
Exploring Mycosporine-like Amino Acid UV-Absorbing Natural Products for a New Generation of Environmentally Friendly Sunscreens
 ,
,  , , and
, , and
Abstract
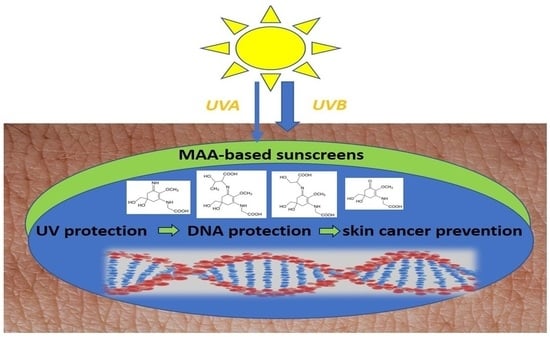
:
1. Introduction
2. Fitzpatrick Phototype and UV Protection Strategies
3. Natural UV-Absorbing Compounds

{kind=link}
{kind=link}
| UV-Protective Natural Products | Chemical Structure | Key Features/Bioactive Properties | Source of the Compounds | |
|---|---|---|---|---|
| Flavonoids | Quercetin (C15H10O7) |  | UV-absorbing, antioxidant-scavenging reactive oxygen species induced by UVA and UVB radiation [76,84] | Fruits and vegetables [85,86,87] |
| Apigenin (C15H10O5) |  | UV-absorbing, antioxidant against UVA and UVB radiation [88] | Parsley, celery, celeriac, basil, chamomile tea [89,90,91] | |
| Myricetin (C15H10O8) |  | UV-absorbing, suppressing UVB-induced wrinkle formation [77] | Fruits, vegetables, tea, red wine [92] | |
| Kaempferol (C15H10O6) |  | UV-absorbing, antioxidant [93] | Fruits and vegetables: grapes, tomatoes, broccoli, spinach [94] | |
| Taxifolin (C15H12O7) |  | UVA- and UVB-protective [84,95], antioxidant [96] | Citrus fruits and onion [96] | |
| Polyphenols | (−)-Epigallocatechin gallate (C22H18O11) |  | UV-absorbing [97], antioxidant [98] | Green tea [99] |
| Tannic acid (C76H52O46) |  | UV-absorbing, antioxidant [78] | All aerial plant tissues [100] | |
| Resveratrol (C14H12O3) |  | UV-absorbing [101], antioxidant [102] | Grapes, apples, wine, peanuts, and soy [103,104] | |
| Curcumin (C21H20O6) |  | UV-absorbing [105] anti-inflammatory [106] | Plant Curcuma longa [90] | |
| Terpenoids | α-Tocopherol (C29H50O2) |  | UV-absorbing (385 nm) [107], antioxidants [108] | Vegetable oils, nuts, and whole grains [109] |
| Astaxanthin (C40H52O4) |  | Antioxidants, prevented UVA-mediated DNA damage [110] | Fungi, bacteria, algae, crustaceans, and some fishes [111,112] | |
| Mycosporines-like amino acids | Mycosporine-glycine (C10H15NO6) |  | UV-absorbing, antioxidants [79,113] | Cyanobacteria Chlorogloeopsis sp. PCC6912 [114]; Gloeocapsa sp. [115]; Nostoc commune [116] Macroalgae Acanthophora specifera [22,117], species from genus Bostrychia [118], and Devaleraea [119] Arthropoda, Molluscs, Cnidaria, Echinodermata, Protochordata, Phytoplankton, Nemertinea, Porifera, etc. [9] |
| Shinorine (C13H20N2O8) |  | UV-absorbing, antioxidants [38,120] | Cyanobacteria (Chlorogloeopsis sp. PCC6912 [114]; Gloeocapsa sp. [115]; Nostoc commune [53]) Macroalgae Acanthophora specifera [22], species from the genus Asparagopsis [56,57,58] and Bostrychia [118], and Devaleraea ramentacea [119] Arthropoda, Molluscs, Cnidaria, Echinodermata, Protochordata, Phytoplankton, Nemertinea, Porifera etc. [9] | |
| Porphyra-334 (C14H22N2O8) |  | UV-absorbing, antioxidants [38,120] | Cyanobacteria Nostoc harveyana [52] Macroalgae species from the genus Bostrychia [118] and Porphyra [117] Devaleraea ramentacea [119] Arthropoda, Molluscs, Cnidaria, Echinodermata, Protochordata, Phytoplankton Nemertinea, Porifera etc. [9] | |
| Mycosporine-2-glycine (C12H18N2O7) |  | UV-absorbing, antioxidants [42] | Cyanobacteria Euhalothece sp. LK-1 [121] and Aphanothece halophytica [122] Sea anemone Anthopleura elegantissima [123], dinoflagellate Maristentor dinoferus [124] Molluscs, Cnidaria and others [9] | |
| Palythine (C13H20N2O5;) |  | UV-absorbing, antioxidants [80] | Macroalgae Acanthophora specifera [22,117], Bostrychia species [118] Phytoplankton, Porifera Chordata [9] | |
4. Limitations and Challenges in Using UV-Absorbing MAAs in Sunscreens
5. Investigation of the Efficacy of Natural Products for Use as Sunscreens
| Model | Advantages | Disadvantages | |
|---|---|---|---|
| In vitro | Cell culture—primary/immortalized keratinocytes [134,135] |
|
|
| Reconstructed skin [138,139] |
|
| |
| Skin explants [140] |
|
| |
| In vivo | Murine models [141,142,143] |
|
|
| Porcine models [146,147] |
|
| |
| Human volunteers [148] |
|
|
6. Genetics of Marine Organisms Producing MAAs
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bertram, C.; Hass, R. Cellular responses to reactive oxygen species-induced DNA damage and aging. Biol. Chem. 2008, 389, 211. [Google Scholar] [CrossRef] [PubMed]
- Savoye, I.; Olsen, C.M.; Whiteman, D.C.; Bijon, A.; Wald, L.; Dartois, L.; Clavel-Chapelon, F.; Boutron-Ruault, M.C.; Kvaskoff, M. Patterns of Ultraviolet Radiation Exposure and Skin Cancer Risk: The E3N-SunExp Study. J. Epidemiol. 2018, 28, 27. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222. [Google Scholar] [CrossRef]
- Leiter, U.; Eigentler, T.; Garbe, C. Epidemiology of skin cancer. Adv. Exp. Med. Biol. 2014, 810, 120. [Google Scholar]
- Armstrong, B.K.; Kricker, A. The epidemiology of UV induced skin cancer. J. Photochem. Photobiol. B 2001, 63, 8. [Google Scholar] [CrossRef] [PubMed]
- Ikehata, H.; Ono, T. The mechanisms of UV mutagenesis. J. Radiat. Res. 2011, 52, 115. [Google Scholar] [CrossRef]
- Ichihashi, M.; Ueda, M.; Budiyanto, A.; Bito, T.; Oka, M.; Fukunaga, M.; Tsuru, K.; Horikawa, T. UV-induced skin damage. Toxicology 2003, 189, 21. [Google Scholar] [CrossRef]
- Rosic, N.N.; Dove, S. Mycosporine-like amino acids from coral dinoflagellates. Appl. Environ. Microbiol. 2011, 77, 8478. [Google Scholar] [CrossRef]
- Sinha, R.P.; Singh, S.P.; Häder, D.P. Database on mycosporines and mycosporine-like amino acids (MAAs) in fungi, cyanobacteria, macroalgae, phytoplankton and animals. J. Photochem. Photobiol. B Biol. 2007, 89, 29. [Google Scholar] [CrossRef]
- Carreto, J.I.; Carignan, M.O. Mycosporine-like amino acids: Relevant secondary metabolites. chemical and ecological aspects. Mar. Drugs 2011, 9, 387. [Google Scholar]
- Figueroa, F.L. Mycosporine-Like Amino Acids from Marine Resource. Mar. Drugs 2021, 19, 18. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Kumari, S.; Rastogi, R.P.; Singh, K.L.; Sinha, R.P. Mycosporine-like amino acids (MAAs): Chemical structure, biosynthesis and significance as UV-absorbing/screening compounds. Indian J. Exp. Biol. 2008, 46, 7. [Google Scholar]
- Vega, J.; Schneider, G.; Moreira, B.R.; Herrera, C.; Bonomi-Barufi, J.; Figueroa, F.L. Mycosporine-Like Amino Acids from Red Macroalgae: UV-Photoprotectors with Potential Cosmeceutical Applications. Appl. Sci. 2021, 11, 5112. [Google Scholar] [CrossRef]
- Rosic, N.; Ling, E.Y.S.; Chan, C.-K.K.; Lee, H.C.; Kaniewska, P.; Edwards, D.; Dove, S.; Hoegh-Guldberg, O. Unfolding the secrets of coral-algal symbiosis. ISME J. 2015, 9, 844. [Google Scholar] [CrossRef] [PubMed]
- Kaniewska, P.; Chan, C.-K.K.; Kline, D.; Ling, E.Y.S.; Rosic, N.; Edwards, D.; Hoegh-Guldberg, O.; Dove, S. Transcriptomic Changes in Coral Holobionts Provide Insights into Physiological Challenges of Future Climate and Ocean Change. PLoS ONE 2015, 10, e0139223. [Google Scholar] [CrossRef] [PubMed]
- Rosic, N.; Rémond, C.; Mello-Athayde, M.A. Differential impact of heat stress on reef-building corals under different light conditions. Mar. Environ. Res. 2020, 158, 104947. [Google Scholar] [CrossRef]
- Shick, J.M.; Dunlap, W.C. Mycosporine-like amino acids and related gadusols: Biosynthesis, accumulation, and UV-protective functions in aquatic organisms. Annu. Rev. Physiol. 2002, 64, 223–262. [Google Scholar] [CrossRef]
- Lawrence, K.P.; Long, P.F.; Young, A.R. Mycosporine-Like Amino Acids for Skin Photoprotection. Curr. Med. Chem. 2018, 25, 5512. [Google Scholar] [CrossRef]
- Rosic, N.N. Mycosporine-Like Amino Acids: Making the Foundation for Organic Personalised Sunscreens. Mar. Drugs 2019, 17, 638. [Google Scholar] [CrossRef]
- Oren, A.; Gunde-Cimerman, N. Mycosporines and mycosporine-like amino acids: UV protectants or multipurpose secondary metabolites? FEMS Microbiol. Lett. 2007, 269, 1–10. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Incharoensakdi, A. Characterization of UV-screening compounds, mycosporine-like amino acids, and scytonemin in the cyanobacterium Lyngbya sp. CU2555. FEMS Microbiol. Ecol. 2014, 87, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Rosic, N.N.; Braun, C.; Kvaskoff, D. Extraction and Analysis of Mycosporine-Like Amino Acids in Marine Algae. Methods Mol. Biol. 2015, 1308, 119–129. [Google Scholar] [PubMed]
- Núñez-Pons, L.; Avila, C.; Romano, G.; Verde, C.; Giordano, D. UV-Protective Compounds in Marine Organisms from the Southern Ocean. Mar. Drugs 2018, 16, 336. [Google Scholar] [CrossRef]
- Parailloux, M.; Godin, S.; Fernandes, S.C.; Lobinski, R. Untargeted Analysis for Mycosporines and Mycosporine-Like Amino Acids by Hydrophilic Interaction Liquid Chromatography (HILIC)—Electrospray Orbitrap MS2/MS3. Antioxidants 2020, 9, 1185. [Google Scholar] [CrossRef] [PubMed]
- Tartarotti, B.; Sommaruga, R. Seasonal and ontogenetic changes of mycosporine-like amino acids in planktonic organisms from an alpine lake. Limnol. Oceanogr. 2006, 51, 1530–1541. [Google Scholar] [CrossRef]
- Al-Utaibi, A.A.; Niaz, G.R.; Al-Lihaibi, S.S. Mycosporine-like amino acids in six scleractinian coral species. Oceanologia 2009, 51, 93–104. [Google Scholar] [CrossRef]
- Raj, S.; Kuniyil, A.M.; Sreenikethanam, A.; Gugulothu, P.; Jeyakumar, R.B.; Bajhaiya, A.K. Microalgae as a Source of Mycosporine-like Amino Acids (MAAs); Advances and Future Prospects. Int. J. Environ. Res. Public Health 2021, 18, 12402. [Google Scholar] [CrossRef]
- Rosic, N. Molecular mechanisms of stress tolerance in cyanobacteria. In Ecophysiology and Biochemistry of Cyanobacteria; Rastogi, R.P., Ed.; Springer Nature Singapore: Singapore, 2021; p. 131. [Google Scholar]
- Rosic, N.; Kaniewska, P.; Chan, C.-K.K.; Ling, E.Y.S.; Edwards, D.; Dove, S.; Hoegh-Guldberg, O. Early transcriptional changes in the reef-building coral Acropora aspera in response to thermal and nutrient stress. BMC Genom. 2014, 15, 1052. [Google Scholar] [CrossRef]
- Singh, A.; Čížková, M.; Bišová, K.; Vítová, M. Exploring Mycosporine-Like Amino Acids (MAAs) as Safe and Natural Protective Agents against UV-Induced Skin Damage. Antioxidants 2021, 10, 683. [Google Scholar] [CrossRef]
- Morey, J.S.; Monroe, E.A.; Kinney, A.L.; Beal, M.; Johnson, J.G.; Hitchcock, G.L.; Van Dolah, F.M. Transcriptomic response of the red tide dinoflagellate, Karenia brevis, to nitrogen and phosphorus depletion and addition. BMC Genom. 2011, 12, 346. [Google Scholar] [CrossRef]
- Korbee, N.; Mata, M.T.; Figueroa, F.L. Photoprotection mechanisms against ultraviolet radiation in Heterocapsa sp. (Dinophyceae) are influenced by nitrogen availability: Mycosporine-like amino acids vs. xanthophyll cycle. Limnol. Oceanogr. 2010, 55, 899. [Google Scholar]
- Singh, S.P.; Klisch, M.; Sinha, R.P.; Häder, D.-P. Effects of abiotic stressors on synthesis of the mycosporine-like amino acid shinorine in the cyanobacterium anabaena variabilis PCC 7937. Photochem. Photobiol. 2008, 84, 1500–1505. [Google Scholar] [CrossRef] [PubMed]
- Peinado, N.K.; Díaz, R.T.A.; Figueroa, F.L.; Helbling, E.W. Ammonium and uv radiation stimulate the accumulation of mycosporine-like amino acids in porphyra columbina (rhodophyta) from patagonia, argentina1. J. Phycol. 2004, 40, 248. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Richa, R.P.; Sinha Singh, S.P.; Häder, D.P. Photoprotective compounds from marine organisms. J. Ind. Microbiol. Biotechnol. 2010, 37, 537. [Google Scholar] [CrossRef] [PubMed]
- Conde, F.R.; Churio, M.S.; Previtali, C.M. Experimental study of the excited-state properties and photostability of the mycosporine-like amino acid palythine in aqueous solution. Photochem. Photobiol. Sci. 2007, 6, 669–674. [Google Scholar] [CrossRef]
- de la Coba, F.; Aguilera, J.; Figueroa, F.L.; de Gálvez, M.V.; Herrera, E. Antioxidant activity of mycosporine-like amino acids isolated from three red macroalgae and one marine lichen. J. Appl. Phycol. 2008, 21, 161–169. [Google Scholar] [CrossRef]
- Gacesa, R.; Lawrence, K.P.; Georgakopoulos, N.D.; Yabe, K.; Dunlap, W.C.; Barlow, D.J.; Wells, G.; Young, A.R.; Long, P.F. The mycosporine-like amino acids porphyra-334 and shinorine are antioxidants and direct antagonists of Keap1-Nrf2 binding. Biochimie 2018, 154, 35–44. [Google Scholar] [CrossRef]
- Kumar, M.S.; Vijaylaxmi, K.K.; Pal, A.K. Antiinflamatuvar and Antioxidant Properties of Spongosorites halichondriodes, a Marine Sponge. Turk. J. Pharm. Sci. 2014, 11, 285. [Google Scholar]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89. [Google Scholar]
- Rastogi, R.P.; Incharoensakdi, A. UV radiation-induced biosynthesis, stability and antioxidant activity of mycosporine-like amino acids (MAAs) in a unicellular cyanobacterium Gloeocapsa sp. CU2556. J. Photochem. Photobiol. B Biol. 2014, 130, 287–292. [Google Scholar] [CrossRef]
- Tarasuntisuk, S.; Palaga, T.; Kageyama, H.; Waditee-Sirisattha, R. Mycosporine-2-glycine exerts anti-inflammatory and antioxidant effects in lipopolysaccharide (LPS)-stimulated RAW 264.7 macrophages. Arch. Biochem. Biophys. 2018, 662, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Koltover, V.K. Antioxidant biomedicine: From free radical chemistry to systems biology mechanisms. Russ. Chem. Bull. 2010, 59, 37–42. [Google Scholar] [CrossRef]
- Geraldes, V.; Pinto, E. Mycosporine-Like Amino Acids (MAAs): Biology, Chemistry and Identification Features. Pharmaceuticals 2021, 14, 63. [Google Scholar] [CrossRef] [PubMed]
- Rosic, N.N. Recent advances in the discovery of novel marine natural products and mycosporine-like amino acid UV-absorbing compounds. Appl. Microbiol. Biotechnol. 2021, 105, 7053–7067. [Google Scholar] [CrossRef] [PubMed]
- Chrapusta, E.; Kaminski, A.; Duchnik, K.; Bober, B.; Adamski, M.; Bialczyk, J. Mycosporine-Like Amino Acids: Potential Health and Beauty Ingredients. Mar. Drugs 2017, 15, 326. [Google Scholar] [CrossRef] [PubMed]
- Climstein, M.; Doyle, B.; Stapelberg, M.; Rosic, N.; Hertess, I.; Furness, J.; Simas, V.; Walsh, J. Point prevalence of non-melanoma and melanoma skin cancers in Australian surfers and swimmers in Southeast Queensland and Northern New South Wales. PeerJ 2022, 10, e13243. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef]
- Gilchrest, B.A.; Eller, M.S.; Geller, A.C.; Yaar, M. The Pathogenesis of Melanoma Induced by Ultraviolet Radiation. N. Engl. J. Med. 1999, 340, 1341–1348. [Google Scholar] [CrossRef]
- Laberge, G.S.; Duvall, E.; Grasmick, Z.; Haedicke, K.; Galan, A.; Leverett, J.; Baswan, S.; Yim, S.; Pawelek, J. Recent Advances in Studies of Skin Color and Skin Cancer. Yale J. Biol. Med. 2020, 93, 69–80. [Google Scholar]
- He, S.Y.; McCulloch, C.E.; Boscardin, W.J.; Chren, M.-M.; Linos, E.; Arron, S.T. Self-reported pigmentary phenotypes and race are significant but incomplete predictors of Fitzpatrick skin phototype in an ethnically diverse population. J. Am. Acad. Dermatol. 2014, 71, 731–737. [Google Scholar] [CrossRef]
- Gallagher, R.P.; Hill, G.B.; Bajdik, C.D.; Coldman, A.J.; Fincham, S.; McLean, D.I.; Threlfall, W.J. Sunlight Exposure, Pigmentation Factors, and Risk of Nonmelanocytic Skin Cancer. Arch. Dermatol. 1995, 131, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Green, A.; Williams, G.; Neale, R.; Hart, V.; Leslie, D.; Parsons, P.; Marks, G.C.; Gaffney, P.; Battistutta, D.; Frost, C.; et al. Daily sunscreen application and betacarotene supplementation in prevention of basal-cell and squamous-cell carcinomas of the skin: A randomised controlled trial. Lancet 1999, 354, 723. [Google Scholar] [CrossRef] [PubMed]
- Thompson, S.C.; Jolley, D.; Marks, R. Reduction of solar keratoses by regular sunscreen use. N. Engl. J. Med. 1993, 329, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Hung, M.; Beazer, I.R.; Su, S.; Bounsanga, J.; Hon, E.S.; Lipsky, M.S. An Exploration of the Use and Impact of Preventive Measures on Skin Cancer. Healthcare 2022, 10, 743. [Google Scholar] [CrossRef] [PubMed]
- Gambichler, T.; Altmeyer, P.; Hoffmann, K. Role of clothes in sun protection. Recent Results Cancer Res. 2002, 160, 15–25. [Google Scholar]
- Berry, E.G.; Bezecny, J.; Acton, M.; Sulmonetti, T.P.; Anderson, D.M.; Beckham, H.W.; Durr, R.A.; Chiba, T.; Beem, J.; Brash, D.E.; et al. Slip versus Slop: A Head-to-Head Comparison of UV-Protective Clothing to Sunscreen. Cancers 2022, 14, 542. [Google Scholar] [CrossRef]
- Wang, J.; Pan, L.; Wu, S.; Lu, L.; Xu, Y.; Zhu, Y.; Guo, M.; Zhuang, S. Recent Advances on Endocrine Disrupting Effects of UV Filters. Int. J. Environ. Res. Public Health 2016, 13, 782. [Google Scholar] [CrossRef]
- Ruszkiewicz, J.A.; Pinkas, A.; Ferrer, B.; Peres, T.V.; Tsatsakis, A.; Aschner, M. Neurotoxic effect of active ingredients in sunscreen products, a contemporary review. Toxicol. Rep. 2017, 4, 245–259. [Google Scholar] [CrossRef]
- de la Coba, F.; Aguilera, J.; Korbee, N.; de Gálvez, M.V.; Herrera-Ceballos, E.; Álvarez-Gómez, F.; Figueroa, F.L. UVA and UVB Photoprotective Capabilities of Topical Formulations Containing Mycosporine-like Amino Acids (MAAs) through Different Biological Effective Protection Factors (BEPFs). Mar. Drugs 2019, 17, 55. [Google Scholar] [CrossRef]
- Kockler, J.; Oelgemöller, M.; Robertson, S.; Glass, B.D. Photostability of sunscreens. J. Photochem. Photobiol. C Photochem. Rev. 2012, 13, 91–110. [Google Scholar] [CrossRef]
- Couteau, C.; Faure, A.; Fortin, J.; Paparis, E.; Coiffard, L.J. Coiffard, Study of the photostability of 18 sunscreens in creams by measuring the SPF in vitro. J. Pharm. Biomed. Anal. 2007, 44, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Jesus, A.; Sousa, E.; Cruz, M.T.; Cidade, H.; Lobo, J.M.S.; Almeida, I.F. UV Filters: Challenges and Prospects. Pharmaceuticals 2022, 15, 263. [Google Scholar] [CrossRef] [PubMed]
- Afonso, S.; Horita, K.; Sousa e Silva, J.P.; Almeida, I.F.; Amaral, M.H.; Lobão, P.A.; Costa, P.C.; Miranda, M.S.; Esteves da Silva, J.C.G.; Sousa Lobo, J.M. Photodegradation of avobenzone: Stabilization effect of antioxidants. J. Photochem. Photobiol. B Biol. 2014, 140, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.; Bs, C.P.; Smith, J.; Mesinkovska, N.A. The banned sunscreen ingredients and their impact on human health: A systematic review. Int. J. Dermatol. 2020, 59, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Santhra Krishnan, P.; Salian, A.; Dutta, S.; Mandal, S. A roadmap to UV-protective natural resources: Classification, characteristics, and applications. Mater. Chem. Front. 2021, 5, 7696–7723. [Google Scholar]
- Mansuri, R.; Diwan, A.; Kumar, H.; Dangwal, K.; Yadav, D. Potential of Natural Compounds as Sunscreen Agents. Pharmacogn. Rev. 2021, 15, 47–56. [Google Scholar] [CrossRef]
- Wong, H.J.; Mohamad-Fauzi, N.; Rizman-Idid, M.; Convey, P.; Alias, S.A. Protective mechanisms and responses of micro-fungi towards ultraviolet-induced cellular damage. Polar Sci. 2019, 20, 19–34. [Google Scholar] [CrossRef]
- Leach, C.M. Ultraviolet-absorbing substances associated with light-induced sporulation in fungi. Can. J. Bot. 1965, 43, 185–200. [Google Scholar] [CrossRef]
- Korać, R.R.; Khambholja, K.M. Potential of herbs in skin protection from ultraviolet radiation. Pharmacogn. Rev. 2011, 5, 164–173. [Google Scholar] [CrossRef]
- Wei, M.; Qiu, H.; Zhou, J.; Yang, C.; Chen, Y.; You, L. The Anti-Photoaging Activity of Peptides from Pinctada martensii Meat. Mar. Drugs 2022, 20, 770. [Google Scholar] [CrossRef]
- Ghazi, S. Do the polyphenolic compounds from natural products can protect the skin from ultraviolet rays? Results Chem. 2022, 4, 100428. [Google Scholar] [CrossRef]
- Smaoui, S.; Ben Hlima, H.; Ben Chobba, I.; Kadri, A. Development and stability studies of sunscreen cream formulations containing three photo-protective filters. Arab. J. Chem. 2017, 10, S1216–S1222. [Google Scholar] [CrossRef]
- Yarovaya, L.; Waranuch, N.; Wisuitiprot, W.; Khunkitti, W. Chemical and mechanical accelerated and long-term stability evaluation of sunscreen formulation containing grape seed extract. J. Cosmet. Dermatol. 2022, 21, 6400–6413. [Google Scholar] [CrossRef] [PubMed]
- Mancebo, S.E.; Hu, J.Y.; Wang, S.Q. Sunscreens: A review of health benefits, regulations, and controversies. Dermatol. Clin. 2014, 32, 427–438. [Google Scholar] [CrossRef]
- Zhu, X.; Li, N.; Wang, Y.; Ding, L.; Chen, H.; Yu, Y.; Shi, X. Protective effects of quercetin on UVB irradiation-induced cytotoxicity through ROS clearance in keratinocyte cells. Oncol. Rep. 2017, 37, 209–218. [Google Scholar] [CrossRef]
- Jung, S.K.; Lee, K.W.; Kim, H.Y.; Oh, M.H.; Byun, S.; Lim, S.H.; Heo, Y.-S.; Kang, N.J.; Bode, A.M.; Dong, Z.; et al. Myricetin suppresses UVB-induced wrinkle formation and MMP-9 expression by inhibiting Raf. Biochem. Pharmacol. 2010, 79, 1455–1461. [Google Scholar] [CrossRef]
- Daré, R.G.; Nakamura, C.V.; Ximenes, V.F.; Lautenschlager, S.O. Tannic acid, a promising anti-photoaging agent: Evidences of its antioxidant and anti-wrinkle potentials, and its ability to prevent photodamage and MMP-1 expression in L929 fibroblasts exposed to UVB. Free Radic. Biol. Med. 2020, 160, 342–355. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Incharoensakdi, A. Analysis of UV-absorbing photoprotectant mycosporine-like amino acid (MAA) in the cyanobacterium Arthrospira sp. CU2556. Photochem. Photobiol. Sci. 2014, 13, 1016–1024. [Google Scholar] [CrossRef]
- Lawrence, K.; Gacesa, R.; Long, P.; Young, A. Molecular photoprotection of human keratinocytes in vitro by the naturally occurring mycosporine-like amino acid palythine. Br. J. Dermatol. 2017, 178, 1353–1363. [Google Scholar] [CrossRef]
- Lionetti, N.; Rigano, L. The New Sunscreens among Formulation Strategy, Stability Issues, Changing Norms, Safety and Efficacy Evaluations. Cosmetics 2017, 4, 15. [Google Scholar] [CrossRef]
- Del Olmo, M.; Navarro, À.; Garcia, C.; Ehara, T.; Beltran, L. Effects of Structure on the Solubility of UV Filters. Cosmetics 2022, 9, 60. [Google Scholar] [CrossRef]
- Chisvert, A.; Salvador, A. Determination of water-soluble UV-filters in sunscreen sprays by liquid chromatography. J. Chromatogr. A 2002, 977, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Rajnochová Svobodová, A.; Ryšavá, A.; Čížková, K.; Roubalová, L.; Ulrichová, J.; Vrba, J.; Zálešák, B.; Vostálová, J. Effect of the flavonoids quercetin and taxifolin on UVA-induced damage to human primary skin keratinocytes and fibroblasts. Photochem. Photobiol. Sci. 2022, 21, 59. [Google Scholar] [CrossRef]
- Horwitz, R.J. Chapter 30—The Allergic Patient, in Integrative Medicine, 4th ed.; Rakel, D., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; p. 300. [Google Scholar]
- David, A.V.A.; Arulmoli, R.; Parasuraman, S. Overviews of biological importance of quercetin: A bioactive flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar]
- Zhao, B.; Wang, L.; Pang, S.; Jia, Z.; Wang, L.; Li, W.; Jin, B. UV-B promotes flavonoid synthesis in Ginkgo biloba leaves. Ind. Crop. Prod. 2020, 151, 112483. [Google Scholar] [CrossRef]
- Sánchez-Marzo, N.; Pérez-Sánchez, A.; Ruiz-Torres, V.; Martínez-Tébar, A.; Castillo, J.; Herranz-López, M.; Barrajón-Catalán, E. Antioxidant and Photoprotective Activity of Apigenin and its Potassium Salt Derivative in Human Keratinocytes and Absorption in Caco-2 Cell Monolayers. Int. J. Mol. Sci. 2019, 20, 2148. [Google Scholar] [CrossRef]
- Singh, D.; Kumari, K.; Ahmed, S. CHAPTER 17—Natural herbal products for cancer therapy. In Understanding Cancer; Jain, B., Pandey, S., Eds.; Academic Press: Cambridge, MA, USA, 2022; p. 257. [Google Scholar]
- Venigalla, M.; Gyengesi, E.; Münch, G. Curcumin and Apigenin—novel and promising therapeutics against chronic neuroinflammation in Alzheimer’s disease. Neural Regen. Res. 2015, 10, 1181. [Google Scholar]
- Sah, A.; Naseef, P.P.; Kuruniyan, M.S.; Jain, G.K.; Zakir, F.; Aggarwal, G. A Comprehensive Study of Therapeutic Applications of Chamomile. Pharmaceuticals 2022, 15, 1284. [Google Scholar] [CrossRef]
- Park, K.-S.; Chong, Y.; Kim, M.K. Myricetin: Biological activity related to human health. Appl. Biol. Chem. 2016, 59, 259–269. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Zhou, S.-S.; Shen, C.-Y.; Jiang, J.-G. Isolation and identification of four antioxidants from Rhodiola crenulata and evaluation of their UV photoprotection capacity in vitro. J. Funct. Foods 2020, 66, 103825. [Google Scholar] [CrossRef]
- Marotta, P.; Grossini, E.; Farruggio, S.; Panella, M. Chapter 25—Antiaging effects of natural agents in the skin: Focus on mitochondria. In Mitochondrial Physiology and Vegetal Molecules; de Oliveira, M.R., Ed.; Academic Press: Cambridge, MA, USA, 2021; p. 557. [Google Scholar]
- Oi, N.; Chen, H.; Kim, M.O.; Lubet, R.A.; Bode, A.M.; Dong, Z. Taxifolin suppresses UV-induced skin carcinogenesis by targeting EGFR and PI3K. Cancer Prev. Res. 2012, 5, 1103–1114. [Google Scholar] [CrossRef]
- Topal, F.; Nar, M.; Gocer, H.; Kalin, P.; Kocyigit, U.M.; Gülçin, İ.; Alwasel, S.H. Antioxidant activity of taxifolin: An activity-structure relationship. J. Enzym. Inhib. Med. Chem. 2016, 31, 674–683. [Google Scholar] [CrossRef] [PubMed]
- OyetakinWhite, P.; Tribout, H.; Baron, E. Protective Mechanisms of Green Tea Polyphenols in Skin. Oxid. Med. Cell. Longev. 2012, 2012, 560682. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Quon, M.J.; Kim, J.-A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Nair, H.H.; Alex, V.V.; Anto, R.J. 14—Significance of nutraceuticals in cancer therapy. In Evolutionary Diversity as a Source for Anticancer Molecules, Srivastava, A.K., Kannaujiya, V.K., Singh, R.K., Singh, D., Eds.; Academic Press: Cambridge, MA, USA, 2021; p. 309. [Google Scholar]
- Robles, H. Tannic Acid. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014; p. 474. [Google Scholar]
- Chan, C.-M.; Huang, C.-H.; Li, H.-J.; Hsiao, C.-Y.; Su, C.-C.; Lee, P.-L.; Hung, C.-F. Protective Effects of Resveratrol against UVA-Induced Damage in ARPE19 Cells. Int. J. Mol. Sci. 2015, 16, 5789–5802. [Google Scholar] [CrossRef]
- Li, Z.; Chen, X.; Liu, G.; Li, J.; Zhang, J.; Cao, Y.; Miao, J. Antioxidant Activity and Mechanism of Resveratrol and Polydatin Isolated from Mulberry (Morus alba L.). Molecules 2021, 26, 7574. [Google Scholar] [CrossRef]
- Weiskirchen, S.; Weiskirchen, R. Resveratrol: How Much Wine Do You Have to Drink to Stay Healthy? Adv. Nutr. 2016, 7, 706–718. [Google Scholar] [CrossRef]
- Kolb, C.A.; Kopecký, J.; Riederer, M.; Pfündel, E.E. UV screening by phenolics in berries of grapevine (Vitis vinifera). Funct. Plant Biol. 2003, 30, 1177–1186. [Google Scholar] [CrossRef]
- Deng, H.; Wan, M.; Li, H.; Chen, Q.; Li, R.; Liang, B.; Zhu, H. Curcumin protection against ultraviolet-induced photo-damage in Hacat cells by regulating nuclear factor erythroid 2-related factor 2. Bioengineered 2021, 12, 9993–10006. [Google Scholar] [CrossRef]
- Adusumilli, N.C.; Mordorski, B.; Nosanchuk, J.; Friedman, J.M.; Friedman, A.J. Curcumin nanoparticles as a photoprotective adjuvant. Exp. Dermatol. 2021, 30, 705–709. [Google Scholar] [CrossRef]
- Saleh, M.M.; Lawrence, K.P.; Jones, S.A.; Young, A.R. The photoprotective properties of α-tocopherol phosphate against long-wave UVA1 (385 nm) radiation in keratinocytes in vitro. Sci. Rep. 2021, 11, 22400. [Google Scholar] [CrossRef] [PubMed]
- Keen, M.A.; Hassan, I. Vitamin E in dermatology. Indian Dermatol. Online J. 2016, 7, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Astley, S.B. ANTIOXIDANTS|Role of Antioxidant Nutrients in Defense Systems. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; p. 282. [Google Scholar]
- Catanzaro, E.; Bishayee, A.; Fimognari, C. On a Beam of Light: Photoprotective Activities of the Marine Carotenoids Astaxanthin and Fucoxanthin in Suppression of Inflammation and Cancer. Mar. Drugs 2020, 18, 544. [Google Scholar] [CrossRef] [PubMed]
- Aneesh, P.; Ajeeshkumar, K.; Lekshmi, R.; Anandan, R.; Ravishankar, C.; Mathew, S. Bioactivities of astaxanthin from natural sources, augmenting its biomedical potential: A review. Trends Food Sci. Technol. 2022, 125, 81–90. [Google Scholar] [CrossRef]
- Tominaga, K.; Hongo, N.; Fujishita, M.; Takahashi, Y.; Adachi, Y. Protective effects of astaxanthin on skin deterioration. J. Clin. Biochem. Nutr. 2017, 61, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, W.C.; Yamamoto, Y. Small-molecule antioxidants in marine organisms: Antioxidant activity of mycosporine-glycine. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1995, 112, 105–114. [Google Scholar] [CrossRef]
- Portwich, A.; Garcia-Pichel, F. A novel prokaryotic UVB photoreceptor in the cyanobacterium Chlorogloeopsis PCC 6912. Photochem. Photobiol. 2000, 71, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Sommaruga, R.; Garcia-Pichel, F. UV-absorbing mycosporine-like compounds in planktonic and benthic organisms from a high-mountain lake. Fundam. Appl. Limnol. 1999, 144, 255–269. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Bilger, W.; Scherer, S. UV-B-induced synthesis of photoprotective pigments and extracellular polysaccharides in the terrestrial cyanobacterium Nostoc commune. J. Bacteriol. 1997, 179, 1940–1945. [Google Scholar] [CrossRef]
- Karsten, U.; Sawall, T.; Wiencke, C. A survey of the distribution of UV-absorbing substances in tropical macroalgae. Phycol. Res. 1998, 46, 271–279. [Google Scholar]
- Karsten, U.; Sawall, T.; West, J.; Wiencke, C. Ultraviolet sunscreen compounds in epiphytic red algae from mangroves. Hydrobiologia 2000, 432, 159–171. [Google Scholar] [CrossRef]
- Obermüller, B.; Karsten, U.; Abele, D. Response of oxidative stress parameters and sunscreening compounds in Arctic amphipods during experimental exposure to maximal natural UVB radiation. J. Exp. Mar. Biol. Ecol. 2005, 323, 100–117. [Google Scholar] [CrossRef]
- Ngoennet, S.; Nishikawa, Y.; Hibino, T.; Waditee-Sirisattha, R.; Kageyama, H. A Method for the Isolation and Characterization of Mycosporine-Like Amino Acids from Cyanobacteria. Methods Protoc. 2018, 1, 46. [Google Scholar] [CrossRef] [PubMed]
- Kedar, L.; Kashman, Y.; Oren, A. Mycosporine-2-glycine is the major mycosporine-like amino acid in a unicellular cyanobacterium (Euhalothece sp.) isolated from a gypsum crust in a hypersaline saltern pond. FEMS Microbiol. Lett. 2002, 208, 233–237. [Google Scholar]
- Waditee-Sirisattha, R.; Kageyama, H.; Sopun, W.; Tanaka, Y.; Takabe, T. Identification and upregulation of biosynthetic genes required for accumulation of mycosporine-2-glycine under salt stress conditions in the halotolerant cyanobacterium Aphanothece halophytica. Appl. Environ. Microbiol. 2014, 80, 1763–1769. [Google Scholar] [CrossRef]
- Shick, J.M.; Dunlap, W.C.; Pearse, J.S.; Pearse, V.B. Mycosporine-like Amino Acid Content in Four Species of Sea Anemones in the Genus Anthopleura Reflects Phylogenetic but Not Environmental or Symbiotic Relationships. Biol. Bull. 2002, 203, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Sommaruga, R.; Whitehead, K.; Shick, J.M.; Lobban, C.S. Mycosporine-like Amino Acids in the Zooxanthella-Ciliate Symbiosis Maristentor dinoferus. Protist 2006, 157, 185–191. [Google Scholar] [CrossRef]
- Kim, S.; Seock, Y.-K. Impacts of health and environmental consciousness on young female consumers’ attitude towards and purchase of natural beauty products. Int. J. Consum. Stud. 2009, 33, 627–638. [Google Scholar] [CrossRef]
- Schmidt, E.W. An enzymatic route to sunscreens. ChemBioChem 2011, 12, 363–365. [Google Scholar] [CrossRef]
- Meinhardt, M.; Krebs, R.; Anders, A.; Heinrich, U.; Tronnier, H. Wavelength-dependent penetration depths of ultraviolet radiation in human skin. J. Biomed. Opt. 2008, 13, 044030. [Google Scholar] [CrossRef]
- Álvarez-Gómez, F.; Korbee, N.; Casas-Arrojo, V.; Abdala-Díaz, R.T.; Figueroa, F.L. UV Photoprotection, Cytotoxicity and Immunology Capacity of Red Algae Extracts. Molecules 2019, 24, 341. [Google Scholar] [CrossRef] [PubMed]
- Ben Ghedifa, A.; Vega, J.; Korbee, N.; Mensi, F.; Figueroa, F.L.; Sadok, S. Effects of light quality on the photosynthetic activity and biochemical composition of Gracilaria gracilis (Rhodophyta). J. Appl. Phycol. 2021, 33, 3413–3425. [Google Scholar] [CrossRef]
- Schneider, G.; Figueroa, F.L.; Vega, J.; Avilés, A.; Horta, P.A.; Korbee, N.; Bonomi-Barufi, J. Effects of UV–visible radiation on growth, photosynthesis, pigment accumulation and UV-absorbing compounds in the red macroalga Gracilaria cornea (Gracilariales, Rhodophyta). Algal Res. 2022, 64, 102702. [Google Scholar] [CrossRef]
- Kräbs, G.; Bischof, K.; Hanelt, D.; Karsten, U.; Wiencke, C. Wavelength-dependent induction of UV-absorbing mycosporine-like amino acids in the red alga Chondrus crispus under natural solar radiation. J. Exp. Mar. Biol. Ecol. 2002, 268, 69–82. [Google Scholar] [CrossRef]
- Bonomi-Barufi, J.; Figueroa, F.L.; Korbee, N.; Momoli, M.M.; Martins, A.P.; Colepicolo, P.; Van Sluys, M.-A.; Oliveira, M.C. How macroalgae can deal with radiation variability and photoacclimation capacity: The example of Gracilaria tenuistipitata (Rhodophyta) in laboratory. Algal Res. 2020, 50, 102007. [Google Scholar] [CrossRef]
- Franklin, L.A.; Kräbs, G.; Kuhlenkamp, R. Blue light and uv-a radiation control the synthesis of mycosporine-like amino acids in Chondrus crispus (florideophyceae). J. Phycol. 2001, 37, 257–270. [Google Scholar] [CrossRef]
- Gulston, M.; Knowland, J. Illumination of human keratinocytes in the presence of the sunscreen ingredient Padimate-O and through an SPF-15 sunscreen reduces direct photodamage to DNA but increases strand breaks. Mutat. Res. 1999, 444, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Duale, N.; Olsen, A.-K.; Christensen, T.; Butt, S.T.; Brunborg, G. Octyl Methoxycinnamate Modulates Gene Expression and Prevents Cyclobutane Pyrimidine Dimer Formation but not Oxidative DNA Damage in UV-Exposed Human Cell Lines. Toxicol. Sci. 2010, 114, 272–284. [Google Scholar] [CrossRef]
- Menon, G.K. New insights into skin structure: Scratching the surface. Adv. Drug Deliv. Rev. 2002, 54, S3–S17. [Google Scholar] [CrossRef]
- Schuch, A.P.; Moraes, M.C.S.; Yagura, T.; Menck, C.F.M. Highly Sensitive Biological Assay for Determining the Photoprotective Efficacy of Sunscreen. Environ. Sci. Technol. 2014, 48, 11584. [Google Scholar] [CrossRef]
- Bernerd, F.; Vioux, C.; Asselineau, D. Evaluation of the protective effect of sunscreens on in vitro reconstructed human skin exposed to UVB or UVA irradiation. Photochem. Photobiol. 2000, 71, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Bernerd, F.; Vioux, C.; Lejeune, F.; Asselineau, D. The sun protection factor (SPF) inadequately defines broad spectrum photoprotection: Demonstration using skin reconstructed in vitro exposed to UVA, UVBor UV-solar simulated radiation. Eur. J. Dermatol. 2003, 13, 242–249. [Google Scholar] [PubMed]
- Mouret, S.; Bogdanowicz, P.; Haure, M.-J.; Castex-Rizzi, N.; Cadet, J.; Favier, A.; Douki, T. Assessment of the Photoprotection Properties of Sunscreens by Chromatographic Measurement of DNA Damage in Skin Explants. Photochem. Photobiol. 2010, 87, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Cole, C.A.; Forbes, P.D.; Ludwigsen, K. Sunscreen testing using the mouse ear model. Photo-dermatology 1989, 6, 131–136. [Google Scholar]
- Ley, R.D.; Fourtanier, A. Sunscreen protection against ultraviolet radiation-induced pyrimidine dimers in mouse epidermal DNA. Photochem. Photobiol. 1997, 65, 1007–1011. [Google Scholar] [CrossRef]
- Klug, H.L.P.; Tooze, J.A.; Graff-Cherry, C.; Anver, M.R.; Noonan, F.; Fears, T.R.; Tucker, M.A.; De Fabo, E.C.; Merlino, G. Sunscreen prevention of melanoma in man and mouse. Pigment. Cell Melanoma Res. 2010, 23, 835–837. [Google Scholar] [CrossRef]
- Viros, A.; Sanchez-Laorden, B.; Pedersen, M.; Furney, S.J.; Rae, J.; Hogan, K.; Ejiama, S.; Girotti, M.R.; Cook, M.; Dhomen, N.; et al. Ultraviolet radiation accelerates BRAF-driven melanomagenesis by targeting TP53. Nature 2014, 511, 478–482. [Google Scholar] [CrossRef]
- Horiki, S.; Miyauchi-Hashimoto, H.; Tanaka, K.; Nikaido, O.; Horio, T. Protective effects of sunscreening agents on photocarcinogenesis, photoaging, and DNA damage in XPA gene knockout mice. Arch. Dermatol. Res. 2000, 292, 511–518. [Google Scholar] [CrossRef]
- Duracher, L.; Blasco, L.; Abdel Jaoued, A.; Vian, L.; Marti-Mestres, G. Irradiation of skin and contrasting effects on absorption of hydrophilic and lipophilic compounds. Photochem. Photobiol. 2009, 85, 1459–1467. [Google Scholar] [CrossRef]
- Wu, J.; Liu, W.; Xue, C.; Zhou, S.; Lan, F.; Bi, L.; Xu, H.; Yang, X.; Zeng, F.-D. Toxicity and penetration of TiO2 nanoparticles in hairless mice and porcine skin after subchronic dermal exposure. Toxicol. Lett. 2009, 191, 1–8. [Google Scholar] [CrossRef]
- Liardet, S.; Scaletta, C.; Panizzon, R.; Hohlfeld, P.; Laurent-Applegate, L. Protection against pyrimidine dimers, p53, and 8-hydroxy-2’-deoxyguanosine expression in ultraviolet-irradiated human skin by sunscreens: Difference between UVB + UVA and UVB alone sunscreens. J. Investig. Dermatol. 2001, 117, 1437–1441. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, K.H.; Guaratini, T.; Barros, M.P.; Falcão, V.R.; Tonon, A.P.; Lopes, N.P.; Campos, S.; Torres, M.A.; Souza, A.O.; Colepicolo, P.; et al. Metabolites from algae with economical impact. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 146, 60–78. [Google Scholar] [CrossRef] [PubMed]
- Dextro, R.B.; Delbaje, E.; Geraldes, V.; Pinto, E.; Long, P.F.; Fiore, M.F. Exploring the Relationship between Biosynthetic Gene Clusters and Constitutive Production of Mycosporine-like Amino Acids in Brazilian Cyanobacteria. Molecules 2023, 28, 1420. [Google Scholar] [CrossRef] [PubMed]
- Milito, A.; Castellano, I.; Damiani, E. From Sea to Skin: Is There a Future for Natural Photoprotectants? Mar. Drugs 2021, 19, 379. [Google Scholar] [CrossRef]
- Balskus, E.P.; Walsh, C.T. The genetic and molecular basis for sunscreen biosynthesis in cyanobacteria. Science 2010, 329, 1653–1656. [Google Scholar] [CrossRef]
- Portwich, A.; Garcia-Pichel, F. Biosynthetic pathway of mycosporines (mycosporine-like amino acids) in the cyanobacterium Chlorogloeopsis sp. strain PCC 6912. Phycologia 2003, 42, 384–392. [Google Scholar]
- Miyamoto, K.T.; Komatsu, M.; Ikeda, H. Discovery of gene cluster for mycosporine-like amino acid biosynthesis from Actinomycetales microorganisms and production of a novel mycosporine-like amino acid by heterologous expression. Appl. Environ. Microbiol. 2014, 80, 5028–5036. [Google Scholar] [CrossRef]
- Rosic, N. Genome Mining as an Alternative Way for Screening the Marine Organisms for Their Potential to Produce UV-Absorbing Mycosporine-like Amino Acid. Mar. Drugs 2022, 20, 478. [Google Scholar] [CrossRef]
- D’Agostino, P.M.; Woodhouse, J.N.; Liew, H.T.; Sehnal, L.; Pickford, R.; Wong, H.L.; Burns, B.P.; Neilan, B.A. Bioinformatic, phylogenetic and chemical analysis of the UV-absorbing compounds scytonemin and mycosporine-like amino acids from the microbial mat communities of Shark Bay, Australia. Environ. Microbiol. 2019, 21, 702–715. [Google Scholar] [CrossRef]
- Ambrosino, L.; Tangherlini, M.; Colantuono, C.; Esposito, A.; Sangiovanni, M.; Miralto, M.; Sansone, C.; Chiusano, M.L. Bioinformatics for Marine Products: An Overview of Resources, Bottlenecks, and Perspectives. Mar. Drugs 2019, 17, 576. [Google Scholar] [CrossRef]
- Mogany, T.; Kumari, S.; Swalaha, F.M.; Bux, F. In silico analysis of enzymes involved in mycosporine-like amino acids biosynthesis in Euhalothece sp.: Structural and functional characterization. Algal Res. 2022, 66, 102806. [Google Scholar] [CrossRef]
- Llewellyn, C.A.; Greig, C.; Silkina, A.; Kultschar, B.; Hitchings, M.D.; Farnham, G. Mycosporine-like amino acid and aromatic amino acid transcriptome response to UV and far-red light in the cyanobacterium Chlorogloeopsis fritschii PCC 6912. Sci. Rep. 2020, 10, 20638. [Google Scholar] [CrossRef] [PubMed]
- Rosic, N.N. Phylogenetic analysis of genes involved in mycosporine-like amino acid biosynthesis in symbiotic dinoflagellates. Appl. Microbiol. Biotechnol. 2012, 94, 29–37. [Google Scholar] [CrossRef]
- Shoguchi, E. Gene clusters for biosynthesis of mycosporine-like amino acids in dinoflagellate nuclear genomes: Possible recent horizontal gene transfer between species of Symbiodiniaceae (Dinophyceae). J. Phycol. 2022, 58, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Garcia-Pichel, F. An ATP-Grasp Ligase Involved in the Last Biosynthetic Step of the Iminomycosporine Shinorine in Nostoc punctiforme ATCC 29133. J. Bacteriol. 2011, 193, 5923–5928. [Google Scholar] [CrossRef]
- Hoyer, K.; Karsten, U.; Wiencke, C. Induction of sunscreen compounds in Antarctic macroalgae by different radiation conditions. Mar. Biol. 2002, 141, 619–627. [Google Scholar]
- Ingebrigtsen, R.A.; Hansen, E.; Andersen, J.H.; Eilertsen, H.C. Light and temperature effects on bioactivity in diatoms. J. Appl. Phycol. 2016, 28, 939–950. [Google Scholar] [CrossRef]
- Waditee-Sirisattha, R.; Kageyama, H.; Fukaya, M.; Rai, V.; Takabe, T. Nitrate and amino acid availability affects glycine betaine and mycosporine-2-glycine in response to changes of salinity in a halotolerant cyanobacterium Aphanothece halophytica. FEMS Microbiol. Lett. 2015, 362, fnv198. [Google Scholar] [CrossRef]
- Nishida, Y.; Miyabe, Y.; Kishimura, H.; Kumagai, Y. Monthly Variation and Ultraviolet Stability of Mycosporine-like Amino Acids from Red Alga Dulse Palmaria palmata in Japan. Phycology 2021, 1, 119–128. [Google Scholar] [CrossRef]
- Weiss, E.L.; Cape, M.R.; Pan, B.J.; Vernet, M.; James, C.C.; Smyth, T.J.; Ha, S.Y.; Iriarte, J.L.; Mitchell, B.G. The distribution of mycosporine-like amino acids in phytoplankton across a Southern Ocean transect. Front. Mar. Sci. 2022, 9, 2133. [Google Scholar] [CrossRef]
- Jofre, J.; Celis-Plá, P.S.M.; Figueroa, F.L.; Navarro, N.P. Seasonal Variation of Mycosporine-Like Amino Acids in Three Subantarctic Red Seaweeds. Mar. Drugs 2020, 18, 75. [Google Scholar] [CrossRef] [PubMed]

| Fitzpatrick Skin Type | Typical Features | Tanning Ability and Sensitivity | Skin Cancer Risk * |
|---|---|---|---|
| I | Unexposed skin: white Eyes: blue or green Frequent freckling Northern European or British | Always burns with minimal UV exposure Peels Never tans | 4 |
| II | Unexposed skin: white Eyes: blue, hazel or brown Hair: red, blonds or brown European or Scandinavian | Burns easily Peels Tans minimally | 3–4 |
| III | Unexposed skin: fair Eyes: brown; Hair: dark Southern or Central European | Burns moderately Average tanning ability | 3 |
| IV | Unexposed skin: light brown Eyes: dark; Hair: dark Mediterranean, Asian, or Latino | Burns minimally Tans easily | 2 |
| V | Unexposed skin: brown Eyes: dark; Hair: dark East Indian, Native American, Latino, or African | Rarely burns Tans easily and substantially | 1 |
| VI | Unexposed skin: black Eyes: dark; Hair: dark African or Aboriginal | Almost never burns Tans readily and profusely | 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosic, N.; Climstein, M.; Boyle, G.M.; Thanh Nguyen, D.; Feng, Y. Exploring Mycosporine-like Amino Acid UV-Absorbing Natural Products for a New Generation of Environmentally Friendly Sunscreens. Mar. Drugs 2023, 21, 253. https://doi.org/10.3390/md21040253
Rosic N, Climstein M, Boyle GM, Thanh Nguyen D, Feng Y. Exploring Mycosporine-like Amino Acid UV-Absorbing Natural Products for a New Generation of Environmentally Friendly Sunscreens. Marine Drugs. 2023; 21(4):253. https://doi.org/10.3390/md21040253
Chicago/Turabian StyleRosic, Nedeljka, Mike Climstein, Glen M. Boyle, Duy Thanh Nguyen, and Yunjiang Feng. 2023. "Exploring Mycosporine-like Amino Acid UV-Absorbing Natural Products for a New Generation of Environmentally Friendly Sunscreens" Marine Drugs 21, no. 4: 253. https://doi.org/10.3390/md21040253