
Bioconversion of Cheese Whey and Food By-Products by Phaeodactylum tricornutum into Fucoxanthin and n-3 Lc-PUFA through a Biorefinery Approach

 ,
,  , ,
, ,  ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
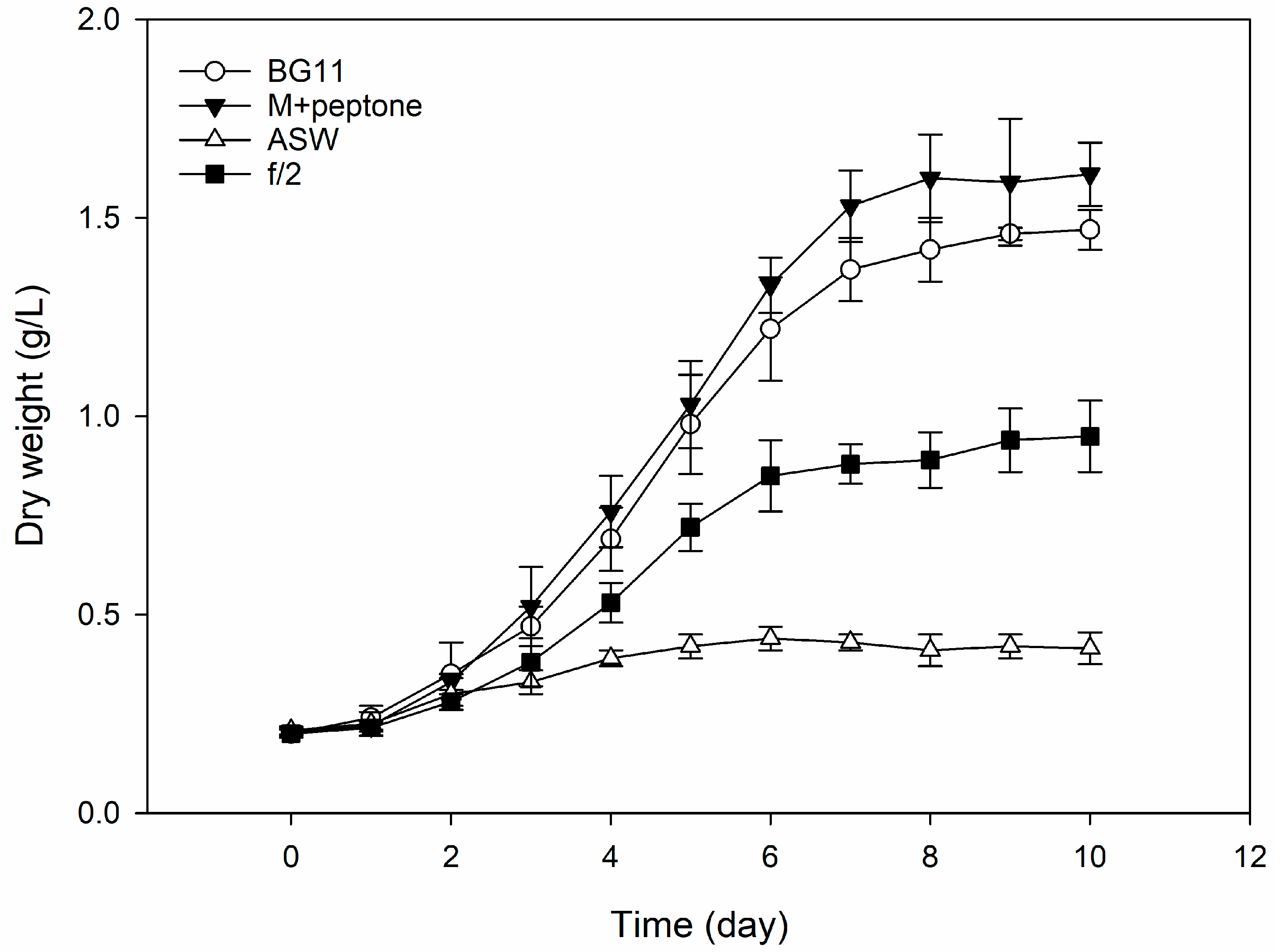
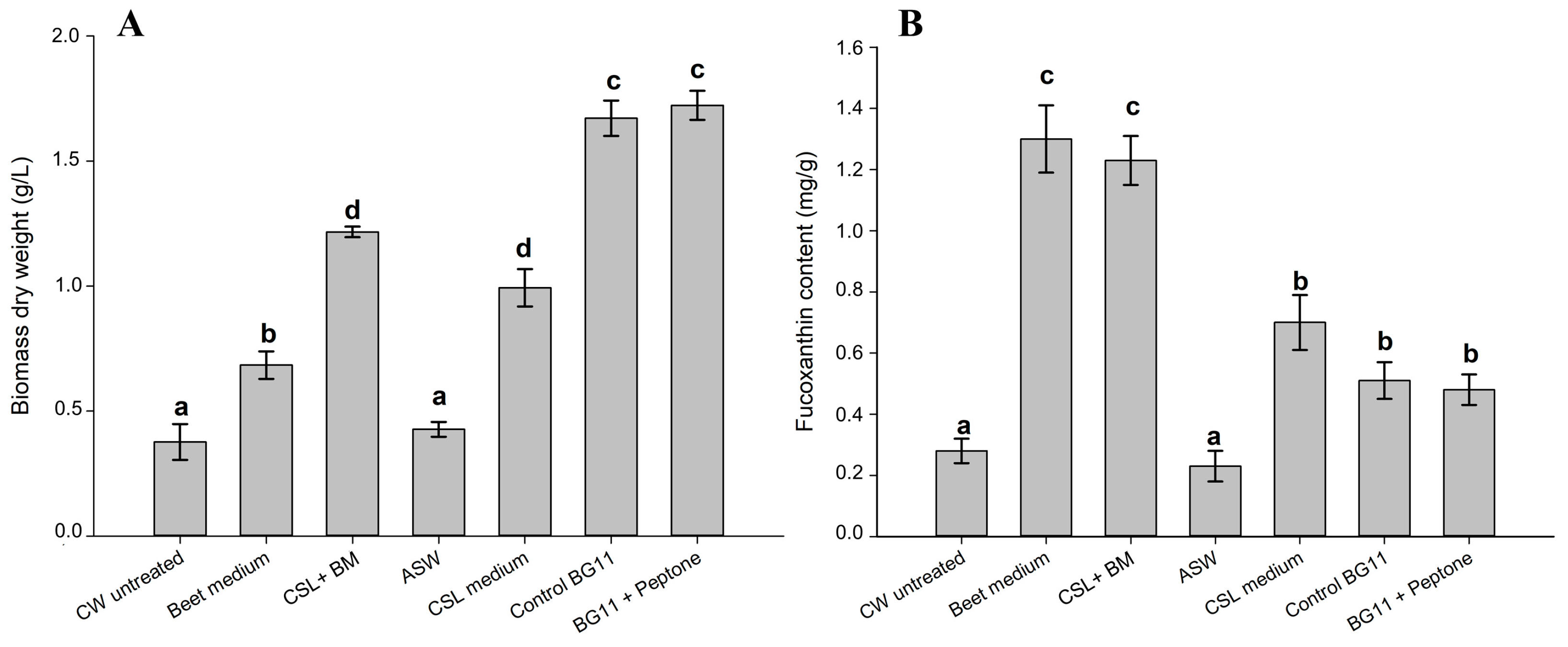
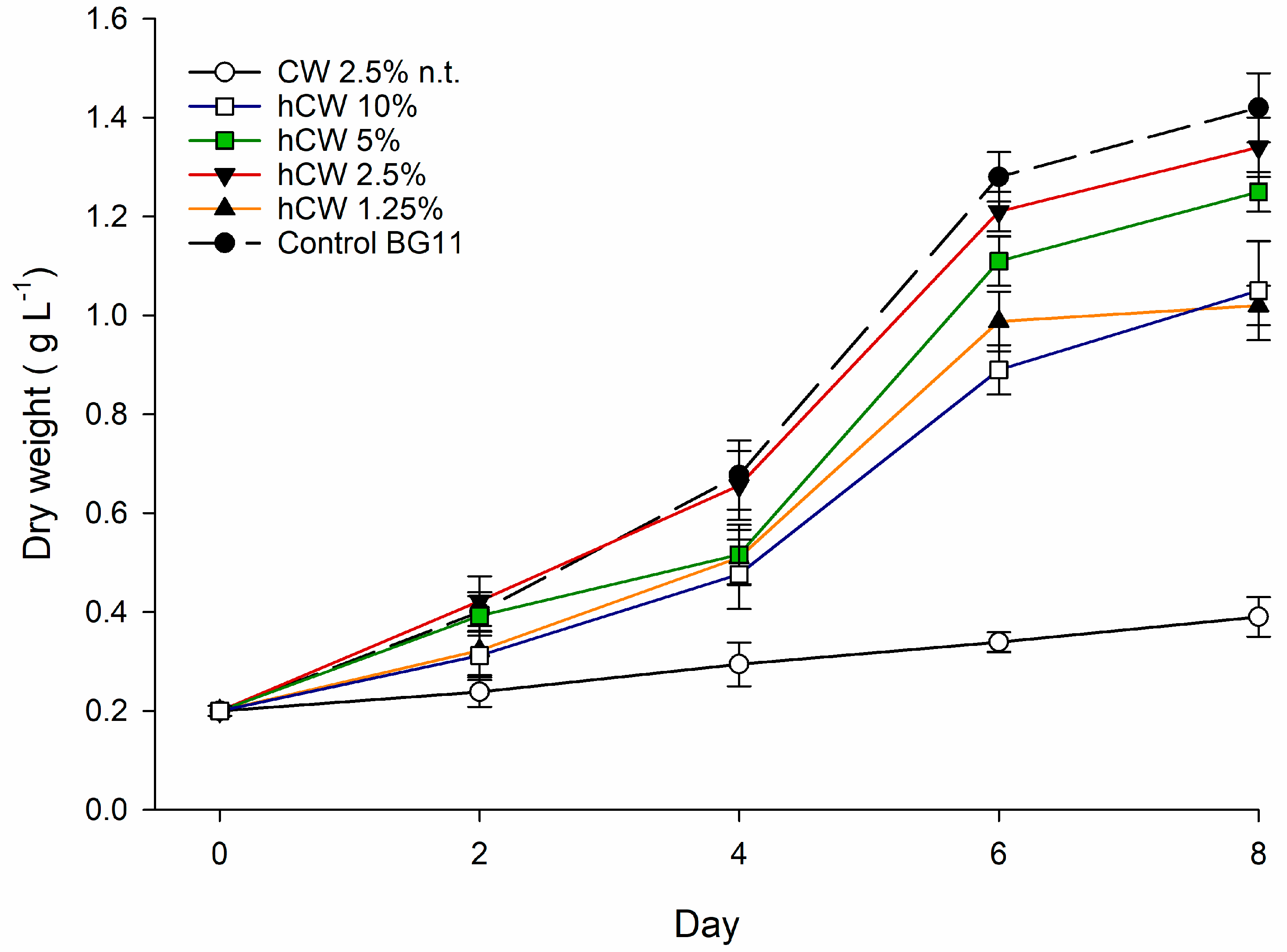
2.1. Characterization of FBW Used and Screening Results
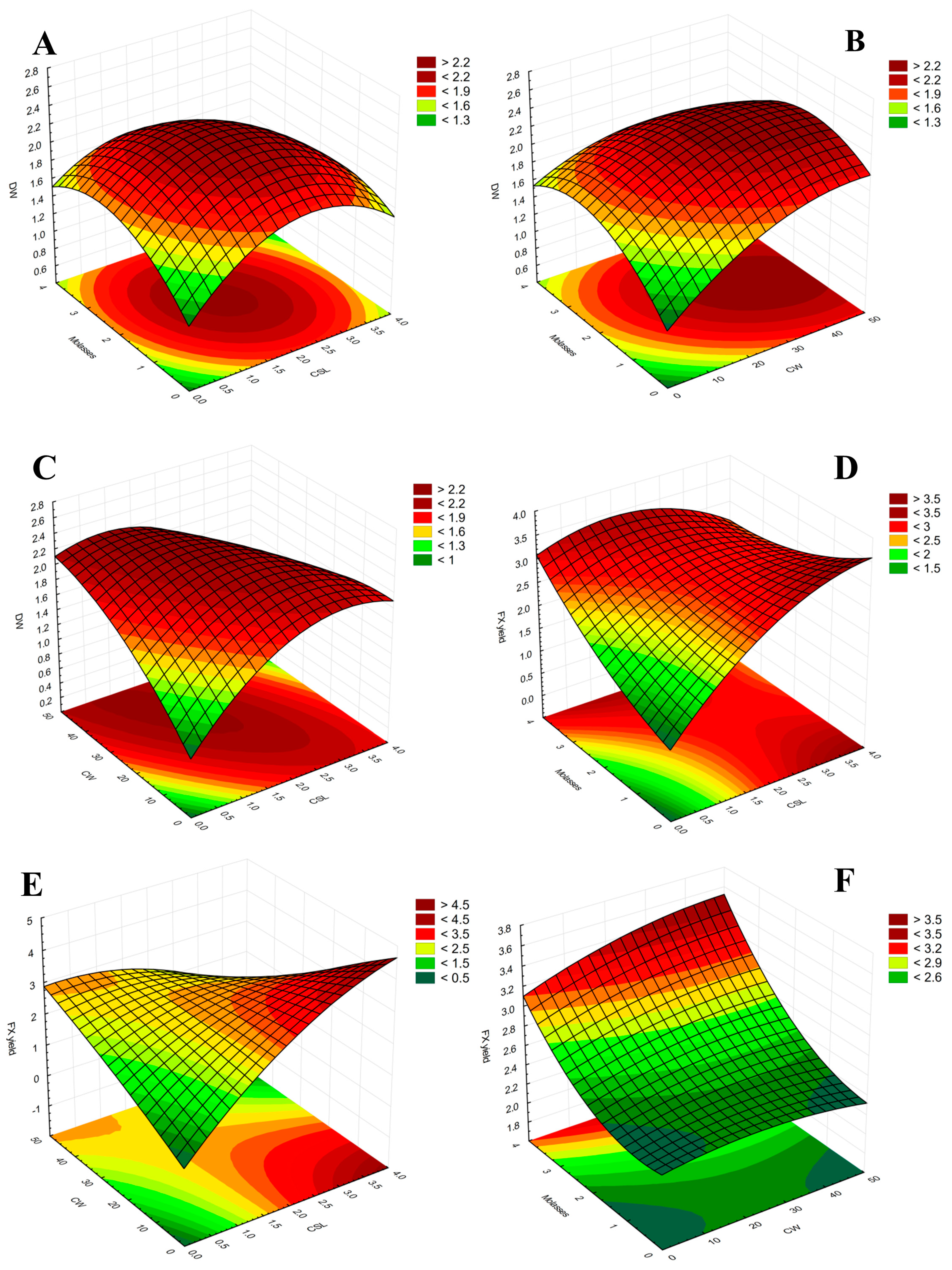
2.2. Results of Medium Optimization Using RSM–CCD
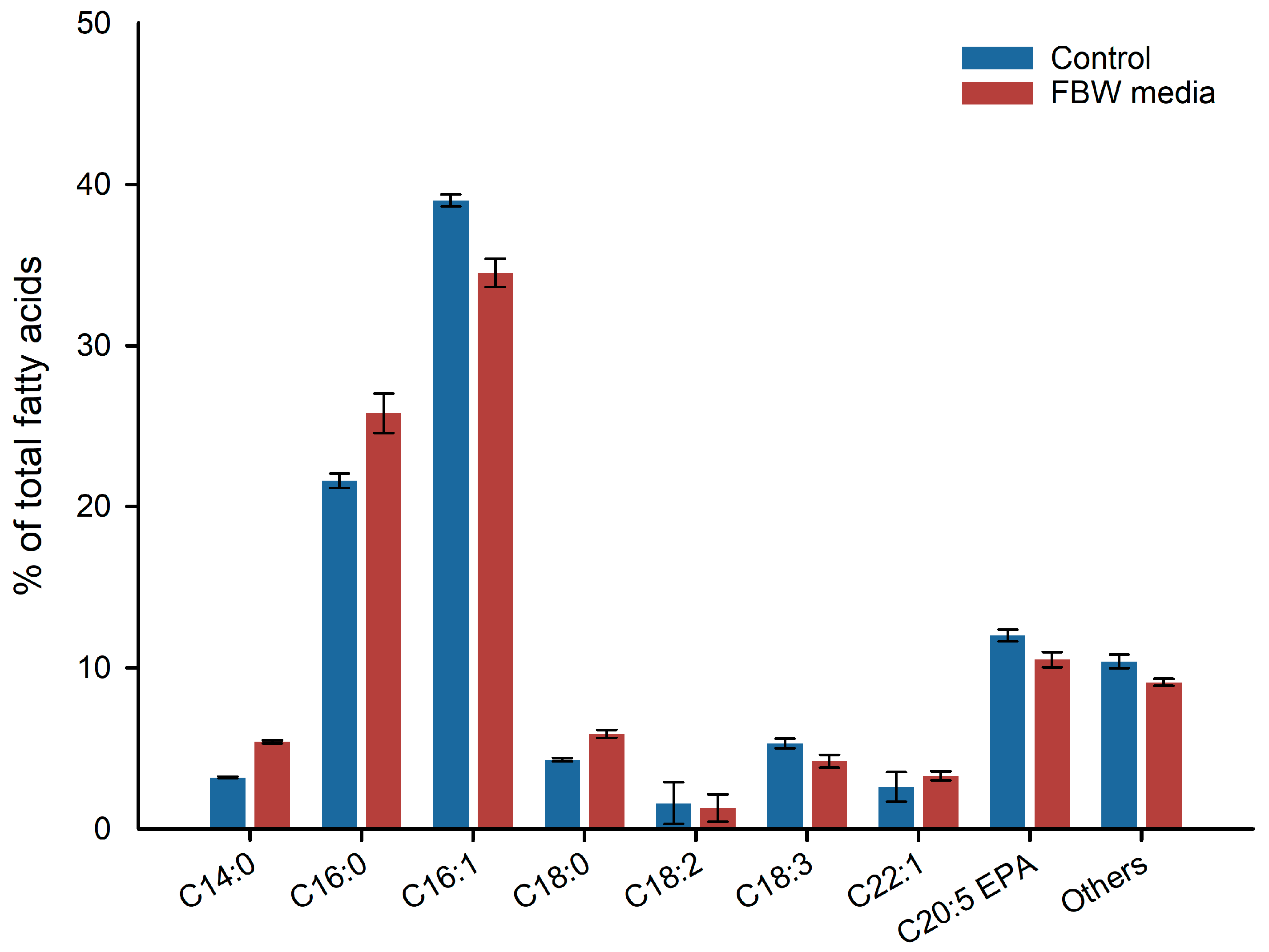
2.3. Fatty Acids Analysis and Carotenoid Profile
3. Discussion
4. Materials and Methods
4.1. Organism and Cultivation
4.2. Growth Parameters
4.3. Food By-Products and Cheese Whey Pre-Treatments
4.4. Chemical Characterization of FBW
4.5. Alternative Growth Media Screening
4.6. Response Surface Analysis and Formulation of Optimized Media
4.7. Lipid Extraction and Fatty Acid Methyl Esters (FAMEs)
4.8. Determination of Carotenoids in Microalgal Extracts by HPLC/MS Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andrade, M.R.; Costa, J.A. V Mixotrophic cultivation of microalga Spirulina platensis using molasses as organic substrate. Aquaculture 2007, 264, 130–134. [Google Scholar] [CrossRef]
- Russo, G.L.; Langellotti, A.L.; Oliviero, M.; Sacchi, R.; Masi, P. Sustainable production of food grade omega-3 oil using aquatic protists: Reliability and future horizons. New Biotechnol. 2021, 62, 32–39. [Google Scholar] [CrossRef]
- Pocha, C.K.R.; Chia, W.Y.; Chew, K.W.; Munawaroh, H.S.H.; Show, P.L. Current advances in recovery and biorefinery of fucoxanthin from Phaeodactylum tricornutum. Algal Res. 2022, 65, 102735. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Chen, L.; Cheng, W.; Liu, T. Combined production of fucoxanthin and EPA from two diatom strains Phaeodactylum tricornutum and Cylindrotheca fusiformis cultures. Bioprocess Biosyst. Eng. 2018, 41, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Leu, S.; Boussiba, S. Advances in the Production of High-Value Products by Microalgae. Ind. Biotechnol. 2014, 10, 169–183. [Google Scholar] [CrossRef]
- McClure, D.D.; Luiz, A.; Gerber, B.; Barton, G.W.; Kavanagh, J.M. An investigation into the effect of culture conditions on fucoxanthin production using the marine microalgae Phaeodactylum tricornutum. Algal Res. 2018, 29, 41–48. [Google Scholar] [CrossRef]
- Pereira, H.; Sá, M.; Maia, I.; Rodrigues, A.; Teles, I.; Wijffels, R.H.; Navalho, J.; Barbosa, M. Fucoxanthin production from Tisochrysis lutea and Phaeodactylum tricornutum at industrial scale. Algal Res. 2021, 56, 102322. [Google Scholar] [CrossRef]
- Leong, Y.K.; Chen, C.Y.; Varjani, S.; Chang, J.S. Producing fucoxanthin from algae—Recent advances in cultivation strategies and downstream processing. Bioresour. Technol. 2022, 344, 126170. [Google Scholar] [CrossRef]
- Liu, X.; Duan, S.; Li, A.; Xu, N.; Cai, Z.; Hu, Z. Effects of organic carbon sources on growth, photosynthesis, and respiration of Phaeodactylum tricornutum. J. Appl. Phycol. 2009, 21, 239–246. [Google Scholar] [CrossRef]
- Wang, X.; Balamurugan, S.; Liu, S.F.; Zhang, M.M.; Yang, W.D.; Liu, J.S.; Li, H.Y.; Lin, C.S.K. Enhanced polyunsaturated fatty acid production using food wastes and biofuels byproducts by an evolved strain of Phaeodactylum tricornutum. Bioresour. Technol. 2020, 296, 122351. [Google Scholar] [CrossRef]
- McDowell, D.; Dick, J.T.; Eagling, L.; Julius, M.; Sheldrake, G.N.; Theodoridou, K.; Walsh, P.J. Recycling nutrients from anaerobic digestates for the cultivation of Phaeodactylum tricornutum: A feasibility study. Algal Res. 2020, 48, 101893. [Google Scholar] [CrossRef]
- Massa, M.; Buono, S.; Langellotti, A.L.; Castaldo, L.; Martello, A.; Paduano, A.; Sacchi, R.; Fogliano, V. Evaluation of anaerobic digestates from different feedstocks as growth media for Tetradesmus obliquus, Botryococcus braunii, Phaeodactylum tricornutum and Arthrospira maxima. New Biotechnol. 2017, 36, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, S.F.; Wang, Z.Y.; Hao, T.B.; Balamurugan, S.; Li, D.W.; He, Y.; Li, H.Y.; Lin, C.S.K. A waste upcycling loop: Two-factor adaptive evolution of microalgae to increase polyunsaturated fatty acid production using food waste. J. Clean. Prod. 2022, 331, 130018. [Google Scholar] [CrossRef]
- Karpagam, R.; Jawaharraj, K.; Ashokkumar, B.; Pugazhendhi, A.; Varalakshmi, P. A cheap two-step cultivation of Phaeodactylum tricornutum for increased TAG production and differential expression of TAG biosynthesis associated genes. J. Biotechnol. 2022, 354, 53–62. [Google Scholar] [CrossRef]
- Wang, X.; Liu, S.F.; Qin, Z.H.; Balamurugan, S.; Li, H.Y.; Lin, C.S.K. Sustainable and stepwise waste-based utilisation strategy for the production of biomass and biofuels by engineered microalgae. Environ. Pollut. 2020, 265, 114854. [Google Scholar] [CrossRef]
- Smith, J.K.P.; Hughes, A.D.; Mcevoy, L.; Day, J.G. Use of crude glycerol for mixotrophic culture of Phaeodactylum tricornutum. Algal Res. 2023, 69, 102929. [Google Scholar] [CrossRef]
- Pleissner, D.; Lam, W.C.; Sun, Z.; Lin, C.S.K. Food waste as nutrient source in heterotrophic microalgae cultivation. Bioresour. Technol. 2013, 137, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.L.; Langellotti, A.L.; Verardo, V.; Martín-García, B.; Di Pierro, P.; Sorrentino, A.; Baselice, M.; Oliviero, M.; Sacchi, R.; Masi, P. Formulation of New Media from Dairy and Brewery Wastes for a Sustainable Production of DHA-Rich Oil by Aurantiochytrium mangrovei. Mar. Drugs 2021, 20, 39. [Google Scholar] [CrossRef]
- Humhal, T.; Kronusová, O.; Kaštánek, P.; Potočár, T.; Kohoutková, J.; Brányik, T. Influence of nitrogen sources on growth of thraustochytrids in waste water from the demineralization of cheese whey. Czech J. Food Sci. 2019, 37, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Abreu, A.P.; Fernandes, B.; Vicente, A.A.; Teixeira, J.; Dragone, G. Mixotrophic cultivation of Chlorella vulgaris using industrial dairy waste as organic carbon source. Bioresour. Technol. 2012, 118, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Russo, G.L.; Langellotti, A.L.; Sacchi, R.; Masi, P. Techno-economic assessment of DHA-rich Aurantiochytrium sp. production using food industry by-products and waste streams as alternative growth media. Bioresour. Technol. Rep. 2022, 18, 100997. [Google Scholar] [CrossRef]
- Sjölin, M.; Thuvander, J.; Wallberg, O.; Lipnizki, F. Purification of sucrose in sugar beet molasses by utilizing ceramic nanofiltration and ultrafiltration membranes. Membranes 2020, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.P.; Jahim, J.M.; Wu, T.Y.; Harun, S.; Mumtaz, T. Use of corn steep liquor as an economical nitrogen source for biosuccinic acid production by Actinobacillus succinogenes. IOP Conf. Ser. Earth Environ. Sci. 2016, 36, 012058. [Google Scholar] [CrossRef]
- Wan, C.; Bai, F.W.; Zhao, X.Q. Effects of nitrogen concentration and media replacement on cell growth and lipid production of oleaginous marine microalga Nannochloropsis oceanica DUT01. Biochem. Eng. J. 2013, 78, 32–38. [Google Scholar] [CrossRef]
- Yuan, X.; Liang, L.; Liu, K.; Xie, L.; Huang, L.; He, W.; Chen, Y.; Xue, T. Spent yeast as an efficient medium supplement for fucoxanthin and eicosapentaenoic acid (EPA) production by Phaeodactylum tricornutum. J. Appl. Phycol. 2020, 32, 59–69. [Google Scholar] [CrossRef]
- Wheeler, P.A.; North, B.B.; Stephens, G.C. Amino acid uptake by marine phytoplankters. Limnol. Oceanogr. 1974, 19, 249–259. [Google Scholar] [CrossRef]
- Yang, R.; Wei, D. Improving Fucoxanthin Production in Mixotrophic Culture of Marine Diatom Phaeodactylum tricornutum by LED Light Shift and Nitrogen Supplementation. Front. Bioeng. Biotechnol. 2020, 8, 820. [Google Scholar] [CrossRef]
- Nur, M.M.A.; Muizelaar, W.; Boelen, P.; Buma, A.G.J. Environmental and nutrient conditions influence fucoxanthin productivity of the marine diatom Phaeodactylum tricornutum grown on palm oil mill effluent. J. Appl. Phycol. 2019, 31, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.J.; Duan, S.S.; Li, A.F.; Sun, K.F. Effects of glycerol on the fluorescence spectra and chloroplast ultrastructure of Phaeodactylum tricornutum (Bacillariophyta). J. Integr. Plant Biol. 2009, 51, 272–278. [Google Scholar] [CrossRef]
- Nymark, M.; Valle, K.C.; Brembu, T.; Hancke, K.; Winge, P.; Andresen, K.; Johnsen, G.; Bones, A.M. An integrated analysis of molecular acclimation to high light in the marine diatom Phaeodactylum tricornutum. PLoS ONE 2009, 4, e7743. [Google Scholar] [CrossRef] [Green Version]
- Ishika, T.; Moheimani, N.R.; Bahri, P.A.; Laird, D.W.; Blair, S.; Parlevliet, D. Halo-adapted microalgae for fucoxanthin production: Effect of incremental increase in salinity. Algal Res. 2017, 28, 66–73. [Google Scholar] [CrossRef]
- Gómez-Loredo, A.; Benavides, J.; Rito-Palomares, M. Growth kinetics and fucoxanthin production of Phaeodactylum tricornutum and Isochrysis galbana cultures at different light and agitation conditions. J. Appl. Phycol. 2016, 28, 849–860. [Google Scholar] [CrossRef]
- Di Lena, G.; Casini, I.; Lucarini, M.; Lombardi-Boccia, G. Carotenoid profiling of five microalgae species from large-scale production. Food Res. Int. 2019, 120, 810–818. [Google Scholar] [CrossRef]
- Humaidah, N.; Nakai, S.; Nishijima, W.; Gotoh, T.; Furuta, M. Application of Aurantiochytrium sp. L3W for food-processing wastewater treatment in combination with polyunsaturated fatty acids production for fish aquaculture. Sci. Total Environ. 2020, 743, 140735. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.L.; Langellotti, A.L.; Mart, B.; Verardo, V.; Romano, R.; Sacchi, R.; Masi, P. New Biotechnological Production of EPA by Pythium irregulare Using Alternative Sustainable Media Obtained from Food Industry By-Products and Waste. Sustainability 2023, 15, 1147. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals; Springer: Berlin/Heidelberg, Germany, 1975. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Thiex, N. Evaluation of Analytical Methods for the Determination of Moisture, Crude Protein, Crude Fat, and Crude Fiber in Distillers Dried Grains with Solubles. J. AOAC Int. 2009, 92, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Lie, S. The EBC-Ninhydrin Method for Determination of Free Alpha Amino Nitrogen. J. Inst. Brew. 1973, 79, 37–41. [Google Scholar] [CrossRef]
- Cha, T.S.; Chen, J.W.; Goh, E.G.; Aziz, A.; Loh, S.H. Differential regulation of fatty acid biosynthesis in two Chlorella species in response to nitrate treatments and the potential of binary blending microalgae oils for biodiesel application. Bioresour. Technol. 2011, 102, 10633–10640. [Google Scholar] [CrossRef]
- Aued-Pimentel, S.; Lago, J.H.G.; Chaves, M.H.; Kumagai, E.E. Evaluation of a methylation procedure to determine cyclopropenoids fatty acids from Sterculia striata St. Hil. Et Nauds seed oil. J. Chromatogr. A 2004, 1054, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Conde, T.A.; Couto, D.; Melo, T.; Costa, M.; Silva, J.; Domingues, M.R.; Domingues, P. Polar lipidomic profile shows Chlorococcum amblystomatis as a promising source of value-added lipids. Sci. Rep. 2021, 11, 4355. [Google Scholar] [CrossRef] [PubMed]
- Castro-Puyana, M.; Herrero, M.; Urreta, I.; Mendiola, J.A.; Cifuentes, A.; Ibáñez, E.; Suárez-Alvarez, S. Optimization of clean extraction methods to isolate carotenoids from the microalga Neochloris oleoabundans and subsequent chemical characterization using liquid chromatography tandem mass spectrometry. Anal. Bioanal. Chem. 2013, 405, 4607–4616. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | CW | hCW | CSL | BM |
|---|---|---|---|---|
| pH | 5.3 ± 0.1 | 5.7 ± 0.1 | 3.6 ± 0.1 | 4.2 ± 0.2 |
| Ash (g kg−1) | 16.5 ± 0.4 | 14.2 ± 0.2 | 137.5 ± 2.2 | 81.9 ± 0.4 |
| Dry weight (%) | 21.9 ± 1.1 | 20.1 ± 0.4 | 62.59 ± 1.8 | 73.2 ± 0.8 |
| N total (g kg−1) | 6.7 ± 0.2 | 6.4 ± 0.2 | 48.45 ± 2.9 | 13.92 ± 0.08 |
| Protein content (g kg−1) | 28.8 ± 0.5 | 5.18 ± 0.3 | 305.83 ± 1.3 | 78.13 ± 1.1 |
| Lactic acid (g kg−1) | 1.5 ± 0.2 | 1.9 ± 0.1 | 121.2± 2.7 | 33.4 ± 1.2 |
| Free amino nitrogen (g kg−1) | 0.31 ± 0.06 | 3.19 ± 0.06 | 18.19 ± 0.6 | 0.11 ± 0.03 |
| Total sugars (g kg−1) | 201.2 ± 2.2 | 197.2 ± 3.1 | 35.1 ± 0.8 | 667.2 ± 4.5 |
| Lactose (g kg−1) | 196.8 ± 2.7 | 8.1 ± 1.1 | n.d. | n.d. |
| Total P (mg kg−1) | 145 ± 3.5 | 132 ± 1.3 | 18.2 ± 0.4 | 26.6 ± 1.1 |
| Run | Factor Assignment | Responses (Y) | |||
|---|---|---|---|---|---|
| X1 (BM) | X2 (CW) | X3 (CSL) | Biomass DW (g L−1) | Fucoxanthin Yield (mg L−1) | |
| 1 | 1.9 | 25 | 1.4 | 2.41 | 2.92 |
| 2 | 1.9 | 25 | 2.379 | 2.54 | 3.22 |
| 3 | 1.9 | 25 | 0.421 | 1.90 | 1.87 |
| 4 | 1.9 | 0.505 | 1.4 | 1.91 | 2.33 |
| 5 | 3.696 | 25 | 1.4 | 2.18 | 3.67 |
| 6 | 1.9 | 25 | 1.4 | 2.25 | 2.50 |
| 7 | 1.9 | 49.495 | 1.4 | 2.31 | 2.84 |
| 8 | 0.103 | 25 | 1.4 | 1.89 | 2.23 |
| 9 | 3 | 40 | 2 | 1.91 | 2.53 |
| 10 | 0.8 | 40 | 0.8 | 2.14 | 2.40 |
| 11 | 3 | 40 | 0.8 | 2.21 | 3.07 |
| 12 | 1.9 | 25 | 1.4 | 2.21 | 2.67 |
| 13 | 0.8 | 10 | 2 | 1.75 | 3.09 |
| 14 | 0.8 | 40 | 2 | 2.16 | 2.45 |
| 15 | 3 | 10 | 2 | 1.94 | 3.22 |
| 16 | 0.8 | 10 | 0.8 | 1.51 | 1.96 |
| 17 | 1.9 | 25 | 1.4 | 2.07 | 2.42 |
| 18 | 1.9 | 25 | 1.4 | 2.31 | 2.74 |
| 19 | 3 | 10 | 0.8 | 1.66 | 2.39 |
| 20 | 1.9 | 25 | 1.4 | 2.27 | 2.67 |
| Source | DF a | Adj SS b | Adj MS c | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 10 | 1.118 | 0.1118 | 5.45 | 0.009 |
| Blocks | 1 | 0.127 | 0.1272 | 6.21 | 0.034 |
| Linear | 3 | 0.519 | 0.1731 | 8.44 | 0.006 |
| Molasses | 1 | 0.029 | 0.0296 | 1.44 | 0.26 |
| CW | 1 | 0.366 | 0.3665 | 17.87 | 0.002 |
| CSL | 1 | 0.123 | 0.1232 | 6.01 | 0.037 |
| Square | 3 | 0.347 | 0.1159 | 5.65 | 0.019 |
| Molasses ×Molasses | 1 | 0.217 | 0.2171 | 10.59 | 0.01 |
| CW × CW | 1 | 0.130 | 0.1301 | 6.34 | 0.033 |
| CSL × CSL | 1 | 0.042 | 0.0424 | 2.07 | 0.184 |
| 2-Way Interaction | 3 | 0.123 | 0.0412 | 2.01 | 0.183 |
| BM × CW | 1 | 0.034 | 0.0340 | 1.66 | 0.23 |
| BM × CSL | 1 | 0.0099 | 0.0099 | 0.48 | 0.504 |
| CW × CSL | 1 | 0.0796 | 0.0796 | 3.88 | 0.08 |
| Error | 9 | 0.1845 | 0.020 | ||
| Lack-of-Fit | 5 | 0.13928 | 0.027856 | 2.46 | 0.202 |
| Pure Error | 4 | 0.04531 | 0.011328 | ||
| Total | 19 | 1.30267 |
| Source | DF a | Adj SS b | Adj MS c | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 10 | 2.829 | 0.283 | 3.900 | 0.021 |
| Blocks | 1 | 0.018 | 0.018 | 0.240 | 0.633 |
| Linear | 3 | 1.851 | 0.617 | 8.490 | 0.005 |
| Molasses | 1 | 0.858 | 0.858 | 11.810 | 0.007 |
| CW | 1 | 0.041 | 0.041 | 0.560 | 0.473 |
| CSL | 1 | 0.953 | 0.953 | 13.120 | 0.006 |
| Square | 3 | 0.151 | 0.050 | 0.690 | 0.579 |
| Molasses × Molasses | 1 | 0.084 | 0.084 | 1.160 | 0.309 |
| CW × CW | 1 | 0.019 | 0.019 | 0.260 | 0.626 |
| CSL × CSL | 1 | 0.037 | 0.037 | 0.510 | 0.492 |
| 2-Way Interaction | 3 | 0.810 | 0.270 | 3.710 | 0.055 |
| BM × CW | 1 | 0.011 | 0.011 | 0.150 | 0.704 |
| BM × CSL | 1 | 0.127 | 0.127 | 1.750 | 0.218 |
| CW × CSL | 1 | 0.671 | 0.671 | 9.240 | 0.014 |
| Error | 9 | 0.654 | 0.073 | ||
| Lack-of-Fit | 5 | 0.507 | 0.101 | 2.770 | 0.212 |
| Pure Error | 4 | 0.146 | 0.037 | ||
| Total | 19 | 3.483 |
| Medium | Fucoxanthin | Violaxanthin | Antheraxanthin | Lutein | β-carotene |
|---|---|---|---|---|---|
| BG11 | 665.1 ± 7.8 b | n.d. | 0.64 ± 0.1 | 0.16 ± 0.01 | 7.42 ± 0.3 a |
| Guillard F/2 | 394 ± 1.5 a | n.d. | 0.95 ± 0.05 | 0.13 ± 0.02 | 3.89 ± 0.5 a |
| BM medium | 1221.1 ± 8.2 c | n.d. | 0.66 ± 0.03 | n.d. | 15.85 ± 0.8 b |
| CW medium | 410.3 ± 2.4 a | n.d. | 0.48 ± 0.02 | 0.35 ± 0.03 | 36.8 ± 1.1 c |
| CSL medium | 808.2 ± 4.1 d | 0.70 ± 0.05 | 0.34 ± 0.01 | n.d. | 34.23 ± 1.2 c |
| Optimized medium | 1785.7 ± 5.1 e | 0.41 ± 0.01 | 0.68 ± 0.03 | 0.25 ± 0.03 | 47.6 ± 0.2 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russo, G.L.; Langellotti, A.L.; Verardo, V.; Martín-García, B.; Oliviero, M.; Baselice, M.; Di Pierro, P.; Sorrentino, A.; Viscardi, S.; Marileo, L.; et al. Bioconversion of Cheese Whey and Food By-Products by Phaeodactylum tricornutum into Fucoxanthin and n-3 Lc-PUFA through a Biorefinery Approach. Mar. Drugs 2023, 21, 190. https://doi.org/10.3390/md21030190
Russo GL, Langellotti AL, Verardo V, Martín-García B, Oliviero M, Baselice M, Di Pierro P, Sorrentino A, Viscardi S, Marileo L, et al. Bioconversion of Cheese Whey and Food By-Products by Phaeodactylum tricornutum into Fucoxanthin and n-3 Lc-PUFA through a Biorefinery Approach. Marine Drugs. 2023; 21(3):190. https://doi.org/10.3390/md21030190
Chicago/Turabian StyleRusso, Giovanni Luca, Antonio Luca Langellotti, Vito Verardo, Beatriz Martín-García, Maria Oliviero, Marco Baselice, Prospero Di Pierro, Angela Sorrentino, Sharon Viscardi, Luis Marileo, and et al. 2023. "Bioconversion of Cheese Whey and Food By-Products by Phaeodactylum tricornutum into Fucoxanthin and n-3 Lc-PUFA through a Biorefinery Approach" Marine Drugs 21, no. 3: 190. https://doi.org/10.3390/md21030190