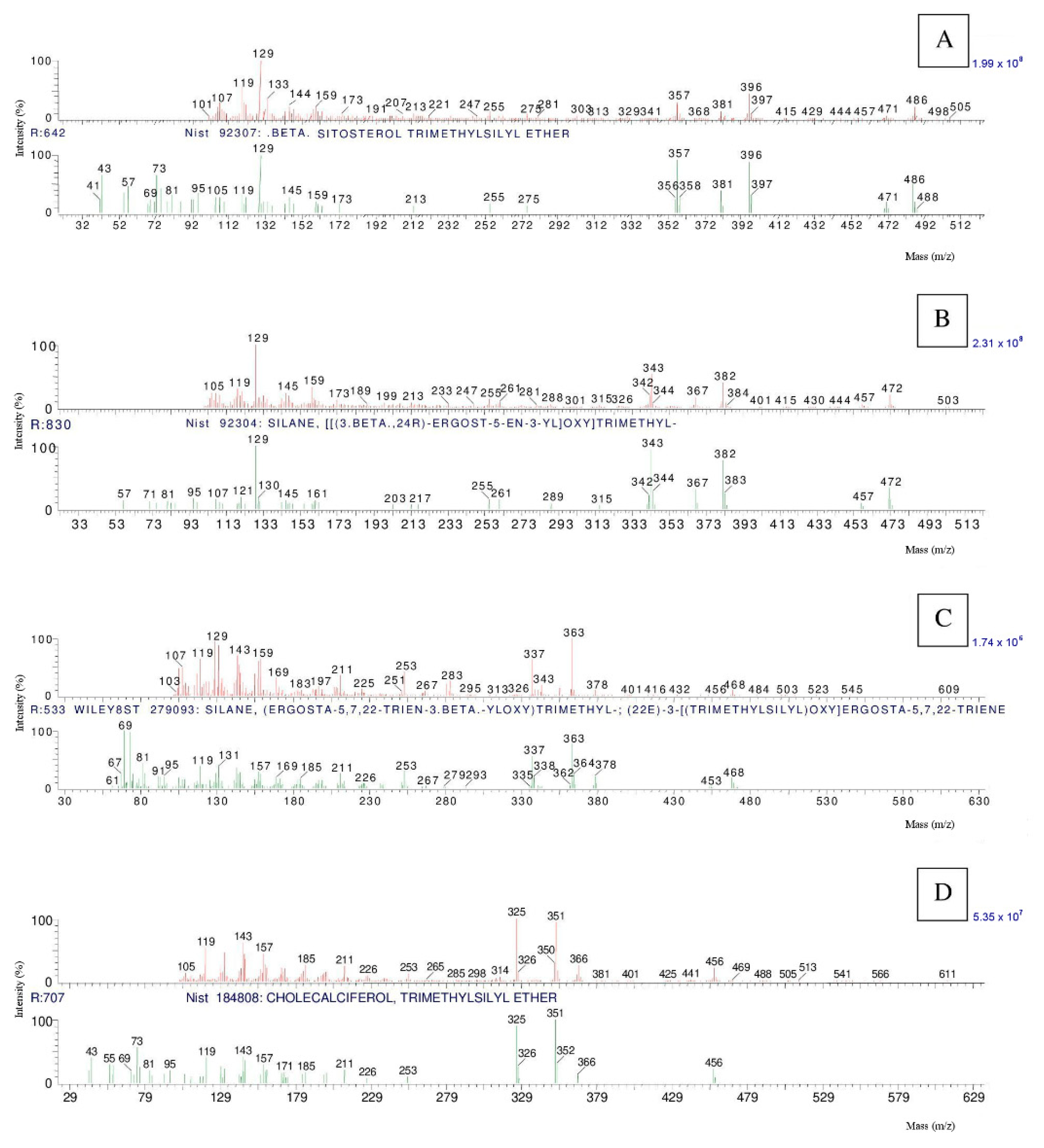
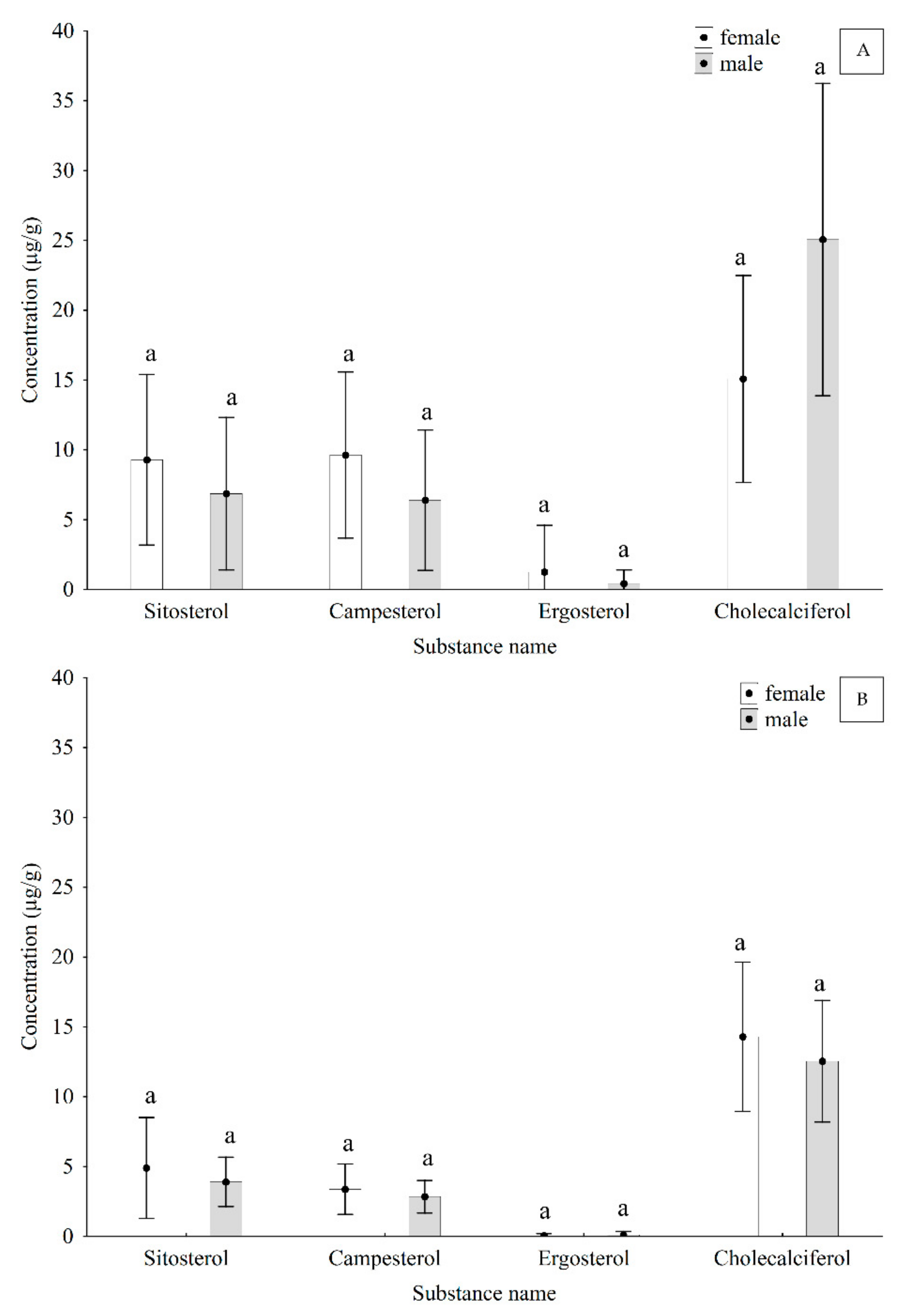
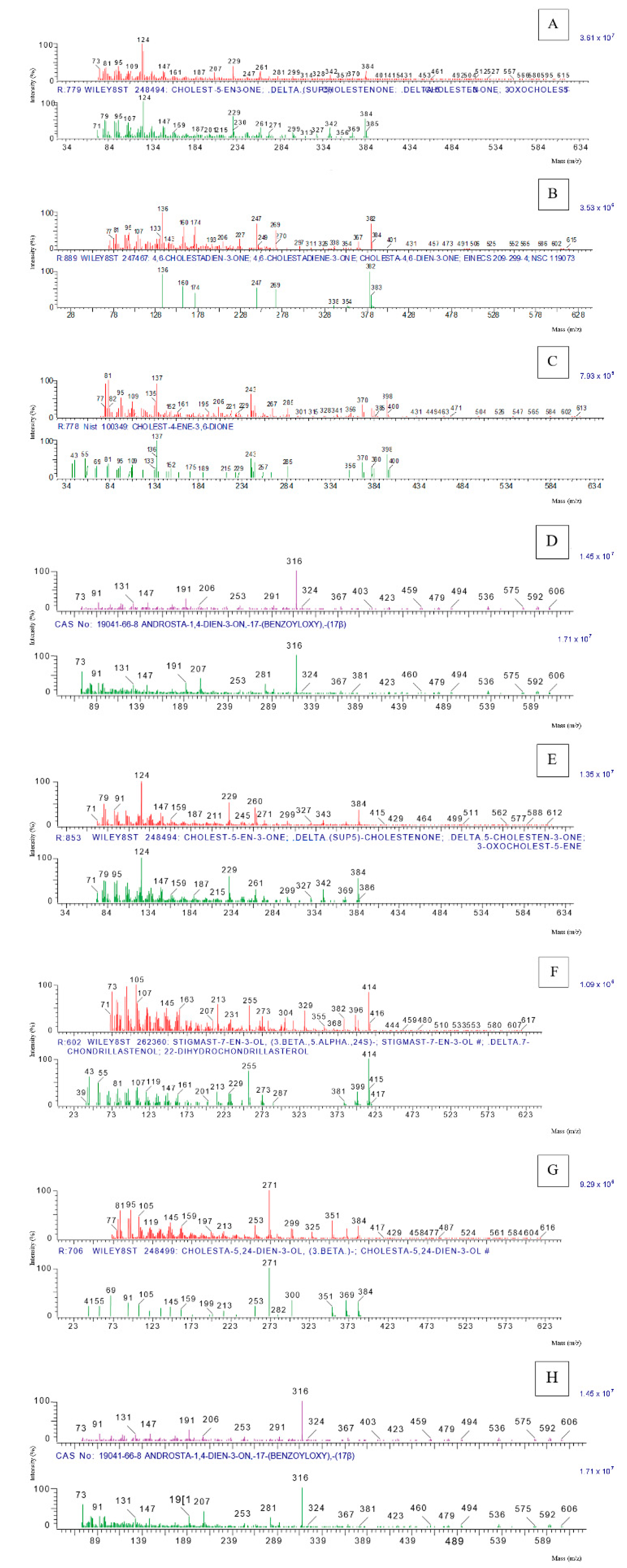
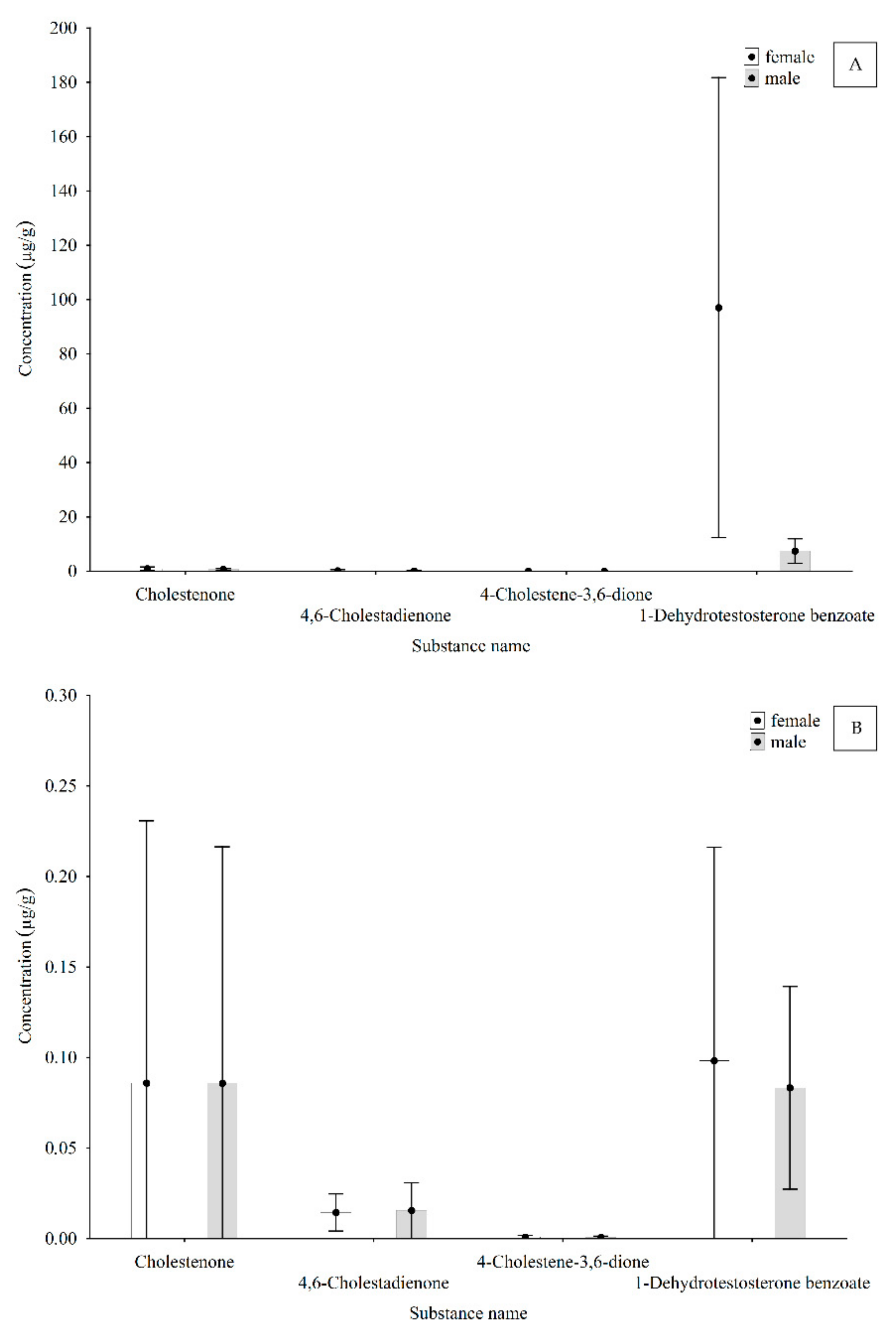
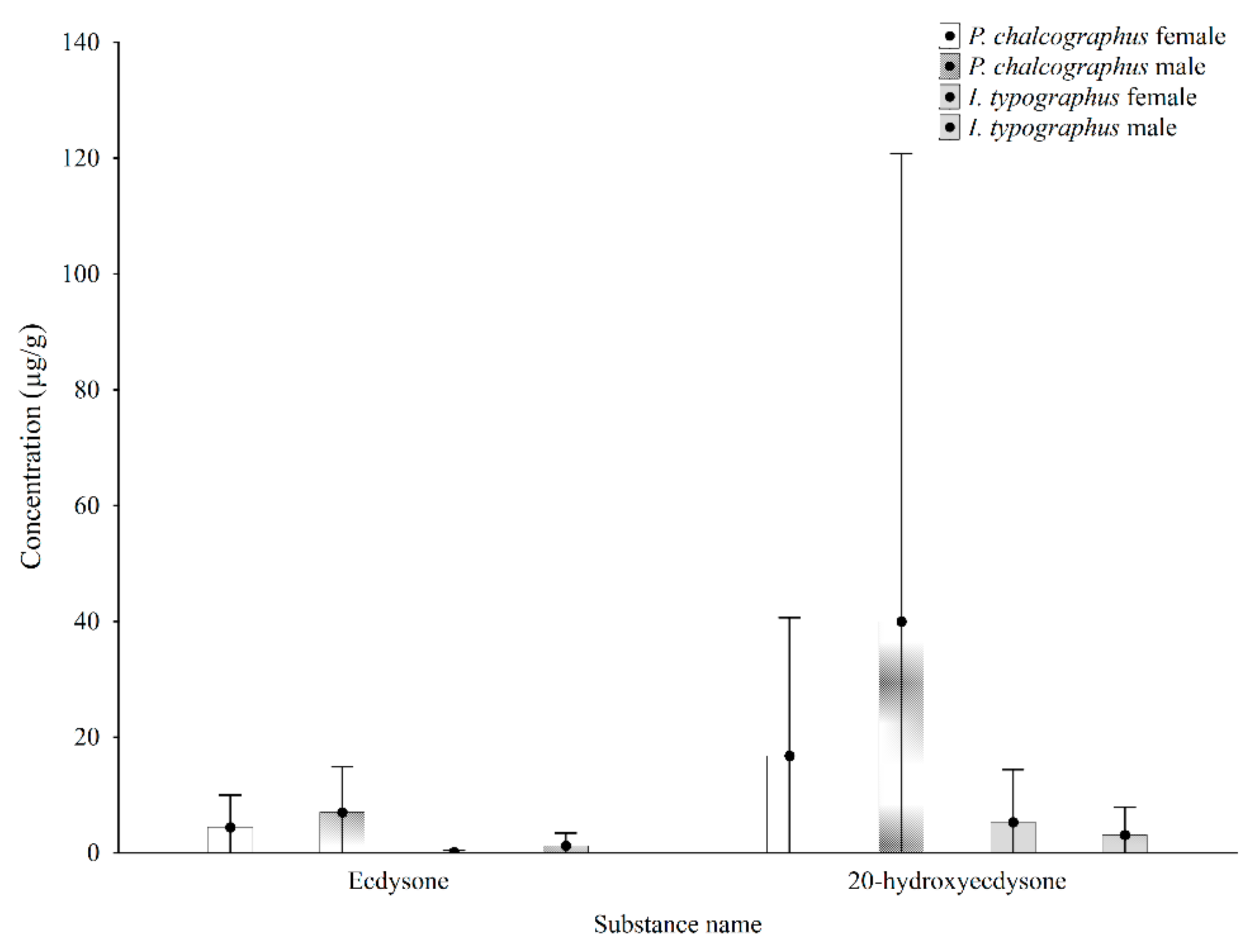
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interests
References
- De Loof, A.; Huybrechts, R. Insects Do Not Have Sex Hormones: A Myth? Gen. Comp. Endocrinol. 1998, 111, 245–260. [Google Scholar] [CrossRef]
- Robinett, C.C.; Vaughan, A.G.; Knapp, J.-M.; Baker, B.S. Sex and the Single Cell. II. There Is a Time and Place for Sex. PLoS Biol. 2010, 8, e1000365. [Google Scholar] [CrossRef]
- Mechoulam, R.; Brueggemeier, R.W.; Denlinger, D.L. Estrogens in insects. Cell. Mol. Life Sci. 1984, 40, 942–944. [Google Scholar] [CrossRef]
- De Loof, A. Ecdysteroids: The overlooked sex steroids of insects? Males: The black box. Insect. Sci. 2006, 13, 325–338. [Google Scholar] [CrossRef]
- Jury, C. Biologia Rozwoju Owadów; PWN: Warszawa, Poland, 1988; p. 312. [Google Scholar]
- Bear, A.; Monteiro, A. Both cell-autonomous mechanisms and hormones contribute to sexual development in vertebrates and insects. BioEssays 2013, 35, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Birgersson, G.; Bergström, G. Volatiles released from individual spruce bark beetle entrance holes quantitative variations during the first week of attack. J. Chem. Ecol. 1989, 15, 2465–2483. [Google Scholar] [CrossRef] [PubMed]
- Lanne, B.S.; Ivarsson, P.; Johnsson, P.; Bergström, G.; Wassgren, A.B. Biosynthesis of 2-methyl-3-buten-2-ol, a pheromone component of Ips typographus (Coleoptera: Scolytidae). Insect Biochem. 1989, 19, 163–167. [Google Scholar] [CrossRef]
- Zhang, Q.-H.; Ma, J.-H.; Zhao, F.-Y.; Song, L.-W.; Sun, J.-H. Aggregation Pheromone of the Qinghai Spruce Bark Beetle, Ips nitidus Eggers. J. Chem. Ecol. 2009, 35, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Baader, E.J. Pityogenes spp. (Col., Scolytidae): Investigations on semiochemicals and their application in forest protection. J. Appl. Entomol. 1989, 107, 1–31. [Google Scholar] [CrossRef]
- Juráň, S.; Pallozzi, E.; Guidolotti, G.; Fares, S.; Šigut, L.; Calfapietra, C.; Alivernini, A.; Savi, F.; Večeřová, K.; Křůmal, K.; et al. Fluxes of biogenic volatile organic compounds above temperate Norway spruce forest of the Czech Republic. Agric. For. Meteorol. 2017, 232, 500–513. [Google Scholar] [CrossRef]
- Engelmann, F. Vitellogenesis controlled by juvenile hormone. In Endocrinology of Insects; Downer, R.G.H., Laufer, H., Eds.; Alan, R. Liss: New York, NY, USA, 1983; pp. 259–270. [Google Scholar]
- Hagedorn, H.H. The role of ecdysteroids in reproduction. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Kerkut, G., Gilbert, L., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 205–261. [Google Scholar]
- Hagedorn, H.H.; O’Connor, J.D.; Fuchs, M.S.; Sage, B.; Schlaeger, D.A.; Bohm, M.K. The ovary as a source of alpha-ecdysone in an adult mosquito. Proc. Natl. Acad. Sci. USA 1975, 72, 3255–3259. [Google Scholar] [CrossRef]
- Bouget, C.; Duelli, P. The effects of windthrow on forest insect communities: A literature review. Biol. Conserv. 2004, 118, 281–299. [Google Scholar] [CrossRef]
- Stadelmann, G.; Bugmann, H.; Wermelinger, B.; Bigler, C. Spatial interactions between storm damage and subsequent infes-tations by the European spruce bark beetle. For. Ecol. Manag. 2014, 318, 167–174. [Google Scholar] [CrossRef]
- Netherer, S.; Matthews, B.; Katzensteiner, K.; Blackwell, E.; Henschke, P.; Hietz, P. Do water-limiting conditions predispose Norway spruce to bark beetle attack? N. Phytol. 2015, 205, 1128–1141. [Google Scholar] [CrossRef]
- Marini, L.; Økland, B.; Jönsson, A.M.; Bentz, B.J.; Carroll, A.; Forster, B.; Grégoire, J.-C.; Hurling, R.; Nageleisen, L.M.; Netherer, S.; et al. Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography 2017, 40, 1426–1435. [Google Scholar] [CrossRef]
- De Groot, M.; Ogris, N.; Kobler, A. The effects of a large-scale ice storm event on the drivers of bark beetle outbreaks and associated management practices. For. Ecol. Manag. 2018, 408, 195–201. [Google Scholar] [CrossRef]
- Hlásny, T.; Krokene, P.; Liebhold, A.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.; Schelhaas, M.-J.; Seidl, R.; Svoboda, M.; et al. Living with bark beetles: Impacts, outlook and management options. From Sci. Policy 2019. [Google Scholar] [CrossRef]
- Jactel, H.; Koricheva, J.; Castagneyrol, B. Responses of forest insect pests to climate change: Not so simple. Curr. Opin. Insect Sci. 2019, 35, 103–108. [Google Scholar] [CrossRef]
- Schlyter, F.; Byers, J.A.; Löfquist, J. Attraction to pheromone sources of different quantity, quality and spacing: Densi-ty-regulation mechanisms in bark beetle Ips typographus. J. Chem. Ecol. 1987, 13, 1503–1523. [Google Scholar] [CrossRef]
- Zumr, V. Reduction of mass outbreaks in spruce bark beetle Ips typographus L. Coleoptera Scolytidae by pheromone traps. Lesnictvi 1987, 33, 49–64. [Google Scholar]
- Dimitri, L.; Gebauer, U.; Lösekrug, R.; Vaupel, O. Influence of mass trapping on the population dynamic and damage-effect of bark beetles. J. Appl. Èntomol. 1992, 114, 103–109. [Google Scholar] [CrossRef]
- Faccoli, M.; Stergulc, F. Damage reduction and performance of mass trapping devices for forest protection against the spruce bark beetle,Ips typographus (Coleoptera Curculionidae Scolytinae). Ann. For. Sci. 2008, 65, 309. [Google Scholar] [CrossRef]
- Bakke, A. Using Pheromones in the Management of Bark Beetle Outbreaks. Available online: http://www.nrs.fs.fed.us/pubs/gtr/gtr_ne153/gtr_ne153_371.pdf (accessed on 21 March 2020).
- Weslien, J.; Bylund, H. The number and sex of spruce bark beetles, Ips typographus (L.), caught in pheromone traps as related to flight season, trap type, and pheromone release. J. Appl. Èntomol. 1988, 106, 488–493. [Google Scholar] [CrossRef]
- Vité, J.P. The European struggle to control Ips typographus-past, present and future. Ecography 1989, 12, 520–525. [Google Scholar] [CrossRef]
- Faccoli, M. A morphological illustrated key to European species of the genus Ips DeGeer (Coleoptera: Scolytidae). The Coleopt. 2004, 13, 103–119. [Google Scholar]
- Byers, J.A. Chemical ecology of bark beetles. Cell. Mol. Life Sci. 1989, 45, 271–283. [Google Scholar] [CrossRef]
- Grégoire, J.C.; Evans, H.F. Damage and control of BAWBILT organisms-an overview. In Bark and Wood Boring Insects in Living Trees in Europe, A Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J.C., Evans, H.F., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 19–37. [Google Scholar]
- Schlyter, F.; Cederholm, I. Separation of the sexes of living spruce bark beetles, Ips typographus (L.), (Coleoptera: Scolytidae)1. Z. Angew. Entomol. 2009, 92, 42–47. [Google Scholar] [CrossRef]
- Pfeffer, A. Kůrovcovití (Scolytidae) a Jádrohlodovití (Platypodidae); Academia: Prague, Czech Republic, 1989; p. 137. [Google Scholar]
- Wermelinger, B. Ecology and management of the spruce bark beetle Ips typographus—A review of recent research. For. Ecol. Manag. 2004, 202, 67–82. [Google Scholar] [CrossRef]
- Bednarz, B.; Kacprzyk, M. An Innovative Method for Sex Determination of the European Spruce Bark Beetle Ips typographus (Coleoptera: Scolytinae). Èntomol. Gen. 2012, 34, 111–118. [Google Scholar] [CrossRef]
- Marciniak, P.; Szymczak, M.; Rosiński, G. Hormony peptydowe owadów-przegląd najważniejszych rodzin. Postępy Biologii Komórki 2011, 38, 43–63. [Google Scholar]
- Sanchez, L. Sex-determining mechanisms in insects. Int. J. Dev. Biol. 2008, 52, 837–856. [Google Scholar] [CrossRef] [PubMed]
- Gempe, T.; Beye, M. Function and evolution of sex determination mechanisms, genes and pathways in insects. BioEssays 2010, 33, 52–60. [Google Scholar] [CrossRef]
- Salz, H.K. Sex determination in insects: A binary decision based on alternative splicing. Curr. Opin. Genet. Dev. 2011, 21, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Negri, I.; Pellecchia, M. Sex Steroids in Insects and the Role of the Endosymbiont Wolbachia: A New Perspective. In Sex Hormones; Dubey, R.K., Ed.; InTech: London, UK, 2012; pp. 353–374. [Google Scholar]
- Raikhel, A.S.; Brown, M.; Belles, X. Hormonal control of reproductive processes. In Comprehensive Insect Physiology, Biochemistry, Pharmacology and Molecular Biology; Red, L., Gilbert, L., Gill, S., Iatrou, K., Eds.; Elsevier Press: Amsterdam, The Netherlands, 2005; pp. 433–491. [Google Scholar]
- Wu, S.J.; Zhang, J.Z.; Ma, M. Monitoring the effects of juvenile hormones and 20 hydroxyecdysone on yolk polypeptide production of Drosophila melanogaster with enzyme immunoassay. Physiol. Entomol. 1987, 12, 355–361. [Google Scholar] [CrossRef]
- Hagedorn, H.H. Physiological roles of hemolymph ecdysteroid in the adult insect. In Ecdysone from Chemistry to Mode of Action; Koolman, J., Ed.; Thieme Medical Publishers Inc.: New York, NY, USA, 1989; pp. 279–289. [Google Scholar]
- Bradfield, J.Y.; Berlin, R.L.; Keeley, L.L. Contrasting modulations of gene expression by a juvenile hormone analog. Insect Biochem. 1990, 20, 105–111. [Google Scholar] [CrossRef]
- Yin, C.-M.; Zou, B.-X.; Yi, S.-X.; Stoffolano, J.G., Jr. Ecdysteroid activity during ögenesis in the black blowfly, Phormia regina (Meigen). J. Insect. Physiol. 1990, 36, 375–382. [Google Scholar] [CrossRef]
- Ito, H.; Sato, K.; Yamamoto, D. Sex-switching of the Drosophila brain by two antagonistic chromatin factors. Fly 2013, 7, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Bonhag, P.F. Ovarian Structure and Vitellogenesis in Insects. Annu. Rev. Èntomol. 1958, 3, 137–160. [Google Scholar] [CrossRef]
- Zumr, V.; Soldán, T. Reproductive cycles of Ips typographus, I. amitinus and Pityogenes chalcographus (Coleptera, Scolytidae). ACTA Entomol. Bohemos. 1981, 78, 280–289. [Google Scholar]
- Kacprzyk, M.; Bednarz, B. The possibilities of six-toothed bark beetle (Pityogenes chalcographus L.) (Coleoptera: Scolytinae) gender determination based on imagines biometric characteristic. J. Entomol. Res. Soc. 2014, 17, 71–82. [Google Scholar]
- Jarošík, V.; Honěk, A.; Fairbairn, D.J.; Blanckenhorn, W.U.; Székely, T. Sexual Differences in Insect Development Time in Relation to Sexual Size Dimorphism; Oxford University Press: Oxford, UK, 2007; pp. 205–212. [Google Scholar]
- Simonet, G.; Poels, J.; Claeys, I.; Van Loy, T.; Franssens, V.; De Loof, A.; Broeck, J.V. Neuroendocrinological and Molecular Aspects of Insect Reproduction. J. Neuroendocr. 2004, 16, 649–659. [Google Scholar] [CrossRef]
- De Loof, A. Ecdysteroids, juvenile hormone and insect neuropeptides: Recent successes and remaining major challenges. Gen. Comp. Endocrinol. 2008, 155, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Vanderplanck, M.; Michez, D.; Vancraenenbroeck, S.; Lognay, G. Micro-Quantitative Method for Analysis of Sterol Levels in Honeybees and Their Pollen Loads. Anal. Lett. 2011, 44, 1807–1820. [Google Scholar] [CrossRef]
- Zhou, J.; Qi, Y.; Hou, Y.; Zhao, J.; Li, Y.; Xue, X.; Wu, L.; Zhang, J.; Chen, F. Quantitative determination of juvenile hormone III and 20-hydroxyecdysone in queen larvae and drone pupae of Apis mellifera by ultrasonic-assisted extraction and liquid chromatography with electrospray ionization tandem mass spectrometry. J. Chromatogr. B 2011, 879, 2533–2541. [Google Scholar] [CrossRef]
- Axelsson, B.-O.; Saraf, A.; Larsson, L. Determination of ergosterol in organic dust by gas chromatography-mass spectrometry. J. Chromatogr. B Biomed. Sci. Appl. 1995, 666, 77–84. [Google Scholar] [CrossRef]
- Parish, E.J.; Nes, W.D. Biochemistry and Function of Sterols; Apple Academic Press: Palm Bay, FL, USA, 2020; p. 288. [Google Scholar]
- Zhang, B.; Sato, K.; Yamamoto, D. Ecdysone signaling regulates specification of neurons with a male-specific neurite in Drosophila. Biol. Open 2018, 7, bio029744. [Google Scholar] [CrossRef] [PubMed]
- Whisenton, L.R.; Warren, J.T.; Manning, M.K.; Bollenbacher, W.E. Ecdysteroid titers during pupal -adult development of Aedes aegypti: Basis for a sexual dimorphism in the rate of development. J. Insect. Physiol. 1989, 35, 67–73. [Google Scholar] [CrossRef]
- Truman, J.W. Hormonal Control of Insect Ecdysis: Endocrine Cascades for Coordinating Behavior with Physiology. Vitam. E 2005, 73, 1–30. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | β sitosterol | (3. β., 24R) ergost-5-en-3-yl | ergosta-5.7.22-trien-3 β -yloxy | Cholecalciferol | ||||
|---|---|---|---|---|---|---|---|---|
| F | p-Value | F | p-Value | F | p-Value | F | p-Value | |
| P. chalcographus | ||||||||
| Sex | 2.027 | 0.1849 | 3.471 | 0.0920 | 0.971 | 0.3476 | 3.973 | 0.0742 |
| Swarming period | 4.959 | 0.0318 | 4.661 | 0.0371 | 2.755 | 0.1114 | 2.018 | 0.1835 |
| Sex and swarming period | 1.886 | 0.2018 | 2.092 | 0.1742 | 0.831 | 0.4635 | 0.141 | 0.8697 |
| I. typographus | ||||||||
| Sex | 1.334 | 0.2632 | 0.791 | 0.3854 | 0.747 | 0.7006 | 0.599 | 0.4490 |
| Swarming period | 2.063 | 0.3565 | 2.400 | 0.1015 | 6.983 | 0.0026 | 7.908 | 0.0014 |
| Sex and swarming period | 2.199 | 0.1233 | 1.528 | 0.2414 | 0.479 | 0.3988 | 1.012 | 0.4104 |
| Compound | RT | Ion Molecular | Cas No. | Function * |
|---|---|---|---|---|
| vitamin E | 19.138 | 430.00 | 59-02-9 | PH |
| cholesterol | 19.850 | 386.00 | 57-88-5 | S |
| provitamin d3 | 20.573 | 384.00 | 434-16-2 | PH |
| campesterol | 21.550 | 400.00 | 474-62-4 | S |
| cholestenone | 22.165 | 384.12 | 601-54-7 | SH |
| 4,6-cholestadien-3-one | 22.977 | 382.14 | 566-93-8 | SH |
| gamma sitosterol | 18.843 | 414.00 | 83-47-6 | S |
| alpha n-hexadecylhydrindane | 23.933 | 384.96 | 55401-73-5 | PH |
| ui not sterol | 24.146 | 321.24 | PH | |
| ui sterol | 25.875 | 316.00 | S | |
| cholest-4-ene-3,6-dione | 26.105 | 398.14 | 984-84-9 | SH |
| ui rather sterol | 29.121 | 347.26 | S | |
| ui rather sterol | 29.750 | 349.26 | S | |
| methyl hexadecanoic acid | 9.479 | 74.00 | 112-39-0 | O |
| methyl-6,9-octadecadienoate | 10.898 | 294.00 | 56599-55-4 | PH |
| methyl 7,10,13-hexdecatrienate | 10.98 | 79.00 | 13058-55-4 | PH |
| methyloctadecanoic acid | 11.09 | 74.00 | 112-61-8 | O |
| unsaturated acid methyl ester | 11.481 | 81.00 | PH | |
| 2-octadecanal (synthetic pheromone) | 11.459 | 83.00 | 56554-96-2 | PH |
| ui silacyclobutane derivative | 11.805 | 72.00 | PH | |
| n-propyl-9-cis,11-trans-octadecanoate | 11.463 | 81.26 | 336-72-6 | PH |
| n-propyl-9-octadecadienoate- | 11.470 | 265.00 | 336-64-8 | PH |
| ethyl 9,12,15-octadecatrienoate | 11.467 | 79.00 | 336-77-4 | PH |
| tricosane (most likely) | 12.269 | 85.00 | 638-67-5 | O |
| methyl 10-oxooctadecanoate | 12.592 | 156.00 | 870-10-0 | PH |
| ui silacyclobutane derivative (most likely) | 13.110 | 72.00 | PH | |
| 1-docosanol | 13.243 | 83.00 | 661-19-8 | PH |
| ui alcohol | 13.559 | 83.00 | PH | |
| ui alcohol | 13.559 | 83.00 | PH | |
| ui saturated hydrocarbon | 13.656 | 71.00 | O | |
| phtalic acid 6-ethyloct-3-yl-2-ethylhexyl ester | 14.138 | 149.00 | 900315-53-8 | PH |
| 17β-hydroxyandrosta-1,4-dien-3-one benzoate | 14.411 | 122.00 | 19041-66-8 | SH |
| ui alcohol | 14.832 | 83.00 | PH | |
| methyl 11,14,17-eicosatrienoate | 14.893 | 108.00 | 90041-66-8 | PH |
| ui saturated hydrocarbon | 14.832 | 71.00 | O | |
| ui aromatic hydrocarbon | 15.044 | 119.00 | O | |
| 5-oxazolidinone, 3-benzoyl-2-(1,1-dimethylethyl)-4-phenylmethyl- | 15.590 | 105.00 | 104057-67-2 | PH |
| 2,6,10,14,18,22-tetracosahexaene, 2,6,10,15,19,23-hexamethyl-, (squalene) | 15.748 | 81.00 | 7683-64-9 | PHT |
| ui cyclic hydrocarbon | 16.040 | 82.00 | PH | |
| ui cyclic hydrocarbon | 16.032 | 82.00 | PH | |
| methyl 2-hydroxy-6-hexadeca-8,11,14-trienyl benzoic acid | 16.036 | 108.00 | PH | |
| ui aromatic hydrocarbon | 16.320 | 119.00 | O | |
| ui aromatic hydrocarbon | 16.320 | 119.00 | O | |
| 2-methyl-2-phenyl pentadecane | 16.403 | 105.00 | PH | |
| trans squalene (most likely) sterol precursor | 16.733 | 137.00 | PHT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kacprzyk, M.; Bednarz, B.; Choczyński, M. Attempt to Identify Sex Hormones in the Bodies of Selected Norway Spruce Bark Beetles. Forests 2021, 12, 536. https://doi.org/10.3390/f12050536
Kacprzyk M, Bednarz B, Choczyński M. Attempt to Identify Sex Hormones in the Bodies of Selected Norway Spruce Bark Beetles. Forests. 2021; 12(5):536. https://doi.org/10.3390/f12050536
Chicago/Turabian StyleKacprzyk, Magdalena, Bartłomiej Bednarz, and Maciej Choczyński. 2021. "Attempt to Identify Sex Hormones in the Bodies of Selected Norway Spruce Bark Beetles" Forests 12, no. 5: 536. https://doi.org/10.3390/f12050536