Domesticating Commercially Important Native Tree Species in the Philippines: Early Growth Performance Level



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Selection of Native Species
2.3. Experimental Design, Planting and Measurements
2.4. Data Analysis
3. Results
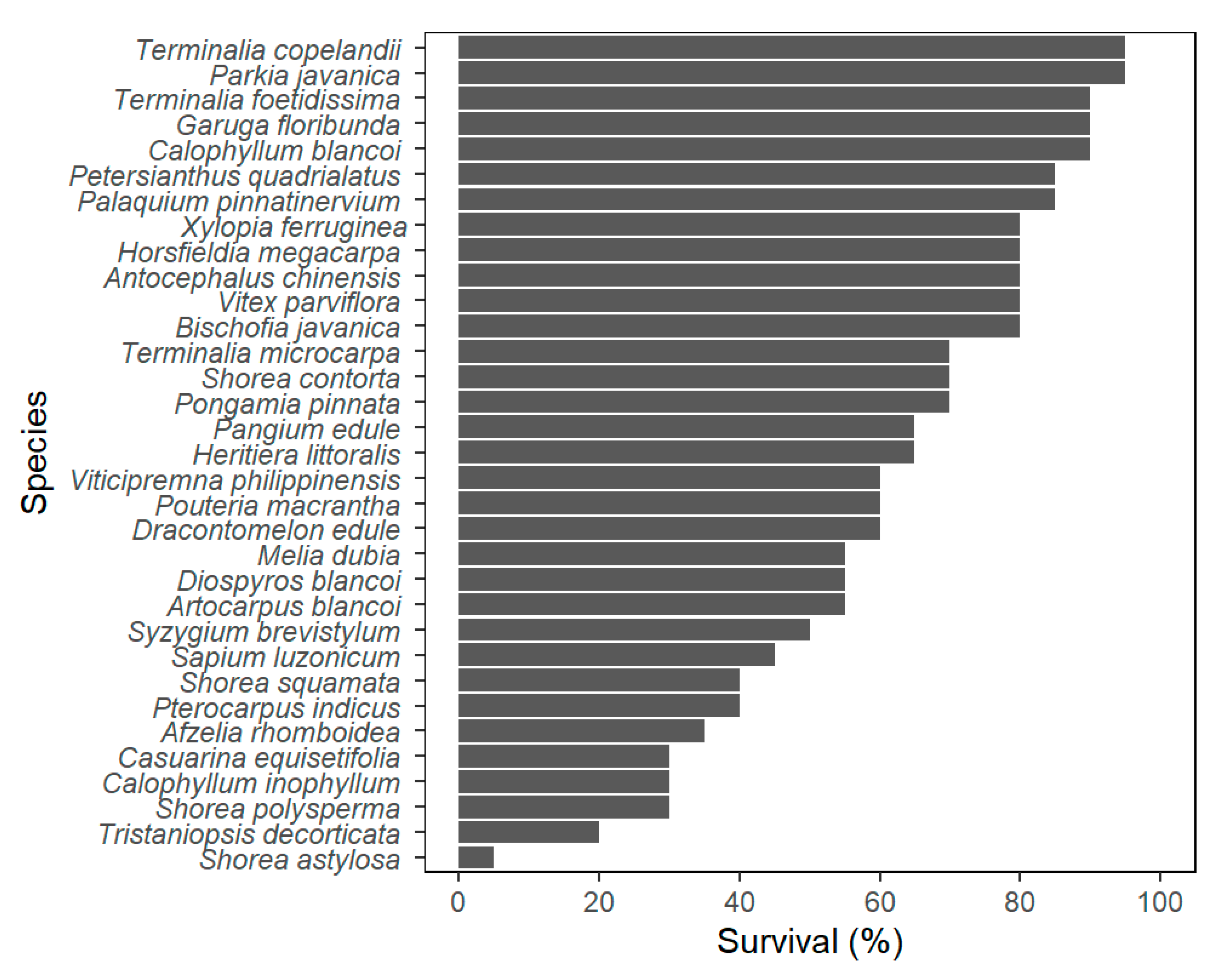
3.1. Survival Percentage of Species on Trial
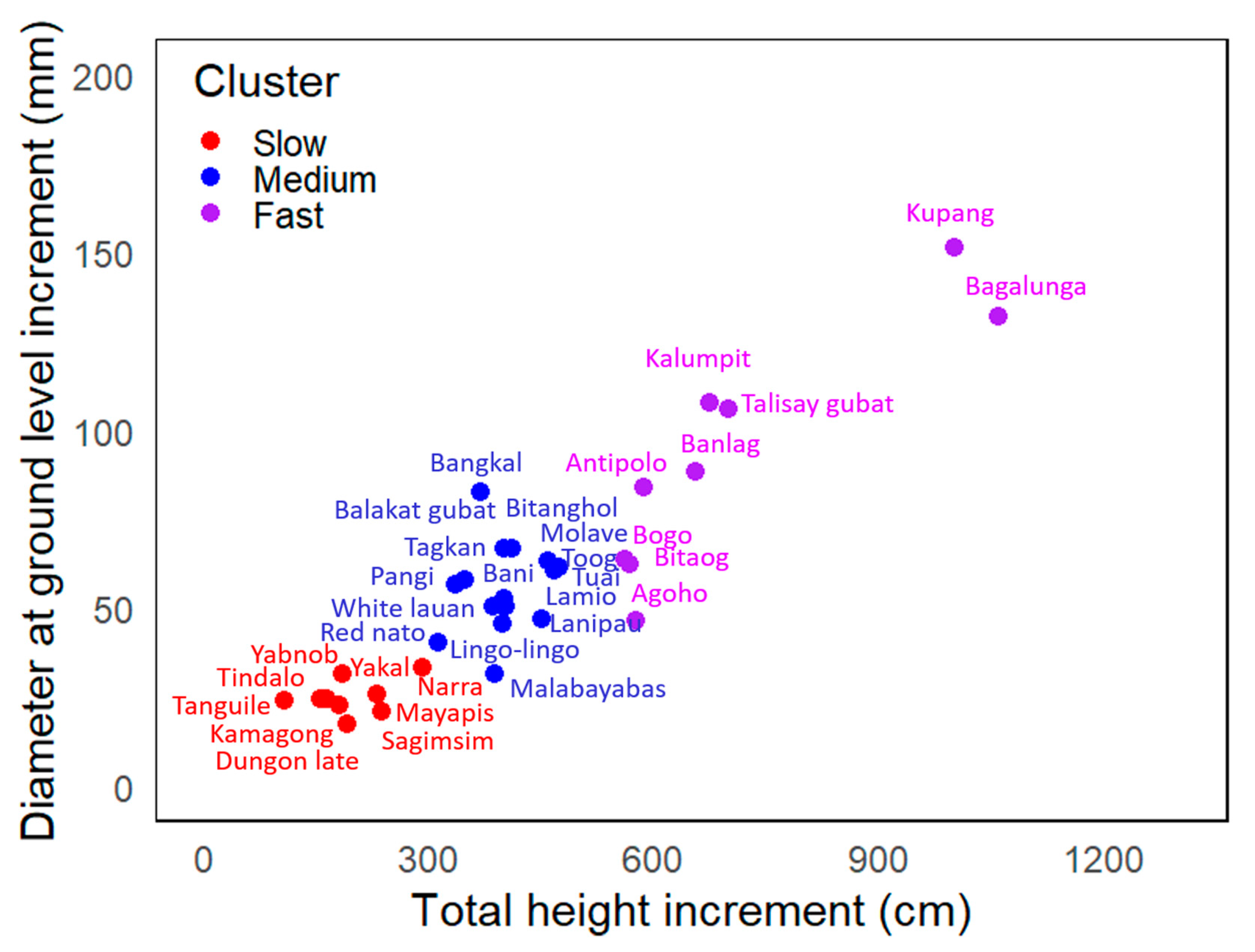
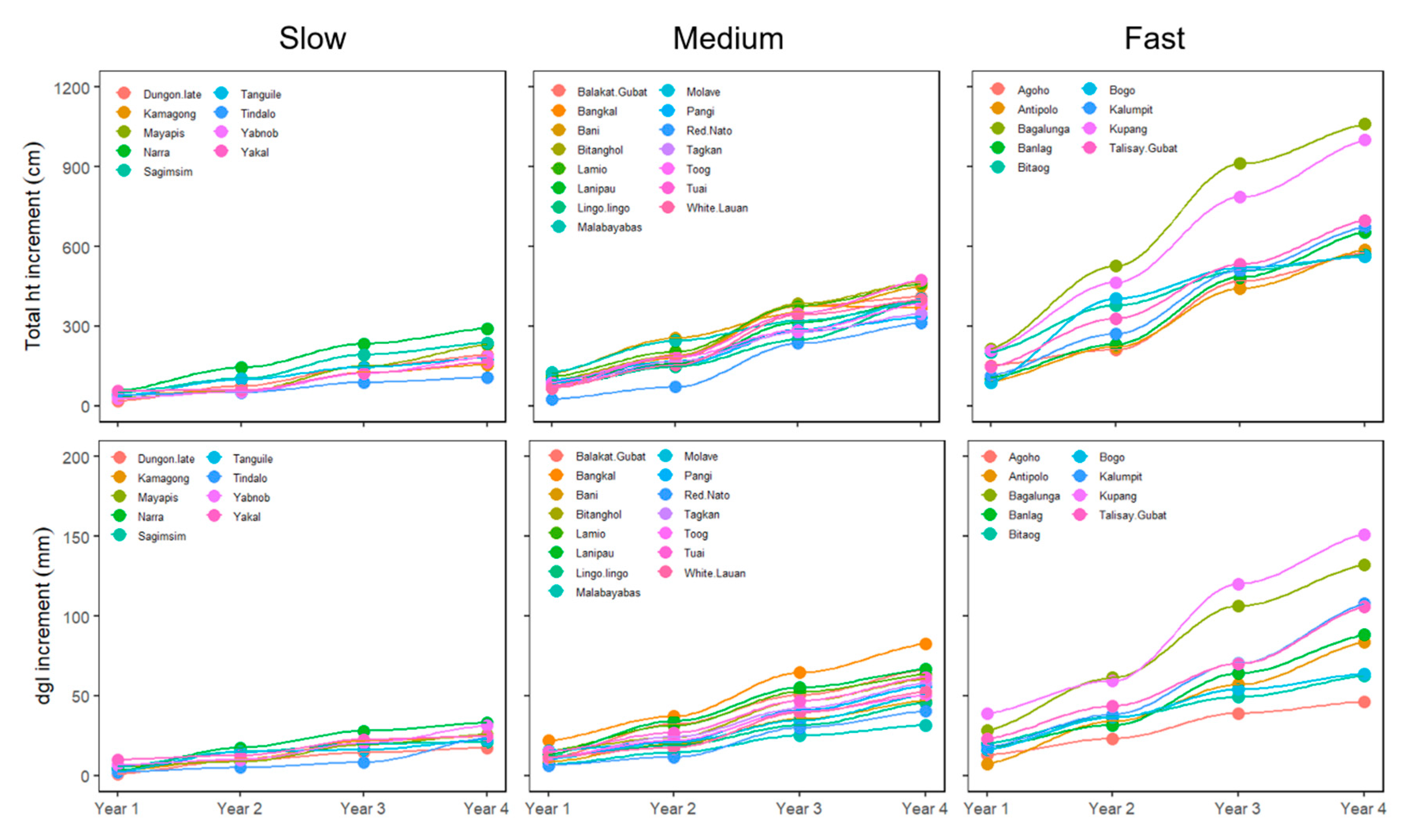
3.2. Growth Performances of Domesticated Species Per Cluster
3.3. Variations in Growth Performances of Clustered Domesticated Species
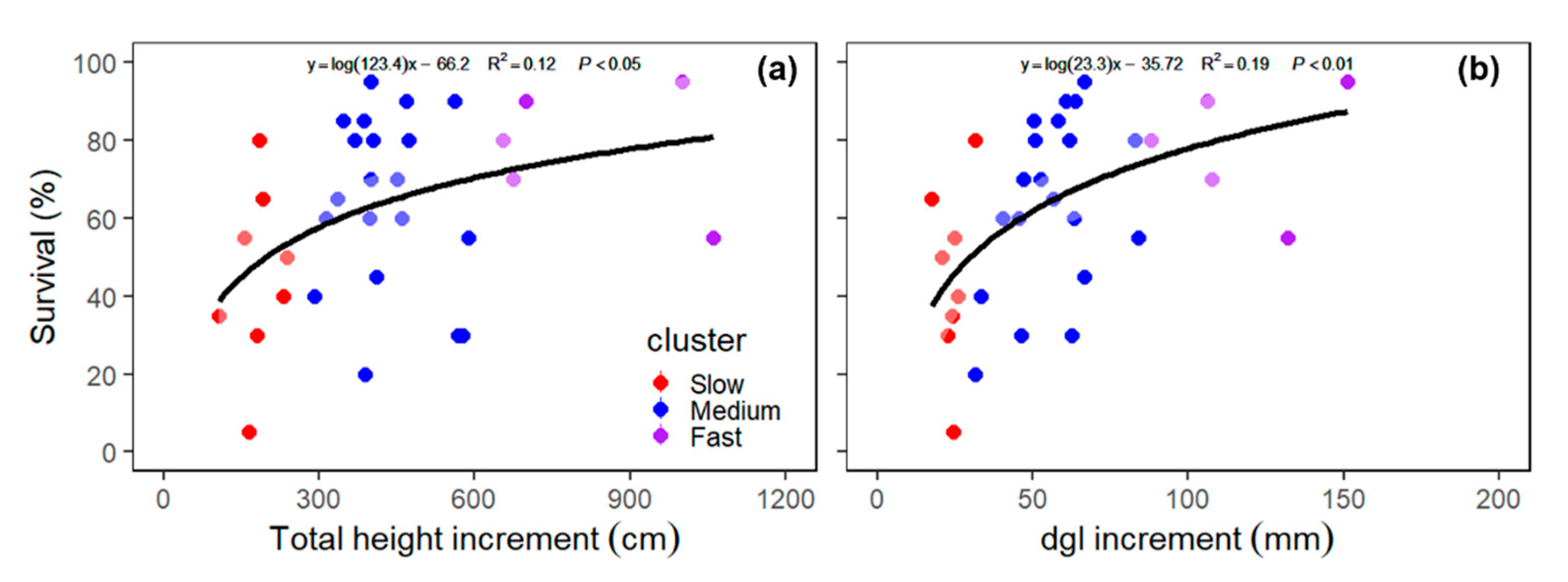
3.4. Sources of Variations in Survival Rate of Clustered Species
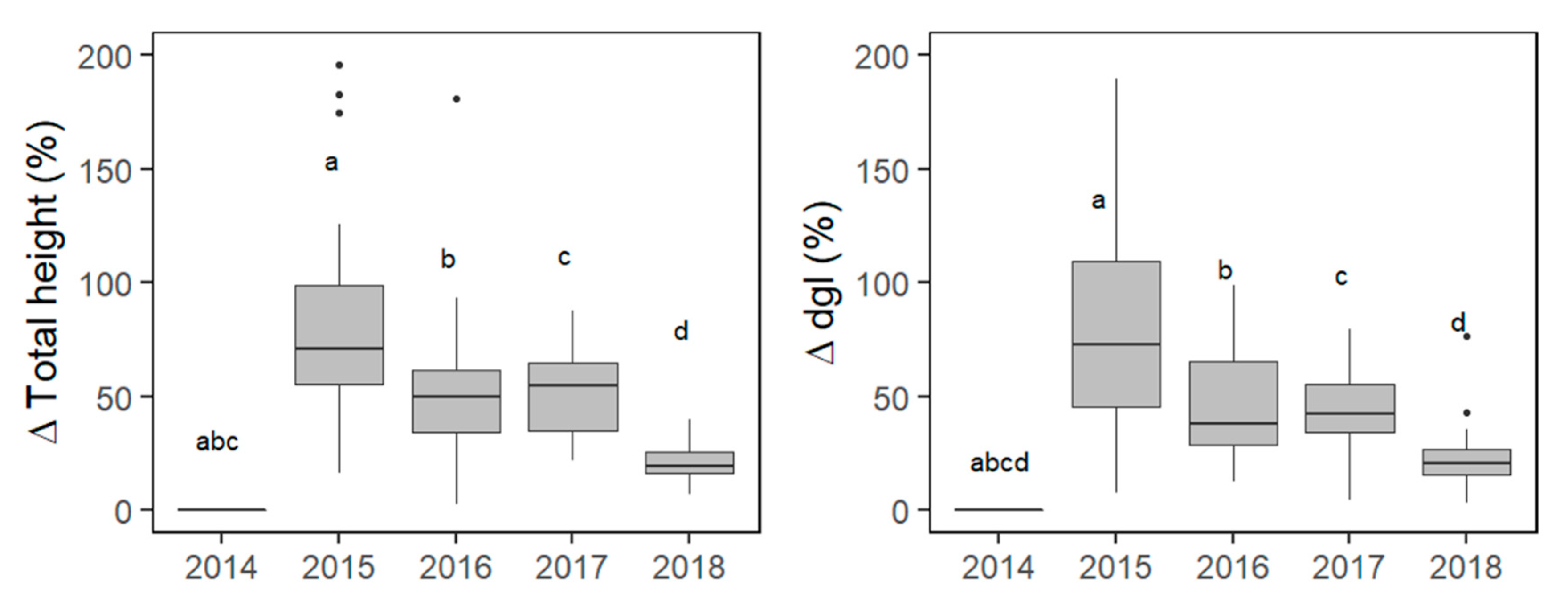
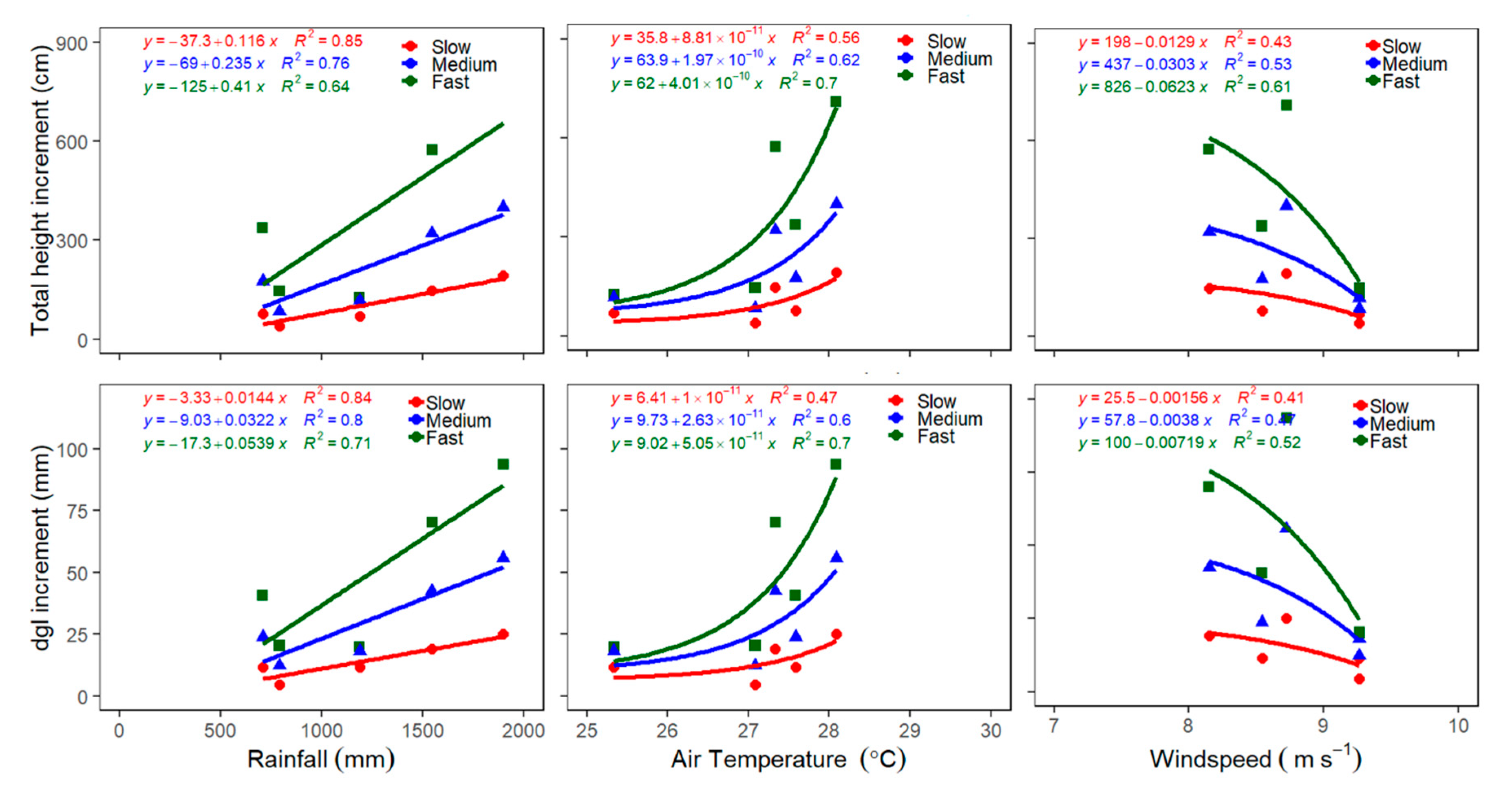
3.5. Climatic Effects on Total Height and Dgl Increments of Clustered species
4. Discussion
4.1. Survival Tendencies of Native Species
4.2. Early Stage of Development of Native Species
4.3. Factors Affecting Survival, Total Height And Dgl Growth
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leakey, R.R.B.; Newton, A.C. Tropical Trees: The Potential for Domestication and the Rebuilding of Forest Resources; HMSO Publications: London, UK, 1994; ISBN 0-11-701562-8. [Google Scholar]
- Nichols, J.D.; Vanclay, J.K. Domestication of native tree species for timber plantations: Key insights for tropical island nations. Int. For. Rev. 2012, 14, 402–413. [Google Scholar] [CrossRef] [Green Version]
- Lamb, D.; Erskine, P.D.; Parrota, J.A. Restoration of Degraded Tropical Forest Landscapes. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jactel, H.; Nicoll, B.C.; Branco, M.; Gonzalez-Olabarria, J.R.; Grodzki, W.; Långström, B.; Moreira, F.; Netherer, S.; Orazio, C.; Piou, D.; et al. The influences of forest stand management on biotic and abiotic risks of damage. Ann. For. Sci. 2009, 66, 701. [Google Scholar] [CrossRef]
- Butterfield, R.P. Early species selection for tropical reforestation: A consideration of stability. For. Ecol. Manag. 1996, 81, 161–168. [Google Scholar] [CrossRef]
- Calvo-Alvarado, J.C.; Arias, D.; Richter, D.D. Early growth performance of native and introduced fast growing tree species in wet to sub-humid climates of the Southern region of Costa Rica. For. Ecol. Manag. 2007, 242, 227–235. [Google Scholar] [CrossRef]
- Philippine Forestry Statistics 2018; Department of Environment and Natural Resources, Forest Management Bureau: Quezon City, Philippines, 2018.
- DENR Annual Report, 2017; Department of Environment and Natural Resources: Quezon City, Philippines, 2017.
- Polinar, A.; City, B. Domesticated Trees of Upland Farmers in Southern Leyte. J. Nat. Stud. 2017, 8, 69–76. [Google Scholar]
- Luna, M.P.G. Impact Assessment of the National Greening Program. of the DENR: Scoping or Process. Evaluation Phase—Institutional Component; Philippine Institute for Development Studies: Quezon City, Philippines, 2016. [Google Scholar]
- Wishnie, M.H.; Dent, D.H.; Mariscal, E.; Deago, J.; Cedeño, N.; Ibarra, D.; Condit, R.; Ashton, P.M.S. Initial performance and reforestation potential of 24 tropical tree species planted across a precipitation gradient in the Republic of Panama. For. Ecol. Manag. 2007, 243, 39–49. [Google Scholar] [CrossRef]
- Potvin, C.; Dutilleul, P. Neighborhood effects and size-assymtric competition in a tree plantation varying in density. Ecology 2009, 90, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Piotto, D.; Víquez, E.; Montagnini, F.; Kanninen, M. Pure and mixed forest plantations with native species of the dry tropics of Costa Rica: A comparison of growth and productivity. For. Ecol. Manag. 2004, 190, 359–372. [Google Scholar] [CrossRef]
- Van Breugel, M.; Hall, J.S.; Craven, D.J.; Gregoire, T.G.; Park, A.; Dent, D.H.; Wishnie, M.H.; Mariscal, E.; Deago, J.; Ibarra, D.; et al. Early growth and survival of 49 tropical tree species across sites differing in soil fertility and rainfall in Panama. For. Ecol. Manag. 2011, 261, 1580–1589. [Google Scholar] [CrossRef]
- Otsamo, R. Effect of nurse tree species on early growth of Anisoptera marginata Korth. (Dipterocarpaceae) on an Imperata cylindrica (L.) Beauv. grassland site in South Kalimantan, Indonesia. For. Ecol. Manag. 1998, 105, 303–311. [Google Scholar] [CrossRef]
- Davis, A.S.; Jacobs, D.F. Quantifying root system quality of nursery seedlings and relationship to outplanting performance. New For. 2005, 30, 295–311. [Google Scholar] [CrossRef]
- Burdeos, K.B.; Lansigan, F.P. Statistical Downscaling of Future Precipitation Scenarios for Agusan del Norte. Clim. Disater Dev. J. Philipp. 2017, 2, 13–22. [Google Scholar] [CrossRef]
- Climate Change in the Philippines; Department of Science and Technology–Philippine Atmospheric Geophysical and Astronomical Services Administration. y and Agrometeorology Division (CAD): Diliman City, Philippines, 2011.
- Collado, W.B.; Obico, M.R. Result of the Soil Analysis on the Physical and Chemical Characteristics of Butuan Series; PhilRice AES: Agusan del Norte, Philippines, 2007. [Google Scholar]
- Rojo, J.P. Revised Lexicon of Philippine Trees; Forest Products Research and Development Inst., Dep. of Science and Technology: Laguna, Philippines, 1999. [Google Scholar]
- Salvosa, F. A Forestry Lexicon of Philippine Trees Author, 1960th ed.; Forest Products Research Institute: Laguna, Philippines, 1960. [Google Scholar]
- Nussbaum, R.; Anderson, J.; Spencer, T. Factors limiting the growth of indigenous tree seedlings planted on degraded rainforest soils in Sabah, Malaysia. For. Ecol. Manag. 1995, 74, 149–159. [Google Scholar] [CrossRef]
- Schneider, T.; Ashton, M.S.; Montagnini, F.; Milan, P.P. Growth performance of sixty tree species in smallholder reforestation trials on Leyte, Philippines. New For. 2014, 45, 83–96. [Google Scholar] [CrossRef]
- Tolentino, E.L. Restoration of Philippine Native Forest by Smallholder Tree Farms; Snelder, D.J., Lasco, R.D., Eds.; Springer: New York, NY, USA, 2008; pp. 319–346. [Google Scholar]
- Redondo-Brenes, A.; Montagnini, F. Growth, productivity, aboveground biomass, and carbon sequestration of pure and mixed native tree plantations in the Caribbean lowlands of Costa Rica. For. Ecol. Manag. 2006, 232, 168–178. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Importance of root growth in overcoming planting stress. New For. 2005, 30, 273–294. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Salifu, K.F.; Seifert, J.R. Relative contribution of initial root and shoot morphology in predicting field performance of hardwood seedlings. New For. 2005, 30, 235–251. [Google Scholar] [CrossRef]
- Kriswanati, H.; Kallio, M.; Kanninen, M. Acacia Mangium Willd: Ecology, Silviculture and Productivity; CIFOR: Bogor, Indonesia, 2011; ISBN1 6028693375. ISBN2 9786028693370. [Google Scholar]
- Tasi, L.M. Growth and Yield (Chapter 8). In Acacia mangium-Growing and Utilization; MPTS Monograph Series No. 3; Winrock International and FAO: Bangkok, Thailand, 1993. [Google Scholar]
- Thompson, J.R.; Schultz, R.C. Root system morphology of Quercus rubra L. planting stock and 3-year field performance in Iowa. New For. 1995, 9, 225–236. [Google Scholar] [CrossRef]
- Bouma, T.J.; Nielsen, K.L.; Koutstaal, B. Sample preparation and scanning protocol for computerised analysis of root length and diameter. Plant Soil 2000, 218, 185–196. [Google Scholar] [CrossRef]
- Costa, C.; Dwyer, L.M.; Hamel, C.; Muamba, D.F.; Wang, X.L.; Nantais, L.; Smith, D.L. Root contrast enhancement for measurement with optical scanner-based image analysis. Can. J. Bot. 2001, 79, 23–29. [Google Scholar] [CrossRef]
- Aguilos, M.; Hérault, B.; Burban, B.; Wagner, F.; Bonal, D. What drives long-term variations in carbon flux and balance in a tropical rainforest in French Guiana? Agric. For. Meteorol. 2018. [Google Scholar] [CrossRef]
- Aguilos, M.; Stahl, C.; Burban, B.; Hérault, B.; Courtois, E.; Coste, S.; Wagner, F.; Ziegler, C.; Takagi, K.; Bonal, D. Interannual and seasonal variations in ecosystem transpiration and water use efficiency in a tropical rainforest. Forests 2019, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Agyeman, V.K.; Swaine, M.D.; Thompson, J. Responses of tropical forest tree seedlings to irradiance and the derivation of a light response index. J. Ecol. 1999, 87, 815–827. [Google Scholar] [CrossRef]
- Mushove, P.T.; Prior, J.A.B.; Gumbie, C.; Cutler, D.F. The effects of different environments on diameter growth increments of Colophospermum mopane and Combretum apiculatum. For. Ecol. Manag. 1995, 72, 287–292. [Google Scholar] [CrossRef]
- Brown, N. The implications of climate and gap microclimate for seedling growth conditions in a Bornean lowland rain forest. J. Trop. Ecol. 1993, 9, 153–168. [Google Scholar] [CrossRef]
- Woods, C.J.; Coutts, M.P.; Grace, J. Wind and Trees: Understanding Wind Forces on Trees; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Telewski, F.W.; Coutts, M.P.; Grace, J. Wind and Wind Related Damage to Trees: Wind-Induced Physiological Responses; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Coutts, M.P.; Nielsen, C.C.N.; Nicoll, B.C. The development of symmetry, rigidity and anchorage in the structural root system of conifers. Plant Soil 1999, 217, 1–15. [Google Scholar] [CrossRef]
- Tamasi, E.; Stokes, A.; Lasserre, B.; Danjon, F.; Berthier, S.; Fourcaud, T. Influence of wind loading on root system development and architecture in oak (Quercus robur L.) seedlings. Trees 2005, 19, 374–384. [Google Scholar] [CrossRef]
- Nicoll, B.C.; Gardiner, B.A.; Rayner, B.; Peace, A.J. Anchorage of coniferous trees in relation to species, soil type, and rooting depth. Can. J. For. Res. 2006, 36, 1871–1883. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Code | Common Name | Family Name | Scientific Name |
|---|---|---|---|
| 1 | Agoho | Casuarinaceae | Casuarina equisetifolia L. |
| 2 | Antipolo | Moraceae | Artocarpus blancoi (Elmer) Merr |
| 3 | Bagalunga | Meliaceae | Melia dubia L. |
| 4 | Balakat Gubat | Euphorbiaceae | Sapium luzonicum (Vidal) |
| 5 | Bani | Fabaceae | Pongamia pinnata (L) Merr |
| 6 | Banlag | Annonaceae | Xylopia ferruginea (Hook. F & Thoms) |
| 7 | Bitanghol | Clusiaceae | Calophyllum blancoi (Pl & Tr.) |
| 8 | Bitaog | Clusiaceae | Calophyllum inophyllum L. |
| 9 | Bogo | Burseraceae | Garuga floribunda (Decne) |
| 10 | Dungon-late | Malvaceae | Heritiera littoralis Ait. |
| 11 | Katoan bangkal | Rubiaceae | Antocephalus chinensis (Lamk) |
| 12 | Kalumpit | Combretaceae | Terminalia microcarpa (Decne) |
| 13 | Kamagong | Ebenaceae | Diospyros blancoi (A. DC) |
| 14 | Kupang | Mimosaceae | Parkia javanica (D.C) Merr |
| 15 | Lamio | Anacardiaceae | Dracontomelon edule (Blanco) Skeels |
| 16 | Lanipau | Combretaceae | Terminalia copelandii Elmer |
| 17 | Lingo-lingo | Lamiaceae | Viticipremna philippinensis Merr. |
| 18 | Malabayabas | Myrtaceae | Tristaniopsis decorticata (Merr) |
| 19 | Mayapis | Dipterocarpaceae | Shorea squamata (Blanco) Merr |
| 20 | Molave | Lamiaceae | Vitex parviflora Juss. |
| 21 | Narra | Fabaceae | Pterocarpus indicus Wild |
| 22 | Pangi | Achariaceae | Pangium edule Reinw. |
| 23 | Red Nato | Sapotaceae | Pouteria macrantha (Merr.) |
| 24 | Sagimsim | Myrtaceae | Syzygium brevistylum (C.B. Rob) Merr. |
| 25 | Tagkan | Sapotaceae | Palaquium pinnatinervium Elmer |
| 26 | Talisay Gubat | Combretaceae | Terminalia foetidissima Griff. |
| 27 | Tanguile | Dipterocarpaceae | Shorea polysperma (Blanco) Merr. |
| 28 | Tindalo | Fabaceae | Afzelia rhomboidea (Blanco) |
| 29 | Toog | Lecythidaceae | Petersianthus quadrialatus (Merr.) Merr. |
| 30 | Tuai | Phyllanthaceae | Bischofia javanica Blume |
| 31 | White Lauan | Dipterocarpaceae | Shorea contorta Vidal |
| 32 | Yabnob | Myristicaceae | Horsfieldia megacarpa (Miq.) Warb |
| 33 | Yakal | Dipterocarpaceae | Shorea astylosa Foxw. |
| Cluster | Scientific Name | Height Increment (cm) | Diameter at Ground Level Increment (mm) | % Survival | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Year 1 | Year 2 | Year 3 | Year 4 | Year 1 | Year 2 | Year 3 | Year 4 | (Age 5 Years) | ||
| Fast | Casuarina equisetifolia | 152 | 211 | 470 | 577 | 13 | 23 | 39 | 46 | 30 |
| Fast | Artocarpus blancoi | 92 | 222 | 442 | 588 | 7 | 34 | 57 | 84 | 55 |
| Fast | Melia dubia | 214 | 527 | 912 | 1061 | 28 | 61 | 106 | 132 | 55 |
| Fast | Xylopia ferruginea | 106 | 231 | 485 | 655 | 18 | 32 | 64 | 88 | 80 |
| Fast | Calophyllum inophyllum | 202 | 378 | 509 | 569 | 20 | 37 | 49 | 63 | 30 |
| Fast | Garuga floribunda | 87 | 404 | 520 | 562 | 16 | 37 | 54 | 64 | 90 |
| Fast | Terminalia microcarpa | 114 | 273 | 510 | 675 | 18 | 38 | 70 | 108 | 70 |
| Fast | Parkia javanica | 208 | 465 | 786 | 1001 | 39 | 60 | 120 | 151 | 95 |
| Fast | Terminalia foetidissima | 147 | 330 | 533 | 699 | 23 | 44 | 70 | 106 | 90 |
| Medium | Sapium luzonicum | 76 | 190 | 376 | 411 | 10 | 32 | 51 | 67 | 45 |
| Medium | Antocephalus chinensis | 71 | 180 | 377 | 369 | 22 | 37 | 65 | 83 | 80 |
| Medium | Pongamia pinnata | 123 | 256 | 350 | 450 | 8 | 20 | 36 | 47 | 70 |
| Medium | Calophyllum blancoi | 97 | 187 | 384 | 468 | 15 | 24 | 47 | 61 | 90 |
| Medium | Dracontomelon edule | 110 | 204 | 377 | 460 | 15 | 31 | 52 | 63 | 60 |
| Medium | Terminalia copelandii | 94 | 158 | 315 | 399 | 13 | 34 | 55 | 67 | 95 |
| Medium | Viticipremna philippinensis | 69 | 148 | 249 | 399 | 12 | 19 | 32 | 46 | 60 |
| Medium | Tristaniopsis decorticata | 126 | 245 | 320 | 389 | 7 | 14 | 25 | 32 | 20 |
| Medium | Vitex parviflora | 96 | 168 | 286 | 403 | 16 | 21 | 35 | 51 | 80 |
| Medium | Pangium edule | 85 | 154 | 280 | 335 | 10 | 21 | 41 | 57 | 65 |
| Medium | Pouteria macrantha | 25 | 71 | 235 | 312 | 6 | 12 | 30 | 40 | 60 |
| Medium | Palaquium pinnatinervium | 67 | 167 | 278 | 347 | 11 | 24 | 42 | 58 | 85 |
| Medium | Petersianthus quadrialatus | 89 | 184 | 285 | 386 | 12 | 22 | 40 | 50 | 85 |
| Medium | Bischofia javanica | 72 | 185 | 349 | 474 | 15 | 27 | 47 | 62 | 80 |
| Medium | Shorea contorta | 67 | 153 | 343 | 400 | 12 | 18 | 40 | 53 | 70 |
| Slow | Heritiera littoralis | 17 | 75 | 149 | 191 | 1 | 9 | 14 | 17 | 65 |
| Slow | Diospyros blancoi | 40 | 58 | 126 | 156 | 3 | 10 | 22 | 25 | 55 |
| Slow | Shorea squamata | 29 | 55 | 150 | 231 | 6 | 9 | 19 | 26 | 40 |
| Slow | Pterocarpus indicus | 56 | 144 | 234 | 292 | 3 | 18 | 28 | 33 | 40 |
| Slow | Syzygium brevistylum | 49 | 103 | 193 | 238 | 6 | 15 | 20 | 21 | 50 |
| Slow | Shorea polysperma | 35 | 100 | 146 | 181 | 2 | 15 | 16 | 23 | 30 |
| Slow | Afzelia rhomboidea | 41 | 51 | 89 | 107 | 2 | 5 | 8 | 24 | 35 |
| Slow | Horsfieldia megacarpa | 27 | 53 | 122 | 185 | 7 | 10 | 20 | 32 | 80 |
| Slow | Shorea astylosa | 57 | 59 | 124 | 164 | 10 | 13 | 23 | 25 | 5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilos, R.; Marquez, C.; Adornado, H.; Aguilos, M. Domesticating Commercially Important Native Tree Species in the Philippines: Early Growth Performance Level. Forests 2020, 11, 885. https://doi.org/10.3390/f11080885
Aguilos R, Marquez C, Adornado H, Aguilos M. Domesticating Commercially Important Native Tree Species in the Philippines: Early Growth Performance Level. Forests. 2020; 11(8):885. https://doi.org/10.3390/f11080885
Chicago/Turabian StyleAguilos, Rustum, Conrado Marquez, Henry Adornado, and Maricar Aguilos. 2020. "Domesticating Commercially Important Native Tree Species in the Philippines: Early Growth Performance Level" Forests 11, no. 8: 885. https://doi.org/10.3390/f11080885