

Water Quality, Heavy Metals, and Antifungal Susceptibility to Fluconazole of Yeasts from Water Systems

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
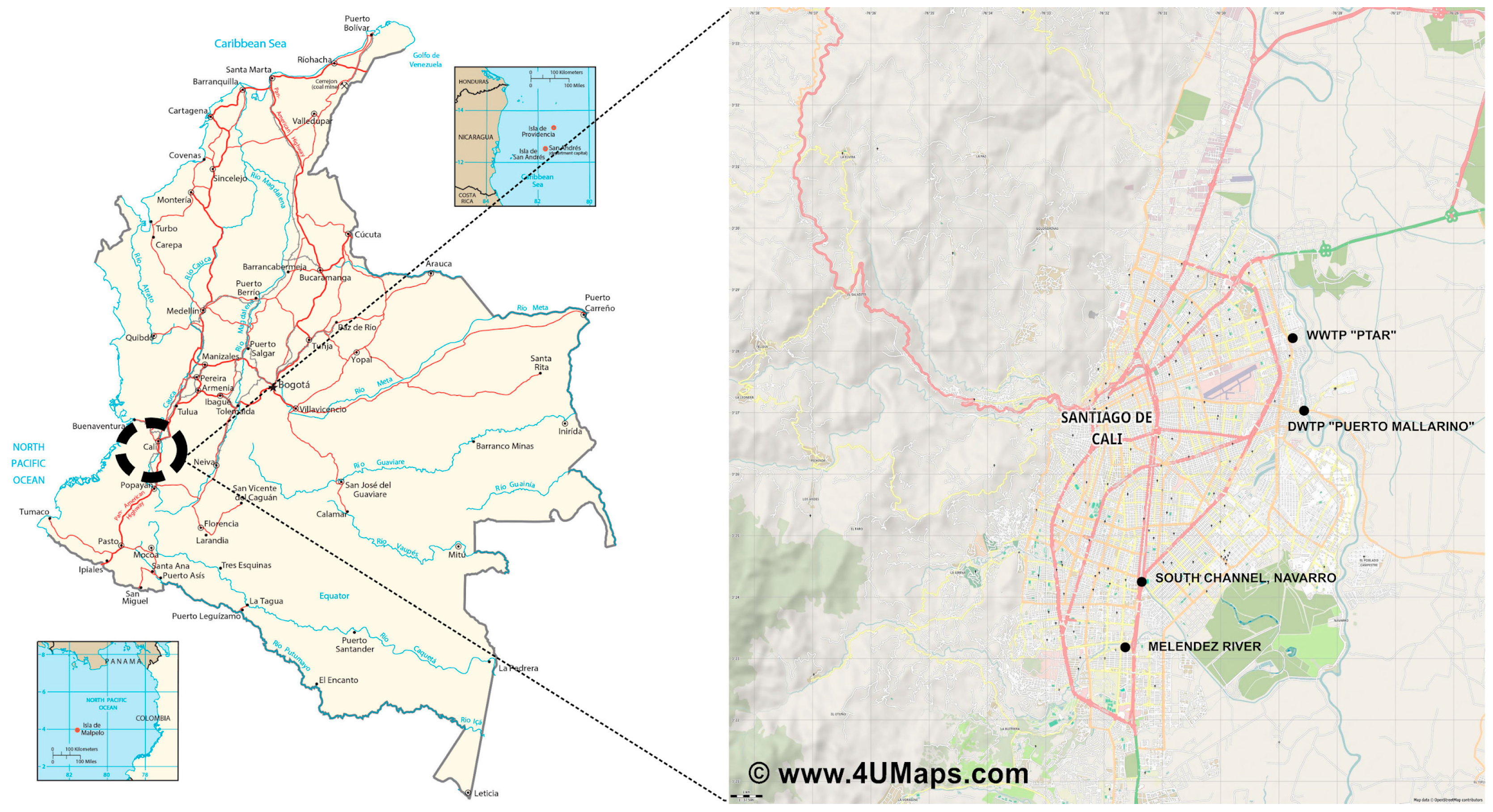
2.1. Study Area and Sampling
2.2. Yeast Isolation and API20C Tests
2.3. Molecular Identification
2.4. Antifungal Susceptibility Tests to Fluconazole and Amphotericin B
2.5. Analysis of Physico-Chemical Parameters and Heavy Metals
2.6. Statistical Analysis
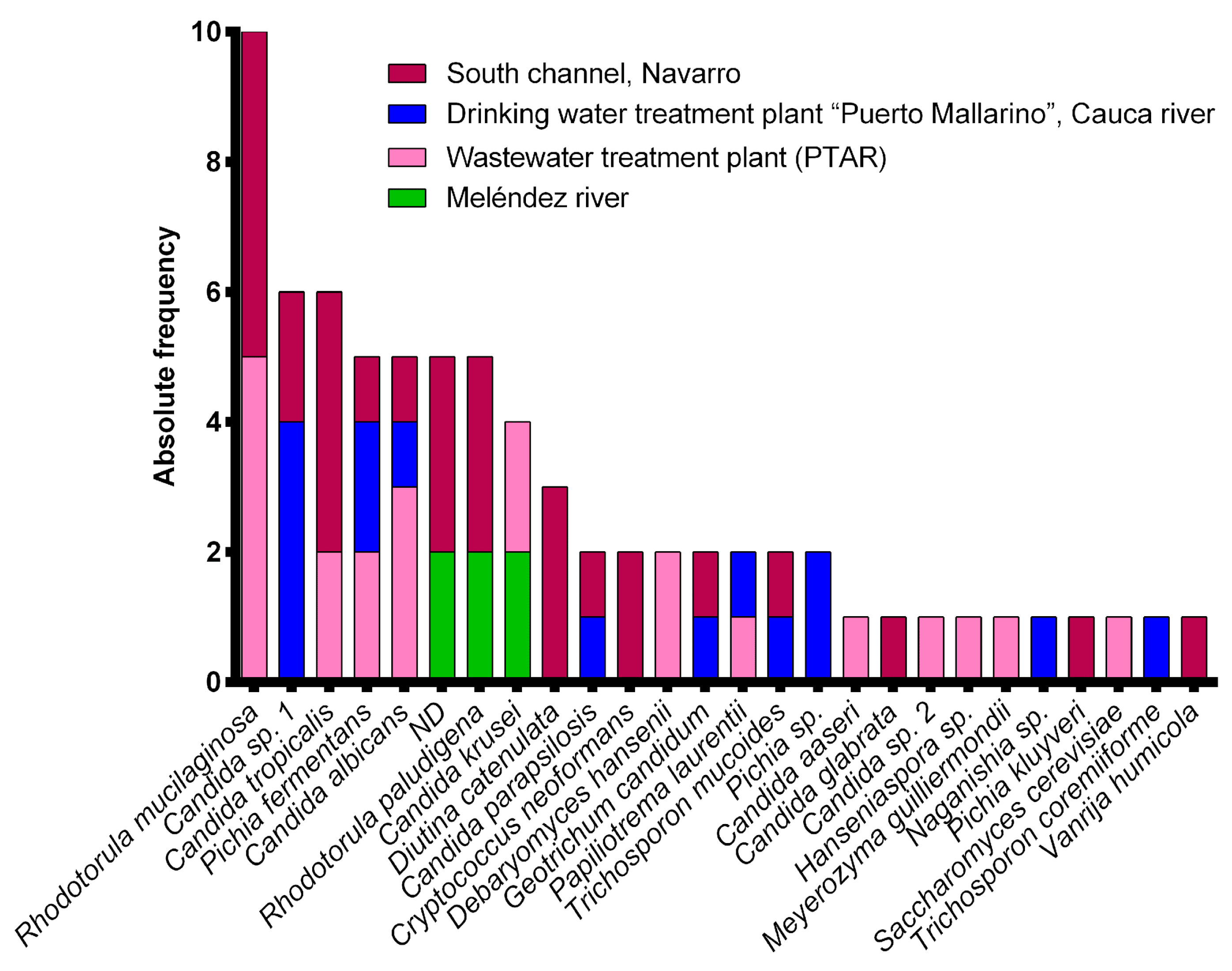
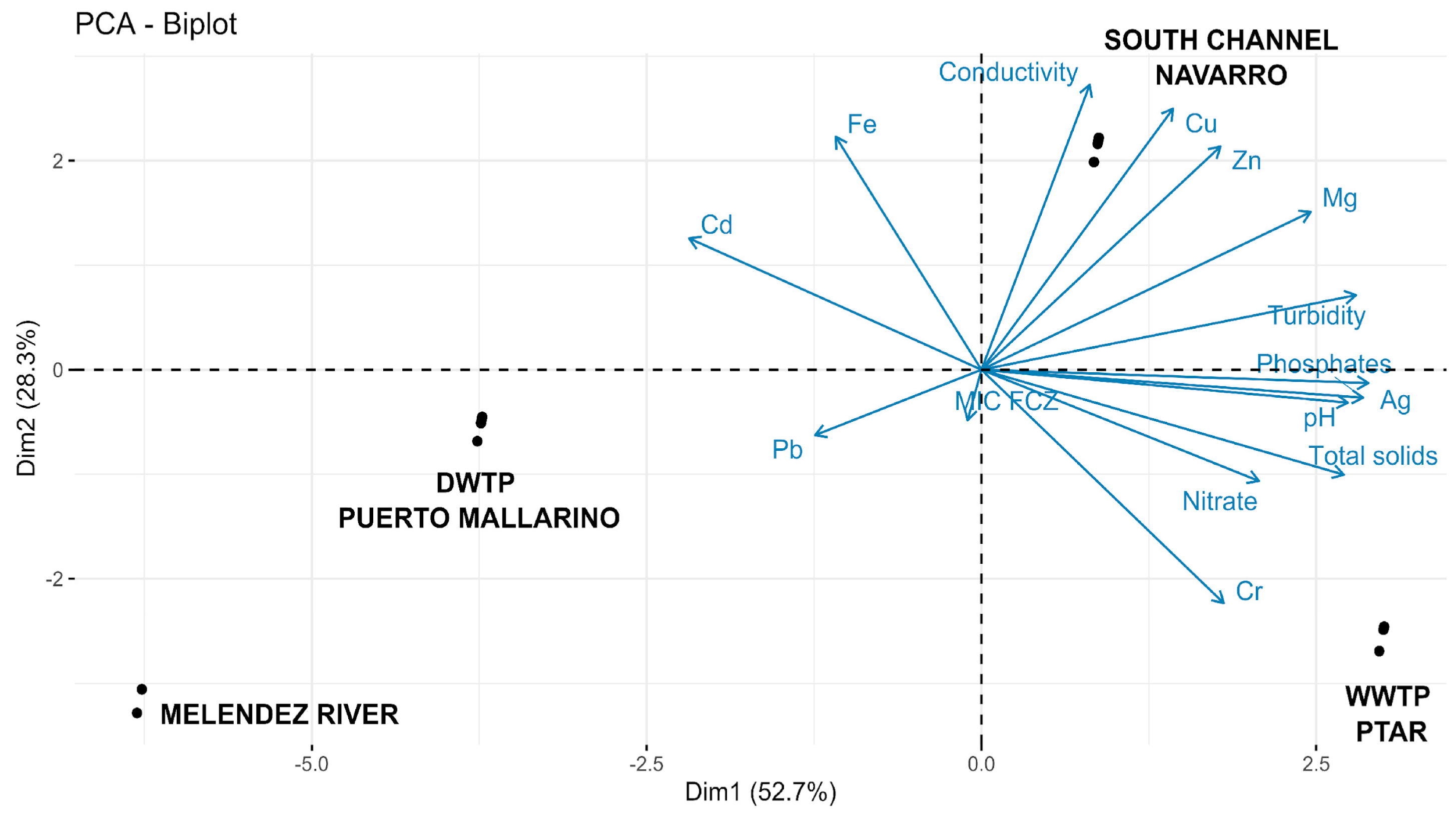
3. Results
4. Discussion
5. Study Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| WWTP | Wastewater Treatment Plant |
| DWTP | Drinking Water Treatment Plant |
| PTAR | Planta de Tratamiento de Aguas Residuales |
| ITS | Internal Transcribed Spacer 1, gene rDNA 5.8S and Internal Transcribed Spacer 2 |
| LSU | D1/D2 Domain of the Large Subunit of the Ribosome 26S |
| CFU | Colony Forming Units |
| MIC | Minimal Inhibitory Concentration |
| FCZ | Fluconazole |
| AMPB | Amphotericin B |
| PCA | Principal Component Analysis |
References
- Monapathi, M.; Bezuidenhout, C.; Rhode, O. Physico-chemical parameters and culturable yeast diversity in surface water: A consequence of pollution. Water SA 2020, 46, 593–601. [Google Scholar] [CrossRef]
- Yurkov, A.; Pozo, M.I. Yeasts community composition and structure. In Yeasts in Natural Ecosystems: Ecology; Buzzini, P., Lachance, M.-A., Yurkov, A., Eds.; Springer Nature: Cham, Switzerland, 2017; pp. 73–100. [Google Scholar]
- Silva-Bedoya, L.M.; Ramírez-Castrillón, M.; Osorio-Cadavid, E. Yeast diversity associated to sediments and water from two colombian artificial lakes. Braz. J. Microbiol. 2014, 45, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milanezi, A.C.M.; Witusk, J.P.D.; van der Sand, S.T. Antifungal susceptibility of yeasts isolated from anthropogenic watershed. An. Acad. Bras. Cienc. 2019, 91, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, N.H.; Reinhard, M.; Gin, K.Y. Occurrence and fate of emerging contaminants in municipal wastewater treatment plants from different geographical regions—A review. Water Res. 2018, 133, 182–207. [Google Scholar] [CrossRef] [PubMed]
- Pagani, D.M.; Heidrich, D.; Tormente, F.; Milani, G.; Jank, L.; Haig, N.H.; Valente, P.; Scroferneker, M.L. High MICs for antifungal agents in yeasts from an anthropized lagoon in South America. Microbiol. Res. 2022, 262, 127083. [Google Scholar] [CrossRef]
- Monapathi, M.; Horn, S.; Vogt, T.; van Wyk, D.; Mienie, C.; Ezeokoli, O.T.; Coertze, R.; Rhode, O.; Bezuidenhout, C.C. Antifungal agents, yeast abundance and diversity in surface water: Potential risks to water users. Chemosphere 2021, 274, 129718. [Google Scholar] [CrossRef]
- Brandao, L.R.; Medeiros, A.O.; Duarte, M.C.; Barbosa, A.C.; Rosa, C.A. Diversity and antifungal susceptibility of yeasts isolated by multiple-tube fermentation from three freshwater lakes in Brazil. J. Water Health 2010, 8, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, A.O.; Missagia, B.S.; Brandão, L.R.; Callisto, M.; Barbosa, F.A.R.; Rosa, C.A. Water quality and diversity of teasts from tropical lakes and rivers from the Rio Doce Basin in Southeastern Brazil. Braz. J. Microbiol. 2012, 43, 1582–1594. [Google Scholar] [CrossRef] [Green Version]
- Monapathi, M.E.; Bezuidenhout, C.C.; Rhode, O.H.J. Water Quality and Antifungal susceptibility of opportunistic yeast pathogens from rivers. Water Sci. Technol. 2017, 75, 1319–1331. [Google Scholar] [CrossRef]
- Van Wyk, D.A.B.; Bezuidenhout, C.C.; Rhode, O.H.J. Diversity and characteristics of yeasts from water sources in the North West Province, South Africa. Water Sci. Technol. 2012, 12, 422–430. [Google Scholar] [CrossRef]
- Assress, H.A.; Selvarajan, R.; Nyoni, H.; Ntushelo, K.; Mamba, B.B.; Msagati, T.A.M. Diversity, Co-occurrence and implications of fungal communities in wastewater treatment plants. Sci. Rep. 2019, 9, 14056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nucci, M.; Queiroz-Telles, F.; Alvarado-Matute, T.; Tiraboschi, I.N.; Cortes, J.; Zurita, J.; Guzman-Blanco, M.; Santolaya, M.E.; Thompson, L.; Sifuentes-Osornio, J.; et al. Epidemiology of candidemia in Latin America: A laboratory-based survey. PLoS ONE 2013, 8, e59373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelo-Branco, D.; Lockhart, S.R.; Chen, Y.; Santos, D.A.; Hagen, F.; Hawkins, N.J.; Lavergne, R.; Meis, J.F.; Pape, L.P.; Rocha, M.F.G.; et al. Collateral consequences of agricultural fungicides on pathogenic yeasts: A one health perspective to tackle azole resistance. Mycoses 2022, 65, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Spader, T.B.; Ramírez-Castrillón, M.; Valente, P.; Alves, S.H.; Severo, L.C. In vitro interactions of amphotericin B combined with non-antifungal agents against Rhodotorula mucilaginosa Strains. Mycopathologia 2019, 184, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Desnos-Ollivier, M.; Lortholary, O.; Bretagne, S.; Dromer, F. Azole susceptibility profiles of more than 9000 clinical yeast isolates belonging to 40 common and rare species. Antimicrob. Agents Chemother. 2021, 65, e02615-20. [Google Scholar] [CrossRef]
- Polvi, E.J.; Li, X.; O´Meara, T.R.; Leach, M.D.; Cowen, L.E. Opportunistic yeast pathogens: Reservoirs, virulence mechanisms, and therapeutic strategies. Cell. Mol. Life Sci. 2015, 72, 2261–2287. [Google Scholar] [CrossRef]
- López-Ávila, K.; Dzul-Rosado, K.R.; Lugo-Caballero, C.; Arias-León, J.J.; Zavala-Castro, J.E. Mecanismos de resistencia antifúngica de los azoles en Candida albicans. Una revisión. Rev. Bioméd. 2016, 27, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Castro, L.Á.; Álvarez, M.I.; Martínez, E. Candida en la cavidad oral de pacientes con VIH en Cali, Colombia: Determinación de especies y sensibilidad al fluconazol. Iatreia 2015, 28, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Demirci-Duarte, S.; Arikan-Akdagli, S.; Gülmez, D. Species distribution, azole resistance and related molecular mechanisms in invasive Candida parapsilosis complex isolates: Increase in fluconazole resistance in 21 years. Mycoses 2021, 64, 823–830. [Google Scholar] [CrossRef]
- Alvarez-Moreno, C.A.; Cortes, J.A.; Denning, D.W. Burden of fungal infections in Colombia. J. Fungi 2018, 4, 41. [Google Scholar] [CrossRef]
- Chen, Z.-F.; Ying, G.-G. Occurrence, fate and ecological risk of five typical azole fungicides as therapeutic and personal care products in the environment: A review. Environ. Int. 2015, 84, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Huang, Q.; Zhang, K.; Yu, Y.; Wang, Z.; Wang, C. Distribution, behavior and fate of azole antifungals during mechanical, biological, and chemical treatments in sewage treatment plants in China. Sci. Total Environ. 2012, 426, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Assress, H.A.; Nyoni, H.; Mamba, B.B.; Msagati, T.A.M. Occurrence and risk assessment of azole antifungal drugs in water and wastewater. Ecotoxicol. Environ. Saf. 2020, 187, 109868. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, Z.; Wang, C.; Peng, X. Chiral profiling of azole antifungals in municipal wastewater and recipient rivers of the Pearl River Delta, China. Environ. Sci. Pollut. Res. 2013, 20, 8890–8899. [Google Scholar] [CrossRef] [PubMed]
- Villota, C.S.V.; Vargas, S.A.F.; Martínez, G.C.A.; Caicedo, B.L.D.; Osorio, C.E.; Ramírez-Castrillón, M. Producción de carotenoides en levaduras nativas aisladas de sistemas acuáticos en Cali, Colombia. Rev. Asoc. Colomb. Cien. Biol. 2020, 1, 103–114. [Google Scholar] [CrossRef]
- Uasapud Garcia, J.D. Instructivo de Toma y Preservación de Muestras Sedimentos y Agua Superficial para la Red de Monitoreo de Calidad del IDEAM 2020, 21. Available online: http://sgi.ideam.gov.co (accessed on 9 June 2018).
- Otero Silva, J.; Candido, R.C. Avaliação do sistema API20C AUX na identificação de leveduras de interesse clínico. Rev. Soc. Bras. Med. Trop. 2005, 38, 261–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, D.; Groenewald, M.; Szöke, S.; Cardinali, G.; Eberhardt, U.; Stielow, B.; de Vries, M.; Verkleij, G.J.M.; Crous, P.W.; Boekhout, T.; et al. DNA barcoding analysis of more than 9 000 yeast isolates contributes to quantitative thresholds for yeast species and genera delimitation. Stud. Mycol. 2016, 85, 91–105. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Perfomance standards for antifungal susceptibility testing of yeasts. In CLSI Supplement M27M44S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022; p. 40. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Epidemiological cutoff values for antifungal susceptibility testing. In CLSI Supplement M57S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022; p. 40. [Google Scholar]
- Libkind, D.; Buzzini, P.; Turchetti, B.; Rosa, C.A. Yeasts in continental and seawater. In Yeasts in Natural Ecosystems: Diversity; Buzzini, P., Lachance, M.-A., Yurkov, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–61. ISBN 9783319626833. [Google Scholar]
- Montanari, L.B.; Garcia Sartori, F.; Montrazi Ribeiro, D.B.; Leandro, L.F.; Pires, R.H.; Melhem, M.d.S.C.; de Mello, C.A.; Gomes Martins, C.H. Yeast isolation and identification in water used in a brazilian hemodialysis unit by classic microbiological techniques and Raman spectroscopy. J. Water Health 2018, 16, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Hagler, A.N. Yeasts as indicators of environmental quality. In Biodiversity and Ecophysiology of Yeasts; Péter, G., Rosa, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 515–532. [Google Scholar]
- Ma, X.; Baron, J.L.; Vikram, A.; Stout, J.E.; Bibby, K. Fungal Diversity and presence of potentially pathogenic fungi in a hospital hot water system treated with on-site monochloramine. Water Res. 2015, 71, 197–206. [Google Scholar] [CrossRef]
- Departamento Administrativo de la Función Pública, Colombia. Decreto 1575 de 2007. Sistema para la Protección y Control de la Calidad del Agua para Consumo Humano. Bogota, Colombia. 9 May 2007. Available online: https://www.funcionpublica.gov.co/eva/gestornormativo/norma.php?i=30007 (accessed on 21 January 2023).
- Ministerio de la Protección Social, Colombia; Ministerio de Ambiente, Vivienda y Desarrollo Territorial, Colombia. Resolución 2115 de 2007 Caracteristicas, Instrumentos Basicos y Frecuencias del Sistema de Control y Vigilancia para la Calidad del Agua para Consumo Humano. Bogota, Colombia. 22 June 2007; p. 23. Available online: https://www.minsalud.gov.co/sites/rid/Lists/BibliotecaDigital/RIDE/DE/DIJ/Resolución_2115_de_2007.pdf (accessed on 21 January 2023).
- Kauffmann–Lacroix, C.; Costa, D.; Imbert, C. Fungi, Water Supply and Biofilms. In Fungal Biofilms and Related Infections; Imbert, C., Ed.; Springer: Cham, Switzerland, 2016; Volume 931, pp. 49–61. [Google Scholar]
- Christa Steffen, H.; Smith, K.; van Deventer, C.; Weiskerger, C.; Bosch, C.; Brandão, J.; Wolfaardt, G.; Botha, A. Health risk posed by direct ingestion of yeasts from polluted river water. Water Res. 2023, 231, 119599. [Google Scholar] [CrossRef]
- Sláviková, E.; Vadkertiová, R. Seasonal Occurrence of yeasts and yeast-like organisms in the River Danube. Antonie Van Leeuwenhoek 1997, 72, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Brandão, L.R.; Libkind, D.; Vaz, A.B.M.; Espírito Santo, L.C.; Moliné, M.; de García, V.; van Broock, M.; Rosa, C.A. Yeasts from an oligotrophic lake in Patagonia (Argentina): Diversity, distribution and synthesis of photoprotective compounds and extracellular enzymes. FEMS Microbiol. Ecol. 2011, 76, 1–13. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, J.M.G.C.F. Yeast community survey in the Tagus estuary. FEMS Microbiol. Ecol. 2005, 53, 295–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez Cespedes, L.; Cespedes Fonseca, Y.M.; Rojas Sanchez, Y.V. Rhodotorula an emergent pathogen. J. Exp. Clin. Microbiol. 2022, 6, 51–54. [Google Scholar] [CrossRef]
- Mejía-Arango, M.A.; Santa-Vélez, C.; Manuela, C.-S.; Vélez, L.M.; Colmenares, L.M.; Restrepo-Jaramillo, B.N.; Cardona-Castro, N. Estudio etiológico y epidemiológico de las micosis cutáneas en un laboratorio de referencia-Antioquia-Colombia. Rev. CES Med. 2013, 27, 7–19. [Google Scholar]
- Rathore, S.S.; Sathiyamoorthy, J.; Lalitha, C.; Ramakrishnan, J. A holistic review on Cryptococcus neoformans. Microb. Pathog. 2022, 166, 105521. [Google Scholar] [CrossRef]
- Hamed, S. Isolation and identification of yeasts along wastewater treatment lines at Zagazig plant. Ann. Agric. Sci. Moshtohor 2016, 54, 77–84. [Google Scholar] [CrossRef]
- Rajasingham, R.; Govender, N.P.; Jordan, A.; Loyse, A.; Shroufi, A.; Denning, D.W.; Meya, D.B.; Chiller, T.M.; Boulware, D.R. The global burden of HIV-associated cryptococcal infection in adults in 2020: A modelling analysis. Lancet Infect. Dis. 2022, 22, 1748–1755. [Google Scholar] [CrossRef]
- Melendres, J.; Carvajal-Valencia, S.; Escandon, P.; Firacative, C. S8.5d Cryptococcus neoformans and Cryptococcus gattii clinical isolates from Colombia develop heteroresistance to fluconazole at high concentrations. Med. Mycol. 2022, 60, 2022. [Google Scholar] [CrossRef]
- Cebeci Güler, N.; Tosun, İ.; Aydin, F. The identification of Meyerozyma guilliermondii from blood cultures and surveillance samples in a university hospital in Northeast Turkey: A ten-year survey. J. Mycol. Med. 2017, 27, 506–513. [Google Scholar] [CrossRef]
- Zhang, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Potential biocontrol activity of a strain of Pichia guilliermondii against grey mold of apples and its possible modes of action. Biol. Control 2011, 57, 193–201. [Google Scholar] [CrossRef]
- Bohner, F.; Papp, C.; Gácser, A. The effect of antifungal resistance development on the virulence of Candida species. FEMS Yeast Res. 2022, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tevyashova, A.; Efimova, S.; Alexandrov, A.; Omelchuk, O.; Ghazy, E.; Bychkova, E.; Zatonsky, G.; Grammatikova, N.; Dezhenkova, L.; Solovieva, S.; et al. Semisynthetic amides of amphotericin B and nynstatin A1: A comparative study of in vitro activity/toxicity ratio in relation to selectivity to ergosterol membranes. Antibiotics 2023, 12, 151–170. [Google Scholar] [CrossRef]
- Brilhante, R.S.N.; Paiva, M.A.N.; Sampaio, C.M.S.; Castelo-Branco, D.S.C.M.; Teixeira, C.E.C.; de Alencar, L.P.; Bandeira, T.J.P.G.; Monteiro, A.J.; Cordeiro, R.A.; Pereira-Neto, W.A.; et al. Azole resistance in Candida spp. isolated from Catú Lake, Ceará, Brazil: An efflux-pump-mediated mechanism. Braz. J. Microbiol. 2016, 47, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros, A.O.; Kohler, L.M.; Hamdan, J.S.; Missagia, B.S.; Barbosa, F.A.R.; Rosa, C.A. Diversity and antifungal susceptibility of yeasts from tropical freshwater environments in Southeastern Brazil. Water Res. 2008, 42, 3921–3929. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Z.; Li, J.; Zhu, Z.; Pang, S.; Xu, J.; Wu, J. Extensive diversity and prevalent fluconazole resistance among environmental yeasts from tropical China. Genes 2022, 13, 444–467. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.; Ribeiro, F.C.; Colombo, A.L.; de Almeida, J.N., Jr. The emerging threat antifungal-resistant Candida tropicalis in humans, animals, and environment. Front. Fungal Biol. 2022, 3, 957021. [Google Scholar] [CrossRef]
- Paul, S.; Shaw, D.; Joshi, H.; Singh, S.; Chakrabarti, A.; Rudramurthy, S.M.; Ghosh, A.K. Mechanisms of azole antifungal resistance in clinical isolates of Candida tropicalis. PLoS ONE 2022, 17, e0269721. [Google Scholar] [CrossRef]
- Forastiero, A.; Mesa-Arango, A.C.; Alastruey-Izquierdo, A.; Alcazar-Fuoli, L.; Bernal-Martinez, L.; Pelaez, T.; Lopez, J.F.; Grimalt, J.O.; Gomez-Lopez, A.; Cuesta, I.; et al. Candida tropicalis antifungal cross-resistance is related to different azole target (Erg11p) modifications. Antimicrob. Agents Chemother. 2013, 57, 4769–4781. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro, R.d.A.; de Oliveira, J.S.; Castelo-Branco, S.C.M.; Cordeiro Teixeira, C.E.; de Farias Marques, F.J.; Bittencourt, P.V.; Carvalho, V.L.; Gomes Bandeira, T.d.J.P.; Nogueira Brilhante, R.S.; Bezerra Moreira, J.L.; et al. Candida tropicalis isolates obtained from veterinary sources show resistance to azoles and produce virulence factors. Med. Mycol. 2015, 53, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Subramanya, S.H.; Sharan, N.K.; Baral, B.P.; Hamal, D.; Nayak, N.; Prakash, P.Y.; Sathian, B.; Bairy, I.; Gokhale, S. Diversity, in-vitro virulence traits and antifungal susceptibility pattern of gastrointestinal yeast flora of healthy poultry, Gallus Gallus Domesticus. BMC Microbiol. 2017, 17, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Boguslawska-Was, E.; Dabrowski, W. The seasonal variability of yeasts and yeast-like organisms in water and bottom sediment of the Szczecin lagoon. Int. J. Hyg. Environ. Health 2001, 203, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Pietryczuk, A.; Cudowski, A.; Hauschild, T. Effect of trophic status in lakes on fungal species diversity and abundance. Ecotoxicol. Environ. Saf. 2014, 109, 32–37. [Google Scholar] [CrossRef]
- World Health Organization W.H.O. Fungal Priority Pathogens List to Guide Research, Development and Public Health Action, 1st ed.; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Baffico, G.D. Variations in the periphytic community structure and dynamics of lake Nahuel Huapi (Patagonia, Argentina). Hydrobiologia 2001, 455, 79–85. [Google Scholar] [CrossRef]
- IDEAM Grupo de Climatología y Agrometereología. Mapas de Precipitación Promedio en Colombia. Available online: http://atlas.ideam.gov.co/visorAtlasClimatologico.html (accessed on 21 January 2023).
- Ortiz-Vera, M.P.; Olchanheski, L.R.; da Silva, E.G.; de Lima, F.R.; Rada Martinez, L.R.d.P.; Zanoli Sato, M.I.; Jaffé, R.; Alves, R.; Ichiwaki, S.; Padilla, G.; et al. Influence of water quality on diversity and composition of fungal communities in a tropical river. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Olaniran, A.O.; Balgobind, A.; Pillay, B. Bioavailability of heavy metals in soil: Impact on microbial biodegradation of organic compounds and possible improvement strategies. Int. J. Mol. Sci. 2013, 14, 10197–10228. [Google Scholar] [CrossRef] [Green Version]
- Shukla, P.K.; Misra, P.; Maurice, N.; Ramteke, P.W. Heavy metal toxicity and possible functional aspects of microbial diversity in heavy metal-contaminated sites. In Microbial Genomics in Sustainable Agroecosystems; Tripathi, V., Kumar, P., Tripathi, P., Kishore, A., Kamle, M., Eds.; Springer Nature: Singapore, 2019; pp. 255–317. ISBN 9789813298606. [Google Scholar]
- Varol, M.; Ustaoğlu, F.; Tokatlı, C. Ecological risks and controlling factors of trace elements in sediments of Dam Lakes in the Black Sea region (Turkey). Environ. Res. 2022, 205, 112478. [Google Scholar] [CrossRef]
- Wang, Q.; He, X.; Xiong, H.; Chen, Y.; Huang, L. Structure, Mechanism, and Toxicity in Antibiotics Metal Complexation: Recent Advances and Perspectives. Sci. Total Environ. 2022, 848, 1–14. [Google Scholar] [CrossRef]
- Dickinson, A.W.; Power, A.; Hansen, M.G.; Brandt, K.K.; Piliposian, G.; Appleby, P.; O’Neill, P.A.; Jones, R.T.; Sierocinski, P.; Koskella, B.; et al. Heavy Metal Pollution and Co-Selection for Antibiotic Resistance: A Microbial Palaeontology Approach. Environ. Int. 2019, 132, 105117. [Google Scholar] [CrossRef]
- Gupta, S.K.; Shin, H.; Han, D.; Hur, H.G.; Unno, T. Metagenomic Analysis Reveals the Prevalence and Persistence of Antibiotic- and Heavy Metal-Resistance Genes in Wastewater Treatment Plant. J. Microbiol. 2018, 56, 408–415. [Google Scholar] [CrossRef]
- Jia, J.; Guan, Y.; Cheng, M.; Chen, H.; He, J.; Wang, S.; Wang, Z. Occurrence and Distribution of Antibiotics and Antibiotic Resistance Genes in Ba River, China. Sci. Total Environ. 2018, 642, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Seiler, C.; Berendonk, T.U. Heavy Metal Driven Co-Selection of Antibiotic Resistance in Soil and Water Bodies Impacted by Agriculture and Aquaculture. Front. Microbiol. 2012, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajasekar, A.; Qiu, M.; Wang, B.; Murava, R.T.; Norgbey, E. Relationship between Water Quality, Heavy Metals and Antibiotic Resistance Genes among Three Freshwater Lakes. Environ. Monit. Assess. 2022, 194, 64–73. [Google Scholar] [CrossRef]
- Buechel, E.R.; Pinkett, H.W. Transcription Factors and ABC Transporters: From Pleiotropic Drug Resistance to Cellular Signaling in Yeast. FEBS Lett. 2020, 594, 3943–3964. [Google Scholar] [CrossRef]
- Hamdoun, A.; Hellmich, U.A.; Szakacs, G.; Kuchler, K. The Incredible Diversity of Structures and Functions of ABC Transporters. FEBS Lett. 2021, 595, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Bazzicalupo, A.L.; Ruytinx, J.; Ke, Y.H.; Coninx, L.; Colpaert, J.V.; Nguyen, N.H.; Vilgalys, R.; Branco, S. Fungal Heavy Metal Adaptation through Single Nucleotide Polymorphisms and Copy-Number Variation. Mol. Ecol. 2020, 29, 4157–4169. [Google Scholar] [CrossRef] [PubMed]
- Mi, L.; Wang, Z.; Yang, W.; Huang, C.; Zhou, B.; Hu, Y.; Liu, S. Cytochromes P450 in Biosensing and Biosynthesis Applications: Recent Progress and Future Perspectives. TrAC Trends Anal. Chem. 2023, 158, 116791. [Google Scholar] [CrossRef]
- Dunkel, N.; Blaß, J.; Rogers, P.D.; Morschhäuser, J. Mutations in the Multi-Drug Resistance Regulator MRR1, Followed by Loss of Heterozygosity, Are the Main Cause of MDR1 Overexpression in Fluconazole-Resistant Candida albicans Strains. Mol. Microbiol. 2008, 69, 827–840. [Google Scholar] [CrossRef]
- Ramirez-Castrillon, M.; Barona-Colorado, A.; Bados-Lopez, M.C.; Bolaños-Burbano, D. Chapter 7: Diversity of Environmental Yeasts of Colombia: A Systematic Review. In Catalogue of Fungi of Colombia, 1st ed.; de Almeida, R.F., Lücking, R., Vasco-Palacios, A., Gaya, E., Diazgranados, M., Eds.; Royal Botanic Gardens,: Kew, UK; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2022; pp. 92–103. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Strain | Species | Susceptibility to Fluconazole | FCZ MIC (μg/mL) | AMPB MIC (μg/mL) |
|---|---|---|---|---|
| CS11 | Rh. paludigena | Sensitive | 2 | 2 |
| CS14 | ND | Sensitive | 2 | 2 |
| CS15 | V. humicola | Sensitive | 0.5 | 0.25 |
| CS16 | C. parapsilosis | Sensitive | 2 | 0.5 |
| CS17 | D. catenulata | Sensitive | 0.5 | 1 |
| CS18 | D. catenulata | Sensitive | 0.5 | 0.5 |
| CS19 | C. tropicalis | DDS | 4 | 2 |
| CS1A | C. glabrata | DDS | 4 | 0.5 |
| CS1B | Rh. mucilaginosa | Resistant * | 1 | 2 |
| CS21 | P. fermentans | DDS | 4 | 2 |
| CS22 | ND | Sensitive | 0.25 | 2 |
| CS23 | Cr. neoformans | Sensitive | 0.125 | 2 |
| CS24A | Cr. neoformans | Sensitive | 2 | 1 |
| CS4 | T. mucoides | Sensitive | 2 | 2 |
| CS45 | P. kluyveri | Sensitive | 0.5 | 0.5 |
| CS51A | D. catenulata | Resistant | 8 | 2 |
| CS51B | C. tropicalis | Resistant | 16 | 2 |
| CS55 | C. albicans | Resistant | 64 | 2 |
| CS7 | C. tropicalis | Sensitive | 1 | 2 |
| CS7B | G. candidum | Resistant | 2 | 2 |
| CS7C | ND | Resistant | 64 | 2 |
| CS9 | Candida sp. 1 | Sensitive | 0.25 | 1 |
| M2 | ND | Sensitive | 2 | 2 |
| M12 | ND | Resistant | 64 | 2 |
| P1 | C. albicans | Resistant | 8 | 1 |
| P13B | C. albicans | DDS | 4 | 1 |
| P14 | C. aaseri | Sensitive | 0.5 | 0.5 |
| P16A | C. tropicalis | Sensitive | 2 | 2 |
| P16B | P. laurentii | Sensitive | 0.5 | 2 |
| P20A | D. hansenii | DDS | 4 | 2 |
| P22 | P. kudriavzevii (C. krusei) | Resistant * | 4 | 1 |
| P24A | D. hansenii | Resistant | 64 | 0.5 |
| P24B | Rh. mucilaginosa | Resistant | 64 | 2 |
| P3 | S. cerevisiae | DDS | 4 | 2 |
| P3A | Hanseniaspora pseudoguilliermondii | Sensitive | 1 | 2 |
| P4 | C. tropicalis | Resistant | 64 | 0.5 |
| P46 | Candida sp. 2 | Sensitive | 0.25 | 2 |
| P9A | P. fermentans | Sensitive | 2 | 2 |
| PM14 | G. candidum | Sensitive | 0.5 | 1 |
| PM15 | Pichia sp. | DDS | 4 | 2 |
| PM18 | T. coremiiforme | Sensitive | 2 | 0.5 |
| PM19 | P. fermentans | DDS | 4 | 2 |
| PM20 | Naganishia sp. | Sensitive | 0.125 | 2 |
| PM22 | C. albicans | Sensitive | 0.5 | 2 |
| PM24 | Candida sp. 1 | DDS | 4 | 2 |
| PM4A | T. mucoides | Resistant | 8 | 2 |
| PM4B | P. laurentii | Resistant | 16 | 2 |
| PM54 | Candida sp. 1 | Resistant | 64 | 0.5 |
| PM54A | C. parapsilosis | Resistant | 16 | 2 |
| Heavy Metals/ Parameters | Cp | Ct | Ca | Pk | Rhm | Tm | Gc | Dh | Pl | Dc | Csp1 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fe | 0.469 | 0.090 | 0.251 | 0.192 | −0.816 | 0.543 | 0.352 | −0.135 | 0.406 | 0.816 | 0.333 |
| Ag | 0.020 | 0.404 | 0.249 | 0.095 | 0.000 | −0.089 | −0.027 | 0.456 | −0.188 | −0.816 | 0.333 |
| Zn | 0.506 | 0.269 | 0.470 | 0.095 | −0.816 | −0.134 | 0.027 | 0.026 | 0.295 | 0.816 | −0.333 |
| Cu | 0.506 | 0.000 | 0.249 | 0.190 | −0.816 | −0.134 | 0.188 | −0.134 | 0.188 | −0.816 | −0.666 |
| Cd | −0.292 | −0.324 | 0.805 | 0.024 | 0.816 | 0.208 | 0.000 | −0.194 | −0.388 | −0.816 | 0.912 |
| Cr | −0.179 | 0.173 | 0.000 | 0.079 | 0.500 | −0.198 | −0.208 | 0.505 | −0.148 | −1.000 | −0.182 |
| Pb | −0.066 | 0.172 | 0.331 | 0.239 | 0.000 | 0.172 | 0.147 | 0.000 | −0.147 | 0.000 | 0.333 |
| Mg | 0.303 | 0.224 | 0.359 | 0.190 | −0.816 | −0.044 | 0.080 | 0.080 | 0.134 | 0.000 | −0.333 |
| TP | 0.060 | 0.449 | 0.304 | −0.047 | 0.000 | −0.269 | −0.241 | 0.510 | −0.134 | −0.816 | 0.000 |
| Turbidity | 0.263 | 0.224 | 0.359 | 0.191 | −0.816 | 0.000 | 0.080 | 0.080 | 0.080 | 0.000 | 0.000 |
| TDS | 0.183 | −0.022 | −0.700 | −0.024 | −0.816 | −0.135 | −0.027 | −0.054 | 0.135 | 0.000 | −0.666 |
| Nitrate | −0.060 | −0.045 | −0.415 | −0.429 | 0.816 | −0.674 | −0.403 | 0.134 | −0.188 | −0.816 | −1.000 |
| NT | 0.101 | 0.314 | 0.304 | 0.000 | −0.816 | 0.000 | −0.134 | 0.241 | 0.026 | 0.000 | 0.333 |
| pH | 0.067 | 0.522 | 0.464 | 0.237 | 0.000 | −0.124 | 0.029 | 0.534 | 0.029 | 0.000 | −0.235 |
| COND | 0.604 | 0.074 | 0.402 | 0.342 | −0.816 | 0.322 | 0.386 | −0.178 | 0.326 | 0.000 | −0.235 |
| TEMP | 0.469 | 0.372 | 0.526 | 0.289 | −0.816 | 0.024 | 0.207 | 0.178 | 0.148 | 0.000 | −0.235 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caicedo-Bejarano, L.D.; Osorio-Vanegas, L.S.; Ramírez-Castrillón, M.; Castillo, J.E.; Martínez-Garay, C.A.; Chávez-Vivas, M. Water Quality, Heavy Metals, and Antifungal Susceptibility to Fluconazole of Yeasts from Water Systems. Int. J. Environ. Res. Public Health 2023, 20, 3428. https://doi.org/10.3390/ijerph20043428
Caicedo-Bejarano LD, Osorio-Vanegas LS, Ramírez-Castrillón M, Castillo JE, Martínez-Garay CA, Chávez-Vivas M. Water Quality, Heavy Metals, and Antifungal Susceptibility to Fluconazole of Yeasts from Water Systems. International Journal of Environmental Research and Public Health. 2023; 20(4):3428. https://doi.org/10.3390/ijerph20043428
Chicago/Turabian StyleCaicedo-Bejarano, Luz Dary, Lizeth Stefania Osorio-Vanegas, Mauricio Ramírez-Castrillón, Jorge Enrique Castillo, Carlos Andrés Martínez-Garay, and Mónica Chávez-Vivas. 2023. "Water Quality, Heavy Metals, and Antifungal Susceptibility to Fluconazole of Yeasts from Water Systems" International Journal of Environmental Research and Public Health 20, no. 4: 3428. https://doi.org/10.3390/ijerph20043428