Effect of Temperature, Light, and Storage Time on the Seed Germination of Pinus bungeana Zucc. ex Endl.: The Role of Seed-Covering Layers and Abscisic Acid Changes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Materials
2.2. The Effect of Temperature and Light on Seed Germination
2.3. The Effect of Dry Cold Storage on Germination
2.4. Germination Recovery Experiments
2.5. Embryo Coverings Contributed to High-Temperature Inhibition
2.6. Changes in Structure and ABA Content of Nucellar Membrane During Germination at Different Temperature
2.7. Statistical Analyses
3. Results
3.1. Temperature and Light Effect on Seed Germination
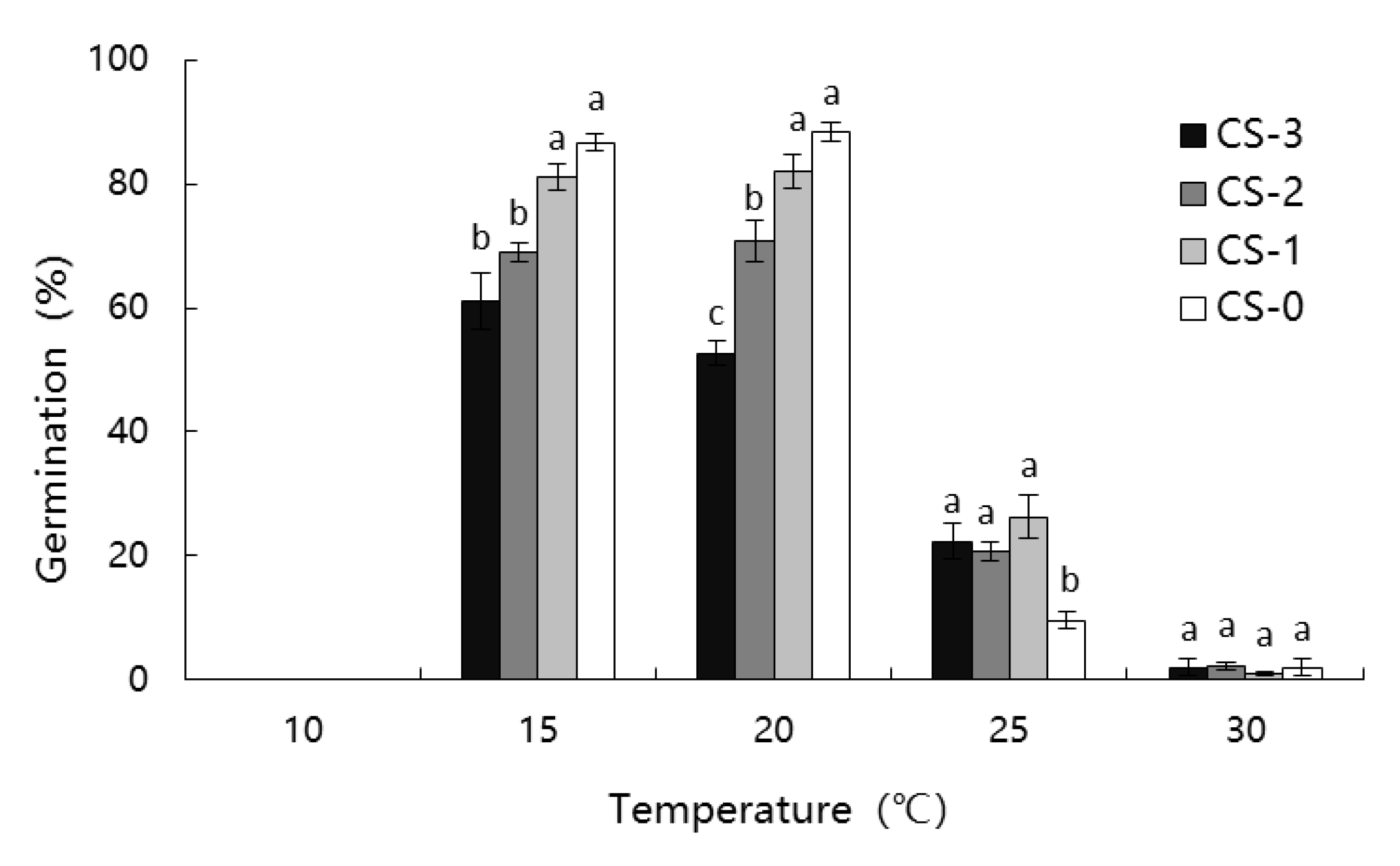
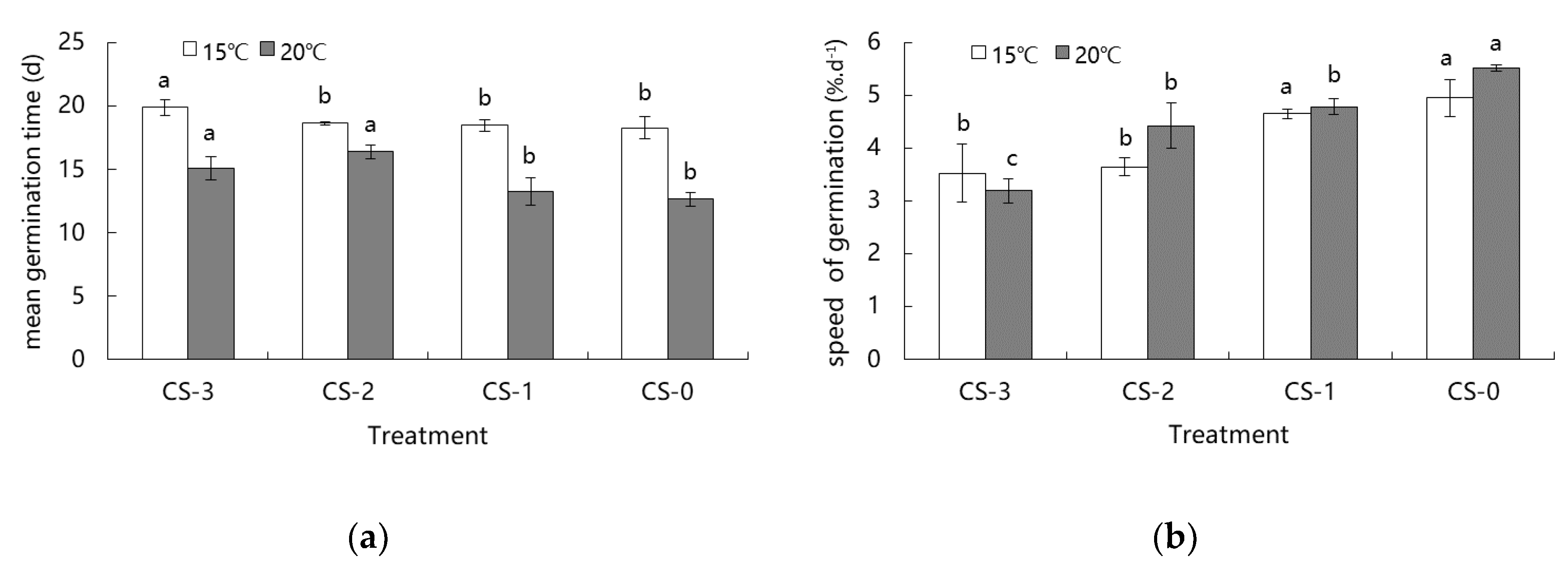
3.2. Effect of Dry Cold Storage on Germination
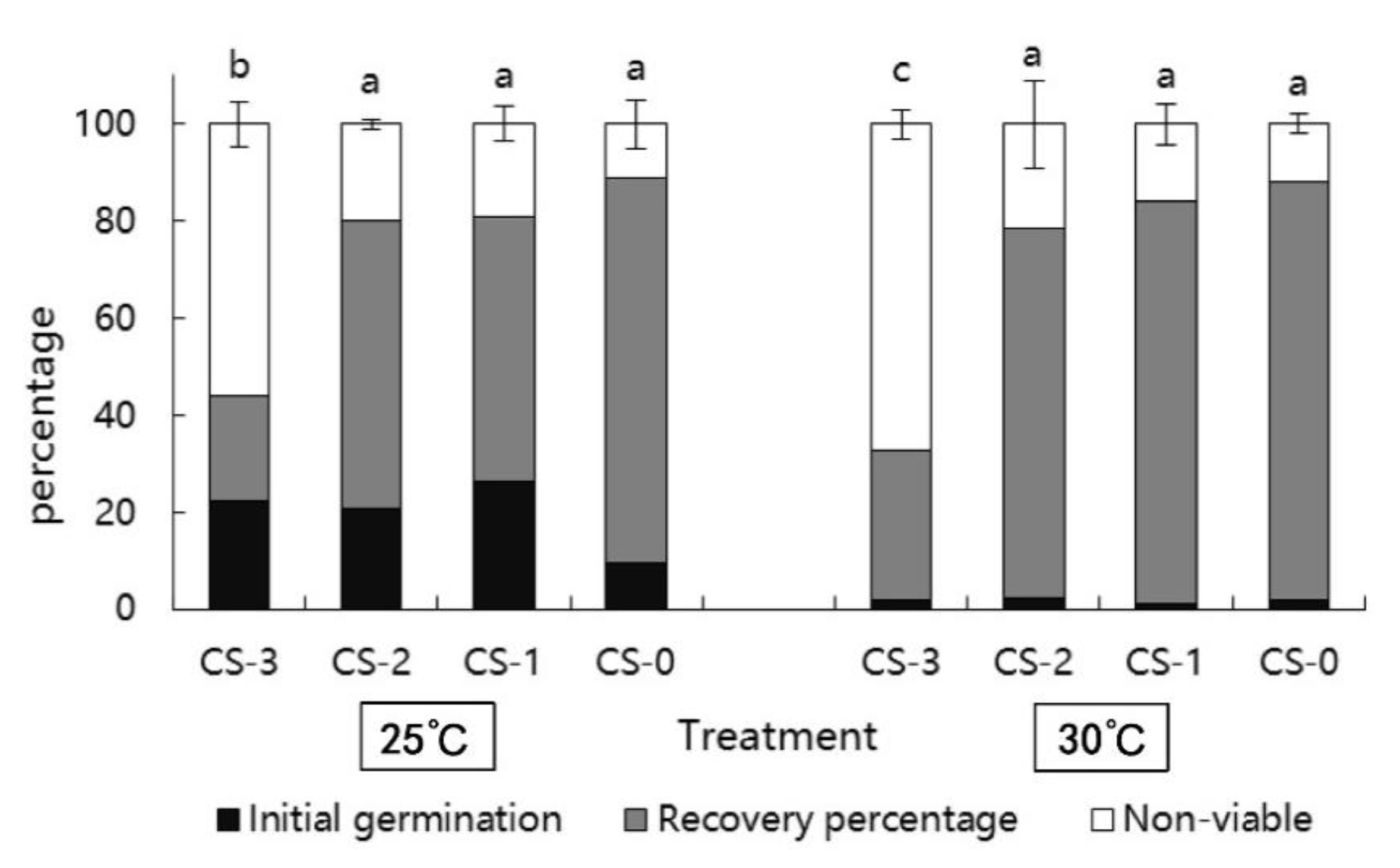
3.3. Seed Recovery Responses
3.4. Embryo Coverings Contributed to High-Temperature Inhibition
3.5. Changes in Morphology of Nucellar Membrane During Germination at Different Temperature
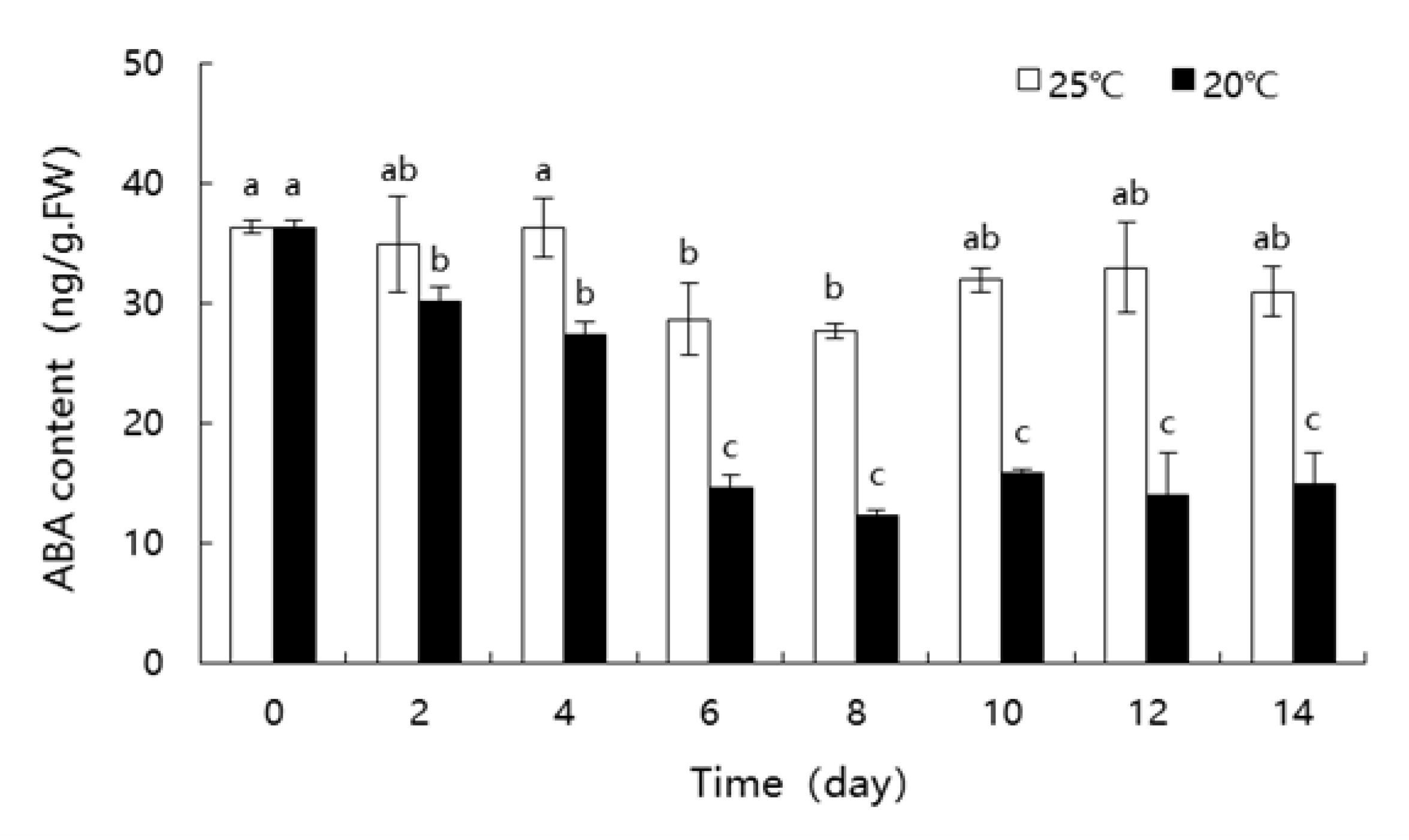
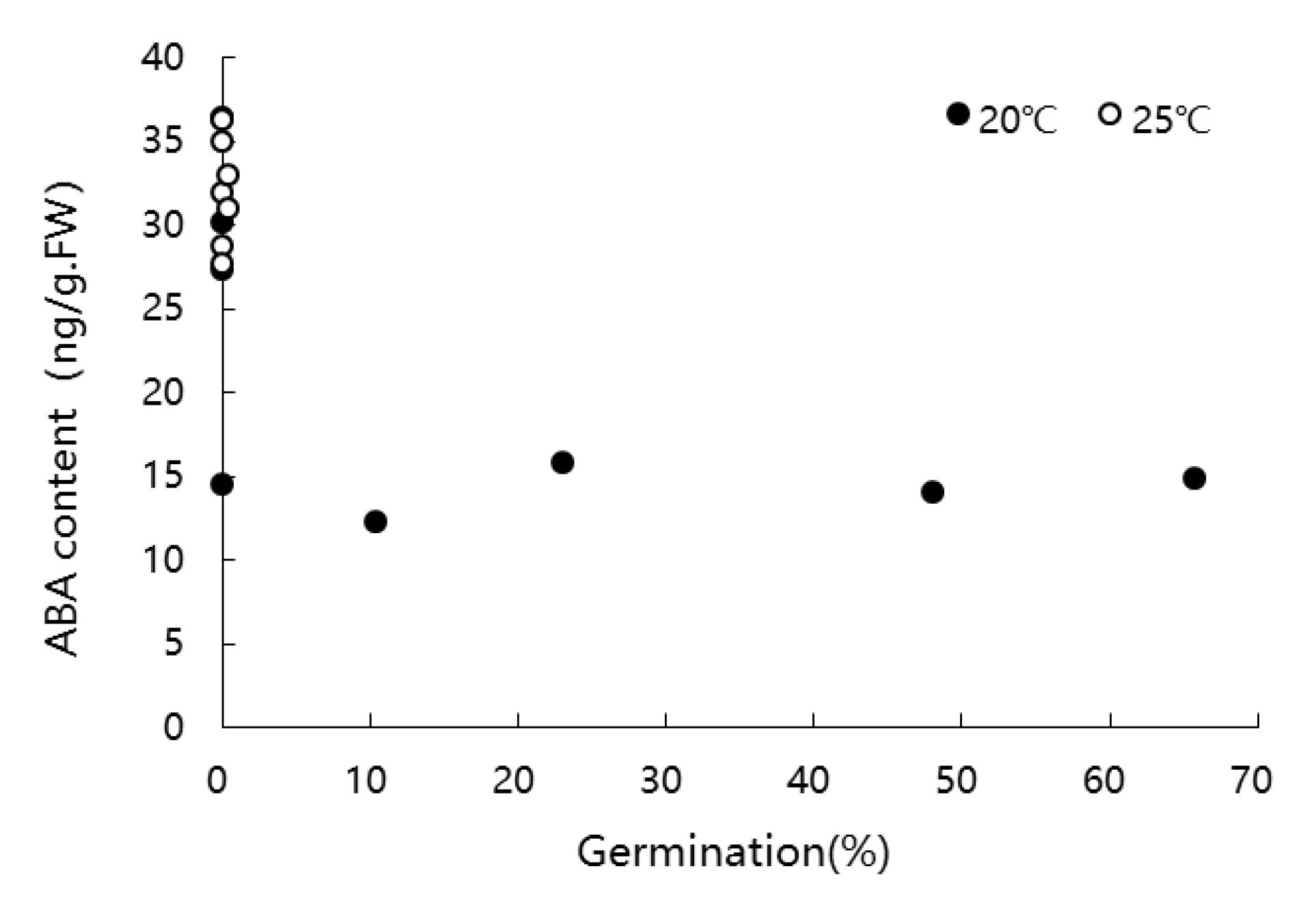
3.6. Changes in ABA Content of Nucellar Membrane during Germination at Different Temperature
4. Discussion
4.1. Seed Germination Responses
4.2. Germination Requirements after Storage Treatments
4.3. The Reversibility of Germination Inhibition from High Temperatures
4.4. Seed Tissues Involved in Inhibited Germination at High Temperatures
4.5. The Role of ABA Played in Germination at High Temperatures
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bewley, J.D.; Bradford, K.; Hilhorst, H.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Plenum Press: New York, NY, USA, 2014. [Google Scholar]
- Marcos Filho, J.M.F. Fisiologia de Sementes de Plantas Cultivadas; Abrates: Londrina, Brazil, 2015. [Google Scholar]
- Cristaudo, A.; Gresta, F.; Restuccia, A.; Catara, S.; Onofri, A. Germinative response of redroot pigweed (Amaranthus retroflexus L.) to environmental conditions: Is there a seasonal pattern? Plant Biosyst. Int. J. Dealing Asp. Plant Biol. 2016, 150, 583–591. [Google Scholar]
- Tolyat, M.A.; Afshari, R.T.; Jahansoz, M.R.; Nadjafi, F.; Naghdibadi, H.A. Determination of cardinal germination temperatures of two ecotypes of Thymus daenensis subsp. daenensis. Seed Sci. Technol. 2014, 42, 28–35. [Google Scholar] [CrossRef]
- Fallahi, H.-R.; Mohammadi, M.; Aghhavani-Shajari, M.; Ranjbar, F. Determination of germination cardinal temperatures in two basil (Ocimum basilicum L.) cultivars using non-linear regression models. J. Appl. Res. Med. Aromat. Plants 2015, 2, 140–145. [Google Scholar] [CrossRef]
- Laghmouchi, Y.; Belmehdi, O.; Bouyahya, A.; Skali Senhaji, N.; Abrini, J. Effect of temperature, salt stress and PH on seed germination of medicinal plant Origanum compactum. Biocatal. Agric. Biotechnol. 2017, 10, 156–160. [Google Scholar] [CrossRef]
- Gresta, F.; Cristaudo, A.; Onofri, A.; Restuccia, A.; Avola, G. Germination response of four pasture species to temperature, light, and post-harvest period. Plant Biosyst. 2010, 144, 849–856. [Google Scholar] [CrossRef]
- Geshnizjani, N.; Ghaderi-Far, F.; Willems, L.A.J.; Hilhorst, H.W.M.; Ligterink, W. Characterization of and genetic variation for tomato seed thermo-inhibition and thermo-dormancy. BMC Plant Biol. 2018, 18, 229. [Google Scholar] [CrossRef]
- Washitani, I.; Masuda, M. A comparative study of the germination characteristics of seeds from a moist tall grassland community. Func. Ecol. 1990, 4, 543. [Google Scholar] [CrossRef]
- Huo, H.; Dahal, P.; Kunusoth, K.; McCallum, C.; Bradford, K. Expression of 9-cis-EPOXYCAROTENOID DIOXYGENASE4 is essential for thermoinhibition of lettuce seed germination but not for seed development or stress tolerance. Plant Cell 2013, 25, 884–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristaudo, A.; Catara, S.; Mingo, A.; Restuccia, A.; Onofri, A. Temperature and storage time strongly affect the germination success of perennial Euphorbia species in Mediterranean regions. Ecol. Evol. 2019, 9, 10984–10999. [Google Scholar] [CrossRef] [Green Version]
- Taiz, L.; Zeiger, E.; Maffei, M. Fisiologia Vegetal; Artimed: Porto Alegre, Brazil, 2013. [Google Scholar]
- Aud, F.F.; Ferraz, I.D.K. Seed size influence on germination responses to light and temperature of seven pioneer tree species from the Central Amazon. Anais da Academia Brasileira de Ciências 2012, 84, 759–766. [Google Scholar] [CrossRef] [Green Version]
- Benvenuti, S.; Dinelli, G.; Bonetti, A. Germination ecology of Leptochloa chinensis: A new weed in the Italian rice agro-environment. Weed Res. Oxf. 2004, 44, 87–96. [Google Scholar] [CrossRef]
- Toh, S.; Imamura, A.; Watanabe, A.; Nakabayashi, K.; Okamoto, M.; Jikumaru, Y.; Hanada, A.; Aso, Y.; Ishiyama, K.; Tamura, N.; et al. High temperature-induced abscisic acid biosynthesis and its role in the inhibition of gibberellin action in Arabidopsis seeds. Plant Physiol. 2008, 146, 1368–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, R.; Nahal, H.; Provart, N.; Gazzarrini, S. The role of the Arabidopsis FUSCA3 transcription factor during inhibition of seed germination at high temperature. BMC Plant Biol. 2012, 12, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miransari, M.; Smith, D. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Chien, C.-T.; Chung, J.-D.; Yang, Y.-S.; Kuo, S.-R. Dormancy-break and germination in seeds of Prunus campanulata (Rosaceae): Role of covering layers and changes in concentration of abscisic acid and gibberellins. Seed Sci. Res. 2007, 17, 21–32. [Google Scholar] [CrossRef]
- Sawada, Y.; Aoki, M.; Nakaminami, K.; Mitsuhashi, W.; Tatematsu, K.; Kushiro, T.; Koshiba, T.; Kamiya, Y.; Inoue, Y.; Nambara, E.; et al. Phytochrome- and gibberellin-mediated regulation of abscisic acid metabolism during germination of photoblastic lettuce seeds. Plant Physiol. 2008, 146, 1386–1396. [Google Scholar] [CrossRef] [Green Version]
- Ali-Rachedi, S.; Bouinot, D.; Wagner, M.-H.; Bonnet, M.; Sotta, B.; Grappin, P.; Jullien, M. Changes in endogenous abscisic acid levels during dormancy release and maintenance of mature seeds: Studies with the Cape Verde Islands ecotype, the dormant model of Arabidopsis thaliana. Planta 2004, 219, 479–488. [Google Scholar] [CrossRef]
- Benech-Arnold, R.L.; Gualano, N.; Leymarie, J.; Côme, D.; Corbineau, F. Hypoxia interferes with ABA metabolism and increases ABA sensitivity in embryos of dormant barley grains. J. Exp. Bot. 2006, 57, 1423–1430. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Shao, C.; Zhang, Z. Characteristics of embryo dormancy and germination of Pinus bungeana. Sci. Silvae Sinicae 2003, 39, 47–54. [Google Scholar]
- Wang, X.; Liu, J.; Wang, J. Variation of dormancy characteristic of different Pinus bungeana seed sources. Chin. J. App. Ecol. 2000, 11, 9–12. [Google Scholar]
- Xin, Z. Seed dormancy mechanism of six pine species. J. Southwest For. Univ. 2008, 28, 5–9. [Google Scholar]
- Dong, L.; Xiao, B.; Cao, C. Investigating seed dormancy in Pinus bungeana. Shaanxi For. Sci. Technol. 1987, 1, 15–17. [Google Scholar]
- Wang, X.; Wang, J. Extraction, separation and biological measurement of endogenous growth regulating substance of seeds of Pinus bungeana. Seed 1998, 5, 19–22. [Google Scholar]
- Guo, C.; Shen, Y.; Shi, F. Investigating seed dormancy in Pinus bungeana Zucc. ex Endl.: Understanding the contributions of enclosing tissues and temperature on germination. Forests 2018, 9, 401. [Google Scholar] [CrossRef] [Green Version]
- Dangasuk, O.; Panetsos, K.P. Altitudinal and longitudinal variations in Pinus brutia (Ten.) of Crete Island, Greece: Some needle, cone and seed traits under natural habitats. New For. 2004, 27, 269–284. [Google Scholar] [CrossRef]
- Ma, Q.Y.; Chen, L.; Hou, J.; Liu, H.; Li, S. Seed viability tests for Acer pictum and A. Rubrum. Eur. J. Hortic. Sci. 2016, 81, 44–48. [Google Scholar] [CrossRef]
- ISTA. International Seed Testing Association; The International Seed Testing Association (ISTA): Bassersdorf, Switzerland, 2005. [Google Scholar]
- Zhao, J.; Li, G.; Yi, G.-X.; Wang, B.-M.; Deng, A.-X.; Nan, T.-G.; Li, Z.-H.; Li, Q.X. Comparison between conventional indirect competitive enzyme-linked immunosorbent assay (icELISA) and simplified icELISA for small molecules. Anal Chim Acta 2006, 571, 79–85. [Google Scholar] [CrossRef]
- Escudero, A.; Pérez-García, F.; Luzuriaga, A. Effects of light, temperature and population variability on the germination of seven Spanish pines. Seed Sci. Res. 2002, 12, 261–271. [Google Scholar] [CrossRef]
- Skordilis, A.; Thanos, C.A. Comparative Ecophysiology of Seed Germination Strategies in the Seven Pine Species Naturally Growing in Greece; Springer: Dordrecht, The Netherlands, 1997. [Google Scholar]
- Ganatsas, P.; Tsakaldimi, M. Effect of light conditions and salinity on germination behaviour and early growth of Umbrella pine (Pinus pinea L.) seed. J. Hortic. Sci. Biotechnol. 2007, 82, 605–610. [Google Scholar] [CrossRef]
- Feng, J.; Shen, Y.; Shi, F.; Li, C. Embryo development, seed germination, and the kind of dormancy of Ginkgo biloba L. Forests 2018, 9, 700. [Google Scholar] [CrossRef] [Green Version]
- Offord, C.; Meagher, P. Effects of temperature, light and stratification on seed germination of Wollemi pine (Wollemia nobilis, Araucariaceae). Aust. J. Bot. 2001, 49, 699–704. [Google Scholar] [CrossRef]
- Thanos, C.; Skordilis, A. The effects of light, temperature and osmotic stress on the germination of Pinus halepensis and P. Brutia seeds. Seed Sci. Technol. 1987, 15, 163–174. [Google Scholar]
- Chanyenga, T.F.; Geldenhuys, C.J.; Sileshi, G.W. Germination response and viability of an endangered tropical conifer Widdringtonia whytei seeds to temperature and light. South Afr. J. Bot. 2012, 81, 25–28. [Google Scholar] [CrossRef] [Green Version]
- Gutterman, Y.; Shem-Tov, S.; Gozlan, S. The effect of post-maturation temperatures and duration on seed germinability of Plantago coronopus occurring in natural populations in the Negev Desert highlands, Israel. J. Arid Environ. 1998, 38, 451–463. [Google Scholar] [CrossRef]
- El-Keblawy, A.; Al-Rawai, A. Effects of seed maturation time and dry storage on light and temperature requirements during germination in invasive Prosopis juliflora. Flora Morphol. Distrib. Funct. Ecol. Plants 2006, 201, 135–143. [Google Scholar] [CrossRef]
- Meyer, S.E.; Allen, P.S. Ecological genetics of seed germination regulation in Bromus tectorum L. Oecologia 1999, 120, 27–34. [Google Scholar] [CrossRef]
- Derkx, M.P.M.; Karssen, C.M. Variability in light-, gibberellin- and nitrate requirement of Arabidopsis thaliana seeds due to harvest time and conditions of dry storage. J. Plant Physiol. 1993, 141, 574–582. [Google Scholar] [CrossRef]
- Yilmaz, M.; Tonguc, F. Effects of temperature on the germination of Fraxinus ornus subsp. cilicica seeds. Dendrobiology 2013, 69, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Suszka, B.; Muller, C.; Bonnet-Masimbert, M. Seeds of Forest Broadleaves, from Harvest to Sowing; INRA: Paris, France, 1996. [Google Scholar]
- Piotto, B.; Di Noi, A. Seed Propagation of Mediterranean Trees and Shrubs; ANPA: Rome, Italy, 2003. [Google Scholar]
- Ribeiro, J.; Costa, C. The effect of temperature regulation on seed germination of the tropical tree Myrsine parvifolia A. Dc near its southern limit. South Afr. J. Bot. 2015, 98, 128–133. [Google Scholar] [CrossRef]
- Kabar, K.; Baltepe, S. Effect of kinetin and gibberellic acid in overcoming high temperature and salinity (NaCl) stresses on the germination of barley and lettuce seeds. Phyton 1990, 30, 65–74. [Google Scholar]
- Tillman-Sutela, E.; Kauppi, A. The morphological background to imbibition in seeds of Pinus sylvestris L. of different provenances. Trees 1995, 9, 123–133. [Google Scholar] [CrossRef]
- Bianco, J.; Garello, G.; Le Page-Degivry, M.T. De novo ABA synthesis and expression of seed dormancy in a gymnosperm: Pseudotsuga menziesii. Plant Growth Regul. 1997, 21, 115–119. [Google Scholar] [CrossRef]
- Hoff, R. Dormancy in Pinus monticola seed related to stratification time, seed coat, and genetics. Can. J. For. Res. 2011, 17, 294–298. [Google Scholar] [CrossRef]
- Zhao, T.; Qian, C.; Gao, Y.; Chen, L.; Zhu, M.; Pan, Y.; Li, S. Germination inhibitors detected in Sapium sebiferum seeds. J. For. Res. 2019, 30, 2305–2312. [Google Scholar] [CrossRef]
- Gonai, T.; Kawahara, S.; Tougou, M.; Satoh, S.; Hashiba, T.; Hirai, N.; Kawaide, H.; Kamiya, Y.; Yoshioka, T. Abscisic acid in the thermoinhibition of lettuce seed germination and enhancement of its catabolism by gibberellin. J. Exp. Bot. 2004, 55, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, N.; Abrams, S.; Kermode, A. Changes in ABA turnover and sensitivity that accompany dormancy termination of yellow-cedar (Chamaecyparis nootkatensis) seeds. J. Exp. Bot. 2002, 53, 89–101. [Google Scholar]
- Corbineau, F.; Bianco, J.; Garello, G.; Côme, D. Breakage of Pseudotsuga menziesii seed dormancy by cold treatment as related to changes in seed ABA sensitivity and ABA levels. Physiol. Plant 2002, 114, 313–319. [Google Scholar] [CrossRef]
- Taylor, N.; Hills, P.; Gold, J.; Stirk, W.; van Staden, J. Factors contributing to the regulation of thermoinhibition in Tagetes minuta L. J. Plant Physiol. 2005, 162, 1270–1279. [Google Scholar] [CrossRef]
- Chiu, R.; Saleh, Y.; Gazzarrini, S. Inhibition of FUSCA3 degradation at high temperature is dependent on ABA signaling and is regulated by the ABA/GA ratio. Plant Signal. Behav. 2016, 11, e1247137. [Google Scholar] [CrossRef] [Green Version]
- Barthe, P.; Garello, G.; Bianco-Trinchant, J.; le Page-Degivry, M.T. Oxygen availability and ABA metabolism in Fagus sylvatica seeds. Plant Growth Regul. 2000, 30, 185–191. [Google Scholar] [CrossRef]
- Viémont, J.D.; Crabbé, J. Dormancy in Plants: From Whole Plant Behaviour to Cellular Control; CABI Publishing: New York, NY, USA, 2000. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Germination (%) | Rot Rate (%) |
|---|---|---|
| Intact seeds | 5.33 ± 1.15 c | 10.67 ± 3.05 a |
| Seed coat cracked at the micropylar end | 11.33 ± 3.45 c | 14.00 ± 4.00 a |
| Seed coat removed (nucellar membrane intact) | 34.67 ± 5.03 b | 12.00 ± 2.00 a |
| Seed coat and nucellar membrane removed | 84.00 ± 2.00 a | 10.67 ± 3.05 a |
| Seed coat, nucellar membrane, and nucellar cap removed | 89.33 ± 3.05 a | 9.33 ± 2.00 a |
| Isolated embryos | 86.67 ± 1.52 a | 13.33 ± 1.52 a |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| TGP | |||||
| Light (L) | 8.533 | 1 | 8.533 | 1.446 | 0.243 |
| Temperature (T) | 54,750.867 | 4 | 13,687.717 | 2319.952 | 0.000 * |
| L × T | 9.800 | 4 | 2.450 | 0.415 | 0.796 |
| MGT | |||||
| Light (L) | 0.642 | 1 | 0.642 | 0.478 | 0.499 |
| Temperature (T) | 415.517 | 3 | 138.506 | 103.157 | 0.000 * |
| L × T | 2.443 | 3 | 0.814 | 0.607 | 0.620 |
| SG | |||||
| Light (L) | 0.009 | 1 | 0.009 | 0.342 | 0.567 |
| Temperature (T) | 173.010 | 3 | 57.670 | 2272.303 | 0.000 * |
| L × T | 0.175 | 3 | 0.058 | 2.304 | 0.116 |
| Temperature (°C) | TGP (%) | MGT (day) | SG (% day−1) | |||
|---|---|---|---|---|---|---|
| Dark | Light | Dark | Light | Dark | Light | |
| 10 | 0.00 ± 0.00 d | 0.00 ± 0.00 c | / | / | / | / |
| 15 | 84.00 ± 2.73 b | 87.33 ± 3.79 b | 17.20 ± 0.15 b | 17.61 ± 0.84 b | 5.11 ± 0.21 b | 5.23 ± 0.08 b |
| 20 | 91.00 ± 3.21 a | 92.67 ± 3.21 a | 13.85 ± 0.92 c | 13.45 ± 0.39 c | 5.95 ± 0.31 a | 5.62 ± 0.07 a |
| 25 | 4.33 ± 2.08 c | 5.00 ± 4.00 c | 23.39 ± 2.11 a | 22.07 ± 1.89 a | 0.18 ± 0.08 c | 0.24 ± 0.21 c |
| 30 | 0.67 ± 0.58 d | 1.00 ± 1.15 c | 24.00 ± 0.00 a | 24.00 ± 1.00 a | 0.04 ± 0.00 c | 0.04 ± 0.00 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, C.; Shen, Y.; Shi, F. Effect of Temperature, Light, and Storage Time on the Seed Germination of Pinus bungeana Zucc. ex Endl.: The Role of Seed-Covering Layers and Abscisic Acid Changes. Forests 2020, 11, 300. https://doi.org/10.3390/f11030300
Guo C, Shen Y, Shi F. Effect of Temperature, Light, and Storage Time on the Seed Germination of Pinus bungeana Zucc. ex Endl.: The Role of Seed-Covering Layers and Abscisic Acid Changes. Forests. 2020; 11(3):300. https://doi.org/10.3390/f11030300
Chicago/Turabian StyleGuo, Congcong, Yongbao Shen, and Fenghou Shi. 2020. "Effect of Temperature, Light, and Storage Time on the Seed Germination of Pinus bungeana Zucc. ex Endl.: The Role of Seed-Covering Layers and Abscisic Acid Changes" Forests 11, no. 3: 300. https://doi.org/10.3390/f11030300