
High Genetic Diversity of Shorea acuminata Dyer in the Rehabilitated Area of a Degraded Lowland Dipterocarp Tropical Rainforest

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Plot and Sample Collection
2.2. DNA Extraction and Microsatellite Analysis
2.3. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akujärvi, A.; Repo, A.; Akujärvi, A.M.; Liski, J. Bridging mapping and simulation modelling in the ecosystem service assessments of boreal forests: Effects of bioenergy production on carbon dynamics. For. Ecosyst. 2021, 8, 4. [Google Scholar] [CrossRef]
- Rusterholz, H.-P.; Studer, M.; Zwahlen, V.; Baur, B. Plant-mycorrhiza association in urban forests: Effects of the degree of urbanisation and forest size on the performance of sycamore (Acer pseudoplatanus) saplings. Urban For. Urban Green. 2020, 56, 126872. [Google Scholar] [CrossRef]
- Silva, L.N.; Freer-Smith, P.; Madsen, P. Production, restoration, mitigation: A new generation of plantations. New For. 2019, 50, 153–168. [Google Scholar] [CrossRef]
- The Star. Ways to Improve the Sustainability of the Timber Industry. 21 March 2021. Available online: https://www.thestar.com.my/news/nation/2021/03/21/charting-the-direction-for-timber (accessed on 11 June 2021).
- United Nations Department of Economic and Social Affairs. The World Population Prospects: 2015 Revision. 29 July 2015. Available online: https://www.un.org/en/development/desa/publications/world-population-prospects-2015-revision.html (accessed on 1 June 2021).
- Food and Agriculture Organization of the United Nations (FAO). The State of the World’s Forests 2018—Forest Pathways to Sustainable Development. 2018. Available online: http://www.fao.org/documents/card/en/c/I9535EN (accessed on 25 June 2021).
- Keenan, R.J. Climate change impacts and adaptation in forest management: A review. Ann. For. Sci. 2015, 72, 145–167. [Google Scholar] [CrossRef] [Green Version]
- Ken, S.; Sasaki, N.; Entani, T.; Ma, H.O.; Thuch, P.; Tsusaka, T.W. Assessment of the local perceptions on the drivers of deforestation and forest degradation, agents of drivers, and appropriate activities in Cambodia. Sustainability 2020, 12, 9987. [Google Scholar] [CrossRef]
- Potter, K.M.; Jetton, R.M.; Bower, A.; Jacobs, D.F.; Man, G.; Hipkins, V.D.; Westwood, M. Banking on the future: Progress, challenges and opportunities for the genetic conservation of forest trees. New For. 2017, 48, 153–180. [Google Scholar] [CrossRef]
- Kettle, C.J. Ecological considerations for using dipterocarps for restoration of lowland rainforest in Southeast Asia. Biodivers. Conserv. 2010, 19, 1137–1151. [Google Scholar] [CrossRef]
- Espeland, E.K.; Emery, N.C.; Mercer, K.L.; Woolbright, S.A.; Kettenring, K.M.; Gepts, P.; Etterson, J.R. Evolution of plant materials for ecological restoration: Insights from the applied and basic literature. J. Appl. Ecol. 2017, 54, 102–115. [Google Scholar] [CrossRef]
- Gauli, A.; Gailing, O.; Stefenon, V.M.; Finkeldey, R. Genetic similarity of natural populations and plantations of Pinus roxburghii Sarg. in Nepal. Ann. For. Sci. 2009, 66, 703. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.H.; Chen, X.Y.; Zhang, X.; Shen, D.W. A population genetic evaluation of ecological restoration with the case study on Cyclobalanopsis myrsinaefolia (Fagaceae). Plant Ecol. 2008, 197, 31–41. [Google Scholar] [CrossRef]
- Sujii, P.S.; Schwarcz, K.D.; Grando, C.; de Aguiar Silvestre, E.; Mori, G.M.; Brancalion, P.H.S.; Zucchi, M.I. Recovery of genetic diversity levels of a Neotropical tree in Atlantic Forest restoration plantations. Biol. Conserv. 2017, 211, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Cheah, L.C. A note on taungya in Negeri Sembilan with particulars reference to the incidence of damage by oviposition of insects in plantation in Kenaboi Forest Reserve. Malay. For. 1971, 34, 133–153. [Google Scholar]
- Shaharuddin, M.I.; Mohd Basri, H.; Samsudin, S. An evaluation of a 19-year old taungya planting in Negeri Sembilan Darul Khusus. Malays. For. 1992, 55, 1–12. [Google Scholar]
- Abd Ghani, A.R.; Najib, L.A. Agroforestry approach of planting timber and non-timber species in Malaysia. In Development of Agroforestry Technology for the Rehabilitation of Tropical Forests; JIRCAS Working Paper No. 60; Gotoh, T., Yakota, Y., Eds.; Japan International Research Centre for Agricultural Sciences (JIRCAS): Tsukuba, Japan, 2009; pp. 125–135. [Google Scholar]
- Krishnapillay, D.B.; Razak, M.A.A.; Appanah, S. Forest rehabilitation—the Malaysian experience. In Proceeding of Keep Asia Green; Don, K.L., Ed.; IUFRO Headquaters: Vienna, Austria, 2007; Volume 1, pp. 85–123. [Google Scholar]
- Fatma, N.A.H.; Wan Juliana, W.A.; Shaharuddin, M.I.; Wickneswari, R. Stand structure of Shorea and spatial distribution of Shorea acuminata in a rehabilitated area of Kenaboi Forest Reserve. J. Trop. For. Sci. 2020, 32, 257–267. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Pythochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Lee, S.L.; Tani, N.; Ng, K.K.S.; Tsumura, Y. Isolation and characterization of 20 microsatellite loci for an important tropical tree Shorea leprosula (Dipterocarpaceae) and their applicability to S. parvifolia. Mol. Ecol. Notes 2004, 4, 222–225. [Google Scholar] [CrossRef]
- Ng, K.K.S.; Lee, S.L.; Tsumura, Y.; Ueno, S.; Ng, C.H.; Lee, C.T. Expressed sequence tag–simple sequence repeats isolated from Shorea leprosula and their transferability to 36 species within the Dipterocarpaceae. Mol. Ecol. Resour. 2009, 9, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Appanah, S.; Rasol, M.M.A. Smaller trees can fruit in logged dipterocarp forests. J. Trop. For. Sci. 1990, 3, 80–87. [Google Scholar]
- Marshall, T.C.; Slate, J.; Kruuk, L.E.B.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef] [Green Version]
- Dakin, E.E.; Avise, J.C. Microsatellite null alleles in parentage analysis. Heredity 2004, 93, 504–509. [Google Scholar] [CrossRef]
- Liu, X.H.; Yao, Y.G. Characterization of 12 polymorphic microsatellite markers in the Chinese tree shrew (Tupaia belangeri chinensis). Dongwuxue Yanjiu (Zool.Res.) 2013, 34, E62–E68. [Google Scholar] [CrossRef] [Green Version]
- Naito, Y.; Kanzaki, M.; Iwata, H.; Obayashi, K.; Lee, S.L.; Muhammad, N.; Okuda, T.; Tsumura, Y. Density-dependent selfing and its effects on seed performance in a tropical canopy tree species, Shorea acuminata (Dipterocarpaceae). For. Ecol. Manag. 2008, 256, 375–383. [Google Scholar] [CrossRef]
- Kondo, T.; Nishimura, S.; Tani, N.; Ng, K.K.S.; Lee, S.L.; Muhammad, N.; Okuda, T.; Tsumura, Y.; Isagi, Y. Complex pollination of a tropical Asian rainforest canopy tree by flower-feeding thrips and thrips-feeding predators. Am. J. Bot. 2016, 103, 1912–1920. [Google Scholar] [CrossRef] [PubMed]
- Casu, R.; Dimmock, C.; Thomas, M.; Bower, N.; Knight, D.; Grof, C.; McIntyre, L.; Jackson, P.; Jordan, D.; Whan, V.; et al. Genetic and expression profiling in sugarcane. In International Society of Sugar Cane Technologists, Vol. Ii, Proceedings, Brisbane, Australia, 17–21 September 2001; Australian Society of Sugar Cane Technologists: Mackay, Australia, 2001; pp. 542–546. [Google Scholar]
- Fukue, Y.; Kado, T.; Lee, S.L.; Ng, K.K.S.; Muhammad, N.; Tsumura, Y. Effects of flowering tree density on matinng system and gene flow in Shorea leprosula (Dipterocarpaceae) in Peninsular Malaysia. J. Plant Res. 2007, 120, 413–420. [Google Scholar] [CrossRef]
- Zeng, X.; Fischer, G.A. Using multiple seedlots in restoration planting enhances genetic diversity compared to natural regeneration in fragmented tropical forests. For. Ecol. Manag. 2021, 482, 118819. [Google Scholar] [CrossRef]
- Ang, C.C.; O’Brien, M.J.; Ng, K.K.S.; Lee, P.C.; Hector, A.; Schmid, B.; Shimizu, K.K. Genetic diversity of two tropical tree species of the Dipterocarpaceae following logging and restoration in Borneo: High genetic diversity in plots with high species diversity. Plant Ecol. Divers. 2016, 9, 459–469. [Google Scholar] [CrossRef]
- Widiyatno; Indrioko, S.; Na’iem, M.; Uchiyama, K.; Numata, S.; Ohtani, M.; Matsumoto, A.; Tsumura, Y. Effects of different silvicultural systems on the genetic diversity of Shorea parvifolia populations in the tropical rainforest of Southeast Asia. Tree Genet. Genomes 2016, 12, 73. [Google Scholar] [CrossRef]
- André, T.; Lemes, M.R.; Grogan, J.; Gribel, R. Post-logging loss of genetic diversity in a mahogany (Swietenia macrophylla King, Meliaceae) population in Brazilian Amazonia. For. Ecol. Manag. 2008, 255, 340–345. [Google Scholar] [CrossRef]
- Shaharudin, M.I. Forest management systems in Southest Asia. In Managing the Future of Southest Asia’s Valuable Tropical Rainforest: A Practitioner’s Guide to Forest Genetics; Wickneswari, R., Cannon, C., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 57–68. ISBN 978-94-007-2175-3. [Google Scholar]




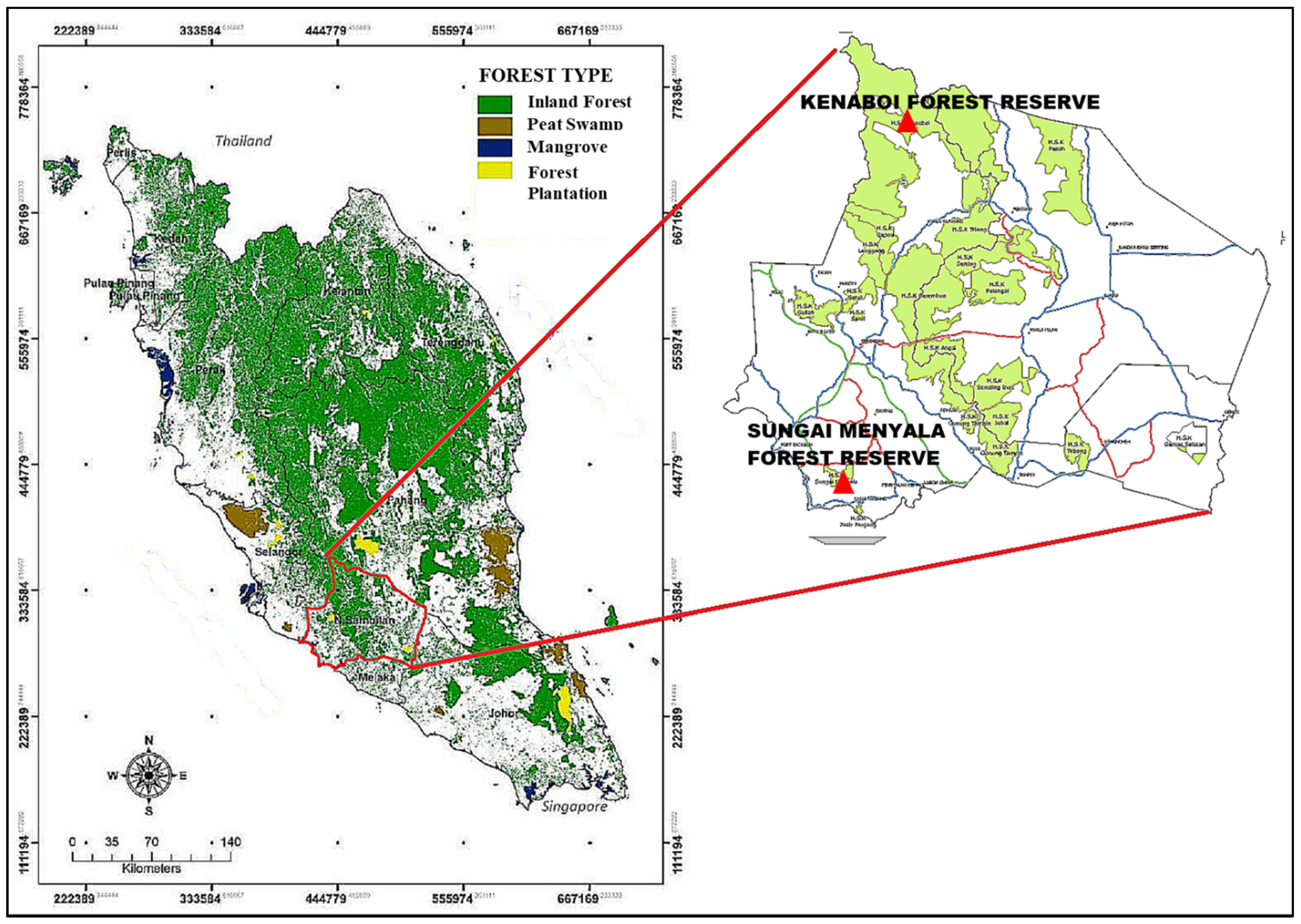{kind=link}
{kind=link}
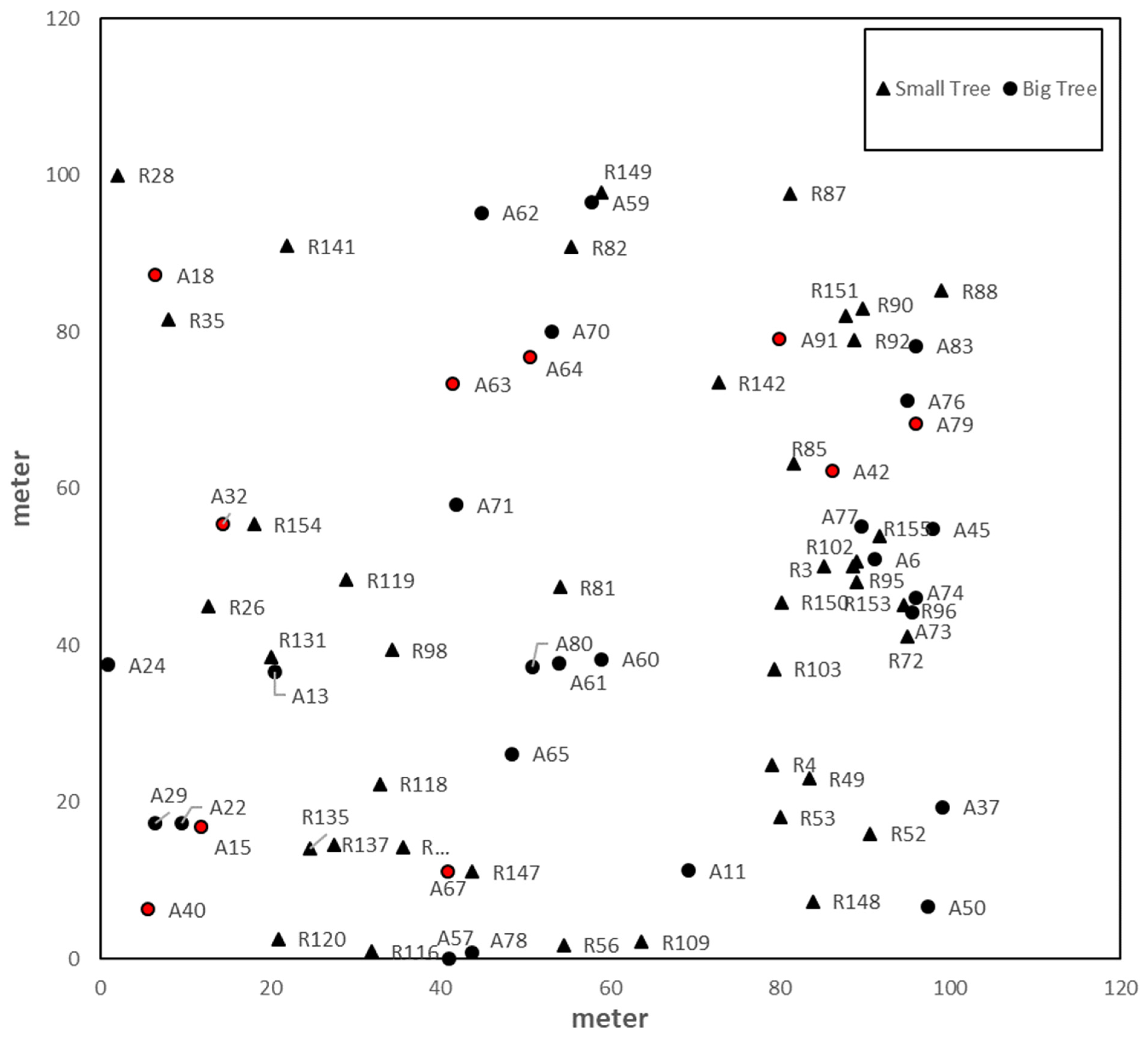{kind=link}
| Loci | Accession No. | Primer Sequence (5′-3′) | Repeat | Allele Size Range (bp) | Annealing Temp. (°C) |
|---|---|---|---|---|---|
| Sle280 | AJ616880 | F: GCAACTAAAATGGACCAGA | |||
| R: GAGTAAGGTGGCAGATATAGAG | (CT)7 | 107–137 | 52 | ||
| Sle 392 | AJ616886 | F: ATGTCCTTGAAGATGTAAAGTGGGTG | |||
| R: AATAATGGAAGTGAGACGAGGCTG | (GA)11 | 161–231 | 55 | ||
| Sle 475 | AJ616888 | F: AGCGAAACCCTTGTGGAGA | |||
| R: GAGACTACGGTGGCGACGA | (GA)10 | 129–139 | 50 | ||
| Sle 566 | AJ616890 | F: TGAGTAACAAGTAATGAGGG | |||
| R: GCAGAGATTGAAACAGAAG | (GA)13 | 59–104 | 52 | ||
| SleE02 | DC649188 | F: GGAGGAGAGAAACGAAG | |||
| R: GTTTGAGGTAGTGAATAACGAGC | (AGC)9 | 142–160 | 45 | ||
| SleE07 | DC649404 | F: AGAAGAATATGGGTACGACTG | |||
| R: GTTTGAATCAACTGGCACCTCTAT | (GAA)7 | 175–190 | 45 | ||
| SleE16 | DC651058 | F: TCGTCAACCTCCGTAGTCC | |||
| R: GTTTGCGCAATAAATAGAGCAATCA | (CT)12 | 184–192 | 45 |
| Panel | Loci | Allele Size | Fluorescent | Peak Colour |
|---|---|---|---|---|
| Panel 1 | Sle280 | 107–137 | TAMRA | Yellow |
| Sle392 | 161–231 | TAMRA | Yellow | |
| Sle566 | 59–104 | FAM | Blue | |
| Panel 2 | Sle475 | 129–139 | FAM | Blue |
| SleE02 | 142–160 | TAMRA | Yellow | |
| SleE07 | 175–190 | ROX | Red | |
| SleE16 | 184–192 | HEX | Green |
| Locus | Number of Alleles | Allele Size Range (bp) | He | Ho | Fis | PIC | Null Allele Frequency | HW |
|---|---|---|---|---|---|---|---|---|
| Big tree | n = 34 | |||||||
| Sle280 | 4 | 111–128 | 0.335 | 0.324 | 0.033 | 0.309 | 0.05 | NS |
| Sle392 | 7 | 175–187 | 0.784 | 0.765 | 0.024 | 0.738 | 0.00 | NS |
| Sle475 | 8 | 126–187 | 0.593 | 0.647 | −0.091 | 0.557 | −0.08 | NS |
| Sle566 | 12 | 63–114 | 0.697 | 0.676 | 0.030 | 0.652 | 0.01 | NS |
| SleE07 | 5 | 180–188 | 0.683 | 0.794 | −0.163 | 0.606 | −0.08 | NS |
| SleE16 | 9 | 190–203 | 0.845 | 0.529 | 0.374 | 0.813 | 0.23 | NS |
| Mean | 6.5 | 0.656 (±0.180) | 0.623 (±0.174) | 0.035 (±0.184) | 0.613 (±0.178) | |||
| Small tree | n = 41 | |||||||
| Sle280 | 3 | 111–128 | 0.319 | 0.268 | 0.160 | 0.293 | 0.06 | NS |
| Sle392 | 6 | 175–187 | 0.773 | 0.78 | −0.009 | 0.728 | −0.01 | NS |
| Sle475 | 7 | 126–187 | 0.533 | 0.39 | 0.268 | 0.505 | 0.13 | NS |
| Sle566 | 10 | 63–114 | 0.765 | 0.634 | 0.171 | 0.725 | 0.09 | NS |
| SleE07 | 5 | 180–188 | 0.700 | 0.561 | 0.199 | 0.635 | 0.11 | NS |
| SleE16 | 9 | 190–203 | 0.820 | 0.659 | 0.196 | 0.786 | 0.10 | NS |
| Mean | 6.7 | 0.652 (±0.191) | 0.549 (±0.188) | 0.164 (±0.09) | 0.612 (±0.186) |
| Observed Allele | Tree < 30 cm DBH * | ||||
|---|---|---|---|---|---|
| Diameter | Locus | Tree ≥ 30 cm DBH | Tree < 30 cm DBH | In Study Plot (%) | Outside Study Plot (%) |
| Biggest 10% | SLEE07 | A, B, C, D | A, B, C, D | 100 | 0 |
| (52–60 cm) | SLEE16 | A, B, C, D, E, F | A, B, C, D, E, F, G | 86 | 14 |
| n = 8 | SLE280 | A, B, C | A, B, C | 100 | 0 |
| SLE392 | A, B, C, D, E | A, B, C, D, E | 100 | 0 | |
| SLE475 | A, B, C, D | A, B, C, D, E, F, G | 57 | 43 | |
| SLE566 | A, B, C, D, E | A, B, C, D, E, F, G | 71 | 29 | |
| Mean | 86 | 14 | |||
| Biggest 20% | SLEE07 | A, B, C, D | A, B, C, D, | 100 | 0 |
| (46–60 cm) | SLEE16 | A, B, C, D, E, F | A, B, C, D, E, F, G | 86 | 14 |
| n = 15 | SLE280 | A, B, C | A, B, C | 100 | 0 |
| SLE392 | A, B, C, D, E | A, B, C, D, E | 100 | 0 | |
| SLE475 | A, B, C, D, E, F | A, B, C, D, E, F, G | 86 | 14 | |
| SLE566 | A, B, C, D, E | A, B, C, D, E, F, G | 71 | 29 | |
| Mean | 90.5 | 9.5 | |||
| Biggest 30% | SLEE07 | A, B, C, D | A, B, C, D | 100 | 0 |
| (42–60 cm) | SLEE16 | A, B, C, D, E, F, G, H | A, B, C, D, E, F, G, H | 100 | 0 |
| n = 24 | SLE280 | A, B, C | A, B, C | 100 | 0 |
| SLE392 | A, B, C, D, E | A, B, C, D, E | 100 | 0 | |
| SLE475 | A, B, C, D, E, F, G | A, B, C, D, E, F, G | 100 | 0 | |
| SLE566 | A, B, C, D, E, F, G | A, B, C, D, E, F, G | 100 | 0 | |
| Mean | 100 | 0 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd Hamid, F.N.; Wan Ahmad, W.J.; Mohamad Ismail, S.; Ratnam, W. High Genetic Diversity of Shorea acuminata Dyer in the Rehabilitated Area of a Degraded Lowland Dipterocarp Tropical Rainforest. Forests 2021, 12, 1344. https://doi.org/10.3390/f12101344
Abd Hamid FN, Wan Ahmad WJ, Mohamad Ismail S, Ratnam W. High Genetic Diversity of Shorea acuminata Dyer in the Rehabilitated Area of a Degraded Lowland Dipterocarp Tropical Rainforest. Forests. 2021; 12(10):1344. https://doi.org/10.3390/f12101344
Chicago/Turabian StyleAbd Hamid, Fatma Nadiah, Wan Juliana Wan Ahmad, Shaharuddin Mohamad Ismail, and Wickneswari Ratnam. 2021. "High Genetic Diversity of Shorea acuminata Dyer in the Rehabilitated Area of a Degraded Lowland Dipterocarp Tropical Rainforest" Forests 12, no. 10: 1344. https://doi.org/10.3390/f12101344