
The Effect of Polyhydroxy Fullerene Derivative on Human Myeloid Leukemia K562 Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Experiment Procedures
2.1. Materials
2.2. Cell Proliferation Inhibition Assay
2.3. Cell Cycle Assay
2.4. Cell Morphology Assay
2.5. Detection of Apoptosis
2.6. Induction of Apoptosis and AO/EB Double Staining
2.7. Analysis of Mitochondrial Membrane Potential
2.8. Electron Microscopy of Cell Apoptosis
2.9. Data Analysis
3. Results
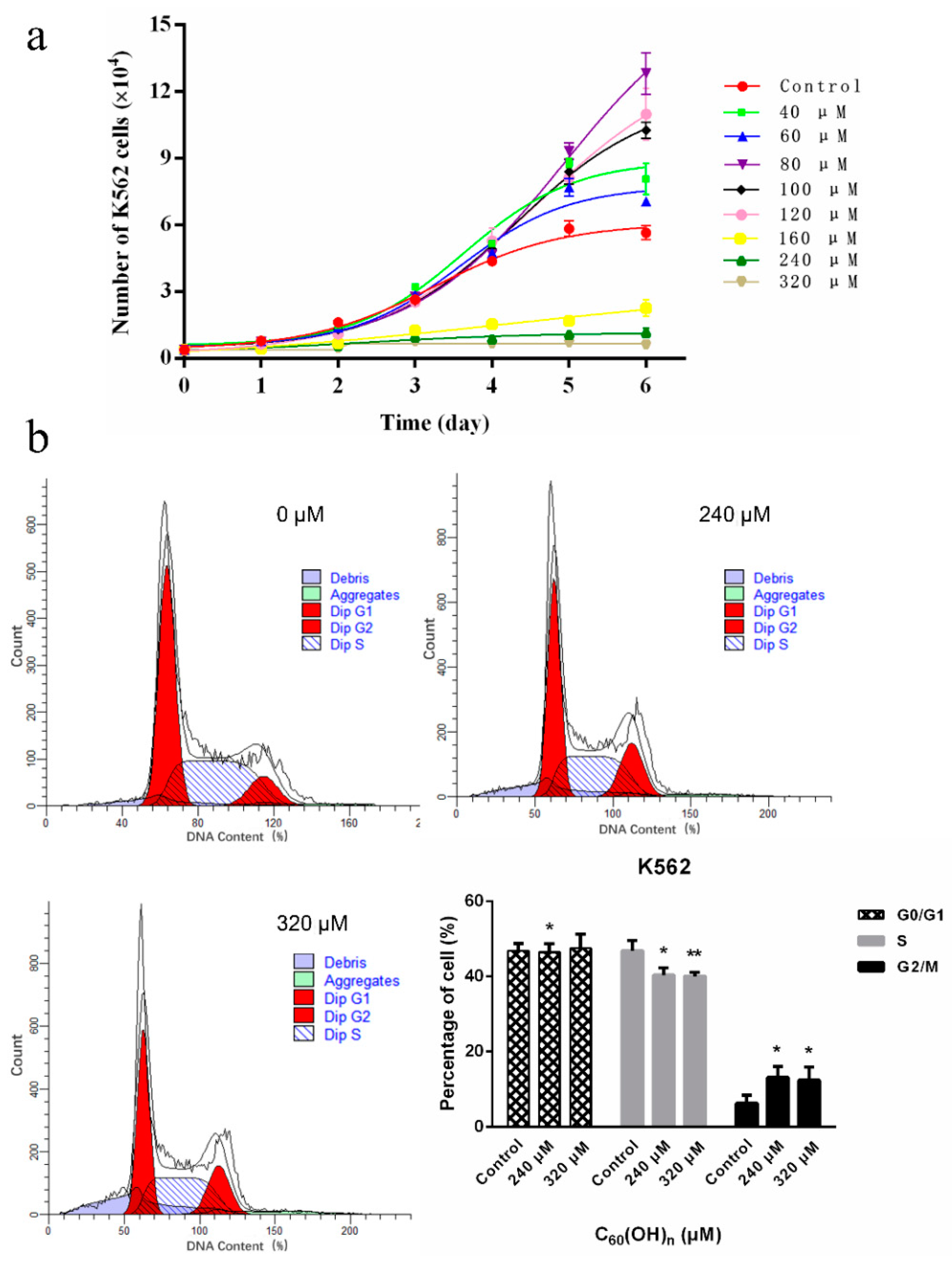
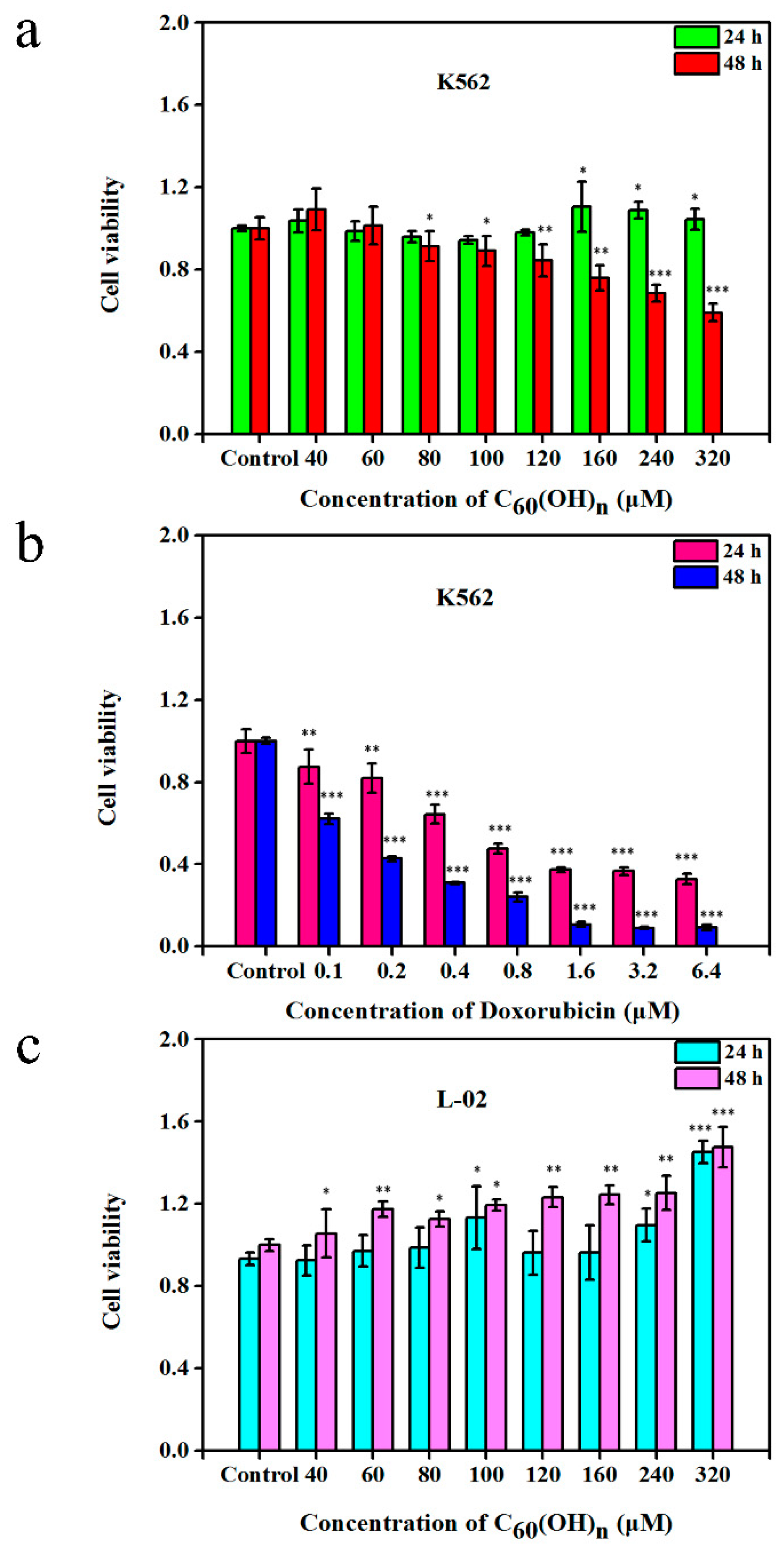
3.1. Effects of C60(OH)n on the Proliferation of K562 Cells
3.2. Effects of C60(OH)n on the Cell Cycle of K562
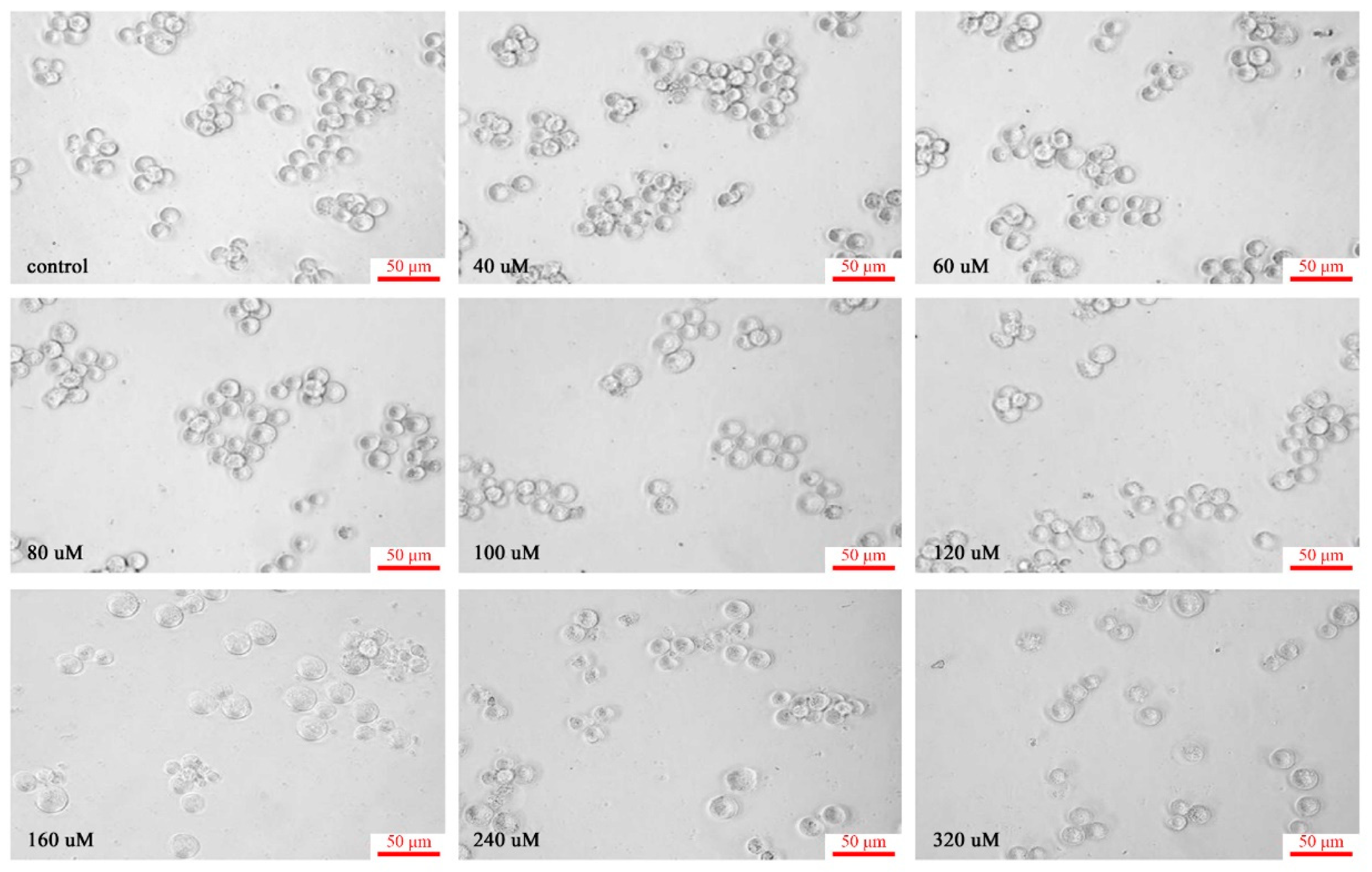
3.3. Morphological Study of C60(OH)n on K562 Cells
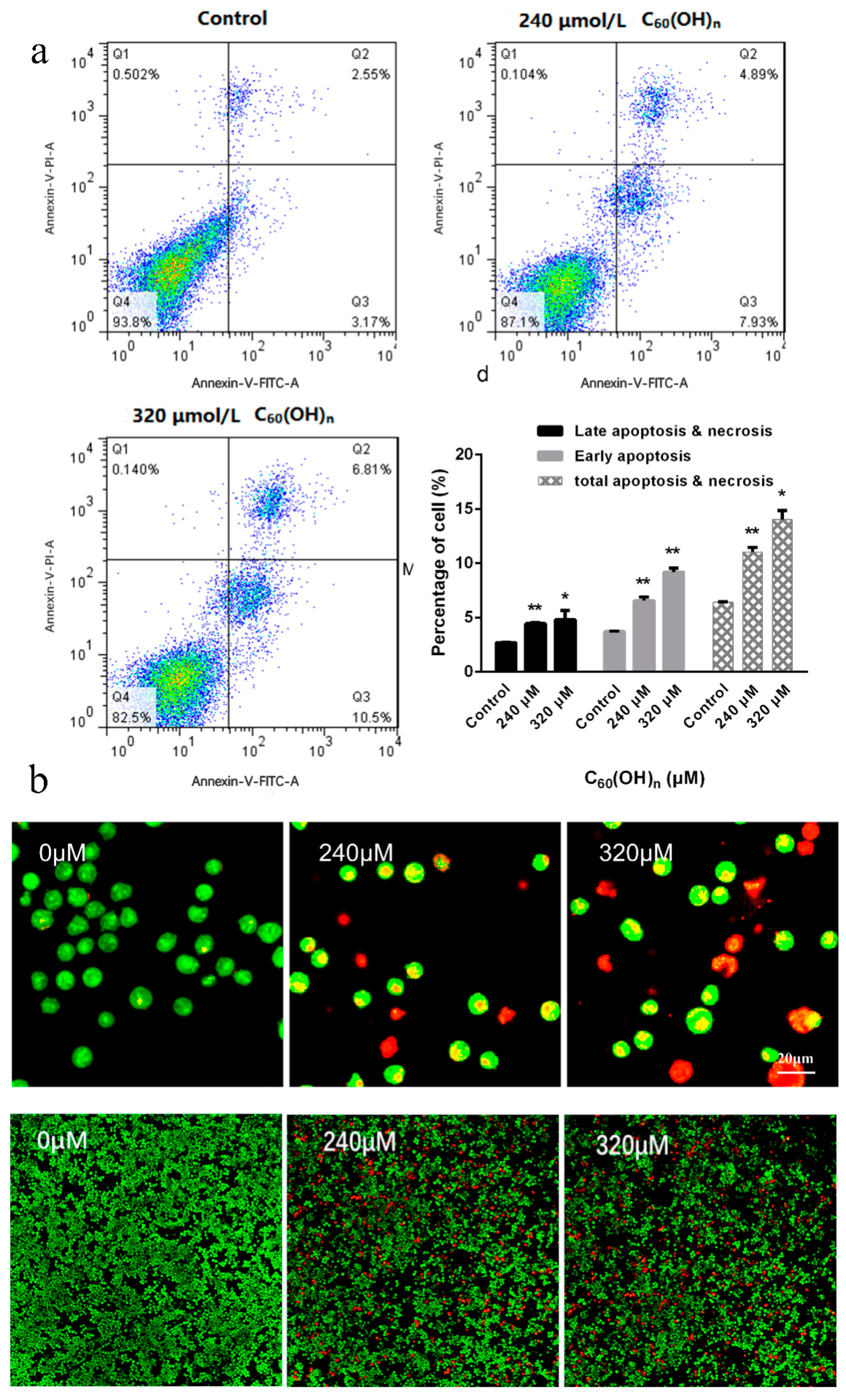
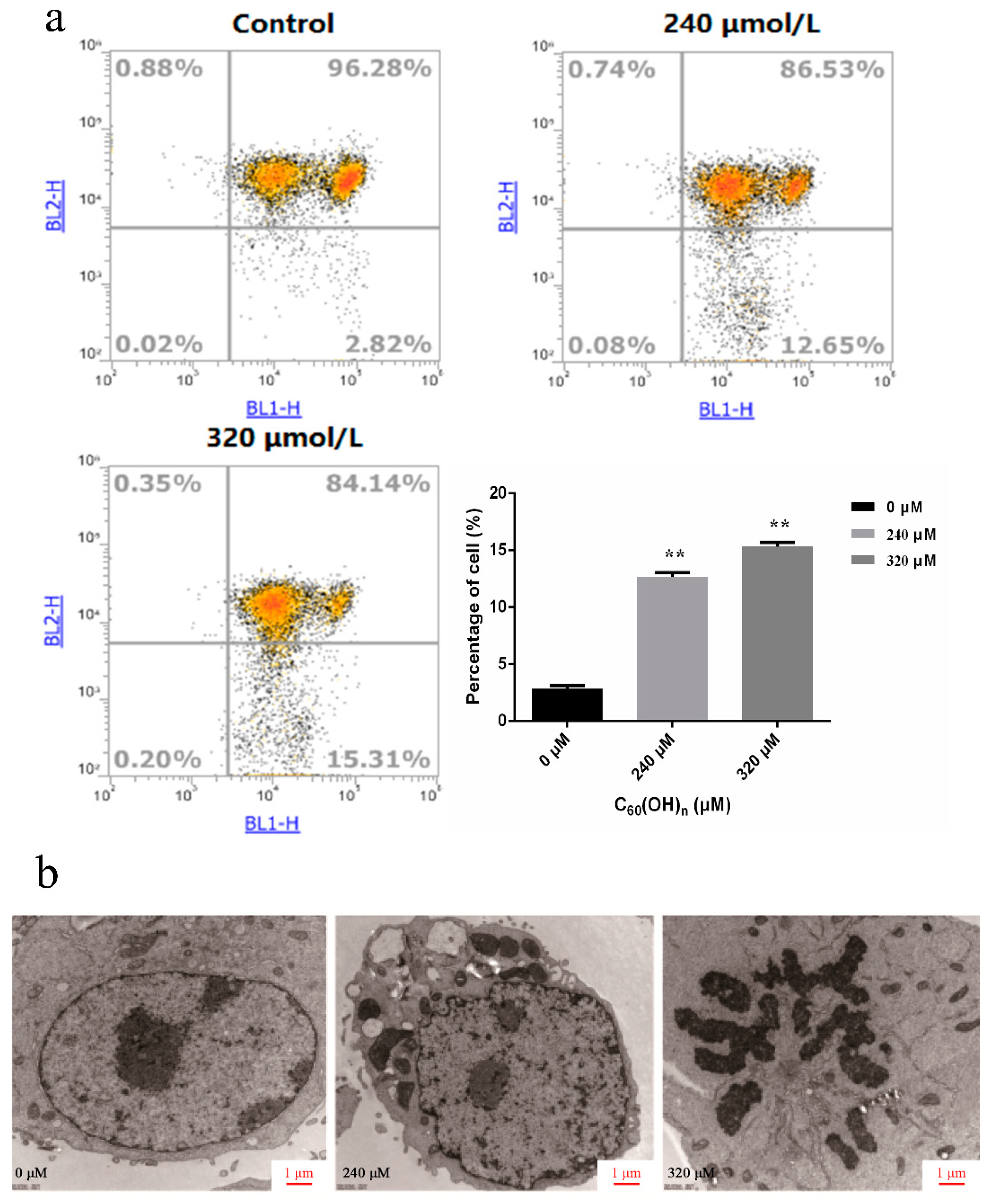
3.4. Effect of C60(OH)n on Apoptosis of K562 Cells
3.5. Effects of C60(OH)n on Mitochondrial Membrane Potential in K562 Cells
3.6. Ultrastructural Changes of Cell Apoptosis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kroto, H.W.; Heath, J.R.; O’Brien, S.C.; Curl, R.F.; Smalley, R.E. C60: Buckminsterfullerene. Nature 1985, 318, 162–163. [Google Scholar] [CrossRef]
- Dugan, L.L.; Turetsky, D.M.; Du, C. Carboxyfullerenes as neuroprotectiveagents. Proc. Natl. Acad. Sci. USA 1997, 94, 9434–9439. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Hong, K.; Roberson, C.; Ding, M.; Fernandez, A.; Shen, F.; Jin, L.; Sonkusare, S.; Li, X. Hydroxylated Fullerene: A Stellar Nanomedicine to Treat Lumbar Radiculopathy via Antagonizing TNF-alpha-Induced Ion Channel Activation, Calcium Signaling, and Neuropeptide Production. ACS Biomater. Sci. Eng. 2018, 4, 266–277. [Google Scholar] [CrossRef]
- Li, J.; Guan, M.; Wang, T.; Zhen, M.; Zhao, F.; Shu, C.; Wang, C. Gd@C82-(ethylenediamine)8 Nanoparticle: A New High-Efficiency Water-Soluble ROS Scavenger. ACS Appl. Mater. Interfaces 2016, 8, 25770–25776. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhen, M.; Guan, M.; Yu, T.; Ma, L.; Li, W.; Zheng, J.; Shu, C.; Wang, C. Amino acid modified [70] fullerene derivatives with high radical scavenging activity as promising bodyguards for chemotherapy protection. Sci. Rep. 2018, 8, 16573. [Google Scholar] [CrossRef]
- Meng, H.; Xing, G.; Sun, B.; Zhao, F.; Lei, H.; Li, W.; Song, Y.; Chen, Z.; Yuan, H.; Wang, X.; et al. Potent Angiogenesis Inhibition by the Particulate Form of Fullerene Derivatives. Acs Nano 2010, 4, 2773–2783. [Google Scholar] [CrossRef]
- Zhu, J.; Ji, Z.; Wang, J.; Sun, R.; Zhang, X.; Gao, Y.; Sun, H.; Liu, Y.; Wang, Z.; Li, A.; et al. Tumor-inhibitory effect and immunomodulatory activity of fullerol C60(OH)x. Small 2008, 4, 1168–1175. [Google Scholar] [CrossRef]
- Balogh, L.P. Caging cancer. Nanomedicine 2015, 11, 867–869. [Google Scholar] [CrossRef]
- Chen, Y.W. Fullerene derivatives protect against oxidative stress in RAW 264.7 cells and ischemia-reperfused lungs. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R21. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Suzuki, T.; Ishii, H.; Nakae, D.; Ogata, A. Cytotoxic effects of hydroxylated fullerenes on isolated rat hepatocytes via mitochondrial dysfunction. Arch. Toxicol. 2011, 85, 1429–1440. [Google Scholar] [CrossRef]
- Markman, J.L.; Rekechenetskiy, A.; Holler, E.; Ljubimova, J.Y. Nanomedicine therapeutic approaches to overcome cancer drug resistance. Adv. Drug Deliv. Rev. 2013, 65, 1866–1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, F.; Liu, Y.; Qu, Y.; Li, W.; Zhou, G.; Ge, C.; Li, Y.; Sun, B.; Chen, C. Studies on anti-tumor and antimetastatic activities of fullerenol in a mouse breast cancer model. Carbon 2010, 48, 2231–2243. [Google Scholar] [CrossRef]
- Ueno, H.; Yamakura, S.; Arastoo, R.S.; Oshima, T.; Kokubo, K. Systematic Evaluation and Mechanistic Investigation of Antioxidant Activity of Fullerenols Using β-Carotene Bleaching Assay. J. Nanomater. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Semenov, K.N.; Charykov, N.A.; Postnov, V.N.; Sharoyko, V.V.; Vorotyntsev, I.V.; Galagudza, M.M.; Murin, I.V. Fullerenols: Physicochemical properties and applications. Prog. Solid State Chem. 2016, 44, 59–74. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhen, M.; Ma, H.; Li, J.; Shu, C.; Wang, C. Inhalable gadofullerenol/[70] fullerenol as high-efficiency ROS scavengers for pulmonary fibrosis therapy. Nanomedicine 2018, 14, 1361–1369. [Google Scholar] [CrossRef]
- Guan, M.; Zhou, Y.; Liu, S.; Chen, D.; Ge, J.; Deng, R.; Li, X.; Yu, T.; Xu, H.; Sun, D.; et al. Photo-triggered gadofullerene: Enhanced cancer therapy by combining tumor vascular disruption and stimulation of anti-tumor immune responses. Biomaterials 2019, 213, 119218. [Google Scholar] [CrossRef]
- Meng, J.; Liang, X.; Chen, X.; Zhao, Y. Biological characterizations of [Gd@C82(OH)22]n nanoparticles as fullerene derivatives for cancer therapy. Integr. Biol. 2013, 5, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, C.; Qian, P.; Lu, X.; Sun, B.; Zhang, X.; Wang, L.; Gao, X.; Li, H.; Chen, Z.; et al. Gd-metallofullerenol nanomaterial as non-toxic breast cancer stem cell-specific inhibitor. Nat. Commun. 2015, 6, 5988. [Google Scholar] [CrossRef]
- Li, L.; Zhen, M.; Wang, H.; Sun, Z.; Jia, W.; Zhao, Z.; Zhou, C.; Liu, S.; Wang, C.; Bai, C. Functional Gadofullerene Nanoparticles Trigger Robust Cancer Immunotherapy Based on Rebuilding an Immunosuppressive Tumor Microenvironment. Nano Lett. 2020, 20, 4487–4496. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, Z.; Wang, Z.; Liu, Y.; Jacobson, O.; Shen, Z.; Fu, X.; Chen, Z.Y.; Chen, X. Gadolinium Metallofullerene-Based Activatable Contrast Agent for Tumor Signal Amplification and Monit, oring of Drug Release. Small 2019, 15, e1900691. [Google Scholar] [CrossRef]
- Chaudhuri, P.; Harfouche, R.; Soni, S.; Hentschel, D.M.; Sengupta, S. Shape Effect of Carbon Nano-vectors on angiogenesis. ACS Nano 2010, 4, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, H. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar]
- Krusic, P.J.; Wasserman, E.; Keizer, P.N.; Morton, J.R.; Preston, K.F. Radical Reactions of C60. Science 1991, 254, 1183–1185. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.C.; Jiang, X.; Wang, J.; Chen, C.; Liu, R.S. Nano-bio effects: Interaction of nanomaterials with cells. Nanoscale 2013, 5, 3547–3569. [Google Scholar] [CrossRef] [PubMed]
- Sawyers, C.L. Chronic myeloid leukemia. N. Engl. J. Med. 1999, 340, 1330–1340. [Google Scholar] [CrossRef] [PubMed]
- Hochhaus, A.; Baccarani, M.; Silver, R.T.; Schiffer, C.; Apperley, J.F.; Cervantes, F.; Clark, R.E.; Cortes, J.E.; Deininger, M.W.; Guilhot, F.; et al. European LeukemiaNet 2020 recommendations for treating chronic myeloid leukemia. Leukemia 2020, 34, 966–984. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, H.; Radich, J.; Garcia-Gonzalez, P. Meeting the needs of CML patients in resource-poor countries. Hematology. American Society of Hematology. Educ. Program 2019, 2019, 433–442. [Google Scholar]
- Liu, W.J.; Zhang, T.; Guo, Q.L.; Liu, C.Y.; Bai, Y.Q. Effect of ATRA on the expression of HOXA5 gene in K562 cells and its relationship with cell cycle and apoptosis. Mol. Med. Rep. 2016, 13, 4221–4228. [Google Scholar] [CrossRef] [Green Version]
- Lozzio, B.B.; Lozzio, C.B. Properties of the K562 cell line derived from a patient with chronic myeloid leukemia. Int. J. Cancer 1977, 19, 136. [Google Scholar] [CrossRef]
- Lee, C.W.; Chi, M.C.; Peng, K.T.; Chiang, Y.C.; Hsu, L.F.; Yan, Y.L.; Li, H.Y.; Chen, M.C.; Lee, I.T.; Lai, C.H. Water-Soluble Fullerenol C60(OH)36 toward Effective Anti-Air Pollution Induced by Urban Particulate Matter in HaCaT Cell. Int. J. Mol. Sci. 2019, 20, 4259. [Google Scholar] [CrossRef] [Green Version]
- Ihara, T.; Yamamoto, T.; Sugamata, M.; Okumura, H.; Ueno, Y. The process of ultrastructural changes from nuclei to apoptotic body. Virchows Arch. 1998, 433, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Arvizo, R.R.; Bhattacharyya, S.; Kudgus, R.A.; Giri, K.; Bhattacharya, R.; Mukherjee, P. Intrinsic therapeutic applications of noble metal nanoparticles: Past, present and future. Chem. Soc. Rev. 2012, 41, 2943–2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zheng, F.; Yang, T.; Zhou, W.; Liu, Y.; Man, N.; Zhang, L.; Jin, N.; Dou, Q.; Zhang, Y.; et al. Tuning the autophagy-inducing activity of lanthanide-based nanocrystals through specifific surface-coating peptides. Nat. Mater. 2012, 11, 817–826. [Google Scholar] [CrossRef]
- Srdjenovic, B.U.; Slavic, M.N.; Stankov, K.M.; Kladar, N.V.; Jović, D.S.; Seke, M.N.; Bogdanović, V.V.; Hemijska, I. Size distribution of fullerenol nanoparticles in cell culture medium and their influence on antioxidative enzymes in Chinese hamster ovary cells. Hem. Ind. 2015, 69, 425–431. [Google Scholar] [CrossRef] [Green Version]
- Jović, D.S.; Seke, M.N.; Djordjevic, A.N.; Mrđanović, J.Ž.; Aleksić, L.D.; Bogdanović, G.M.; Pavić, A.B.; Plavec, J. Fullerenol nanoparticles as a new delivery system for doxorubicin. RSC Adv. 2016, 6, 38563–38578. [Google Scholar] [CrossRef]
- Liu, Y.; Jiao, F.; Qiu, Y.; Li, W.; Qu, Y.; Tian, C.; Li, Y.; Bai, R.; Lao, F.; Zhao, Y.; et al. Immunostimulatory properties and enhanced TNF- alpha mediated cellular immunity for tumor therapy by C60(OH)20 nanoparticles. Nanotechnology 2009, 20, 415102. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, W.; Liu, X.; Ye, L.; Liu, J.; Larwubah, K.; Meng, G.; Shen, W.; Ying, X.; Zhu, J.; Yang, S.; et al. The Effect of Polyhydroxy Fullerene Derivative on Human Myeloid Leukemia K562 Cells. Materials 2022, 15, 1349. https://doi.org/10.3390/ma15041349
Guo W, Liu X, Ye L, Liu J, Larwubah K, Meng G, Shen W, Ying X, Zhu J, Yang S, et al. The Effect of Polyhydroxy Fullerene Derivative on Human Myeloid Leukemia K562 Cells. Materials. 2022; 15(4):1349. https://doi.org/10.3390/ma15041349
Chicago/Turabian StyleGuo, Wei, Xing Liu, Lianjie Ye, Jie Liu, Kollie Larwubah, Ge Meng, Weiqiang Shen, Xiangxian Ying, Jun Zhu, Shengjie Yang, and et al. 2022. "The Effect of Polyhydroxy Fullerene Derivative on Human Myeloid Leukemia K562 Cells" Materials 15, no. 4: 1349. https://doi.org/10.3390/ma15041349