
First Field Record of the Tropical Red-Banded Thrips Selenothrips rubrocinctus (Thripidae: Panchaetothripinae) in Europe

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Specimens and Morphological Identification
2.2. DNA extraction and Amplification
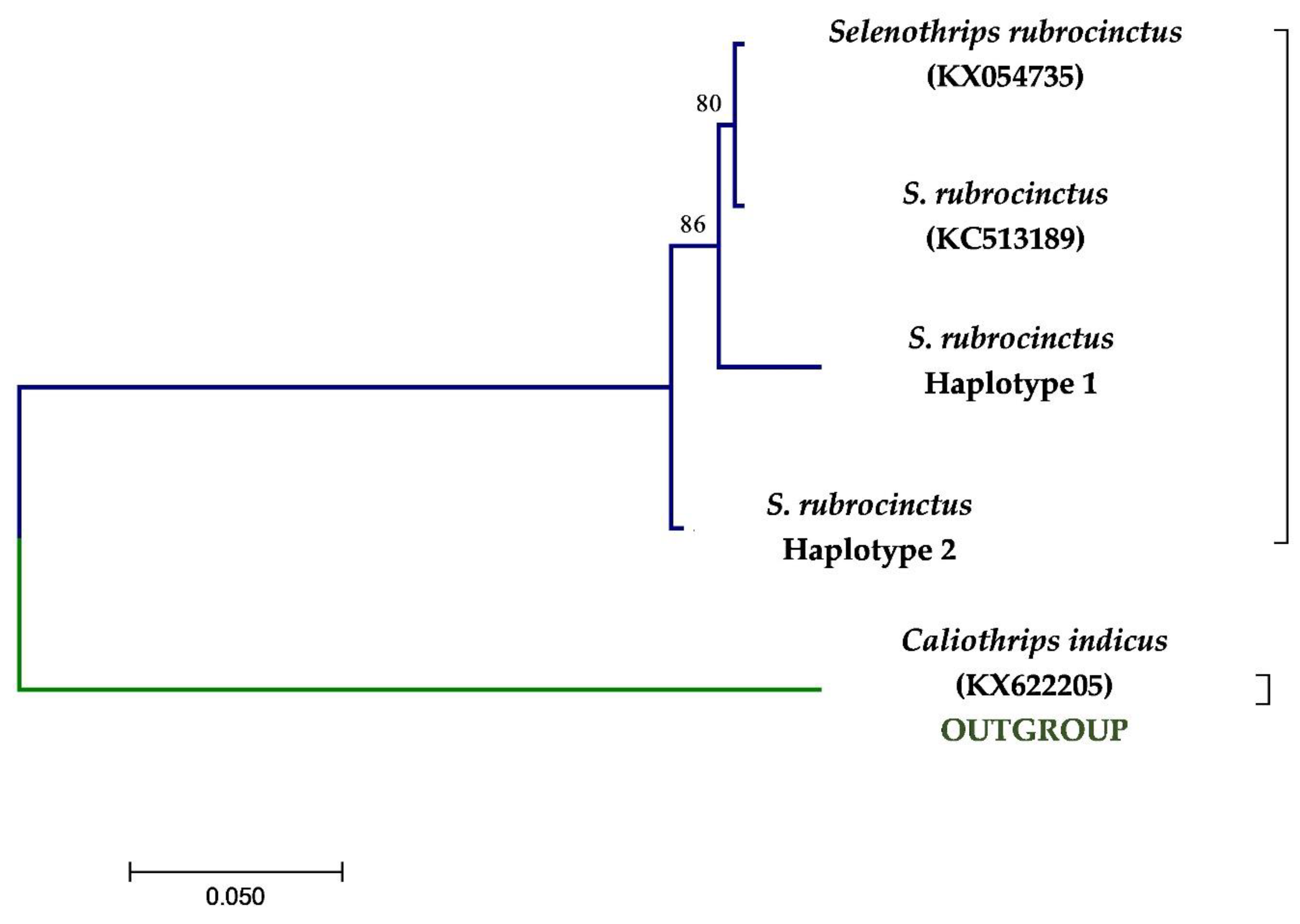
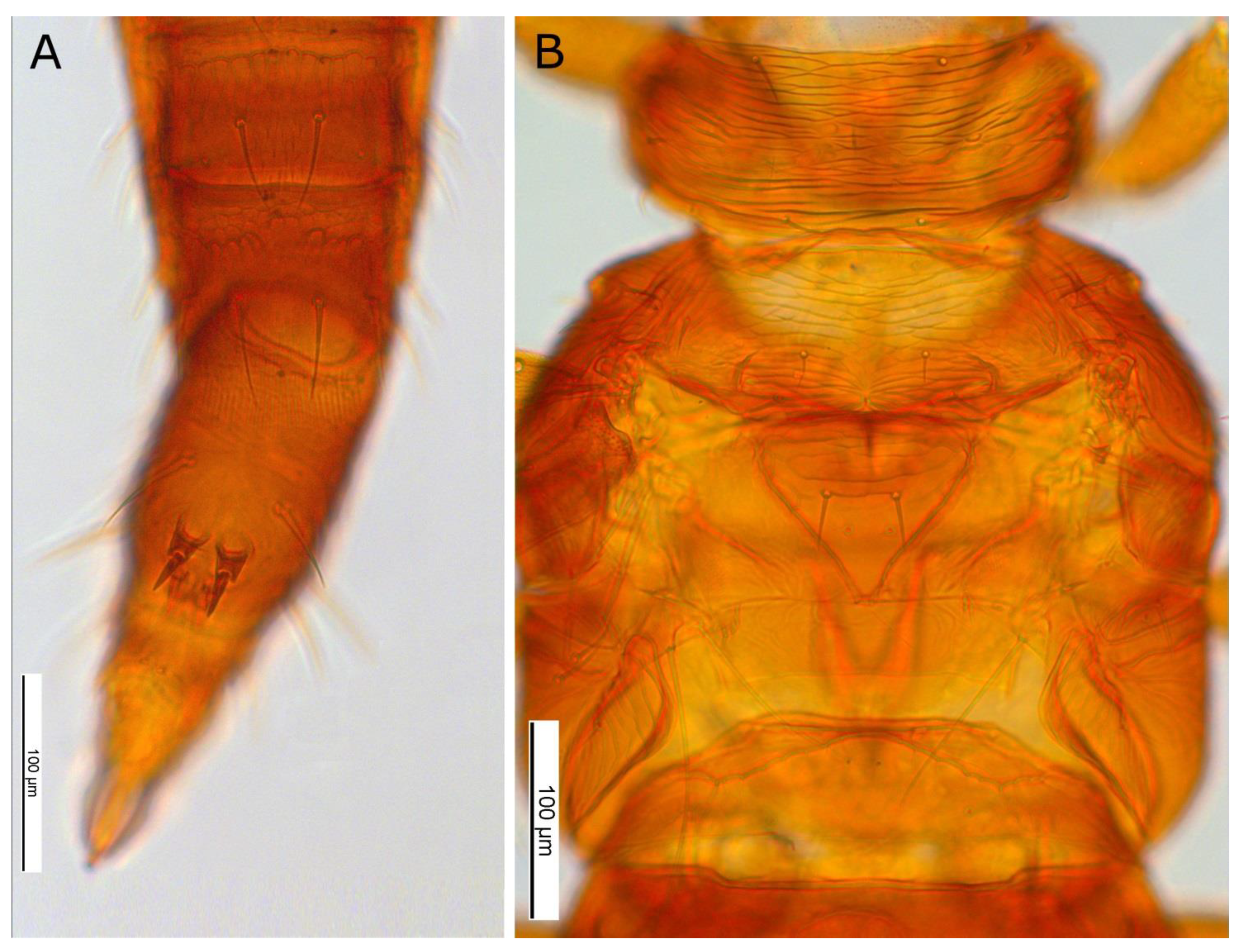
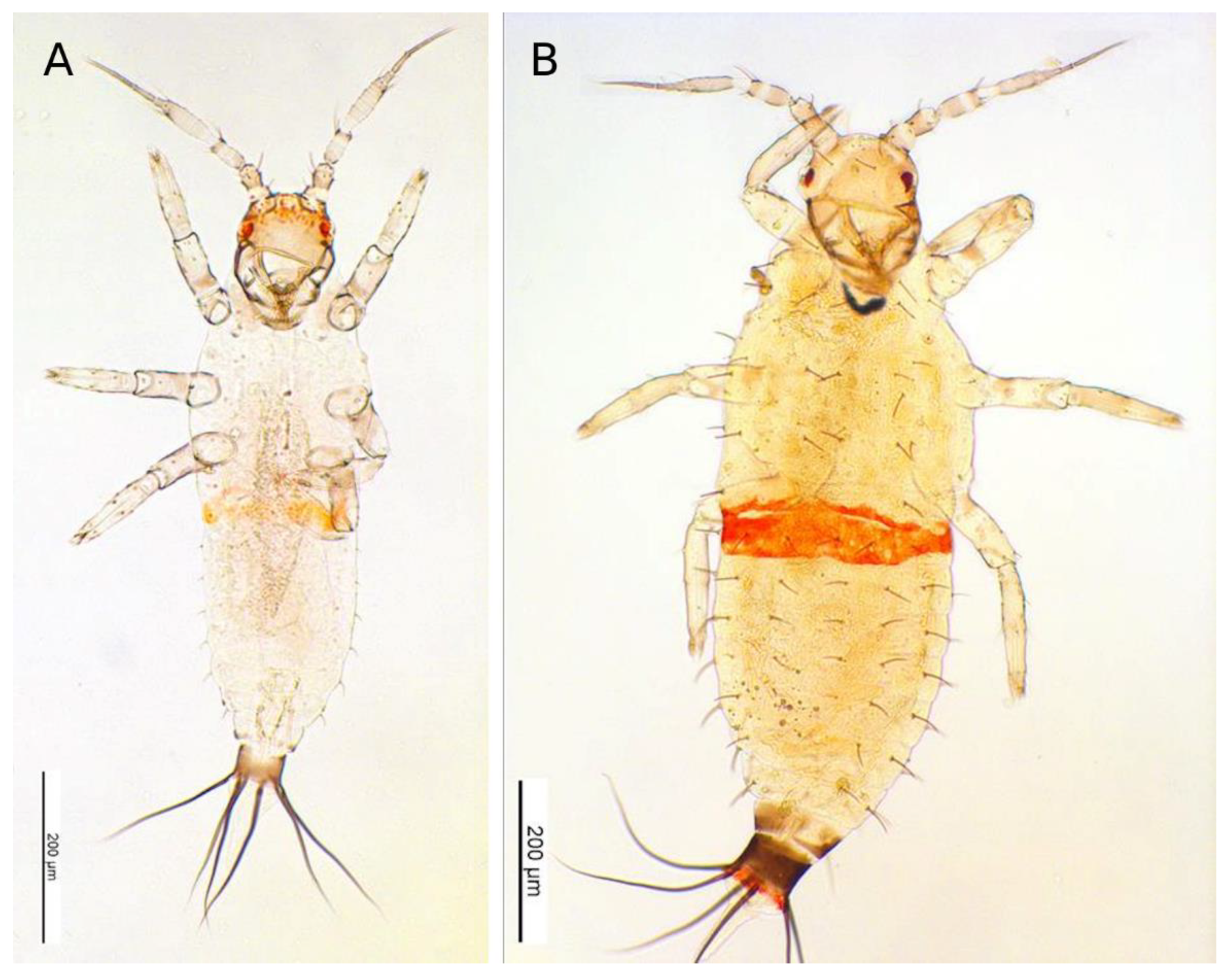
3. Results
3.1. Morphological Characteristics
3.2. Molecular Analysis
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A





References
- Mound, L.A.; Marullo, R. The Thrips of Central and South America: An Introduction. Mem. Entomol. Int. 1996, 6, 1–488. [Google Scholar]
- Grove, T.; Giliomee, J.H.; Pringle, K.L. Thrips (Thysanoptera) species associated with mango trees in South Africa. Afr. Entomol. 2001, 9, 153–162. [Google Scholar]
- Denmark, H.A.; Wolfenbarger, D.O. Redbanded Thrips, Selenothrips rubrocinctus (Giard) (Insects: Thysanoptera: Thripidae); Entomology Circular No. 108; Division of Plant Industry, Florida Department of Agriculture and Consumer Service: Gainesville, FL, USA, 2013.
- Giard, A. Sur un Thrips (Physopus rubrocincta n. sp.) nuisible au cacaoyer.(Thys.). Bull. Soc. Entomol. Fr. 1901, 6, 263–265. [Google Scholar] [CrossRef]
- Dennill, G.B.; Erasmus, M.J. Basis for a practical technique for monitoring thrips in avocado orchards. Crop. Prot. 1992, 11, 89–91. [Google Scholar] [CrossRef]
- Hill, D.S. Agricultural Insect Pests of the Tropicsand their Control, 1st ed.; Cambridge University Press: Cambridge, UK, 1975; p. 528. [Google Scholar]
- Peng, R.K.; Christian, K. The weaver ant, Oecophylla smaragdina (Hymenoptera: Formicidae), an effective biological control agent of the red-banded thrips, Selenothrips rubrocinctus (Thysanoptera: Thripidae) in mango crops in the Northern Territory of Australia. Int. J. Pest. Manag. 2004, 50, 107–114. [Google Scholar] [CrossRef]
- Sánchez-Soto, S.; Nakano, O. Primeiro registro de Selenothrips rubrocinctus (Giard)(Thysanoptera: Thripidae) atacando lichia no Brasil. Neotrop. Entomol. 2004, 33, 395–396. [Google Scholar] [CrossRef] [Green Version]
- Lima, É.F.B.; Thomazini, M.; Santos, R.S.; Lopes, E.N.; Saito, L.; Zucchi, R.A. New findings of thrips (Thysanoptera: Thripidae) on plants in Brazil. Fla. Entomol. 2016, 99, 146–150. [Google Scholar] [CrossRef] [Green Version]
- Haji, F.N.P.; Moreira, A.N.; Ferreira, R.C.F.; Lopes, L.D.M.; de Alencar, J.A.; Barbosa, F.R. Monitoramento e determinação do nível de ação para tripes na cultura da uva. Embrapa Semiárido-Circ. Técnica 2001, 70, 1–8. [Google Scholar]
- Nali, L.R.; Barbosa, F.R.; de Carvalho, C.A.L.; dos Santos, J.B.C. Eficiência de inseticidas naturais e tiametoxam no controle de tripes em videira e seletividade para inimigos naturais. Pestic. Rev. Ecotoxicologia Meio Ambiente 2004, 14, 103–108. [Google Scholar] [CrossRef]
- Moreira, A.N.; Oliveira, J.V.D.; Oliveira, J.E.D.M.; Torres, J.B.; Souza, G.; França, S. Sampling technique for thrips in vineyards. Rev. Bras. Frutic. 2017, 39, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.X. Bionomics and control of Selenothrips rubrocinctus. Acta Entomol. Sinica 1984, 27, 81–86. [Google Scholar]
- Mirab-Balou, M.; Tong, X.L.; Chen, X.X. Thrips species diversity in urban green spaces of Hangzhou (Zhejiang Province), China. J. Entomol. Acarol. Res. 2014, 46, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Etienne, J.; Ryckewaert, P.; Michel, B. Thrips (Insecta: Thysanoptera) of Guadeloupe and Martinique: Updated check-list with new information on their ecology and natural enemies. Fla. Entomol. 2015, 298–304. [Google Scholar] [CrossRef] [Green Version]
- Walter, N.T.; Adeleye, V.O.; Muthomi, P.K.; Rojas, R.J.O.; Strzyzewski, I.; Funderburk, J.; Martini, X. Toxicity of different insecticides against two thrips (Thysanoptera: Thripidae) pests of concern in Central America. Fla. Entomol. 2018, 101, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Reyes, C.P. Thysanoptera (Hexapoda) of the Philippine islands. Raffles Bull. Zool. 1994, 42, 107–507. [Google Scholar]
- Mirab-Balou, M.; Chen, X. The subfamily Panchaetothripinae (Thysanoptera: Thripidae) in Iran, with the first report of genus Selenothrips Karny. Entomotaxonomia 2012, 34, 22–29. [Google Scholar]
- Vierbergen, G. International movement, detection and quarantine of Thysanoptera pests. In Thrips Biology and Management, 1st ed.; Parker, B.L., Skinner, M., Levis, T., Eds.; Plenum Press: New York, NY, USA; London, UK, 1995; Volume 276, pp. 119–132. [Google Scholar]
- Netherlands Plant. Protection Service NPPS. Interceptions at import. Interceptions of plant parasitic insect and mite species. In Netherlands Plant Protection Service Annual Report 1994; Netherlands Plant Protection Service: Utrecht, The Netherlands, 1995; pp. 127–129. [Google Scholar]
- Netherlands Plant Protection Service NPPS. Interceptions at import. Interceptions of plant parasitic insect and mite species. In Netherlands Plant Protection Service Annual Report 1995; Netherlands Plant Protection Service: Utrecht, The Netherlands, 1996; pp. 118–121. [Google Scholar]
- Vierbergen, G. Netherlands Food and Consumer Product Safety Authority, Laboratory of Entomology, P.O. Box 9102, 6700 HC Wageningen, The Netherlands. Unpublished work. 2021. [Google Scholar]
- Collins, D.W. Thysanoptera of Great Britain: A revised and updated checklist. Zootaxa 2010, 2412, 21–41. [Google Scholar] [CrossRef] [Green Version]
- Łabanowski, G. Organizmy Inwazyjne Wykrywane w Polskich Szklarniach. Wciornastki (Thripidae) Instrukcja Rozpoznawania na Podstawie Wyglądu i Objawów Żerowania, 1st ed.; Instytut Ogrodnictwa: Skierniewice, Poland, 2012; pp. 1–40. [Google Scholar]
- Kobro, S. On the Norwegian thrips fauna (Thysanoptera). Nor. J. Entomol. 2003, 50, 17–32. [Google Scholar]
- Mound, L.A.; Kibby, G. Thysanoptera. An. Identification Guide, 2nd ed.; CAB International: Wallingford, UK, 1998; pp. 1–70. [Google Scholar]
- Wilson, T.H. A monography of the subfamily Panchaetothripinae (Thysanoptera: Thripidae). In Monografía de la Subfamilia Panchaetothripinae (Thysanoptera: Thripidae), 1st ed.; Memoirs of the American Entomological Institute: Ann Arbor, MI, USA, 1975; Volume 23, pp. 1–354. [Google Scholar]
- Mirab-Balou, M.; Wang, Z.; Tong, X. Review of the Panchaetothripinae (Thysanoptera: Thripidae) of China, with two new species descriptions. Can. Entomol. 2017, 149, 141–158. [Google Scholar] [CrossRef]
- Vono, G.; Bonsignore, C.P.; Gullo, G.; Marullo, R. Olive production threatened by a resurgent pest Liothrips oleae (Costa, 1857) (Thysanoptera: Phlaeothripidae) in Southern Italy. Insects 2020, 11, 887. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- EPPO. PM 7/129 (1) DNA barcoding as an identification tool for a number of regulated pests. EPPO Bull. 2016, 46, 501–537. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Boil. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA 7: Molecular Evolutionary Genetics Analysis version 7.0 for biggest databases. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Buckman, R.S.; Mound, L.A.; Whiting, M.F. Phylogeny of thrips (Insecta: Thysanoptera) based on five molecular loci. Syst. Entomol. 2013, 38, 123–133. [Google Scholar] [CrossRef]
- Ramage, T.; Martins-Simoes, P.; Mialdea, G.; Allemand, R.; Duplouy, A.; Rousse, P.; Davies, N.; Roderick, G.K.; Charlat, S. A DNA barcode-based survey of terrestrial arthropods in the Society Islands of French Polynesia: Host diversity within the SymbioCode Project. Eur. J. Taxon. 2017, 272, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Morse, J.G.; Hoddle, M.S. Invasion biology of thrips. Annu. Rev. Entomol. 2006, 51, 67–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strapazzon, A. Italian faunal records 368. Boll. Soc. Entomol. Ital. 1999, 131, 259. [Google Scholar]
- Trdan, S. After the first record of Microcephalothrips abdominalis (Crawford) in Slovenia: Presentation of the species and evaluation of its potential economic importance. Zb. Bioteh. Fak. Univ. Ljubl. Kmet. Supl. 2002, 79, 309–316. [Google Scholar]
- Vierbergen, G.; Cean, M.; Szeller, I.B.; Jenser, G.; Masren, T.; Simala, M. Spread of two thrips pests in Europe: Echinothrips americanus and Microcephalothrips abdominalis (Thysanoptera: Thripidae). Acta Phytopathol. Entomol. Hung. 2006, 41, 287–296. [Google Scholar] [CrossRef]
- Zur-Strassen, R. Die Terebranten Thysanoptera Europas und des Mittelmeergebietes; Die Tierwelt Deutschands Verlag Goecke & Evers: Keltern, Germany, 2003; pp. 1–277. [Google Scholar]
- Pizzol, J.; Nammour, D.; Voisin, S.; Ziegler, M.; Desneux, N.; Poncet, C.; Reynaud, P. Survey of thrips species in horticultural greenhouses in Southern France. Acta Hortic. 2012, 952, 785–792. [Google Scholar] [CrossRef]
- Trencheva, K.; Popov, A.; Trenchev, G. Composite thrips, Microcephalothrips abdominalis, a new alien species for Bulgarian fauna. Plant Prot. Sci. 2020, 56, 132–134. [Google Scholar] [CrossRef]
- Reynaud, P.; Balmes, V.; Pizzol, J. Thrips hawaiiensis (Morgan, 1913) (Thysanoptera: Thripidae), an Asian pest thrips now established in Europe. Bull. OEPP 2008, 38, 155–160. [Google Scholar] [CrossRef]
- Goldarazena, A. First record of Thrips hawaiiensis (Morgan, 1913) (Thysanoptera: Thripidae), an Asian pest thrips in Spain. Bull. OEPP 2011, 41, 170–173. [Google Scholar] [CrossRef]
- Marullo, R.; de Grazia, A. Thrips hawaiiensis a pest thrips from Asia newly introduced into Italy. Bull. Insectology 2017, 70, 27–30. [Google Scholar]
- Diez, J.M.; D’Antonio, C.M.; Dukes, J.S.; Grosholz, E.D.; Olden, J.D.; Sorte, C.J.; Blumental, D.M.; Early, R.; Ibáñez, I.; Jones, S.J.; et al. Will extreme climatic events facilitate biological invasions? Front. Ecol. Environ. 2012, 10, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzales, P.; Grosholz, E.D.; Ibáñez, I.; Miller, L.P.; et al. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- Fennah, R.G. The influence of environmental stress on the cacao tree in predetermining the feeding sites of cacao thrips, Selenothrips rubrocinctus (Giard) on leaves and pods. Bull. Entomol. 1965, 56, 333–349. [Google Scholar] [CrossRef]
- Krueger, S.; Mound, L.A.; Moritz, G.B. Offspring sex ratio and development are determined by copulation activity in Echinothrips americanus Morgan 1913 (Thysanoptera: Thripidae). J. Appl. Entomol. 2016, 140, 462–473. [Google Scholar] [CrossRef]
- Xiao-Wei, L.; Xiao-Chen, Z.; Hong-Xue, J.; Ji-Nian, F. Comparisons of developmental and reproductive biology between parthenogenetic and sexual Echinothrips americanus (Thysanoptera: Thripidae). Environ. Entomol. 2016, 41, 706–713. [Google Scholar] [CrossRef]
- Marullo, R.; Mercati, F.; Vono, G. DNA Barcoding: A Reliable Method for the Identification of Thrips Species (Thysanoptera, Thripidae) Collected on Sticky Traps in Onion Fields. Insects 2020, 11, 489. [Google Scholar] [CrossRef] [PubMed]
- Funderburk, J.; Diffie, S.; Sharma, J.; Hodges, A.; Osborne, L. Thrips of Ornamentals in the Southeastern US; ENY-845 (IN754); Entomology & Nematology Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2007. [Google Scholar]
- Bradshaw, C.J.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Barbet-Massin, M.; Salles, J.M.; Simard, F.; Courchamp, F. Massive yet grossly underestimated global costs of invasive insects. Nat. Commun. 2016, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | Source | Reference |
|---|---|---|---|
| LCO-1490 (fw) | GGTCAACAAATCATAAAGATATTGG | Folmer et al.,1994 | [30] |
| LCO1490puc-t1 (fw) | caggaaacagctatgaccTTTCAACWAATCATAAAGATATTGG | EPPO, 2016 | [31] |
| LCO1490Hem1-t1 (fw) | caggaaacagctatgaccTTTCAACTAAYCATAARGATATYGG | EPPO, 2016 | [31] |
| HCO-2198 (rv) | TAAACTTCAGGGTGACCAAAAAATCA | Folmer et al.,1994 | [30] |
| HCO2198puc-t1 (rv) | tgtaaaacgacggccagtTAAACTTCWGGRTGWCCAAARAATCA | EPPO, 2016 | [31] |
| HCO2198Hem1-t1 (rv) | tgtaaaacgacggccagtTAAACYTCDGGATGBCCAAARAATCA | EPPO, 2016 | [31] |
| HCO2198Hem2-t1 (rv) | tgtaaaacgacggccagtTAAACYTCAGGATGACCAAAAAAYCA | EPPO, 2016 | [31] |
| Sample Nr. | Location | Lat., Long. | Host Plant | Material Sampled |
|---|---|---|---|---|
| 5683090 | Bagnolo Cremasco (1) | 45°21′39.1″ N 9°36′31.4″ E | Carpinus betulus | 2♀, 1♂, 2 larvae II, 4 larvae I |
| 5683146 | Bagnolo Cremasco (2) | 45°21′40.2″ N 9°36′32.2″ E | Carpinus betulus | 4♀, 1♂ |
| 5683082 | Cremosano | 45°22′49.5″ N 9°39′13.8″ E | Koelreuteria paniculata | 3♀, 2♂, 4 larvae II, 6 larvae I |
| 5683121 | Crespiatica (1) | 45°21′37.7″ N 9°34′22.9″ E | Acer campestre | 1♀, 4 larvae I |
| 5683103 * | Crespiatica (2) | 45°21′37.6″ N 9°34′22.4″ E | Liquidambar styraciflua | 7♀, 2 larvae II, 6 larvae I |
| 5683154 | Pandino | 45°24′05.3″ N 9°33′34.4″ E | Liquidambar styraciflua | 5♀, 10 larvae I |
| 5683138 * | Palazzo Pignano (Scannabue) | 45°23′13.9″ N 9°35′29.2″ E | Parrotia persica | 1♂, 3 larvae I |
| 26082016 | Trescore Cremasco | 45°24′06.1″ N 9°37′40.2″ E | Carpinus betulus | 16♀, 5♂, 10 larvae I |
| Origin * | Guadeloupe Neotype | Italy 2019(1) | Italy 2019(2) | Surinam 1960 | Surinam 2011 | Vietnam 2018 | Indonesia 1936 |
|---|---|---|---|---|---|---|---|
| Body length | 1300 | 1515 | 1285 | 1457 | 1215 | 1375 | 1415 |
| Forewing -length -medial width | 755 49 | 837 57 | 800 56 | 728 55 | 745 55 | 746 49 | 714 51 |
| Antenna (l/w) | |||||||
| I II III IV V VI VII VIII | 24/28–0.9 48/36–1.3 62/26–2.4 73/24–3.0 42/24–1.8 29/22–1.3 17/13–1.3 34/6–5.7 | 28/31–0.9 53/36–1.5 65/27–2.4 66/26–2.5 41/25–1.6 32/20–1.6 18/11–1.6 31/6–5.24 | 24/25–1.0 51/33–1.5 58/30–1.9 62/26–2.4 37/26–1.4 27/22–1.2 14/13–1.1 26/6–4.3 | 21/32–0.7 45/42–1.1 59/29–2.0 65/26–2.5 40/25–1.6 28/23–1.2 15/13–1.2 29/6–4.8 | 21/24–0.8 46/34–1.4 55/26–2.1 68/25–2.7 42/23–1.8 26/20–1.3 13/13–1.0 24/6–4.0 | 22/25–0.9 45/36–1.3 60/26–2.3 61/25–2.4 38/25–1.5 29/26–1.1 13/10–1.3 26/6–4.3 | 22/28–0.8 45/33–1.4 58/27–2.1 63/23–2.7 38/23–1.7 29/19–1.5 11/>8–<1.4 26/>3–<8.7 |
| Total length | 329 | 334 | 299 | 302 | 295 | 294 | 292 |
| Haplotype | Number of Samples Sequenced | Host Plant | Location |
|---|---|---|---|
| Haplotype 1 | 1 | Carpinus betulus | Bagnolo Cremasco (1) |
| 1 | Carpinus betulus | Bagnolo Cremasco (2) | |
| 1 | Koelreuteria paniculata | Cremosano | |
| 1 | Acer campestre | Crespiatica (1) | |
| 1 | Liquidambar styraciflua | Crespiatica (2) | |
| 1 | Liquidambar styraciflua | Pandino | |
| 1 | Parrotia persica | Palazzo Pignano (Scannabue) | |
| 12 | Carpinus betulus | Trescore Cremasco | |
| Haplotype 2 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taddei, A.; Vono, G.; Vierbergen, G.; Wojnar, A.; Zugno, M.; Marullo, R. First Field Record of the Tropical Red-Banded Thrips Selenothrips rubrocinctus (Thripidae: Panchaetothripinae) in Europe. Forests 2021, 12, 1484. https://doi.org/10.3390/f12111484
Taddei A, Vono G, Vierbergen G, Wojnar A, Zugno M, Marullo R. First Field Record of the Tropical Red-Banded Thrips Selenothrips rubrocinctus (Thripidae: Panchaetothripinae) in Europe. Forests. 2021; 12(11):1484. https://doi.org/10.3390/f12111484
Chicago/Turabian StyleTaddei, Andrea, Gregorio Vono, Gijsbertus Vierbergen, Andrea Wojnar, Matteo Zugno, and Rita Marullo. 2021. "First Field Record of the Tropical Red-Banded Thrips Selenothrips rubrocinctus (Thripidae: Panchaetothripinae) in Europe" Forests 12, no. 11: 1484. https://doi.org/10.3390/f12111484