
Challenges in the Vaccination of the Elderly and Strategies for Improvement

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
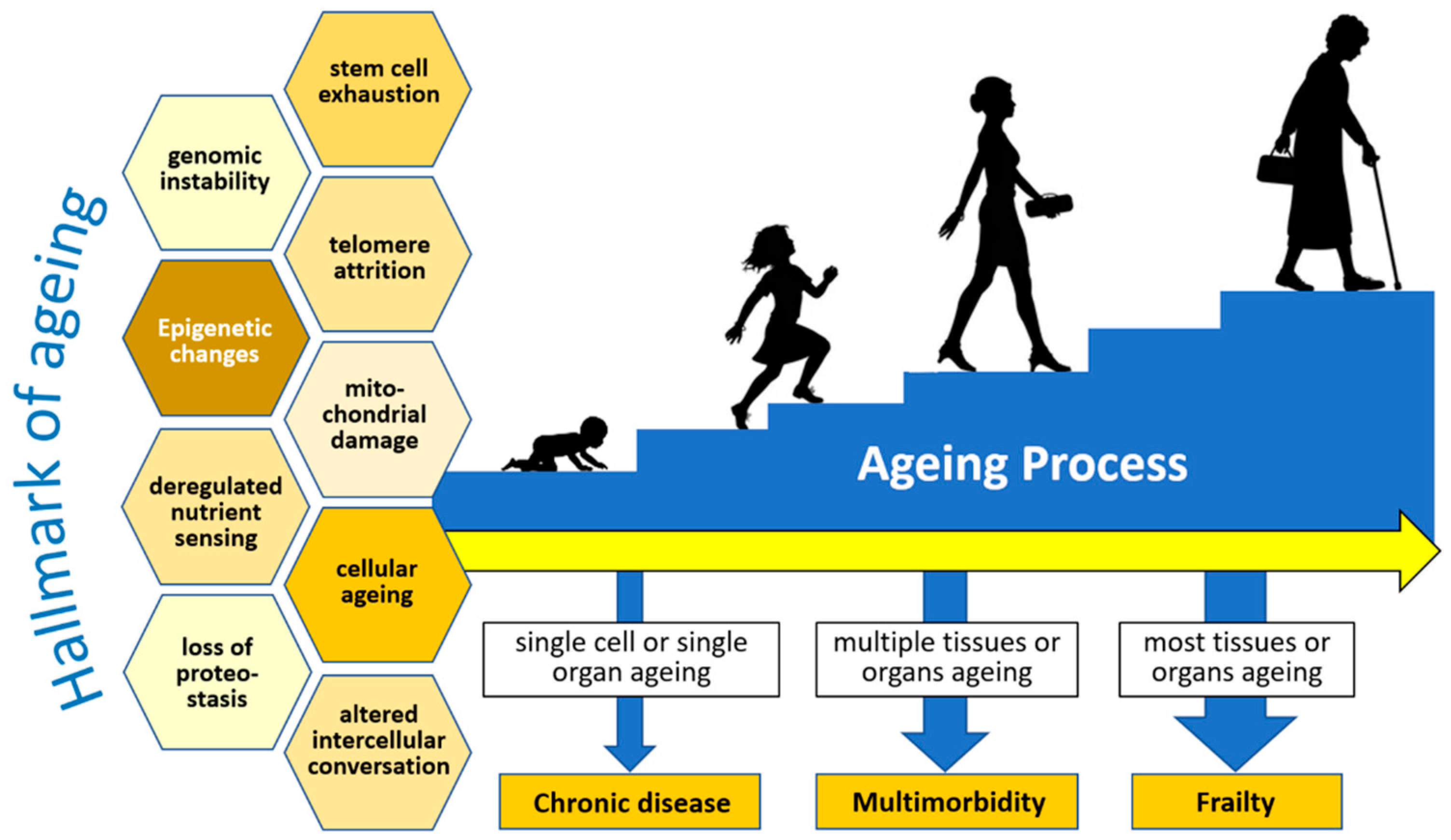
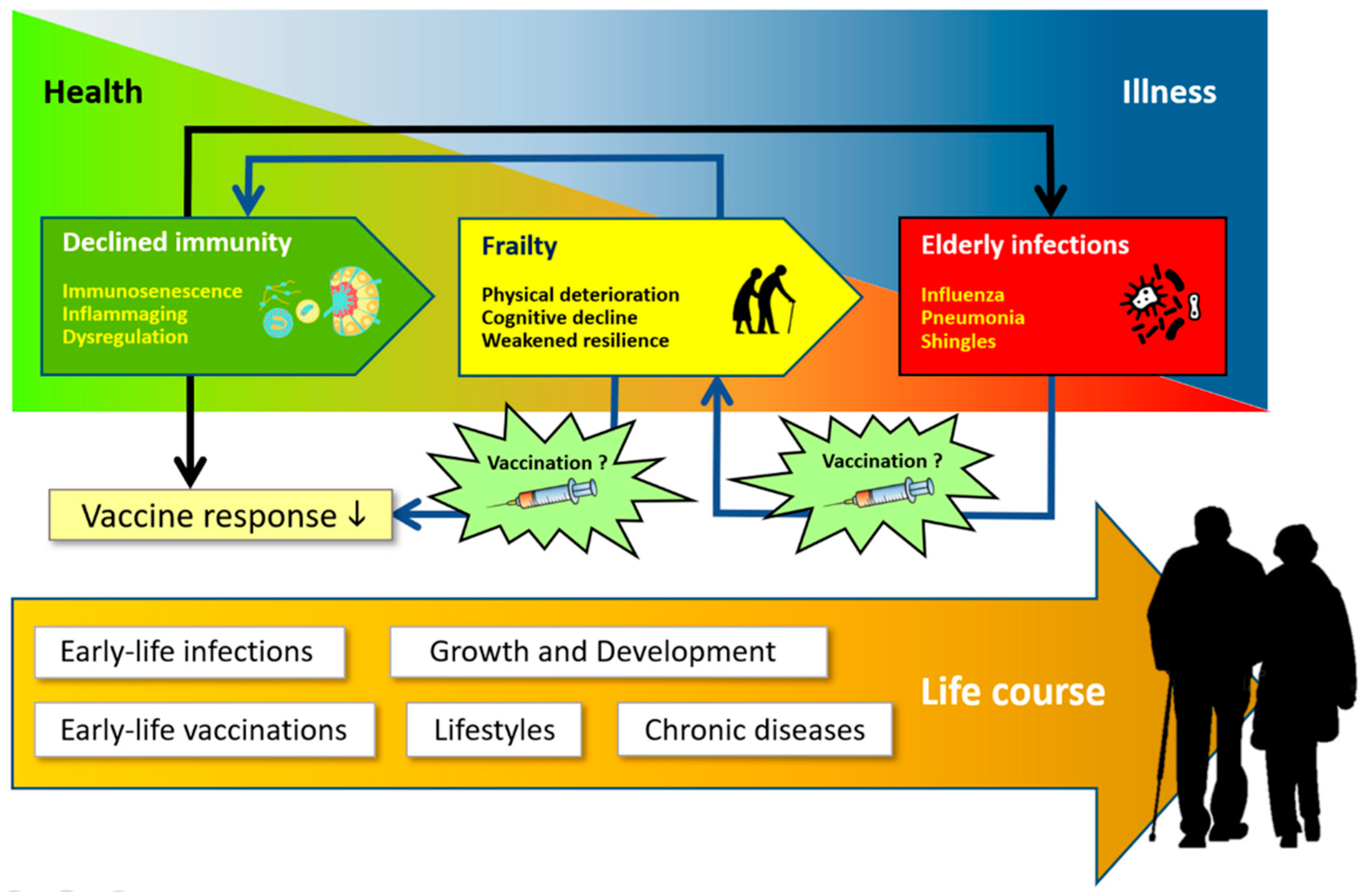
2. Frailty, Immunosenescence, and Inflammaging in the Elderly
3. Infection Threat in the Elderly
3.1. Pneumococcal Pneumonia
3.2. Influenza
3.3. Herpes Zoster
3.4. Coronavirus Disease 2019 (COVID-19)
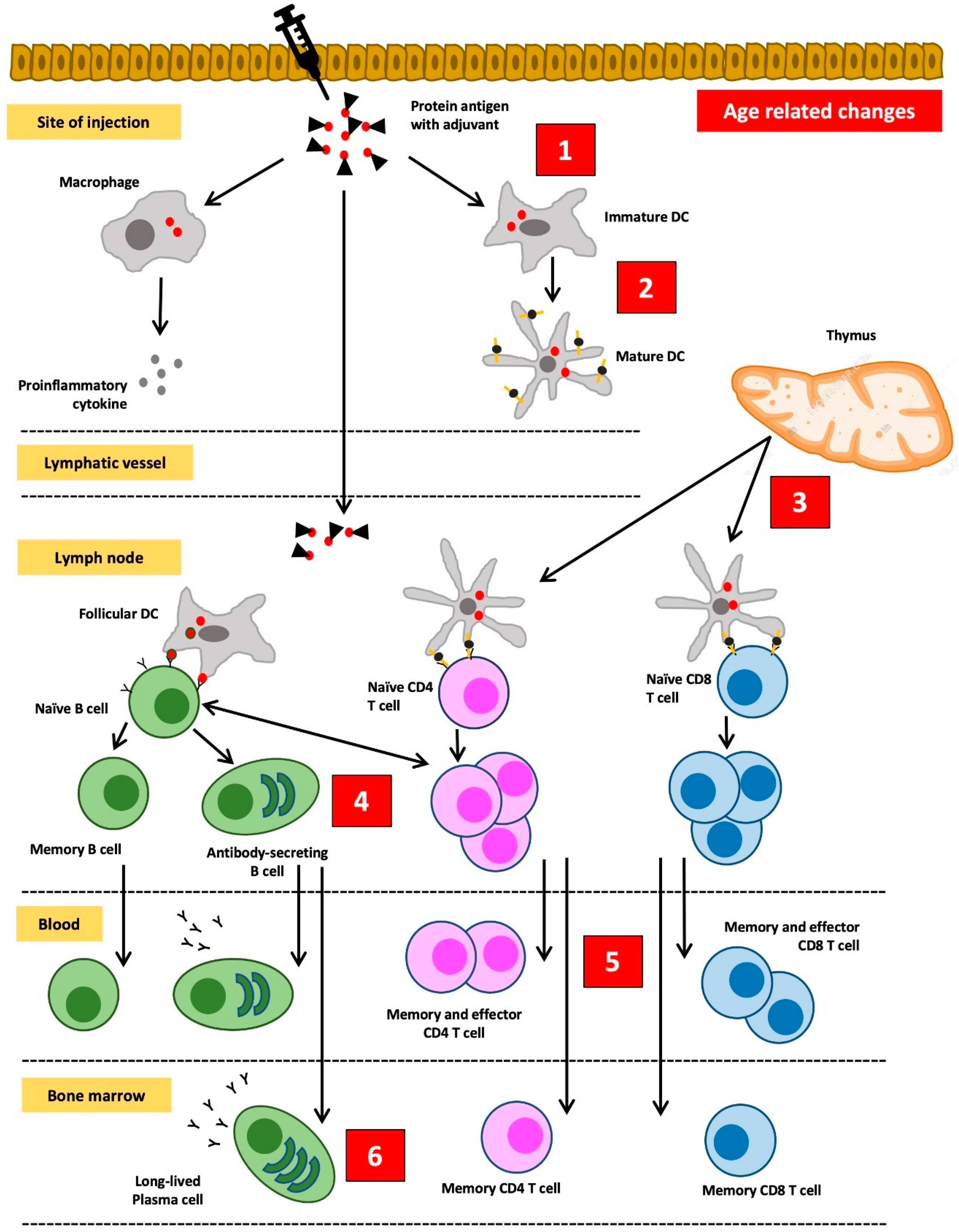
4. Decreased Response to Vaccines among the Elderly
5. Strategies to Increase the Vaccine Response in the Elderly
5.1. The Use of Higher Antigen Doses
5.2. The Use of More Immunogenic Adjuvants
5.3. Delivering Booster Injections
5.4. Exploration of Alternative Routes of Vaccine Administration
5.5. Vaccine Injection Time
5.6. The Use of Senolytic and Immunomodulatory Drugs
6. Recommended Vaccines for the Elderly
6.1. Influenza Vaccine
6.2. Pneumococcal Vaccine
- For elderly ≥65 years who have previously received the PPSV-23 vaccine, it is recommended to receive one dose of the PCV-15 or PCV-20 vaccine at least a year after the previous PPSV-23 injection. There is no longer a need for a booster shot with PPSV-23.
- For elderly ≥65 years who have previously received the PCV-13 vaccine, it is recommended to get the PPSV-23 or PCV-20 vaccine (if PPSV-23 is not available)
- For immunocompromised adults that reach 65 years and have previously received the PCV-13 vaccine followed by the PPSV-23 vaccine before 65 years, it is recommended to get a PPSV-23 booster vaccine with an interval of at least 5 years from the previous PPSV-23 injection.
6.3. COVID-19 Vaccine
6.4. Herpes Zoster Vaccine
6.5. Diphtheria and Tetanus Vaccine
6.6. Other Vaccines
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, W.; Goodkind, D.; Kowal, P. An Aging World; United States Census Bureau: Washington, DC, USA, 2015.
- Aw, D.; Silva, A.B.; Palmer, D.B. Immunosenescence: Emerging challenges for an ageing population. Immunology 2007, 120, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A.; Farzaneh, F.; Candore, G.; Caruso, C.; Davinelli, S.; Gambino, C.M.; Ligotti, M.E.; Zareian, N.; Accardi, G. Immunosenescence and its hallmarks: How to oppose aging strategically? A review of potential options for therapeutic intervention. Front. Immunol. 2019, 10, 2247. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2014, 69, S4–S9. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A new immune–metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 2018, 14, 576–590. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Dupuis, G.; Baehl, S.; Le Page, A.; Bourgade, K.; Frost, E.; Witkowski, J.M.; Pawelec, G.; Larbi, A.; Cunnane, S. From inflamm-aging to immune-paralysis: A slippery slope during aging for immune-adaptation. Biogerontology 2016, 17, 147–157. [Google Scholar] [CrossRef]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Claudio Franceschi, C. Immunosenescence and inflamm-aging as two sides of the same coin: Friends or foes? Front. Immunol. 2018, 8, 1960. [Google Scholar] [CrossRef]
- Fülöp, T.; Dupuis, G.; Witkowski, J.M.; Larbi, A. The role of immunosenescence in the development of age-related diseases. Rev. Investig. Clin. 2016, 68, 84–91. [Google Scholar]
- Fedarko, N.S. The biology of aging and frailty. Clin. Geriatr. Med. 2011, 27, 27–37. [Google Scholar] [CrossRef]
- Vetrano, D.L.; Triolo, F.; Maggi, S.; Malley, R.; Jackson, T.A.; Poscia, A.; Bernabei, R.; Ferrucci, L.; Fratiglioni, L. Fostering healthy aging: The interdependency of infections, immunity and frailty. Ageing Res. Rev. 2021, 69, 101351. [Google Scholar] [CrossRef]
- Kline, K.A.; Bowdish, D.M.E. Infection in an aging population. Curr. Opin. Microbiol. 2016, 29, 63–67. [Google Scholar] [CrossRef]
- Cunningham, A.L.; McIntyre, P.; Subbarao, K.; Booy, R.; Levin, M.J. Vaccines for older adults. BMJ 2021, 372, n188. [Google Scholar] [CrossRef]
- Fulop, T.; Larbi, A.; Pawelec, G.; Cohen, A.A.; Provost, G.; Khalil, A.; Lacombe, G.; Rodrigues, S.; Desroches, M.; Hirokawa, K.; et al. Immunosenescence and altered vaccine efficiency in older subjects: A myth difficult to change. Vaccines 2022, 10, 607. [Google Scholar] [CrossRef]
- Aiello, A.; Ligotti, M.E.; Garnica, M.; Accardi, G.; Calabrò, A.; Pojero, F.; Arasanz, H.; Bocanegra, A.; Blanco, E.; Chocarro, L.; et al. How can we improve vaccination response in old people? Part I: Targeting immunosenescence of innate immunity cells. Int. J. Mol. Sci. 2022, 23, 9880. [Google Scholar] [CrossRef]
- Garnica, M.; Aiello, A.; Ligotti, M.E.; Accardi, G.; Arasanz, H.; Bocanegra, A.; Blanco, E.; Calabrò, A.; Chocarro, L.; Echaide, M.; et al. How can we improve the vaccination response in older people? Part II: Targeting immunosenescence of adaptive immunity cells. Int. J. Mol. Sci. 2022, 23, 9797. [Google Scholar] [CrossRef]
- Loukov, D.; Naidoo, A.; Bowdish, D. Immunosenescence: Implications for vaccination programs in the elderly. Vaccine Dev. Ther. 2015, 5, 17–29. [Google Scholar]
- Pereira, B.; Xu, X.N.; Akbar, A.N. Targeting inflammation and immunosenescence to improve vaccine responses in the elderly. Front. Immunol. 2020, 14, 583019. [Google Scholar] [CrossRef]
- Hägg, S.; Religa, D. COVID vaccination in older adults. Nat. Microbiol. 2022, 7, 1106–1107. [Google Scholar] [CrossRef]
- Thillainadesan, J.; Scott, I.A.; Le Couteur, D.G. Frailty, a multisystem ageing syndrome. Age Ageing 2020, 49, 758–763. [Google Scholar] [CrossRef]
- Teissier, T.; Boulanger, E.; Cox, L.S. Interconnections between inflammageing and immunosenescence during ageing. Cells 2022, 11, 359. [Google Scholar] [CrossRef]
- Bartleson, J.M.; Radenkovic, D.; Covarrubias, A.J.; Furman, D.; Winer, D.A.; Verdin, E. SARS-CoV-2, COVID-19 and the aging immune system. Nat. Aging 2021, 1, 769–782. [Google Scholar] [CrossRef]
- Esme, M.; Topeli, A.; Yavuz, B.B.; Akova, M. Infections in the elderly critically-ill patients. Front. Med. 2019, 6, 118. [Google Scholar] [CrossRef] [PubMed]
- Australian Institute of Health and Welfare. The Burden of Vaccine Preventable Diseases in Australia; Australian Institute of Health and Welfare: Canberra, Australia, 2019; p. 152. [Google Scholar]
- Cassini, A.; Colzani, E.; Pini, A.; Mangen, M.-J.J.; Plass, D.; McDonald, S.A.; Maringhini, G.; van Lier, A.; Haagsma, J.A.; Havelaar, A.H.; et al. Impact of infectious diseases on population health using incidence-based disability-adjusted life years (DALYs): Results from the Burden of Communicable Diseases in Europe Study, European Union and European Economic Area countries, 2009 to 2013. Eurosurveillance 2018, 23, 17-00454. [Google Scholar] [CrossRef] [PubMed]
- Kwong, J.C.; Ratnasingham, S.; Campitelli, M.A.; Daneman, N.; Deeks, S.L.; Manuel, D.G.; Allen, V.G.; Bayoumi, A.M.; Fazil, A.; Fisman, D.N.; et al. The impact of infection on population health: Results of the Ontario Burden of Infectious Diseases Study. PLoS ONE 2012, 7, e44103. [Google Scholar] [CrossRef] [PubMed]
- Tam, E.M.Y.Y.; Kwan, Y.K.; Ng, Y.Y.; Yam, P.W. Clinical course and mortality in older patients with COVID-19: A cluster-based study in Hong Kong. Hong Kong Med. J. 2022, 28, 215–222. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Ageing Collaborators. Global, regional, and national burden of diseases and injuries for adults 70 years and older: Systematic analysis for the Global Burden of Disease 2019 Study. BMJ 2022, 376, e068208. [Google Scholar]
- Li, X.; Mukandavire, C.; Cucunubá, Z.M.; Echeverria-Londono, S.; Abbas, K.; Clapham, H.E.; Jit, M.; Johnson, H.L.; Papadopoulos, T.; Vynnycky, E.; et al. Estimating the health impact of vaccination against ten pathogens in 98 low-income and middle-income countries from 2000 to 2030: A modelling study. Lancet 2021, 397, 398–408. [Google Scholar] [CrossRef]
- Toor, J.; Echeverria-Londono, S.; Li, X.; Abbas, K.; Carter, E.D.; Clapham, H.E.; Clark, A.; de Villiers, M.J.; Eilertson, K.; Ferrari, M.; et al. Lives saved with vaccination for 10 pathogens across 112 countries in a pre-COVID-19 world. eLife 2021, 10, e67635. [Google Scholar] [CrossRef]
- John, D.; Narassima, M.S.; Bhattacharya, P.; Mukherjee, N.; Banerjee, A.; Menon, J. Model-based estimation of burden of COVID-19 with disability-adjusted life years and value of statistical life in West Bengal, India. BMJ Open 2023, 13, e065729. [Google Scholar] [CrossRef]
- Li, Z.W.; Yin, C.N.; Wang, H.T.; Du, W.Y.; Wang, W.J.; Xue, F.Z.; Zhao, L.; Cao, W.-C.; Cheeloo EcoHealth Consortium (CLEC). Incidence trend and disease burden of seven vaccine-preventable diseases in Shandong province, China, 2013–2017: Findings from a population-based observational study. Vaccine X 2022, 10, 100145. [Google Scholar] [CrossRef]
- Wyper, G.M.A.; Fletcher, E.; Grant, I.; McCartney, G.; Fischbacher, C.; Harding, O.; Jones, H.; de Haro Moro, M.T.; Speybroeck, N.; Devleesschauwer, B.; et al. Measuring disability-adjusted life years (DALYs) due to COVID-19 in Scotland, 2020. Arch. Public Health 2022, 80, 105. [Google Scholar] [CrossRef]
- Lebrasseur, A.; Fortin-Bédard, N.; Lettre, J.; Raymond, E.; Bussières, E.L.; Lapierre, N.; Faieta, J.; Vincent, C.; Duchesne, L.; Ouellet, M.-C.; et al. Impact of the COVID-19 pandemic on older adults: Rapid review. JMIR Aging 2021, 4, e26474. [Google Scholar] [CrossRef]
- Wong, M.K.; Brooks, D.J.; Ikejezie, J.; Gacic-Dobo, M.; Dumolard, L.; Nedelec, Y.; Steulet, C.; Kassamali, Z.; Acma, A.; Ajong, B.N.; et al. COVID-19 mortality and progress toward vaccinating older adults—World Health Organization, worldwide, 2020–2022. Morb. Mortal. Wkly. Rep. MMWR 2023, 72, 113–118. [Google Scholar] [CrossRef]
- Lansbury, L.; Lim, B.; McKeever, T.M.; Lawrence, H.; Lim, W.S. Non-invasive pneumococcal pneumonia due to vaccine serotypes: A systematic review and meta-analysis. EClinicalMedicine 2022, 44, 101271. [Google Scholar] [CrossRef]
- Uyeki, T.M.; Hui, D.S.; Zambon, M.; Wentworth, D.E.; Monto, A.S. Influenza. Lancet 2022, 400, 693–706. [Google Scholar] [CrossRef]
- Macias, A.E.; McElhaney, J.E.; Chaves, S.S.; Nealon, J.; Nunes, M.C.; Samson, S.I.; Seet, B.T.; Weinke, T.; Yu, H. The disease burden of influenza beyond respiratory illness. Vaccine 2021, 39, A6–A14. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention (CDC). Clinical Overview of Herpes Zoster (Shingles); Center for Disease Control and Prevention (CDC): Atlanta, GA, USA, 2020.
- Cucinotta, D.; Vanelli, M. WHO declares COVID-19 a pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar] [CrossRef]
- World Health Organization (WHO). WHO Coronavirus (COVID-19) Dashboard; World Health Organization (WHO): Geneva, Switzerland, 2023. [Google Scholar]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef]
- Verity, R.; Okell, L.C.; Dorigatti, I.; Winskill, P.; Whittaker, C.; Imai, N.; Cuomo-Dannenburg, G.; Thompson, H.; Walker, P.G.T.; Fu, H.; et al. Estimates of the severity of coronavirus disease 2019: A model-based analysis. Lancet Infect. Dis. 2020, 20, 669–677. [Google Scholar] [CrossRef]
- Hartantri, Y.; Debora, J.; Widyatmoko, L.; Giwangkancana, G.; Suryadinata, H.; Susandi, E.; Hutajulu, E.; Hakiman, A.P.A.; Pusparini, Y.; Alisjahbana, B. Clinical and treatment factors associated with the mortality of COVID-19 patients admitted to a referral hospital in Indonesia. Lancet Reg. Health Southeast Asia 2023, 11, 100167. [Google Scholar] [CrossRef]
- Nidadavolu, L.S.; Walston, J.D. Underlying vulnerabilities to the cytokine storm and adverse COVID-19 outcomes in the aging immune system. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, e13–e18. [Google Scholar] [CrossRef]
- Sherwani, S.; Khan, M.W.A. Cytokine response in SARS-CoV-2 infection in the elderly. J. Inflamm. Res. 2020, 13, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Meftahi, G.H.; Jangravi, Z.; Sahraei, H.; Bahari, Z. The possible pathophysiology mechanism of cytokine storm in elderly adults with COVID-19 infection: The contribution of “inflame-aging”. Inflamm. Res. 2020, 69, 825–839. [Google Scholar] [CrossRef] [PubMed]
- Tizazu, A.M.; Mengist, H.M.; Demeke, G. Aging, inflammaging and immunosenescence as risk factors of severe COVID-19. Immun. Ageing 2022, 19, 53. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Curtis, N. Factors that influence the immune response to vaccination. Clin. Microbiol. Rev. 2019, 32, e00084-00018. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, B.; Herndler-Brandstetter, D.; Schwanninger, A.; Weiskopf, D.; Grubeck-Loebenstein, B. Biology of immune responses to vaccines in elderly persons. Clin. Infect. Dis. 2008, 46, 1078–1084. [Google Scholar] [CrossRef]
- Coe, C.L.; Lubach, G.R.; Kinnard, J. Immune senescence in old and very old rhesus monkeys: Reduced antibody response to influenza vaccination. Age 2012, 34, 1169–1177. [Google Scholar] [CrossRef]
- Murasko, D. Role of humoral and cell-mediated immunity in protection from influenza disease after immunization of healthy elderly. Exp. Gerontol. 2002, 37, 427–439. [Google Scholar] [CrossRef]
- Ademokun, A.; Wu, Y.-C.; Martin, V.; Mitra, R.; Sack, U.; Baxendale, H.; Kipling, D.; Dunn-Walters, D.K. Vaccination-induced changes in human B-cell repertoire and pneumococcal IgM and IgA antibody at different ages: Vaccine-induced changes in B-cell repertoire. Aging Cell 2011, 10, 922–930. [Google Scholar] [CrossRef]
- Schenkein, J.G.; Park, S.; Nahm, M.H. Pneumococcal vaccination in older adults induces antibodies with low opsonic capacity and reduced antibody potency. Vaccine 2008, 26, 5521–5526. [Google Scholar] [CrossRef]
- Oxman, M.N.; Levin, M.J.; Johnson, G.R.; Schmader, K.E.; Straus, S.E.; Gelb, L.D.; Arbeit, R.D.; Simberkoff, M.S.; Gershon, A.A.; Davis, L.E.; et al. A vaccine to prevent herpes zoster and postherpetic neuralgia in older adults. N. Engl. J. Med. 2005, 352, 2271–2284. [Google Scholar] [CrossRef]
- Weinberger, B. Vaccines for the elderly: Current use and future challenges. Immun. Ageing 2018, 15, 3. [Google Scholar] [CrossRef]
- Robertson, C.A.; DiazGranados, C.A.; Decker, M.D.; Chit, A.; Mercer, M.; Greenberg, D.P. Fluzone® high-dose influenza vaccine. Expert Rev. Vaccines 2016, 15, 1495–1505. [Google Scholar] [CrossRef]
- Chang, L.-J.; Meng, Y.; Janosczyk, H.; Landolfi, V.; Talbot, H.K. Safety and immunogenicity of high-dose quadrivalent influenza vaccine in adults ≥65 years of age: A phase 3 randomized clinical trial. Vaccine 2019, 37, 5825–5834. [Google Scholar] [CrossRef]
- Weinberger, B. Adjuvant strategies to improve vaccination of the elderly population. Curr. Opin. Pharmacol. 2018, 41, 34–41. [Google Scholar] [CrossRef]
- Khurana, S.; Verma, N.; Yewdell, J.W.; Hilbert, A.K.; Castellino, F.; Lattanzi, M.; Del Giudice, G.; Rappuoli, R.; Golding, H. MF59 adjuvant enhances diversity and affinity of antibody-mediated immune response to pandemic influenza vaccines. Sci. Transl. Med. 2011, 3, 85ra48. [Google Scholar] [CrossRef]
- Kayraklioglu, N.; Horuluoglu, B.; Klinman, D.M. CpG Oligonucleotides as Vaccine Adjuvants. In DNA Vaccines; Sousa, Â., Ed.; Springer: New York, NY, USA, 2021; Volume 2197, pp. 51–85. [Google Scholar]
- Fan, J.; Jin, S.; Gilmartin, L.; Toth, I.; Hussein, W.M.; Stephenson, R.J. Advances in infectious disease vaccine adjuvants. Vaccines 2022, 10, 1120. [Google Scholar] [CrossRef]
- Schwarz, T.F.; Volpe, S.; Catteau, G.; Chlibek, R.; David, M.P.; Richardus, J.H.; Lal, H.; Oostvogels, L.; Pauksens, K.; Ravault, S.; et al. Persistence of immune response to an adjuvanted varicella-zoster virus subunit vaccine for up to year nine in older adults. Hum. Vaccines Immunother. 2018, 14, 1370–1377. [Google Scholar] [CrossRef]
- Kang, I.; Hong, M.S.; Nolasco, H.; Park, S.H.; Dan, J.M.; Choi, J.-Y.; Craft, J. Age-associated change in the frequency of memory CD4+ T cells impairs long term CD4+ T cell responses to influenza vaccine. J. Immunol. 2004, 173, 673–681. [Google Scholar] [CrossRef]
- Mwimanzi, F.; Lapointe, H.R.; Cheung, P.K.; Sang, Y.; Yaseen, F.; Umviligihozo, G.; Kalikawe, R.; Datwani, S.; Omondi, F.H.; Burns, L.; et al. Older adults mount less durable humoral responses to two doses of COVID-19 mRNA vaccine but strong initial responses to a third dose. J. Infect. Dis. 2022, 226, 983–994. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Expert Advisory Group Recommends COVID-19 Vaccination Strategy for the Autumn Season; World Health Organization (WHO): Geneva, Switzerland, 2022. [Google Scholar]
- United Kingdom Government. A Guide to the Spring Booster for Those Aged 75 Years and Older and Older Residents in Care Homes—GOV.UK.pdf; United Kingdom Government Press: London, UK, 2022.
- Kaml, M.; Weiskirchner, I.; Keller, M.; Luft, T.; Hoster, E.; Hasford, J.; Young, L.; Bartlett, B.; Neuner, C.; Fischer, K.-H.; et al. Booster vaccination in the elderly: Their success depends on the vaccine type applied earlier in life as well as on pre-vaccination antibody titers. Vaccine 2006, 24, 6808–6811. [Google Scholar] [CrossRef]
- Holland, D.; Booy, R.; Looze Ferdinandus, D.; Eizenberg, P.; McDonald, J.; Karrasch, J.; McKeirnan, M.; Salem, H.; Mills, G.; Reid, J.; et al. Intradermal influenza vaccine administered using a new microinjection system produces superior immunogenicity in elderly adults: A randomized controlled trial. J. Infect. Dis. 2008, 198, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Belshe, R.B.; Newman, F.K.; Cannon, J.; Duane, C.; Treanor, J.; Van Hoecke, C.; Howe, B.J.; Dubin, G. Serum antibody responses after intradermal vaccination against influenza. N. Engl. J. Med. 2004, 351, 2286–2294. [Google Scholar] [CrossRef] [PubMed]
- Kenney, R.T.; Frech, S.A.; Muenz, L.R.; Villar, C.P.; Glenn, G.M. Dose sparing with intradermal injection of influenza vaccine. N. Engl. J. Med. 2004, 351, 2295–2301. [Google Scholar] [CrossRef] [PubMed]
- Marra, F.; Young, F.; Richardson, K.; Marra, C.A. Immunogenicity and adverse events: A meta-analysis of intradermal versus intramuscular influenza vaccines. Influenza Other Respir. Viruses 2013, 7, 584–603. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.M.; Atmar, R.L.; El Sahly, H.M.; Guo, K.; Hill, H.; Keitel, W.A. Direct comparison of an inactivated subvirion influenza A virus subtype H5N1 vaccine administered by the intradermal and intramuscular routes. J. Infect. Dis. 2012, 206, 1069–1077. [Google Scholar] [CrossRef]
- Luria-Pérez, R.; Sánchez-Vargas, L.A.; Muñoz-López, P.; Mellado-Sánchez, G. Mucosal vaccination: A promising alternative against flaviviruses. Front. Cell. Infect. Microbiol. 2022, 12, 887729. [Google Scholar] [CrossRef]
- Ainai, A.; Riet, E.; Ito, R.; Ikeda, K.; Senchi, K.; Suzuki, T.; Tamura, S.-I.; Asanuma, H.; Odagiri, T.; Tashiro, M.; et al. Human immune responses elicited by an intranasal inactivated H5 influenza vaccine. Microbiol. Immunol. 2020, 64, 313–325. [Google Scholar] [CrossRef]
- Scheiermann, C.; Kunisaki, Y.; Frenette, P.S. Circadian control of the immune system. Nat. Rev. Immunol. 2013, 13, 190–198. [Google Scholar] [CrossRef]
- Long, J.E.; Drayson, M.T.; Taylor, A.E.; Toellner, K.M.; Lord, J.M.; Phillips, A.C. Morning vaccination enhances antibody response over afternoon vaccination: A cluster-randomised trial. Vaccine 2016, 34, 2679–2685. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Y.; Liu, D.; Zeng, Q.; Li, L.; Zhou, Q.; Li, M.; Mei, J.; Yang, N.; Mo, S.; et al. Time of day influences immune response to an inactivated vaccine against SARS-CoV-2. Cell Res. 2021, 31, 1215–1217. [Google Scholar] [CrossRef]
- Mannick, J.B.; Del Giudice, G.; Lattanzi, M.; Valiante, N.M.; Praestgaard, J.; Huang, B.; Lonetto, M.A.; Maecker, H.T.; Kovarik, J.; Carson, S.; et al. mTOR inhibition improves immune function in the elderly. Sci. Transl. Med. 2014, 6, 268ra179. [Google Scholar] [CrossRef]
- Netti, G.S.; Infante, B.; Troise, D.; Mercuri, S.; Panico, M.; Spadaccino, F.; Catalano, V.; Gigante, M.; Simone, S.; Pontrelli, P.; et al. mTOR inhibitors improve both humoral and cellular response to SARS-CoV-2 messenger RNA BNT16b2 vaccine in kidney transplant recipients. Am. J. Transpl. 2022, 22, 1475–1482. [Google Scholar] [CrossRef]
- Zhang, H.; Alsaleh, G.; Feltham, J.; Sun, Y.; Napolitano, G.; Riffelmacher, T.; Charles, P.; Frau, L.; Hublitz, P.; Yu, Z.; et al. Polyamines control eIF5A hypusination, TFEB translation, and autophagy to reverse B cell senescence. Mol. Cell 2019, 76, 110–125.e119. [Google Scholar] [CrossRef]
- Kirkland, J.L.; Tchkonia, T. Senolytic drugs: From discovery to translation. J. Intern. Med. 2020, 288, 518–536. [Google Scholar] [CrossRef]
- Domnich, A.; Arata, L.; Amicizia, D.; Puig-Barberà, J.; Gasparini, R.; Panatto, D. Effectiveness of MF59-adjuvanted seasonal influenza vaccine in the elderly: A systematic review and meta-analysis. Vaccine 2017, 35, 513–520. [Google Scholar] [CrossRef]
- Diaz Granados, C.A.; Dunning, A.J.; Kimmel, M.; Kirby, D.; Treanor, J.; Collins, A.; Pollak, R.; Christoff, J.; Earl, J.; Landolfi, V.; et al. Efficacy of high-dose versus standard-dose influenza vaccine in older adults. N. Engl. J. Med. 2014, 371, 635–645. [Google Scholar] [CrossRef]
- Keitel, W.A.; Treanor, J.J.; El Sahly, H.M.; Gilbert, A.; Meyer, A.L.; Patriarca, P.A.; Cox, M.M. Comparative immunogenicity of recombinant influenza hemagglutinin (rHA) and trivalent inactivated vaccine (TIV) among persons ≥65 years old. Vaccine 2009, 28, 379–385. [Google Scholar] [CrossRef]
- Cowling, B.J.; Perera, R.A.P.M.; Valkenburg, S.A.; Leung, N.H.L.; Iuliano, A.D.; Tam, Y.H.; Wong, J.H.F.; Fang, V.J.; Li, A.P.Y.; So, H.C.; et al. Comparative immunogenicity of several enhanced influenza vaccine options for older adults: A randomized, controlled trial. Clin. Infect. Dis. 2020, 71, 1704–1714. [Google Scholar] [CrossRef]
- Talbot, H.K.; Nian, H.; Zhu, Y.; Chen, Q.; Williams, J.V.; Griffin, M.R. Clinical effectiveness of split-virion versus subunit trivalent influenza vaccines in older adults. Clin. Infect. Dis. 2015, 60, 1170–1175. [Google Scholar] [CrossRef]
- Berild, J.D.; Winje, B.A.; Vestrheim, D.F.; Slotved, H.-C.; Valentiner-Branth, P.; Roth, A.; Storsäter, J. A systematic review of studies published between 2016 and 2019 on the effectiveness and efficacy of pneumococcal vaccination on pneumonia and invasive pneumococcal disease in an elderly population. Pathogens 2020, 9, 259. [Google Scholar] [CrossRef]
- Djennad, A.; Ramsay, M.E.; Pebody, R.; Fry, N.K.; Sheppard, C.; Ladhani, S.N.; Andrews, N.J. Effectiveness of 23-valent polysaccharide pneumococcal vaccine and changes in invasive pneumococcal disease incidence from 2000 to 2017 in those aged 65 and over in England and Wales. EClinicalMedicine 2018, 6, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Rappuoli, R.; De Gregorio, E.; Costantino, P. On the mechanisms of conjugate vaccines. Proc. Natl. Acad. Sci. USA 2019, 116, 14–16. [Google Scholar] [CrossRef] [PubMed]
- de Roux, A.; Schmöle-Thoma, B.; Siber, G.R.; Hackell, J.G.; Kuhnke, A.; Ahlers, N.; Baker, S.A.; Razmpour, A.; Emini, E.A.; Fernsten, P.D.; et al. Comparison of pneumococcal conjugate polysaccharide and free polysaccharide vaccines in elderly adults: Conjugate vaccine elicits improved antibacterial immune responses and immunological memory. Clin. Infect. Dis. 2008, 46, 1015–1023. [Google Scholar] [CrossRef] [PubMed]
- Bonten, M.J.M.; Huijts, S.M.; Bolkenbaas, M.; Webber, C.; Patterson, S.; Gault, S.; van Werkhoven, C.H.; van Deursen, A.M.M.; Sanders, E.A.M.; Verheij, T.J.M.; et al. Polysaccharide conjugate vaccine against pneumococcal pneumonia in adults. N. Engl. J. Med. 2015, 372, 1114–1125. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Pneumococcal Vaccination: Who and When to Vaccinate; Centers for Disease Control and Prevention (CDC): Atlanta, GA, USA, 2022.
- Kudlay, D.; Svistunov, A. COVID-19 vaccines: An overview of different platforms. Bioengineering 2022, 9, 72. [Google Scholar] [CrossRef]
- Thi Nhu Thao, T.; Labroussaa, F.; Ebert, N.; V’kovski, P.; Stalder, H.; Portmann, J.; Kelly, J.; Steiner, S.; Holwerda, M.; Kratzel, A.; et al. Rapid reconstruction of SARS-CoV-2 using a synthetic genomics platform. Nature 2020, 582, 561–565. [Google Scholar] [CrossRef]
- Soegiarto, G.; Wulandari, L.; Purnomosari, D.; Fahmita, K.D.; Gautama, H.I.; Hadmoko, S.T.; Prasetyo, M.E.; Mahdi, B.A.; Arafah, N.; Prasetyaningtyas, D.; et al. Hypertension is associated with antibody response and breakthrough infection in health care workers following vaccination with inactivated SARS-CoV-2. Vaccine 2022, 40, 4046–4056. [Google Scholar] [CrossRef]
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning immune humoral response to BNT162b2 COVID-19 vaccine over 6 months. N. Engl. J. Med. 2021, 385, e84. [Google Scholar] [CrossRef]
- Feikin, D.R.; Higdon, M.M.; Abu-Raddad, L.J.; Andrews, N.; Araos, R.; Goldberg, Y.; Groome, M.J.; Huppert, A.; O’Brien, K.L.; Smith, P.G.; et al. Duration of effectiveness of vaccines against SARS-CoV-2 infection and COVID-19 disease: Results of a systematic review and meta-regression. Lancet 2022, 399, 924–944. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; COVID-19 Genomics UK (COG-UK) Consortium; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Ao, D.; Lan, T.; He, X.; Liu, J.; Chen, L.; Baptista-Hon, D.T.; Zhang, K.; Wei, X. SARS-CoV-2 Omicron variant: Immune escape and vaccine development. MedComm 2022, 3, e126. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Hillus, H.; Schwarz, T.; Tober-Lau, P.; Vanshylla, K.; Hastor, H.; Thibeault, C.; Jentzsch, S.; Helbig, E.T.; Lippert, L.J.; Tscheak, P.; et al. Safety, reactogenicity, and immunogenicity of homologous and heterologous prime-boost immunisation with ChAdOx1 nCoV-19 and BNT162b2: A prospective cohort study. Lancet Respir. Med. 2021, 9, 1255–1265. [Google Scholar] [CrossRef]
- Agrati, C.; Capone, S.; Castilletti, C.; Cimini, E.; Matusali, G.; Meschi, S.; Tartaglia, E.; Camerini, R.; Lanini, S.; Milleri, S.; et al. Strong immunogenicity of heterologous prime-boost immunizations with the experimental vaccine GRAd-COV2 and BNT162b2 or ChAdOx1-nCOV19. NPJ Vaccines 2021, 6, 131. [Google Scholar] [CrossRef]
- Sablerolles, R.S.G.; Rietdijk, W.J.R.; Goorhuis, A.; Postma, D.F.; Visser, L.G.; Geers, D.; Schmitz, K.S.; Garcia Garrido, H.M.; Koopmans, M.P.G.; Dalm, V.A.S.H.; et al. for the SWITCH Research Group. Immunogenicity and reactogenicity of vaccine boosters after Ad26.COV2.S priming. N. Engl. J. Med. 2022, 386, 951–963. [Google Scholar] [CrossRef]
- Heinz, F.X.; Stiasny, K. Distinguishing features of current COVID-19 vaccines: Knowns and unknowns of antigen presentation and modes of action. NPJ Vaccines 2021, 6, 104. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, L.; Tian, T.; Li, W.; Pan, Y.; Wang, Y. Efficacy and safety of COVID-19 vaccination in older adults: A systematic review and meta-analysis. Vaccines 2023, 11, 33. [Google Scholar] [CrossRef]
- Chi, W.Y.; Li, Y.D.; Huang, H.C.; Chan, T.E.H.; Chow, S.Y.; Su, J.H.; Ferrall, L.; Hung, C.-F.; Wu, T.-C. COVID-19 vaccine update: Vaccine effectiveness, SARS-CoV-2 variants, boosters, adverse effects, and immune correlates of protection. J Biomed. Sci. 2022, 29, 82. [Google Scholar] [CrossRef]
- James, S.F.; Chahine, E.B.; Sucher, A.J.; Hanna, C. Shingrix: The new adjuvanted recombinant herpes zoster vaccine. Ann. Pharmacother. 2018, 52, 673–680. [Google Scholar] [CrossRef]
- Lacaille-Dubois, M.-A. Updated insights into the mechanism of action and clinical profile of the immunoadjuvant QS-21: A review. Phytomedicine 2019, 60, 152905. [Google Scholar] [CrossRef]
- Lal, H.; Cunningham, A.L.; Godeaux, O.; Chlibek, R.; Diez-Domingo, J.; Hwang, S.-J.; Levin, M.J.; McElhaney, J.E.; Poder, A.; Puig-Barberà, J.; et al. Efficacy of an adjuvanted herpes zoster subunit vaccine in older adults. N. Engl. J. Med. 2015, 372, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.L.; Lal, H.; Kovac, M.; Chlibek, R.; Hwang, S.-J.; Díez-Domingo, J.; Godeaux, O.; Levin, M.J.; McElhaney, J.E.; Puig-Barberà, J.; et al. Efficacy of the herpes zoster subunit vaccine in adults 70 years of age or older. N. Engl. J. Med. 2016, 375, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.L.; Heineman, T.C.; Lal, H.; Godeaux, O.; Chlibek, R.; Hwang, S.-J.; McElhaney, J.E.; Vesikari, T.; Andrews, C.; Choi, W.S.; et al. Immune responses to a recombinant glycoprotein E herpes zoster vaccine in adults aged 50 years or older. J. Infect. Dis. 2018, 217, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Tricco, A.C.; Zarin, W.; Cardoso, R.; Veroniki, A.-A.; Khan, P.A.; Nincic, V.; Ghassemi, M.; Warren, R.; Sharpe, J.P.; Page, A.V.; et al. Efficacy, effectiveness, and safety of herpes zoster vaccines in adults aged 50 and older: Systematic review and network meta-analysis. BMJ 2018, 363, k4029. [Google Scholar] [CrossRef]
- Berkowitz, E.M.; Moyle, G.; Stellbrink, H.-J.; Schürmann, D.; Kegg, S.; Stoll, M.; El Idrissi, M.; Oostvogels, L.; Heineman, T.C.; Zoster-015 HZ/su Study Group. Safety and immunogenicity of an adjuvanted herpes zoster subunit candidate vaccine in HIV-infected adults: A phase 1/2a randomized, placebo-controlled study. J. Infect. Dis. 2015, 211, 1279–1287. [Google Scholar] [CrossRef]
- Curran, D.; Matthews, S.; Rowley, S.D.; Young, J.-A.H.; Bastidas, A.; Anagnostopoulos, A.; Barista, I.; Chandrasekar, P.H.; Dickinson, M.; El Idrissi, M.; et al. Recombinant zoster vaccine significantly reduces the impact on quality of life caused by herpes zoster in adult autologous hematopoietic stem cell transplant recipients: A randomized placebo-controlled trial (ZOE-HSCT). Biol. Blood Marrow Transpl. 2019, 25, 2474–2481. [Google Scholar] [CrossRef]
- Weinberger, B.; Schirmer, M.; Matteucci Gothe, R.; Siebert, U.; Fuchs, D.; Grubeck-Loebenstein, B. Recall responses to tetanus and diphtheria vaccination are frequently insufficient in elderly persons. PLoS ONE 2013, 8, e82967. [Google Scholar] [CrossRef]
- McQuillan, G.M.; Kruszon-Moran, D.; Deforest, A.; Chu, S.Y.; Wharton, M. Serologic immunity to diphtheria and tetanus in the United States. Ann. Intern. Med. 2002, 136, 660–666. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Diphtheria, Tetanus, and Pertussis Vaccine Recommendations; Centers for Disease Control and Prevention (CDC): Atlanta, GA, USA, 2022.
- Torres-Flores, J.M.; Reyes-Sandoval, A.; Salazar, M.I. Dengue vaccines: An update. BioDrugs 2022, 36, 325–336. [Google Scholar] [CrossRef]
- Willyard, C. The slow roll-out of the world’s first malaria vaccine. Nature 2022, 612, S48–S49. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soegiarto, G.; Purnomosari, D. Challenges in the Vaccination of the Elderly and Strategies for Improvement. Pathophysiology 2023, 30, 155-173. https://doi.org/10.3390/pathophysiology30020014
Soegiarto G, Purnomosari D. Challenges in the Vaccination of the Elderly and Strategies for Improvement. Pathophysiology. 2023; 30(2):155-173. https://doi.org/10.3390/pathophysiology30020014
Chicago/Turabian StyleSoegiarto, Gatot, and Dewajani Purnomosari. 2023. "Challenges in the Vaccination of the Elderly and Strategies for Improvement" Pathophysiology 30, no. 2: 155-173. https://doi.org/10.3390/pathophysiology30020014