
Complexity of Running and Its Relationship with Joint Kinematics during a Prolonged Run


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Equipment
2.3. Experimental Protocol
2.4. Data Analysis
2.4.1. Calculation of Spatiotemporal and Kinematic Variables
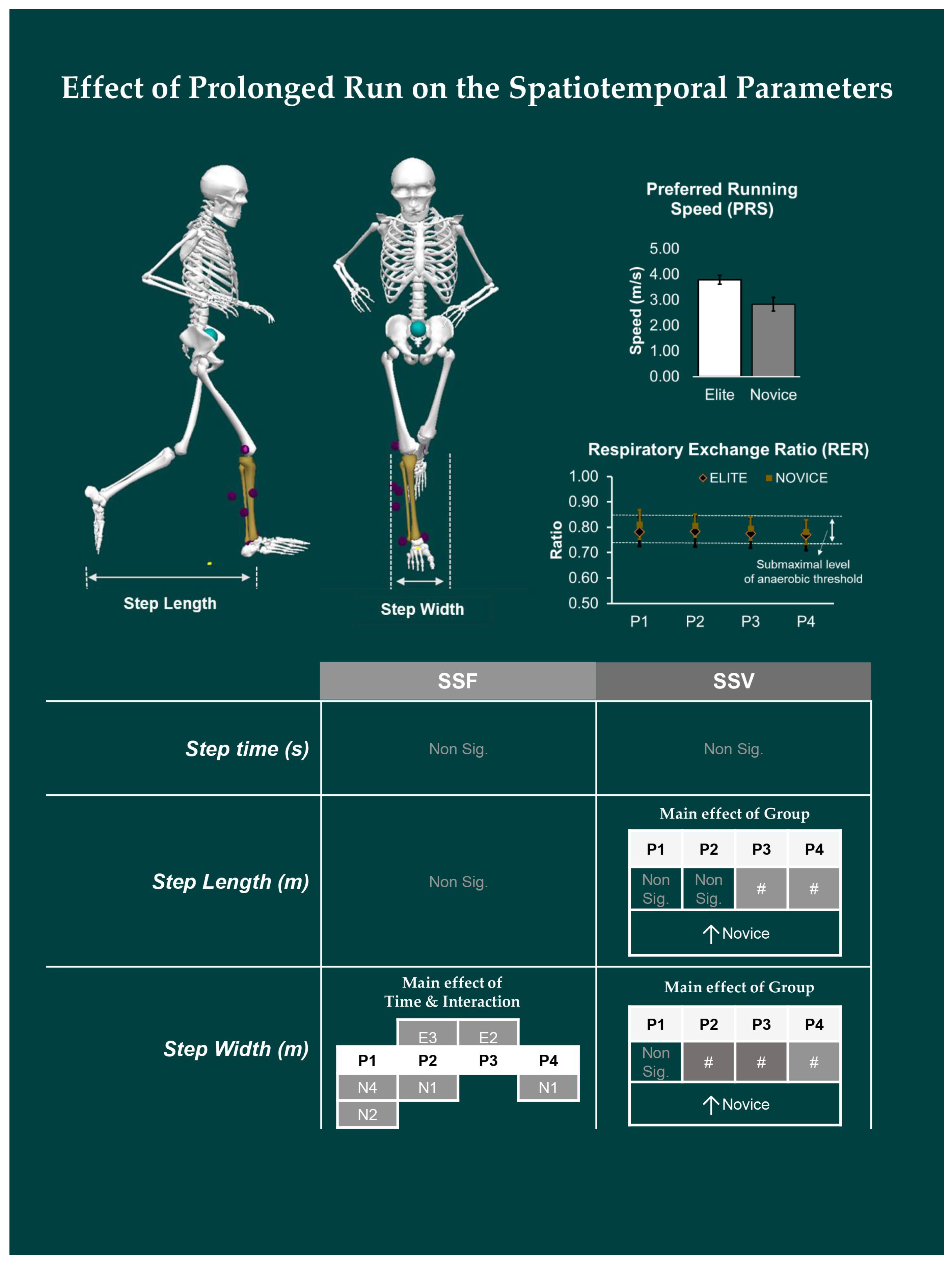
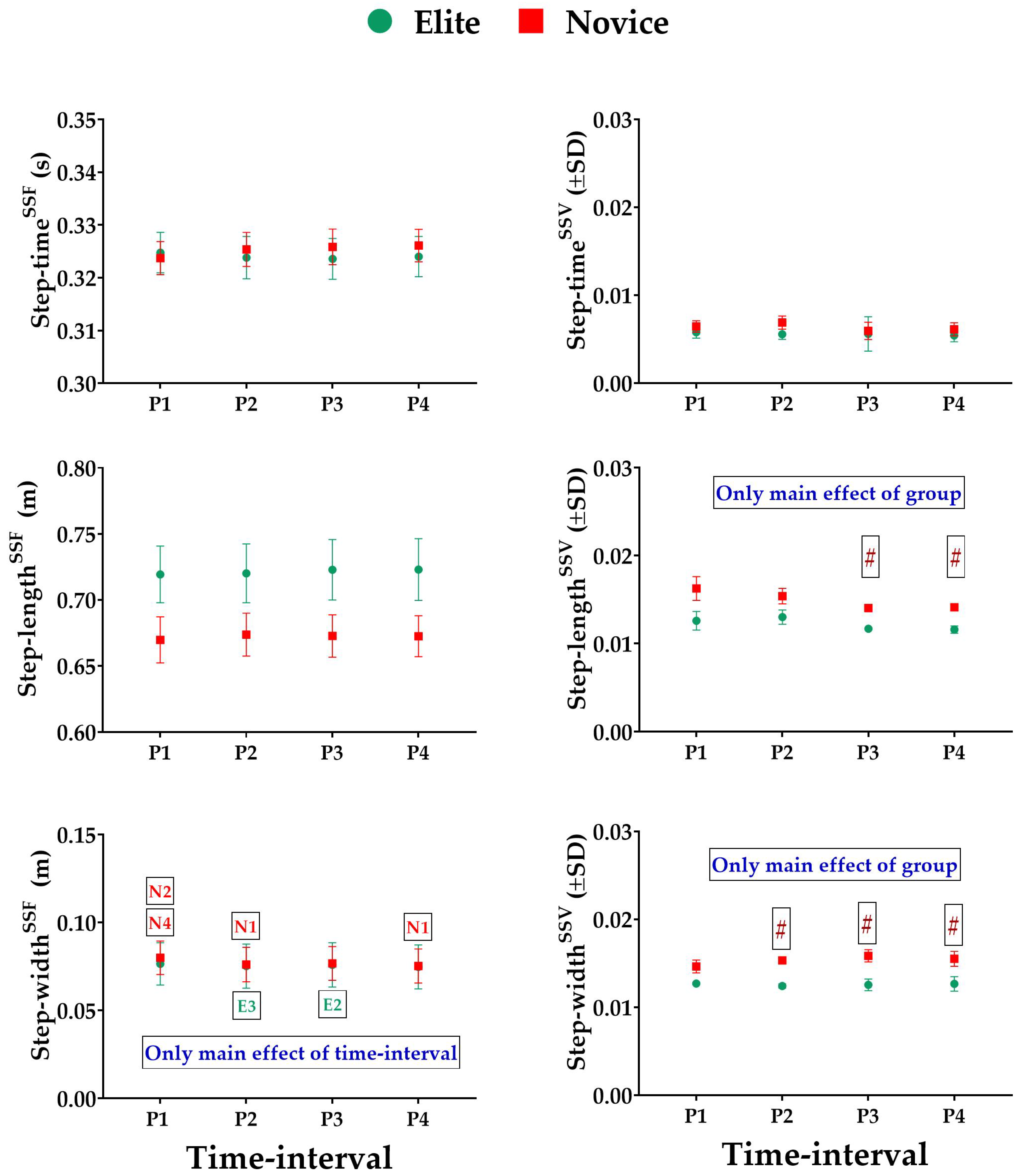
- Spatiotemporal variables: include step time in seconds (s), step length, and step width in centimeters (cm).
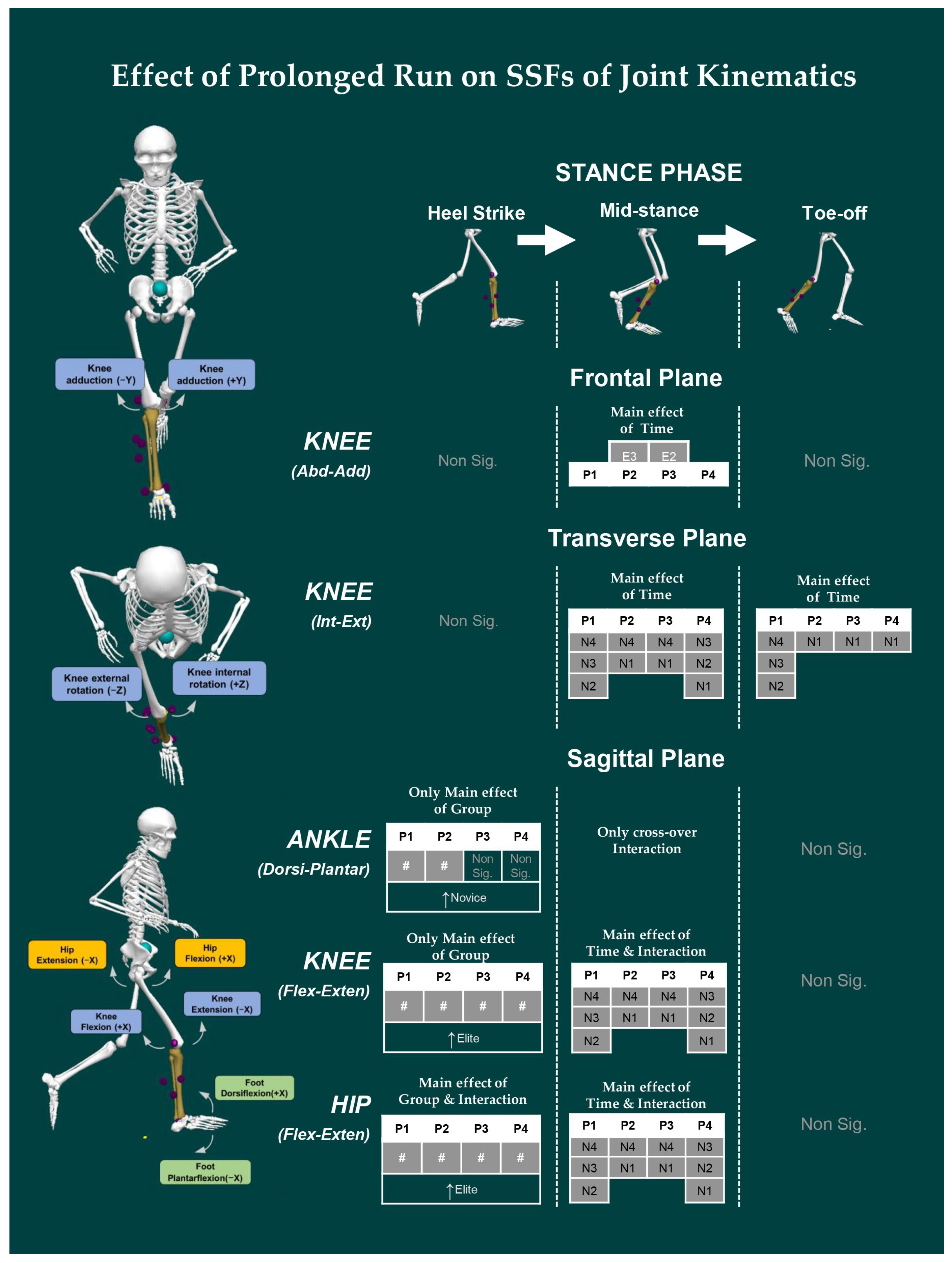
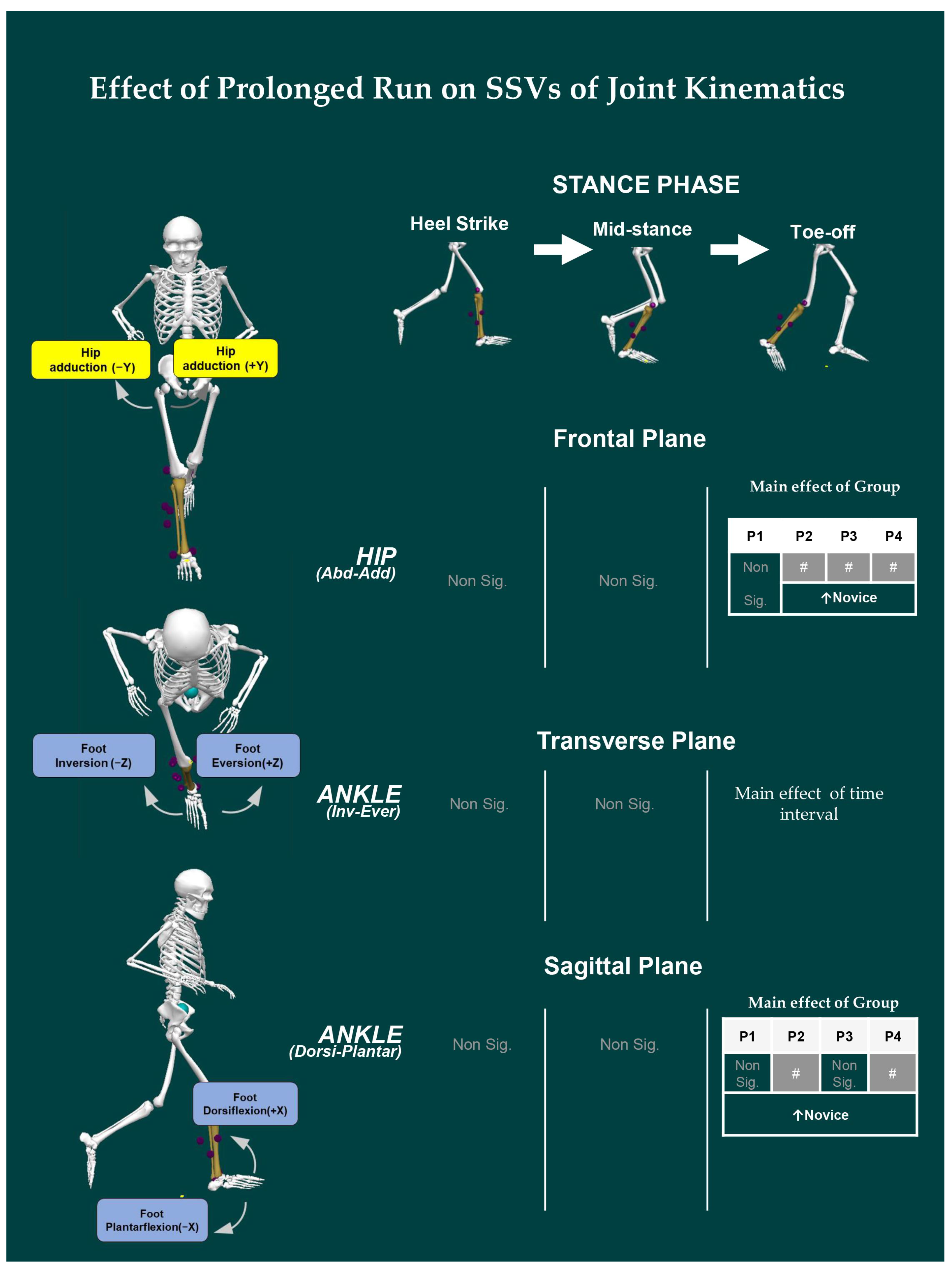
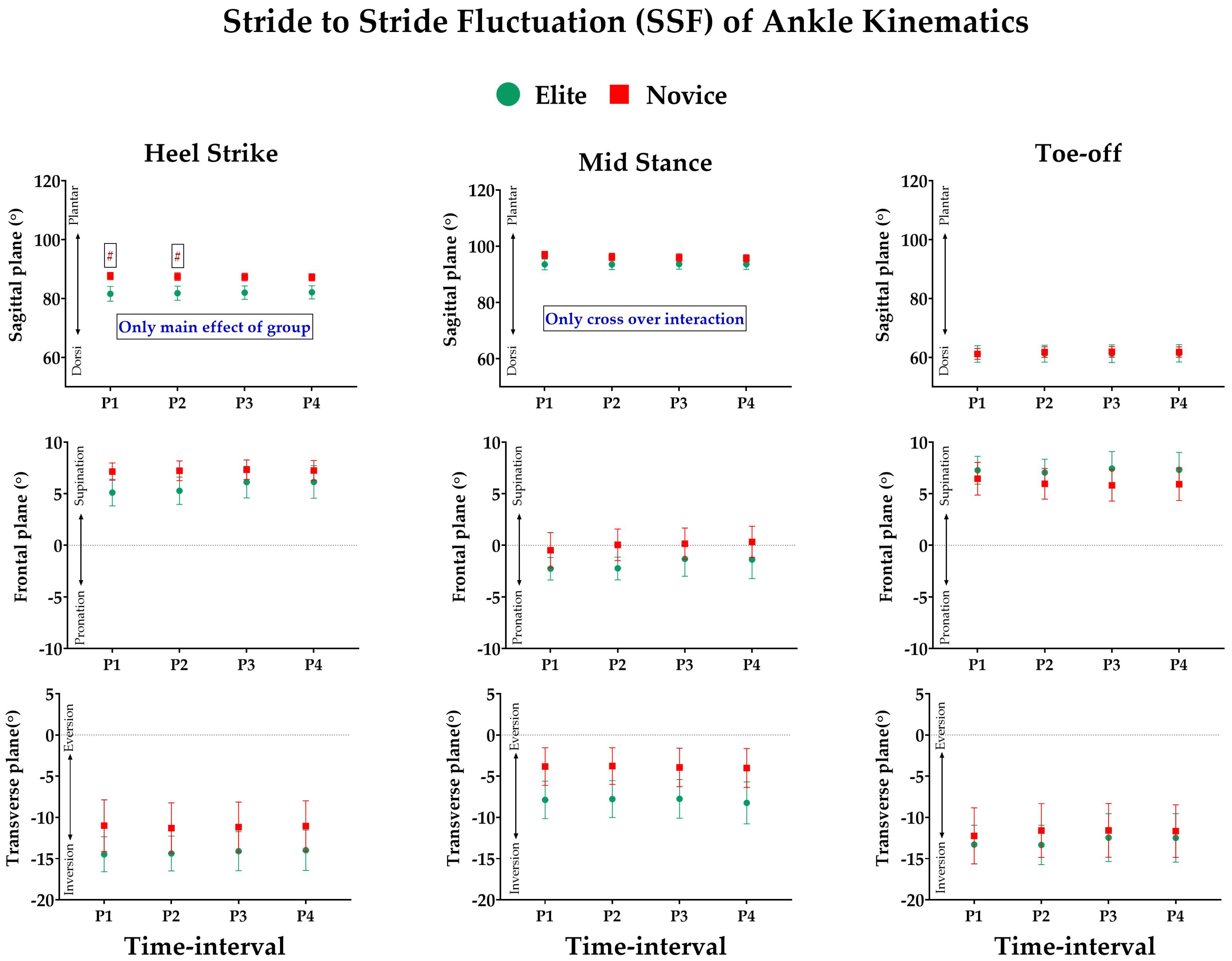
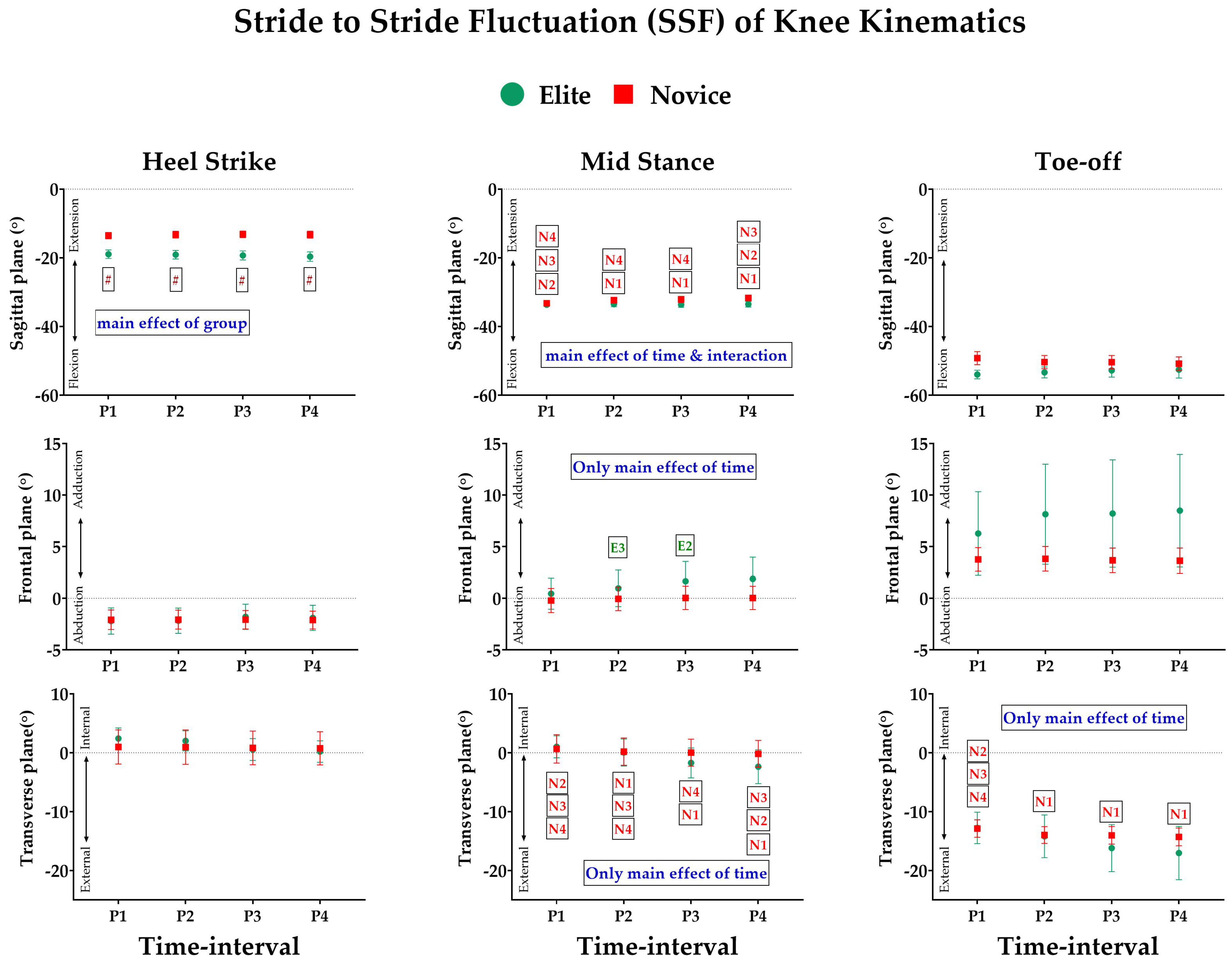
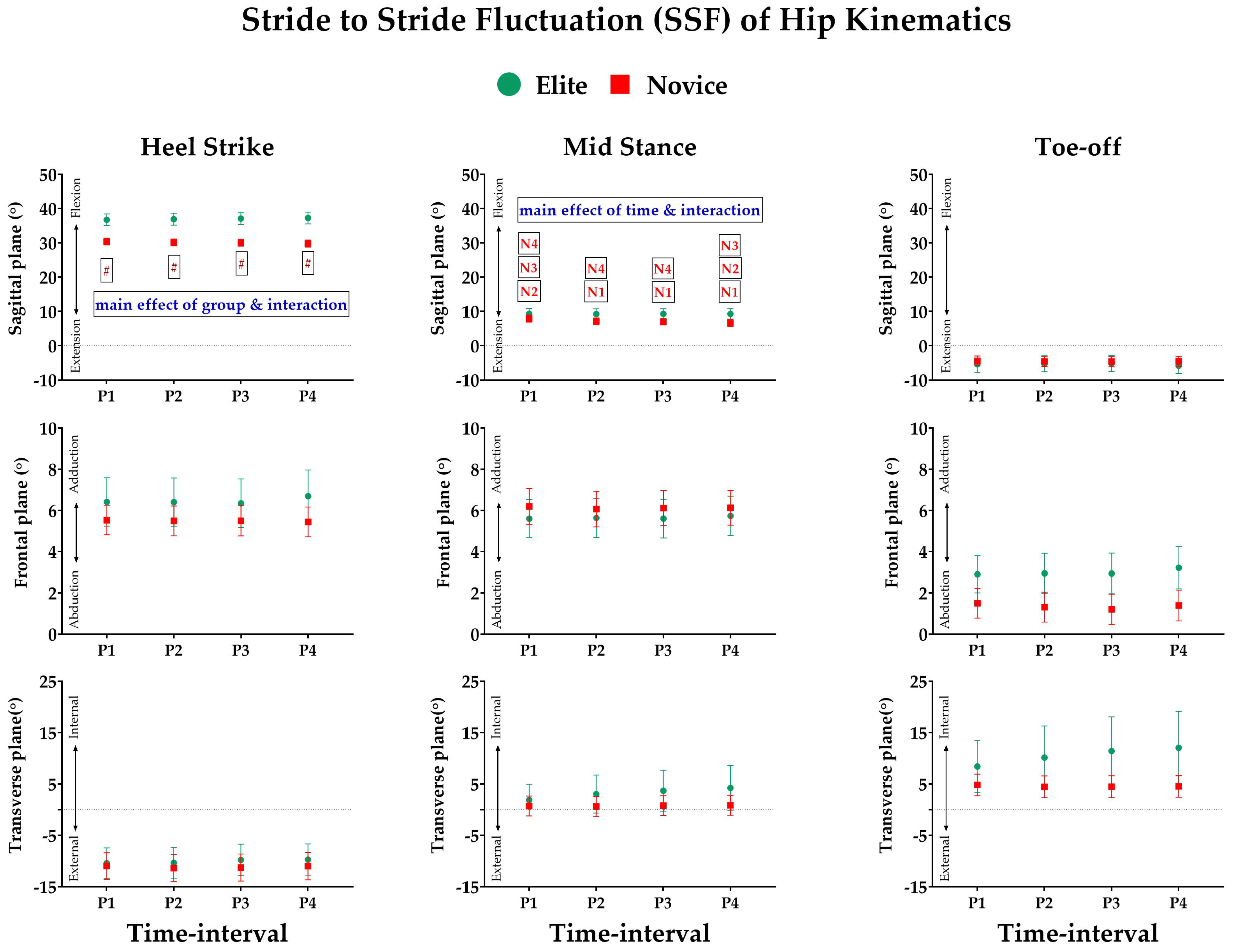
- Joint Kinematics: Similarly, the joint angles of the ankle, knee, and hip were computed based on the Cardan sequence, where the X-, Y-, and Z-axes corresponded to flexion/extension, abduction/adduction, and internal/external rotation angles, respectively. Thereafter, discrete joint angles were assessed, including hip, knee, and ankle angles, in sagittal (X), frontal (Y), and transverse (Z) planes for each event of the stance phase. The units are expressed in degrees (°).
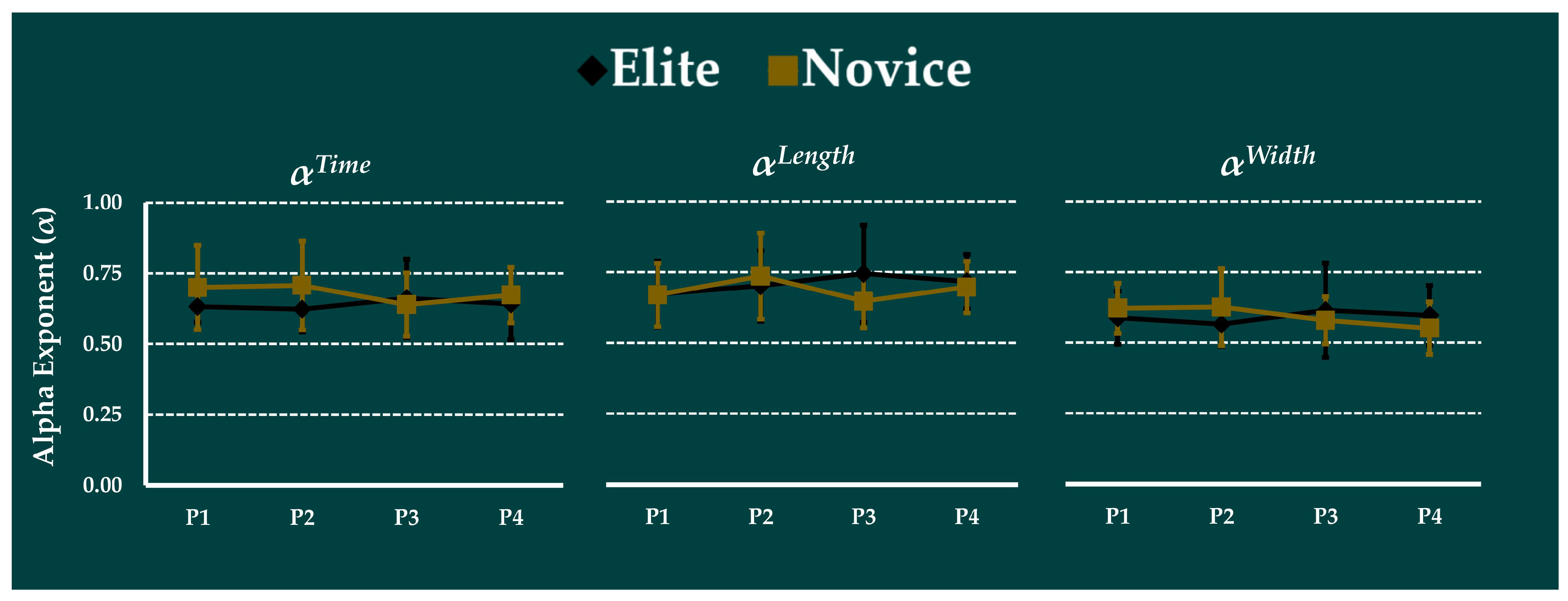
2.4.2. Calculation of Running Complexity
2.5. Statistics
3. Results
3.1. Effect of Prolonged Running on SSFs and SSV of Spatiotemporal Variables
3.2. Effect of Prolonged Running on SSFs of Kinematic Variables
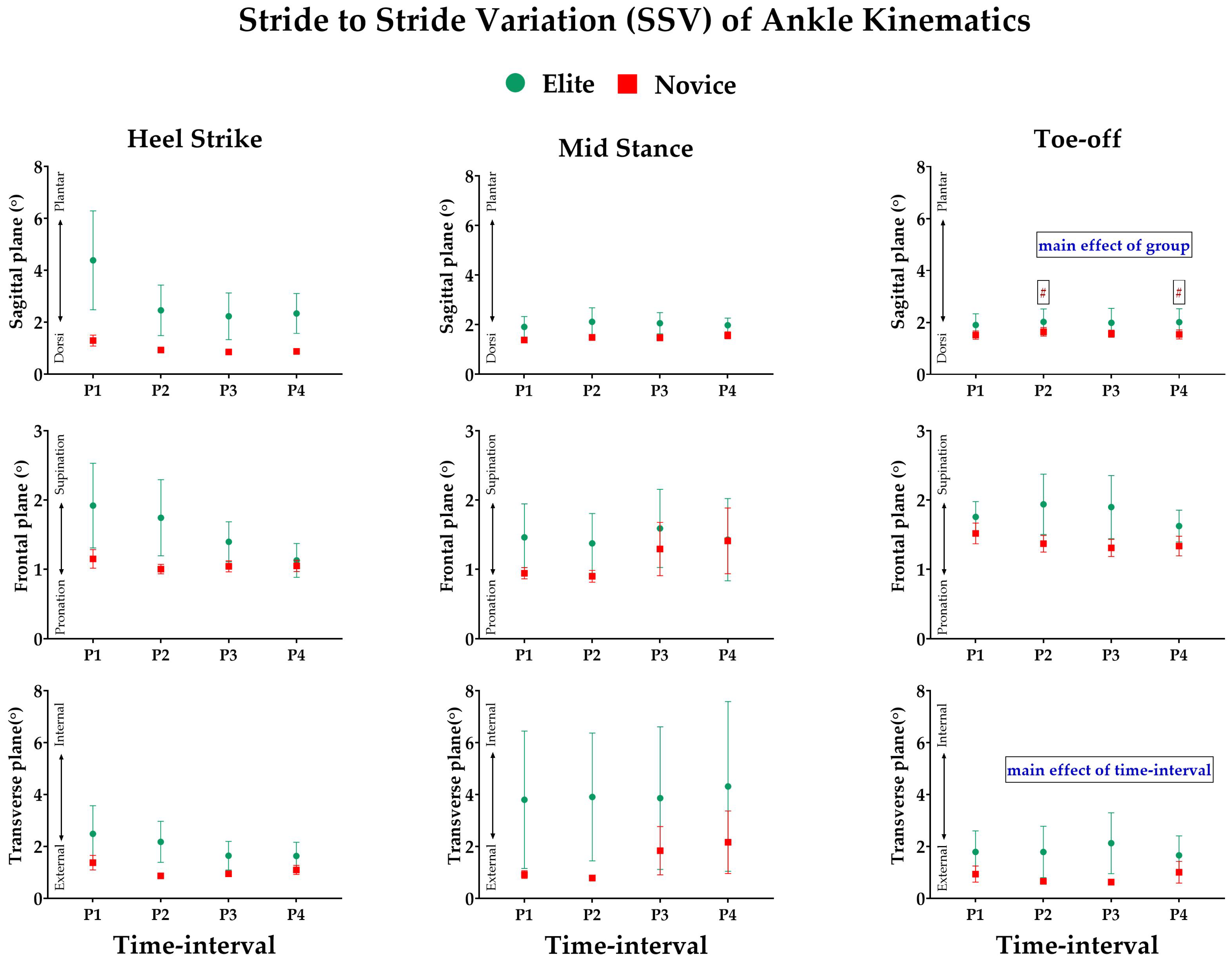
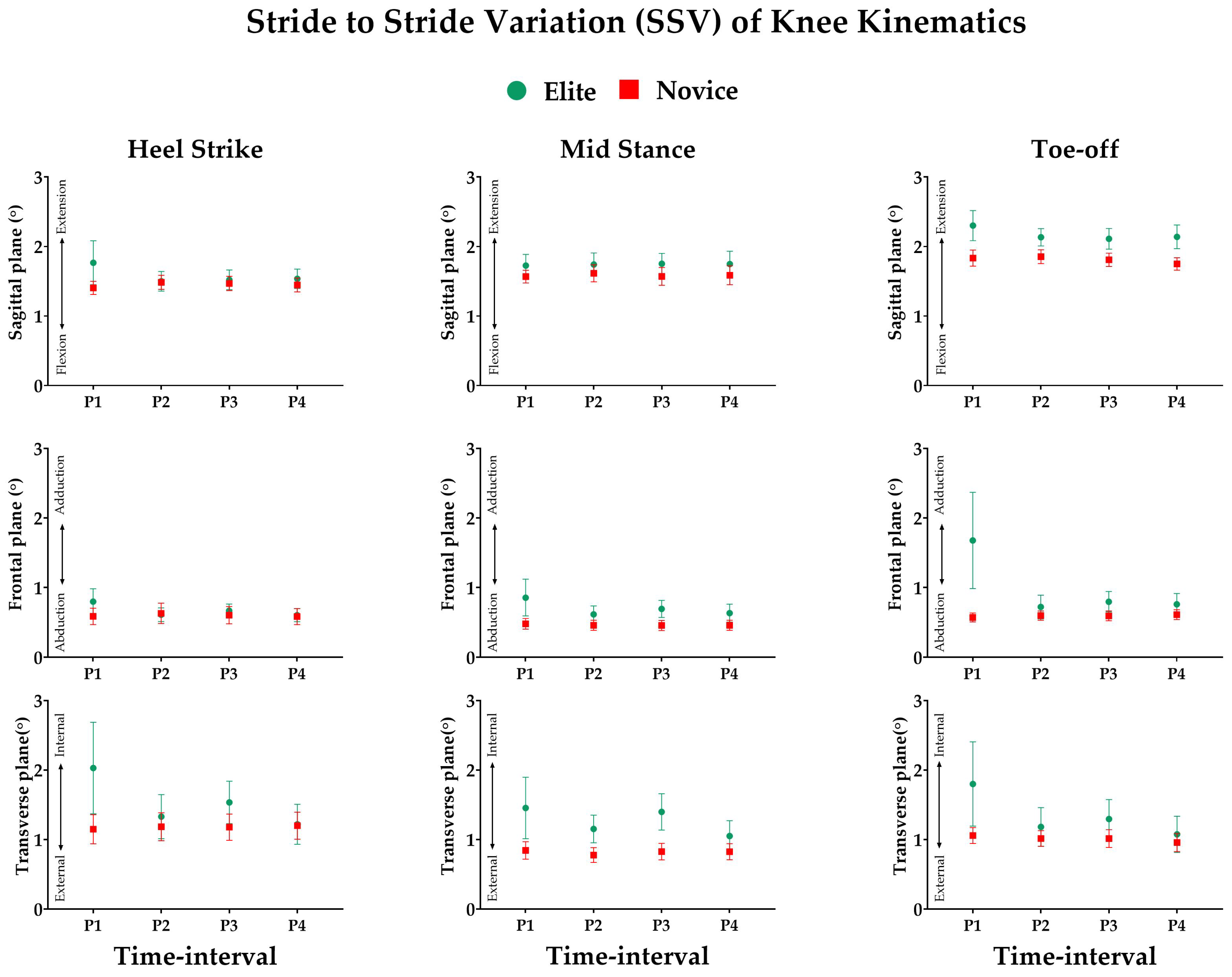
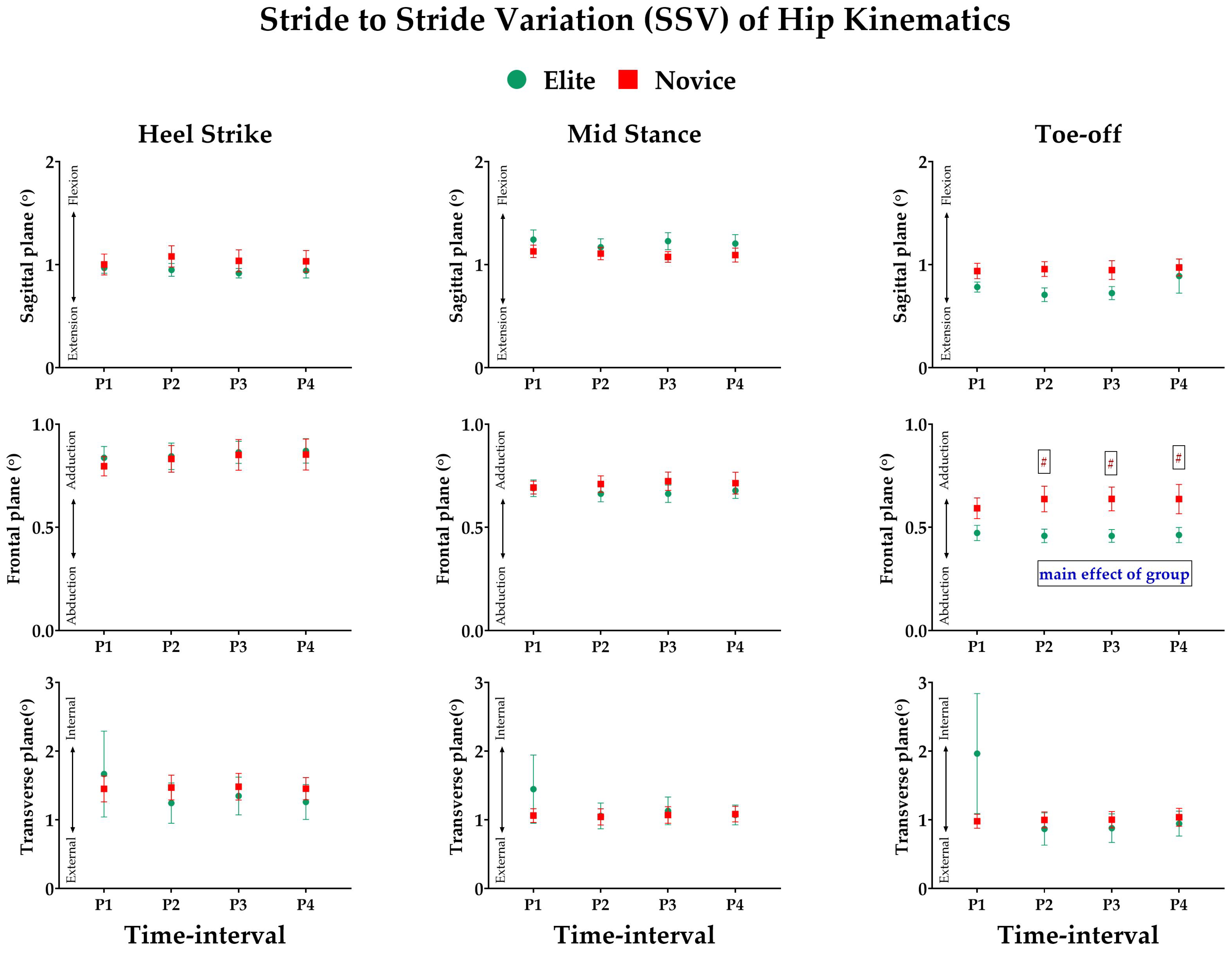
3.3. Effect of Prolonged Running on SSV of Kinematic Variables
3.4. Effect of Prolonged Running on Complexity
3.5. Correlation Analysis of Complexity with Joint Kinematics SSVs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.N | CODE | Age (Years) | Mass (kg) | Height (m) | PRS (m/s) | Half Marathon Record (hh:mm) | Experience (Years) |
|---|---|---|---|---|---|---|---|
| 1 | S#E01 | 34 | 50 | 1.65 | 3.75 | 1:21 | 3 |
| 2 | S#E02 | 23 | 56 | 1.71 | 3.89 | 1:10 | 11 |
| 3 | S#E03 | 33 | 68 | 1.72 | 3.47 | 1:22 | 5 |
| 4 | S#E04 | 28 | 67 | 1.72 | 4.03 | 1:19 | 4 |
| 5 | S#E05 | 26 | 61 | 1.73 | 3.94 | 1:10 | 10 |
| 6 | S#E06 | 23 | 65 | 1.76 | 3.86 | 1:07 | 13 |
| 7 | S#E07 | 27 | 78 | 1.77 | 3.64 | 1:22 | 3 |
| 8 | S#E08 | 26 | 66 | 1.82 | 3.56 | 1:10 | 14 |
| 9 | S#E09 | 30 | 83 | 1.87 | 3.89 | 1:14 | 10 |
| 10 | S#E10 | 30 | 68 | 1.88 | 3.83 | 1:06 | 13 |
| Elite | Mean | 28 | 66 | 1.76 | 3.79 | 1:14 | 8.60 |
| [M71] SD | 4 | 10 | 0.07 | 0.18 | 0:06 | 4.40 | |
| 1 | S#N01 | 20 | 77 | 1.83 | 2.86 | N/A | N/A |
| 2 | S#N02 | 20 | 69 | 1.84 | 3.06 | ||
| 3 | S#N03 | 22 | 75 | 1.74 | 3.06 | ||
| 4 | S#N04 | 23 | 62 | 1.71 | 2.61 | ||
| 5 | S#N05 | 24 | 81 | 1.9 | 2.92 | ||
| 6 | S#N06 | 24 | 76 | 1.8 | 2.92 | ||
| 7 | S#N07 | 25 | 73 | 1.73 | 2.25 | ||
| 8 | S#N08 | 25 | 83 | 1.74 | 2.53 | ||
| 9 | S#N09 | 25 | 79 | 1.87 | 3.06 | ||
| 10 | S#N10 | 26 | 68 | 1.77 | 3.06 | ||
| 11 | S#N11 | 31 | 89 | 1.85 | 2.78 | ||
| Novice | Mean | 24 | 76 | 1.80 | 2.83 | N/A | N/A |
| SD | 3 | 8 | 0.06 | 0.26 |
Appendix B
Appendix B.1. Estimation Procedure of Preferred Running Speed (PRS)
Appendix B.2. Calculation of Submaximal Level of Anaerobic Threshold Percentage Using Respiratory Exchange Ratio (RER)
Appendix B.3. Details on Marker Attachment Location
- Joint markers: left/right anterior superior iliac spine, left/right posterior superior iliac spine, left/right great trochanter, left/right femur lateral epicondyle, left/right femur medial epicondyle, left/right fibula apex of the lateral malleolus, left/right tibia apex of the medial malleolus, left/right posterior surface of the calcaneus, left/right head of the 5th metatarsus, left/right proximal medial phalanx, left/right head of the 1st metatarsus.
- Tracking clusters markers: three tracking markers for each segment of the thighs (left/right) and shanks (left/right).
Appendix C







References
- Meardon, S.A.; Hamill, J.; Derrick, T.R. Running injury and stride time variability over a prolonged run. Gait Posture 2011, 33, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Kudo, K.; Ohtsuki, T. Variability and fluctuation in running gait cycle of trained runners and non-runners. Gait Posture 2010, 31, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Fuller, J.T.; Amado, A.; van Emmerik, R.E.; Hamill, J.; Buckley, J.D.; Tsiros, M.D.; Thewlis, D. The effect of footwear and footfall pattern on running stride interval long-range correlations and distributional variability. Gait Posture 2016, 44, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Hamacher, D.; Singh, N.; Van Dieen, J.; Heller, M.; Taylor, W. Kinematic measures for assessing gait stability in elderly individuals: A systematic review. J. R. Soc. Interface 2011, 8, 1682–1698. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Hogan, N. Improved assessment of orbital stability of rhythmic motion with noise. PLoS ONE 2015, 10, e0119596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausdorff, J.M.; Purdon, P.L.; Peng, C.-K.; Ladin, Z.; Wei, J.Y.; Goldberger, A.L. Fractal dynamics of human gait: Stability of long-range correlations in stride interval fluctuations. J. Appl. Physiol. 1996, 80, 1448–1457. [Google Scholar] [CrossRef]
- Jordan, K.; Challis, J.H.; Newell, K.M. Long range correlations in the stride interval of running. Gait Posture 2006, 24, 120–125. [Google Scholar] [CrossRef]
- Srinivasan, M.; Ruina, A. Computer optimization of a minimal biped model discovers walking and running. Nature 2006, 439, 72–75. [Google Scholar] [CrossRef]
- Roberts, T.J.; Scales, J.A. Adjusting muscle function to demand: Joint work during acceleration in wild turkeys. J. Exp. Biol. 2004, 207, 4165–4174. [Google Scholar] [CrossRef] [Green Version]
- Abbiss, C.R.; Laursen, P.B. Describing and understanding pacing strategies during athletic competition. Sports Med. 2008, 38, 239–252. [Google Scholar] [CrossRef]
- De Leeuw, A.-W.; Meerhoff, L.A.; Knobbe, A. Effects of pacing properties on performance in long-distance running. Big Data 2018, 6, 248–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skorski, S.; Abbiss, C.R. The manipulation of pace within endurance sport. Front. Physiol. 2017, 8, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Lozano, A.; Collado, P.; Foster, C.; Lucia, A.; Garatachea, N. Influence of sex and level on marathon pacing strategy. Insights from the New York City race. Int. J. Sports Med. 2014, 35, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Haney, T.A., Jr.; Mercer, J.A. A description of variability of pacing in marathon distance running. Int. J. Exerc. Sci. 2011, 4, 133. [Google Scholar]
- Kais, Ü.; Mooses, K.; Pind, R.; Pehme, A.; Kaasik, P.; Mooses, M. Pacing strategy of the finishers of the world marathon majors series. Kinesiology 2019, 51, 22–27. [Google Scholar] [CrossRef]
- Hamill, J.; Palmer, C.; Van Emmerik, R.E. Coordinative variability and overuse injury. Sports Med. Arthrosc. Rehabil. Ther. Technol. 2012, 4, 45. [Google Scholar] [CrossRef] [Green Version]
- Hiley, M.J.; Zuevsky, V.V.; Yeadon, M.R. Is skilled technique characterized by high or low variability? An analysis of high bar giant circles. Hum. Mov. Sci. 2013, 32, 171–180. [Google Scholar] [CrossRef]
- Floría, P.; Sánchez-Sixto, A.; Ferber, R.; Harrison, A.J. Effects of running experience on coordination and its variability in runners. J. Sports Sci. 2018, 36, 272–278. [Google Scholar] [CrossRef]
- Sloan, R.S.; Wight, J.T.; Hooper, D.R.; Garman, J.E.; Pujalte, G.G. Metabolic testing does not alter distance running lower body sagittal kinematics. Gait Posture 2020, 76, 403–408. [Google Scholar] [CrossRef]
- Wight, J.T.; Garman, J.E.; Hooper, D.R.; Robertson, C.T.; Ferber, R.; Boling, M.C. Distance running stride-to-stride variability for sagittal plane joint angles. Sports Biomech. 2020, 21, 966–980. [Google Scholar] [CrossRef]
- Mo, S.; Chow, D.H. Stride-to-stride variability and complexity between novice and experienced runners during a prolonged run at anaerobic threshold speed. Gait Posture 2018, 64, 7–11. [Google Scholar] [CrossRef] [PubMed]
- De Ruiter, C.J.; Verdijk, P.W.; Werker, W.; Zuidema, M.J.; de Haan, A. Stride frequency in relation to oxygen consumption in experienced and novice runners. Eur. J. Sport Sci. 2014, 14, 251–258. [Google Scholar] [CrossRef] [PubMed]
- García-Pinillos, F.; García-Ramos, A.; Ramírez-Campillo, R.; Latorre-Román, P.Á.; Roche-Seruendo, L.E. How do spatiotemporal parameters and lower-body stiffness change with increased running velocity? A comparison between novice and elite level runners. J. Hum. Kinet. 2019, 70, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, I.; Lee, K.; Ward, J.; Tracy, J. Self-optimization of stride length among experienced and inexperienced runners. Int. J. Exerc. Sci. 2017, 10, 446. [Google Scholar]
- Jordan, K.; Challis, J.H.; Newell, K.M. Speed influences on the scaling behavior of gait cycle fluctuations during treadmill running. Hum. Mov. Sci. 2007, 26, 87–102. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G* Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Wu, G.; Cavanagh, P.R. ISB recommendations for standardization in the reporting of kinematic data. J. Biomech. 1995, 28, 1257–1261. [Google Scholar] [CrossRef]
- Baldari, C.; Meucci, M.; Bolletta, F.; Gallotta, M.; Emerenziani, G.; Guidetti, L. Accuracy and reliability of COSMED K5 portable metabolic device versus simulating system. Sport Sci. Health 2015, 11, 58. [Google Scholar]
- Panday, S.B. Relationship between Orbital Stability and Efficiency in Running. Ph.D. Thesis, Seoul National University, Seoul, Korea, 2021. [Google Scholar]
- Daniels, J. Daniels’ Running Formula; Human Kinetics: Champaign, IL, USA, 2014. [Google Scholar]
- Dingwell, J.B.; Marin, L.C. Kinematic variability and local dynamic stability of upper body motions when walking at different speeds. J. Biomech. 2006, 39, 444–452. [Google Scholar] [CrossRef]
- Ramos-Jiménez, A.; Hernández-Torres, R.P.; Torres-Durán, P.V.; Romero-Gonzalez, J.; Mascher, D.; Posadas-Romero, C.; Juárez-Oropeza, M.A. The respiratory exchange ratio is associated with fitness indicators both in trained and untrained men: A possible application for people with reduced exercise tolerance. Clin. Med. Circ. Respir. Pulm. Med. 2008, 2, CCRPM-S449. [Google Scholar] [CrossRef]
- Solberg, G.; Robstad, B.; Skjønsberg, O.H.; Borchsenius, F. Respiratory gas exchange indices for estimating the anaerobic threshold. J. Sports Sci. Med. 2005, 4, 29. [Google Scholar] [PubMed]
- Kristianslund, E.; Krosshaug, T.; van den Bogert, A.J. Effect of low pass filtering on joint moments from inverse dynamics: Implications for injury prevention. J. Biomech. 2012, 45, 666–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, N.; Delattre, N.; Berton, E.; Rao, G. Does an increase in energy return and/or longitudinal bending stiffness shoe features reduce the energetic cost of running? Eur. J. Appl. Physiol. 2019, 119, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.D.; Ghoussayni, S.N.; Ewins, D.J.; Kent, J.A. A six degrees-of-freedom marker set for gait analysis: Repeatability and comparison with a modified Helen Hayes set. Gait Posture 2009, 30, 173–180. [Google Scholar] [CrossRef]
- Dugan, S.A.; Bhat, K.P. Biomechanics and analysis of running gait. Phys. Med. Rehabil. Clin. 2005, 16, 603–621. [Google Scholar] [CrossRef] [PubMed]
- Zeni, J., Jr.; Richards, J.; Higginson, J. Two simple methods for determining gait events during treadmill and overground walking using kinematic data. Gait Posture 2008, 27, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Peng, C.-K.; Buldyrev, S.V.; Havlin, S.; Simons, M.; Stanley, H.E.; Goldberger, A.L. Mosaic organization of DNA nucleotides. Phys. Rev. E 1994, 49, 1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ihlen, E.A.F.E. Introduction to multifractal detrended fluctuation analysis in Matlab. Front. Physiol. 2012, 3, 141. [Google Scholar] [CrossRef] [Green Version]
- Bruijn, S.; Meijer, O.; Beek, P.; Van Dieën, J. Assessing the stability of human locomotion: A review of current measures. J. R. Soc. Interface 2013, 10, 20120999. [Google Scholar] [CrossRef]
- Ahn, J.; Hogan, N. Long-range correlations in stride intervals may emerge from non-chaotic walking dynamics. PLoS ONE 2013, 8, e73239. [Google Scholar] [CrossRef] [Green Version]
- Hausdorff, J.M. Gait dynamics, fractals and falls: Finding meaning in the stride-to-stride fluctuations of human walking. Hum. Mov. Sci. 2007, 26, 555–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekkekakis, P.; Parfitt, G.; Petruzzello, S.J. The pleasure and displeasure people feel when they exercise at different intensities. Sports Med. 2011, 41, 641–671. [Google Scholar] [CrossRef] [PubMed]
- Moore, I.S. Is there an economical running technique? A review of modifiable biomechanical factors affecting running economy. Sports Med. 2016, 46, 793–807. [Google Scholar] [CrossRef] [Green Version]
- Arellano, C.J.; Kram, R. The energetic cost of maintaining lateral balance during human running. J. Appl. Physiol. 2012, 112, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brindle, R.A.; Milner, C.E.; Zhang, S.; Fitzhugh, E.C. Changing step width alters lower extremity biomechanics during running. Gait Posture 2014, 39, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Dingwell, J.B.; Cusumano, J.P. Re-interpreting detrended fluctuation analyses of stride-to-stride variability in human walking. Gait Posture 2010, 32, 348–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maas, E.; De Bie, J.; Vanfleteren, R.; Hoogkamer, W.; Vanwanseele, B. Novice runners show greater changes in kinematics with fatigue compared with competitive runners. Sports Biomech. 2018, 17, 350–360. [Google Scholar] [CrossRef]
- De Wit, B.; De Clercq, D.; Aerts, P. Biomechanical analysis of the stance phase during barefoot and shod running. J. Biomech. 2000, 33, 269–278. [Google Scholar] [CrossRef]
- Lohman, E.B., III; Sackiriyas, K.S.B.; Swen, R.W. A comparison of the spatiotemporal parameters, kinematics, and biomechanics between shod, unshod, and minimally supported running as compared to walking. Phys. Ther. Sport 2011, 12, 151–163. [Google Scholar] [CrossRef]
- Kuitunen, S.; Komi, P.V.; Kyröläinen, H. Knee and ankle joint stiffness in sprint running. Med. Sci. Sports Exerc. 2002, 34, 166–173. [Google Scholar] [CrossRef]
- Lieberman, D.E.; Raichlen, D.A.; Pontzer, H.; Bramble, D.M.; Cutright-Smith, E. The human gluteus maximus and its role in running. J. Exp. Biol. 2006, 209, 2143–2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, H.-L.; Powers, C.M. Influence of trunk posture on lower extremity energetics during running. Med. Sci. Sports Exerc. 2015, 47, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Reenalda, J.; Maartens, E.; Buurke, J.H.; Gruber, A.H. Kinematics and shock attenuation during a prolonged run on the athletic track as measured with inertial magnetic measurement units. Gait Posture 2019, 68, 155–160. [Google Scholar] [CrossRef]
- Günther, M.; Blickhan, R. Joint stiffness of the ankle and the knee in running. J. Biomech. 2002, 35, 1459–1474. [Google Scholar] [CrossRef]
- Lees, A.; Bouracier, J. The longitudinal variability of ground reaction forces in experienced and inexperienced runners. Ergonomics 1994, 37, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Hamill, J.; van Emmerik, R.E.; Heiderscheit, B.C.; Li, L. A dynamical systems approach to lower extremity running injuries. Clin. Biomech. 1999, 14, 297–308. [Google Scholar] [CrossRef]
- Mo, S.; Chow, D.H.K. Differences in lower-limb coordination and coordination variability between novice and experienced runners during a prolonged treadmill run at anaerobic threshold speed. J. Sports Sci. 2019, 37, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Tam, N.; Tucker, R.; Santos-Concejero, J.; Prins, D.; Lamberts, R.P. Running economy: Neuromuscular and joint-stiffness contributions in trained runners. Int. J. Sports Physiol. Perform. 2019, 14, 16–22. [Google Scholar] [CrossRef]
- Robergs, R.A.; Dwyer, D.; Astorino, T. Recommendations for improved data processing from expired gas analysis indirect calorimetry. Sports Med. 2010, 40, 95–111. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panday, S.B.; Pathak, P.; Moon, J.; Koo, D. Complexity of Running and Its Relationship with Joint Kinematics during a Prolonged Run. Int. J. Environ. Res. Public Health 2022, 19, 9656. https://doi.org/10.3390/ijerph19159656
Panday SB, Pathak P, Moon J, Koo D. Complexity of Running and Its Relationship with Joint Kinematics during a Prolonged Run. International Journal of Environmental Research and Public Health. 2022; 19(15):9656. https://doi.org/10.3390/ijerph19159656
Chicago/Turabian StylePanday, Siddhartha Bikram, Prabhat Pathak, Jeheon Moon, and Dohoon Koo. 2022. "Complexity of Running and Its Relationship with Joint Kinematics during a Prolonged Run" International Journal of Environmental Research and Public Health 19, no. 15: 9656. https://doi.org/10.3390/ijerph19159656