
Isolation and Characterization of Targeting-HBsAg VNAR Single Domain Antibodies from Whitespotted Bamboo Sharks (Chiloscyllium plagiosum)

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
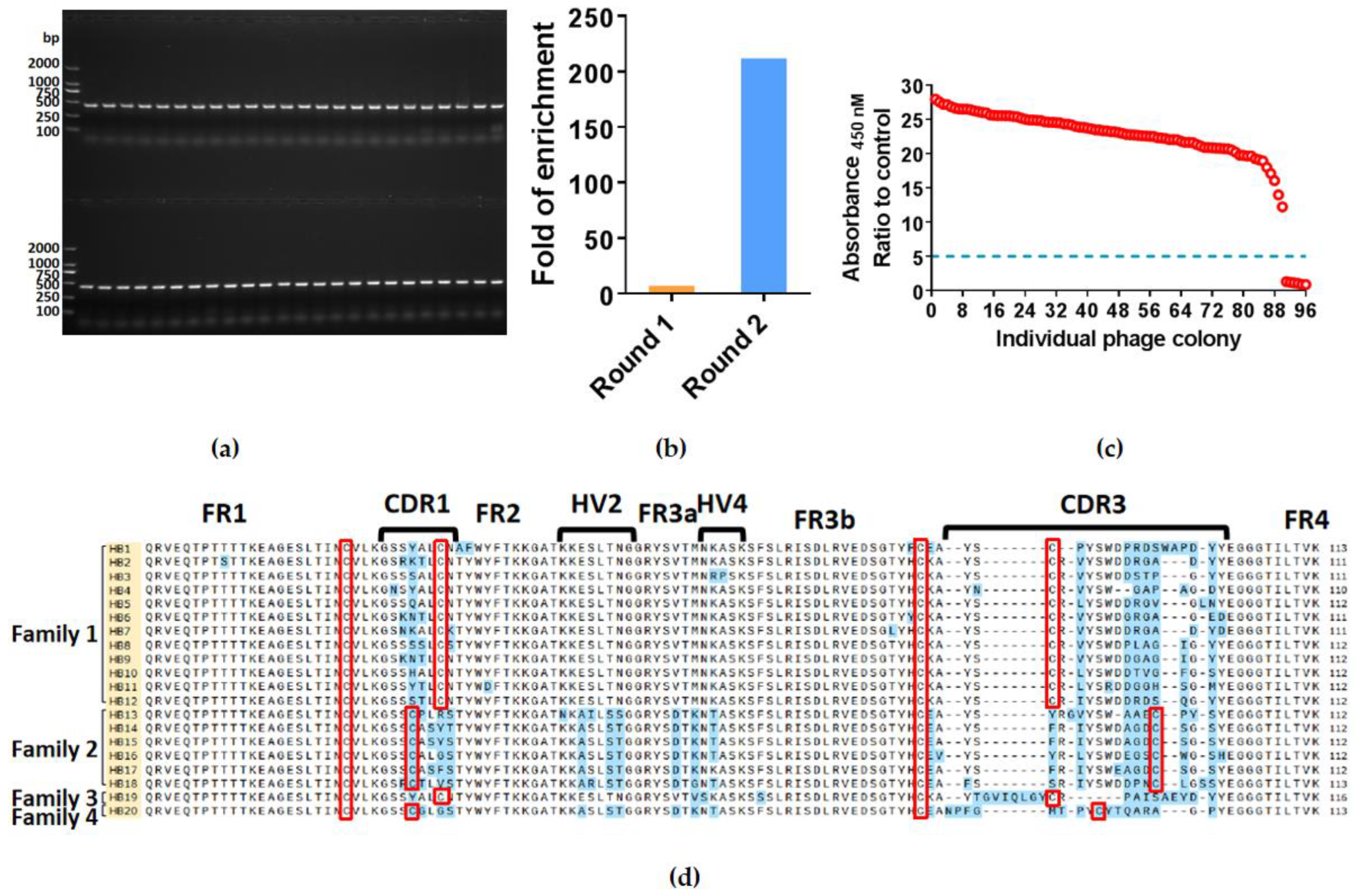
2.1. Isolation of Specific Nanobodies against HBsAg
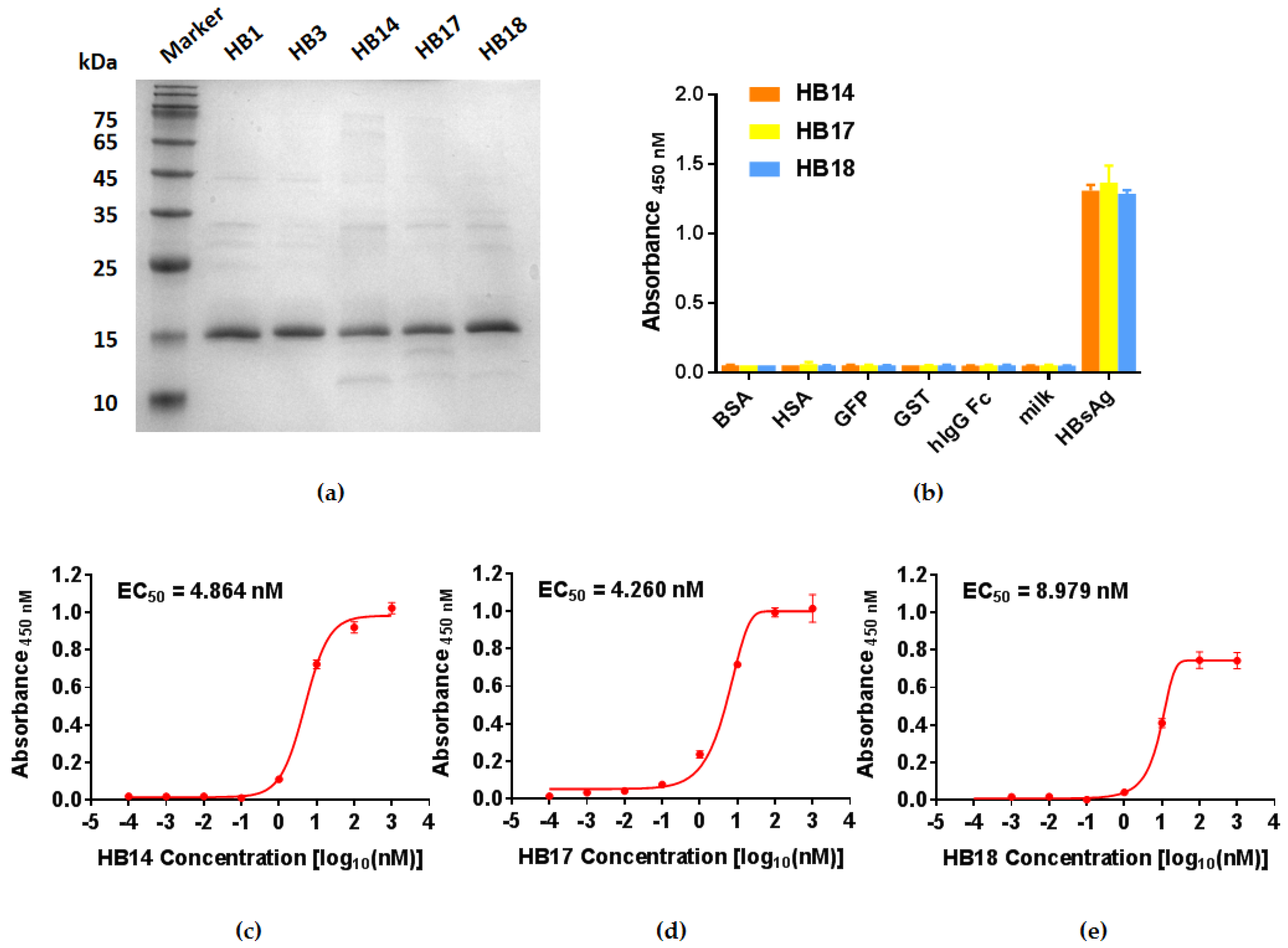
2.2. Expression of Soluble Single Domain Antibodies and Binding Potency Assay
2.3. Testing of HBsAg-Specific VNARs Available for Sandwich ELISA
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Immunization of Whitespotted Bamboo Sharks
4.3. VNAR Library Construction
4.4. Selection of HBsAg-Specific VNAR
4.5. Phage ELISA Assay
4.6. Expression and Purification of VNARs
4.7. Binding Specificity Assay for VNARs
4.8. ELISA Binding Potency Assay for VNARs
4.9. Sandwich ELISA for VNARs
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greenberg, A.S.; Avila, D.; Hughes, M.; Hughes, A.; McKinney, E.C.; Flajnik, M.F. A new antigen receptor gene family that undergoes rearrangement and extensive somatic diversification in sharks. Nature 1995, 374, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, J.; Alzogaray, V.; Reyelt, J.; Unger, M.; Juarez, K.; Urrutia, M.; Cauerhff, A.; Danquah, W.; Rissiek, B.; Scheuplein, F.; et al. Single domain antibodies: Promising experimental and therapeutic tools in infection and immunity. Med. Microbiol. Immunol. 2009, 198, 157–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielonka, S.; Empting, M.; Grzeschik, J.; Könning, D.; Barelle, C.J.; Kolmar, H. Structural insights and biomedical potential of IgNAR scaffolds from sharks. mAbs 2015, 7, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, M.; Bian, H.; Wu, X.; Fu, T.; Fu, Y.; Hong, J.; Fleming, B.D.; Flajnik, M.F.; Ho, M. Construction and next-generation sequencing analysis of a large phage-displayed V(NAR) single-domain antibody library from six naïve nurse sharks. Antib. Ther. 2019, 2, 1–11. [Google Scholar]
- Feige, M.J.; Gräwert, M.A.; Marcinowski, M.; Hennig, J.; Behnke, J.; Ausländer, D.; Herold, E.M.; Peschek, J.; Castro, C.D.; Flajnik, M.; et al. The structural analysis of shark IgNAR antibodies reveals evolutionary principles of immunoglobulins. Proc. Natl. Acad. Sci. USA 2014, 111, 8155–8160. [Google Scholar] [CrossRef] [Green Version]
- Zielonka, S.; Weber, N.; Becker, S.; Doerner, A.; Christmann, A.; Christmann, C.; Uth, C.; Fritz, J.; Schäfer, E.; Steinmann, B.; et al. Shark Attack: High affinity binding proteins derived from shark vNAR domains by stepwise in vitro affinity maturation. J. Biotechnol. 2014, 191, 236–245. [Google Scholar] [CrossRef]
- Juma, S.N.; Gong, X.; Hu, S.; Lv, Z.; Shao, J.; Liu, L.; Chen, G. Shark New Antigen Receptor (IgNAR): Structure, Characteristics and Potential Biomedical Applications. Cells 2021, 10, 1140. [Google Scholar] [CrossRef]
- English, H.; Hong, J.; Ho, M. Ancient species offers contemporary therapeutics: An update on shark V(NAR) single domain antibody sequences, phage libraries and potential clinical applications. Antib. Ther. 2020, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Matz, H.; Dooley, H. Shark IgNAR-derived binding domains as potential diagnostic and therapeutic agents. Dev. Comp. Immunol. 2019, 90, 100–107. [Google Scholar] [CrossRef]
- Cheong, W.S.; Leow, C.Y.; Abdul Majeed, A.B.; Leow, C.H. Diagnostic and therapeutic potential of shark variable new antigen receptor (VNAR) single domain antibody. Int. J. Biol. Macromol. 2020, 147, 369–375. [Google Scholar] [CrossRef]
- Pillay, T.S.; Muyldermans, S. Application of Single-Domain Antibodies ("Nanobodies") to Laboratory Diagnosis. Ann. Lab. Med. 2021, 41, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Goodchild, S.A.; Dooley, H.; Schoepp, R.J.; Flajnik, M.; Lonsdale, S.G. Isolation and characterisation of Ebolavirus-specific recombinant antibody fragments from murine and shark immune libraries. Mol. Immunol. 2011, 48, 2027–2037. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.P.; Teichler, D.D.; Zabetakis, D.; Shriver-Lake, L.C.; Liu, J.L.; Lonsdale, S.G.; Goodchild, S.A.; Goldman, E.R. Importance of Hypervariable Region 2 for Stability and Affinity of a Shark Single-Domain Antibody Specific for Ebola Virus Nucleoprotein. PLoS ONE 2016, 11, e0160534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, R.; Nuttall, S.; Revill, P.; Colledge, D.; Cabuang, L.; Soppe, S.; Dolezal, O.; Griffiths, K.; Bartholomeusz, A.; Locarnini, S. Targeting the hepatitis B virus precore antigen with a novel IgNAR single variable domain intrabody. Virology 2011, 411, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.L.; Anderson, G.P.; Delehanty, J.B.; Baumann, R.; Hayhurst, A.; Goldman, E.R. Selection of cholera toxin specific IgNAR single-domain antibodies from a naïve shark library. Mol. Immunol. 2007, 44, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- The Polaris Observatory Collaborators. Global prevalence, treatment, and prevention of hepatitis B virus infection in 2016: A modelling study. Lancet Gastroenterol. Hepatol. 2018, 3, 383–403. [Google Scholar] [CrossRef]
- Tseng, T.C.; Kao, J.H. Clinical utility of quantitative HBsAg in natural history and nucleos(t)ide analogue treatment of chronic hepatitis B: New trick of old dog. J. Gastroenterol. 2013, 48, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Kao, J.H.; Chen, P.J.; Chen, D.S. Recent advances in the research of hepatitis B virus-related hepatocellular carcinoma: Epidemiologic and molecular biological aspects. Adv. Cancer Res. 2010, 108, 21–72. [Google Scholar]
- Philips, C.A.; Ahamed, R.; Abduljaleel, J.K.; Rajesh, S.; Augustine, P. Critical Updates on Chronic Hepatitis B Virus Infection in 2021. Cureus 2021, 13, e19152. [Google Scholar] [CrossRef]
- Blumberg, B.S.; Sutnick, A.I.; London, W.T. Hepatitis and leukemia: Their relation to Australia antigen. Bull. N. Y. Acad. Med. 1968, 44, 1566–1586. [Google Scholar]
- Leyva, A.; Sánchez, J.C.; Valdes, R.; Font, M.; López, L.; Hernández, N.; Ferro, W.; Medina, Y. A combinatorial strategy of a new monoclonal ELISA and immunoaffinity chromatography using sodium deoxycholate to increase the recovery of multimeric proteins like r-HBsAg. Talanta 2010, 81, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Wolters, G.; Nelissen, P.; Kuijpers, L. Improved ELISA for the detection of HBsAg. J. Virol. Methods 1985, 10, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.; Fish, G.E.; Smith, M.B. Laboratory evaluation of a fully automated chemiluminescence immunoassay for rapid detection of HBsAg, antibodies to HBsAg, and antibodies to hepatitis C virus. J. Clin. Microbiol. 2004, 42, 610–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, J.; Cheng, L.; Yin, B.; Guan, Q.; Liu, Y.; Wu, S.; Wang, B.; Tang, N.; Zhang, B.; Wang, L.; et al. Development of an economic and efficient strategy to detect HBsAg: Application of "gray-zones" in ELISA and combined use of several detection assays. Clin. Chim. Acta Int. J. Clin. Chem. 2011, 412, 2046–2051. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Matsui, T.; Tanaka, Y. Novel strategies for the early diagnosis of hepatitis B virus reactivation. Hepatol. Res. Off. J. Jpn. Soc. Hepatol. 2021, 51, 1033–1043. [Google Scholar] [CrossRef]
- Tsurusawa, N.; Chang, J.; Namba, M.; Makioka, D.; Yamura, S.; Iha, K.; Kyosei, Y.; Watabe, S.; Yoshimura, T.; Ito, E. Modified ELISA for Ultrasensitive Diagnosis. J. Clin. Med. 2021, 10, 5197. [Google Scholar] [CrossRef]
- Hayrapetyan, H.; Tran, T.; Tellez-Corrales, E.; Madiraju, C. Enzyme-Linked Immunosorbent Assay: Types and Applications. Methods Mol. Biol. (Clifton N.J.) 2023, 2612, 1–17. [Google Scholar]
- Liu, J.K. The history of monoclonal antibody development—Progress, remaining challenges and future innovations. Ann. Med. Surg. 2014, 3, 113–116. [Google Scholar] [CrossRef]
- Ji, P.; Wang, K.; Zhang, L.; Yan, Z.; Kong, M.; Sun, X.; Zhang, Q.; Zhou, N.; Liu, B.; Zhou, E.M.; et al. A new nanobody-enzyme fusion protein-linked immunoassay for detecting antibodies against influenza A virus in different species. J. Biol. Chem. 2022, 298, 102709. [Google Scholar] [CrossRef]
- Stanker, L.H.; Hnasko, R.M. A Double-Sandwich ELISA for Identification of Monoclonal Antibodies Suitable for Sandwich Immunoassays. Methods Mol. Biol. (Clifton N.J.) 2015, 1318, 69–78. [Google Scholar]
- Dooley, H.; Flajnik, M.F. Shark immunity bites back: Affinity maturation and memory response in the nurse shark, Ginglymostoma cirratum. Eur. J. Immunol. 2005, 35, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Leow, C.H.; Jones, M.; Cheng, Q.; Mahler, S.; McCarthy, J. Production and characterization of specific monoclonal antibodies binding the Plasmodium falciparum diagnostic biomarker, histidine-rich protein 2. Malar. J. 2014, 13, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Chen, M.; Wang, X.; Kang, S.; Xue, W.; Li, Z. Identification of Anti-TNFα VNAR Single Domain Antibodies from Whitespotted Bambooshark (Chiloscyllium plagiosum). Mar. Drugs 2022, 20, 307. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, X.; Sun, S.; Li, Z.; Chen, M. Isolation and Characterization of Targeting-HBsAg VNAR Single Domain Antibodies from Whitespotted Bamboo Sharks (Chiloscyllium plagiosum). Mar. Drugs 2023, 21, 237. https://doi.org/10.3390/md21040237
Jiang X, Sun S, Li Z, Chen M. Isolation and Characterization of Targeting-HBsAg VNAR Single Domain Antibodies from Whitespotted Bamboo Sharks (Chiloscyllium plagiosum). Marine Drugs. 2023; 21(4):237. https://doi.org/10.3390/md21040237
Chicago/Turabian StyleJiang, Xierui, Shan Sun, Zengpeng Li, and Mingliang Chen. 2023. "Isolation and Characterization of Targeting-HBsAg VNAR Single Domain Antibodies from Whitespotted Bamboo Sharks (Chiloscyllium plagiosum)" Marine Drugs 21, no. 4: 237. https://doi.org/10.3390/md21040237