
Marine Organisms as a Prolific Source of Bioactive Depsipeptides

1
School of Pharmaceutical Sciences, Zhejiang University of Technology, Hangzhou 310014, China
2
College of Life and Environmental Sciences, Hangzhou Normal University, Hangzhou 311121, China
*
Author to whom correspondence should be addressed.
Mar. Drugs 2023, 21(2), 120; https://doi.org/10.3390/md21020120
Submission received: 20 January 2023
/
Revised: 31 January 2023
/
Accepted: 9 February 2023
/
Published: 11 February 2023
(This article belongs to the Special Issue Marine Bioactive Peptides: Structure, Function, and Therapeutic Potential IV)
Abstract
:Depsipeptides, an important group of polypeptides containing residues of hydroxy acids and amino acids linked together by amide and ester bonds, have potential applications in agriculture and medicine. A growing body of evidence demonstrates that marine organisms are prolific sources of depsipeptides, such as marine cyanobacteria, sponges, mollusks, microorganisms and algae. However, these substances have not yet been comprehensively summarized. In order to enrich our knowledge about marine depsipeptides, their biological sources and structural features, as well as bioactivities, are highlighted in this review after an extensive literature search and data analysis.
Keywords:
depsipeptide; marine cyanobacterium; sponge; mollusk; marine microbe; alga; biological activity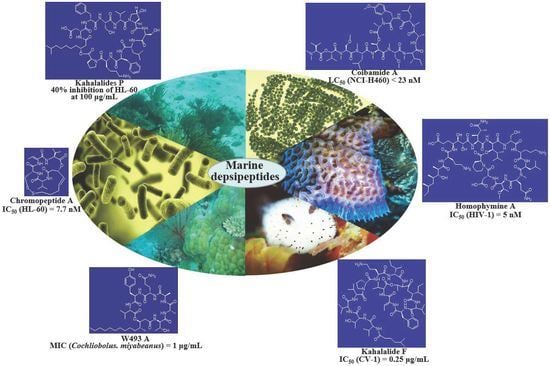
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
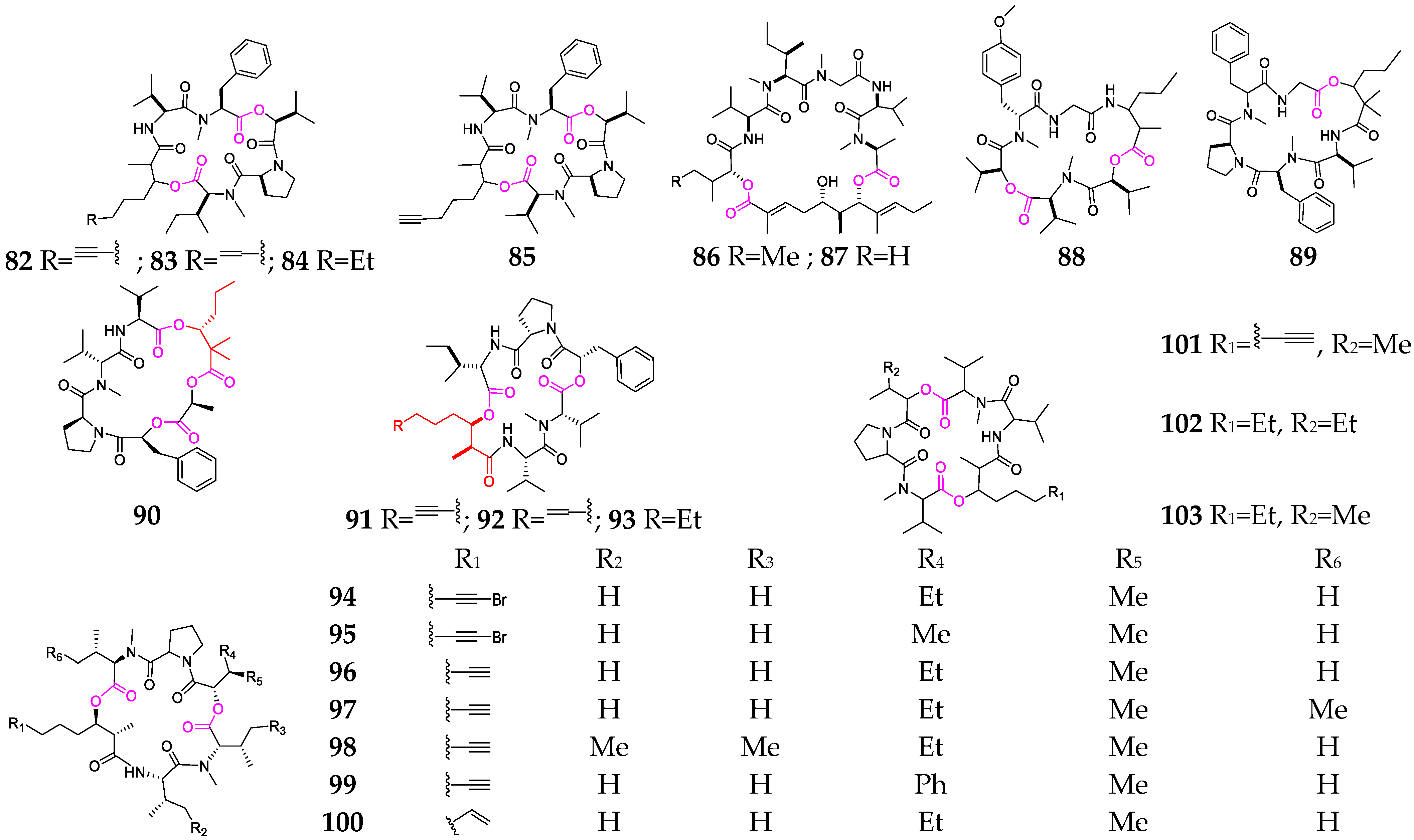{kind=link}
{kind=link}
{kind=link}
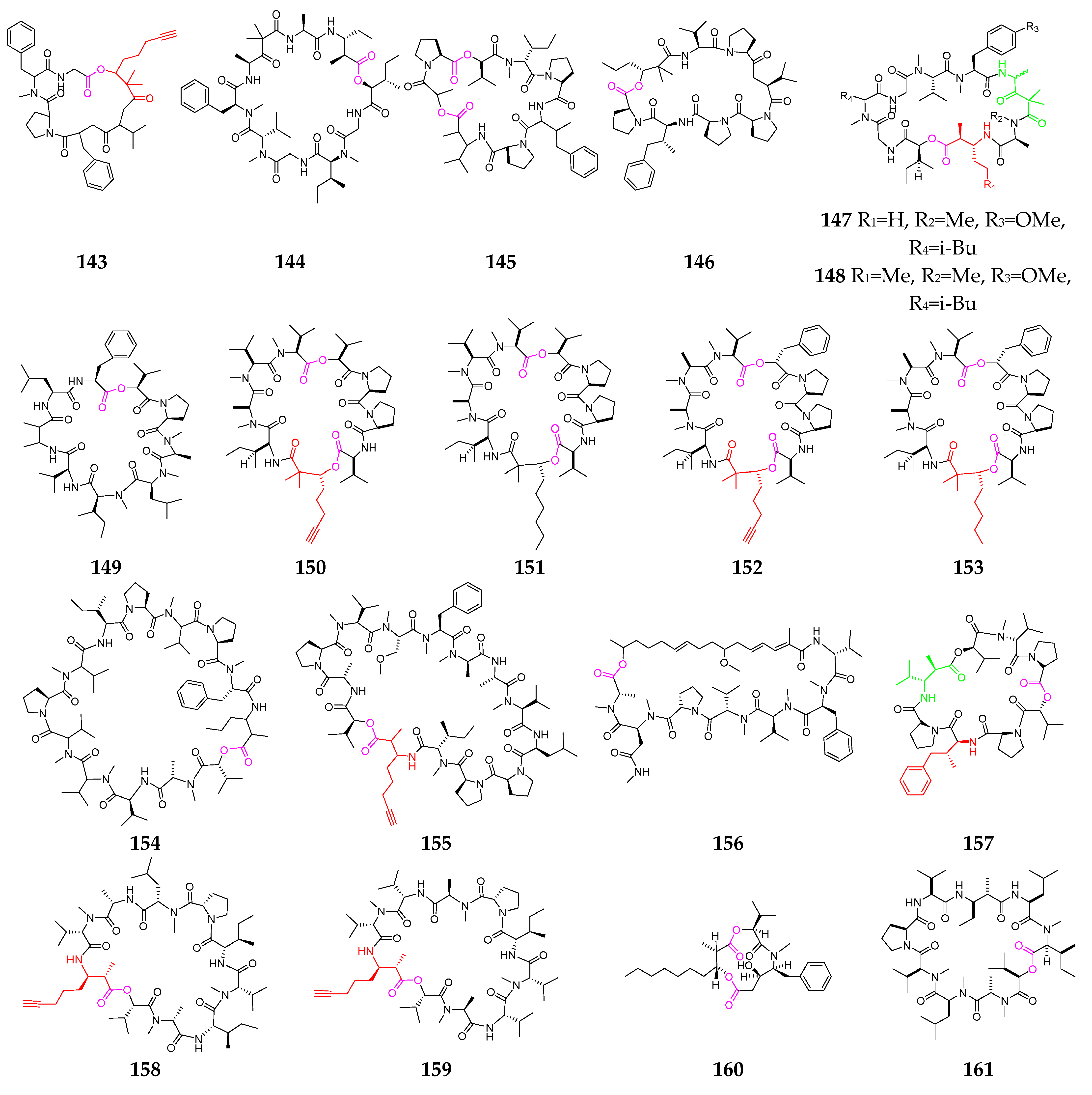{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
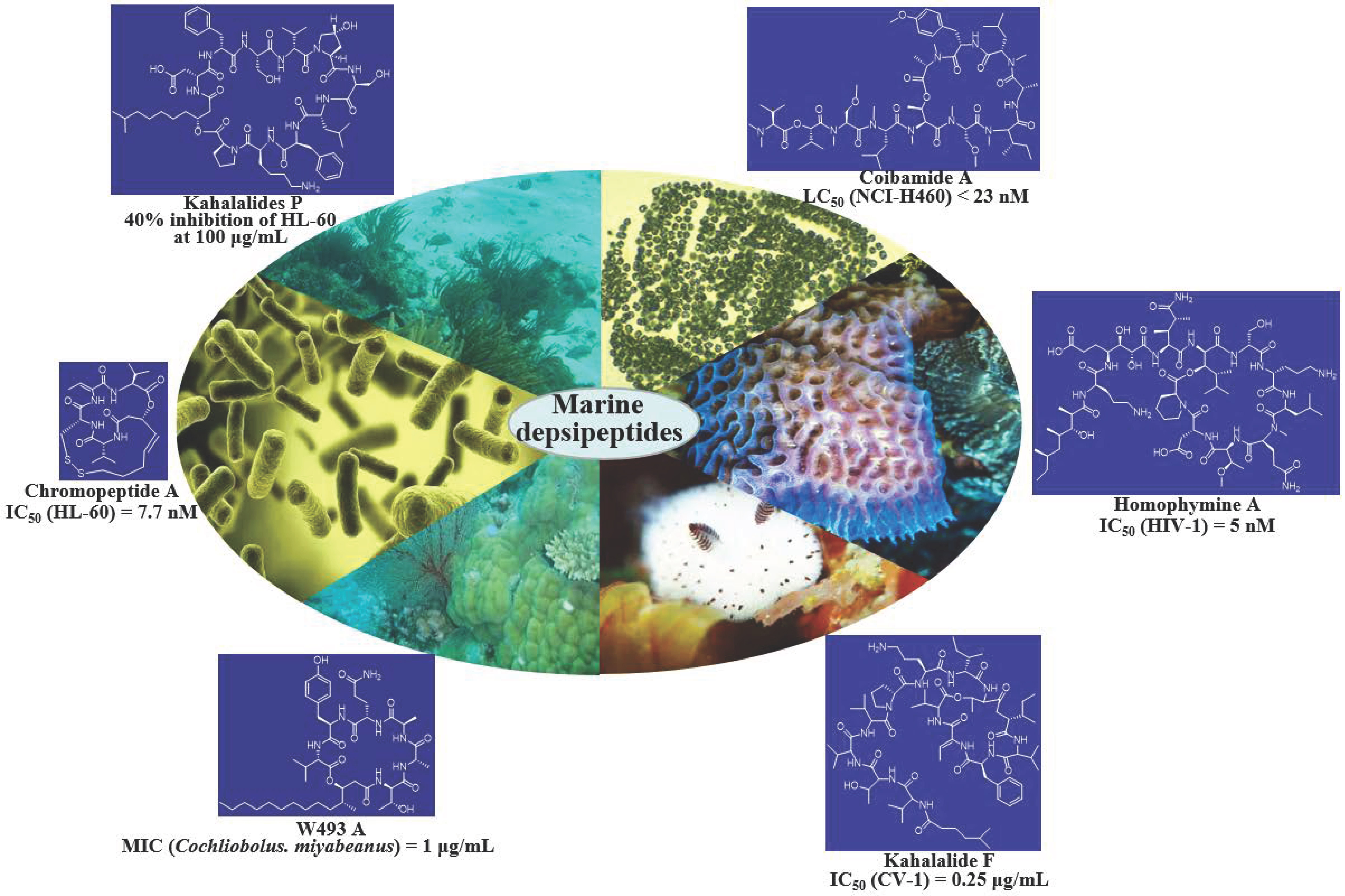
Marine organisms are tremendously important sources of natural products since almost 40,000 compounds have been discovered and recorded in the MarinLit database (https://marinlit.rsc.org/, accessed on 12 December 2022) [1]. Depsipeptides are an important group of polypeptides simultaneously containing ester and amide bonds, and they display a wide variety of biological properties [2,3]. A number of naturally occurring depsipeptides have been successfully developed as new drugs or are being evaluated in clinical trials, such as the antitumor agents romidepsin [4,5], plitidepsin (aplidine) [6,7], kahalalide F [8,9] and OBP-801 (spiruchostatin A) [10]. Generally, these substances are divided into two groups, namely, cyclic and non-cyclic, of which the former tends to display excellent bioactivity [11,12]. However, marine-derived depsipeptides have not yet been comprehensively summarized until now. In order to enrich our knowledge about these compounds, their origins and structural features, as well as their biological properties, are highlighted in this review.
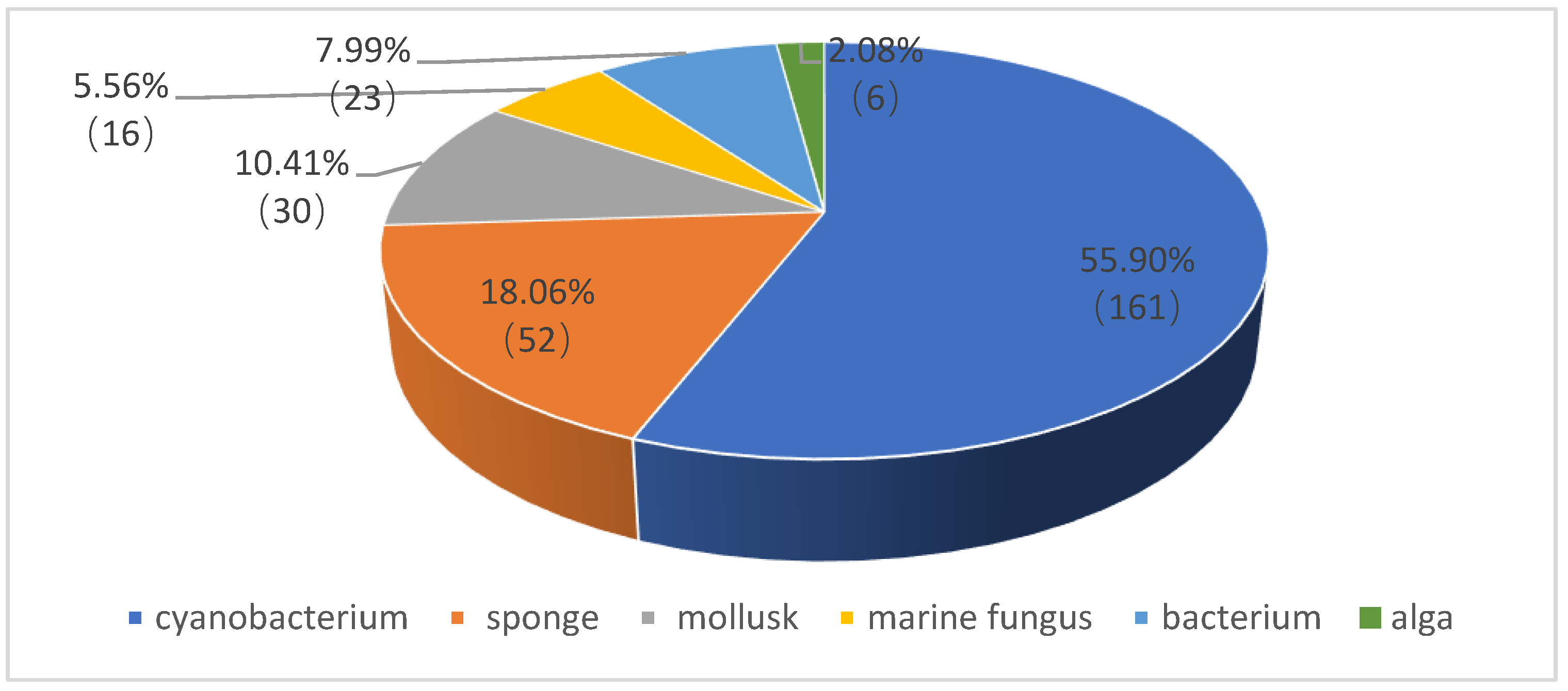
According to an extensive literature search using the DNP (Dictionary of Natural Products) database as well as Web of Science and SciFinder tools, as many as 288 depsipeptides (1–288) have been isolated and characterized from marine organisms. As shown in Figure 1, the major producers of depsipeptides are marine cyanobacteria, which make up 55.90%, followed by marine sponges (18.06%), mollusks (10.41%), bacteria (7.99%), marine fungi (5.56%) and algae (2.08%). On the basis of biological sources and chemical structures, these marine depsipeptides are each introduced herein. Their detailed information is supplied in the Supplementary Materials.
2. Marine Cyanobacteria
Marine-cyanobacterium-derived depsipeptides (1–161) have diverse chemical structures and a wide variety of pharmacological activities, and most of them are cytotoxic [13]. Structurally, these metabolites are linear and cyclic depsipeptides containing ɑ-amino or ɑ-hydroxy carboxylic acid residues, and the latter are the major components and can be further divided into five subgroups, including cyclic penta-, hexa-, and hepta-depsipeptides, thiazole-containing depsipeptides and others.
2.1. Linear Depsipeptides
The marine cyanobacteria Symploca and Lyngbya spp. are the major producers of linear depsipeptides (1–27, Figure 2) [10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25]. Grassystatins D–F (1–3) containing statine units have strong aspartic protease inhibitory activity preferentially targeting cathepsins D and E [14]. Both compounds 4 and 5 possess an acetate extended and S-adenosyl methionine-modified isoleucine unit, a central triheterocyclic system comprising two R-methylated thiazolines and one thiazole, and a highly oxygenated and methylated C-15 polyketide unit [15], and 5 exerted potent inhibitory activity against the p53/MDM2 interaction (EC50 = 4.5 μM), an attractive target for anticancer drug development [15,16]. Malevamide D (7) exhibited a highly cytotoxic effect on P-388, A-549, HT-29 and MEL-28 cell lines in the subnanomolar range [17]. Symplostatins 3 (8) and 4 (9) were discovered as new analogs of dolastatin 10 and were shown to possess excellent cytotoxicity mediated by the disruption of microtubules [18]. Tasiamides A–E (10–14), produced by a Symploca sp., were found to have strong cytotoxicity against KB and LoVo cells [19,20,21].
The key structural feature of tasiamide F (15) is the presence of a Phe-derived statine core, which contributes to its aspartic protease inhibitory activity [22]. Izenamides A, B and C (16–18) were purified from an Okinawan Lyngbya sp. and demonstrated an inhibitory effect on cathepsin D [23]. Grassystatins A–C (19–21) showed potency and selectivity against cathepsins D and E in vivo [24]. Maedamide (22) was reported as a novel chymotrypsin-inhibiting depsipeptide and strongly inhibited the growth of HeLa and HL60 cell lines [25]. Lyngbyabellins D (23) and P (24) are, respectively, produced by Lyngbya sp. and Okeania sp. and displayed strong antifouling and cytotoxic activities [26,27]. Gallinamide A (25) was presented as a new antimalarial pentapeptide from a Schizothrix sp. collected off the north coast of Panama [28]. Veraguamides K (26) and L (27) are two unique cytotoxic depsipeptides containing brominated alkynyls and were isolated from Oscillatoria margaritifera [29].
2.2. Cyclic Depsipeptides
2.2.1. Cyclopentadepsipeptides
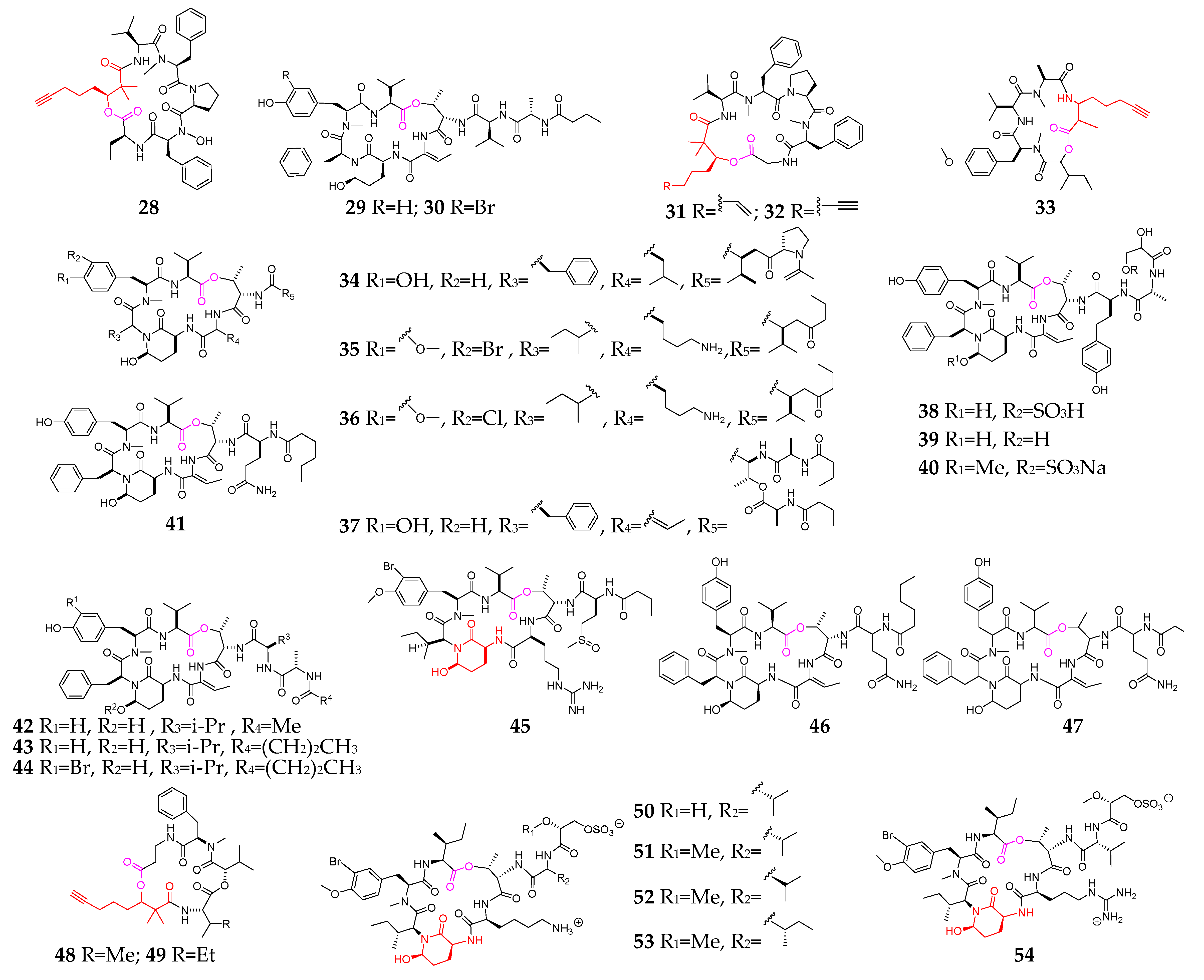
The marine cyanobacteria Lyngbya, Symploca and Dichothrix are the major sources of cyclopentadepsipeptides (28–80, Figure 3), of which compounds 28, 32, 33, 48 and 49 contain a unique 2,2-dimethyl-3-hydroxy-7-octynoic acid (Dhoya) residue. Bouillomides A (29) and B (30) were reported as two new analogs of dolastatin 13 and showed selective inhibitory effects on the serine proteases elastase and chymotrypsin [30]. Cocosamides A (31) and B (32) exhibited weak cytotoxicities toward MCF7 and HT-29 cancer cells [31]. Guineamide C (33) possessed moderate cytotoxicity against a mouse neuroblastoma cell line [32]. Floridian Lyngbya sp.-derived novel cyclodepsipeptides (34–36) exhibited potent inhibitory effects on elastase, chymotrypsin and trypsin [33,34]. Kurahamide (37) was presented as a new dolastatin 13 analog and strongly inhibited elastase and chymotrypsin as well as showed moderately cytotoxic activity against HeLa and HL60 cells [35]. Lyngbyastatins 4–6 (38–40) contain an unusual amino acid homotyrosine residue and selectively inhibited porcine pancreatic elastase and chymotrypsin [36,37]. Lyngbyastatins 8–10 (42–44) had strong inhibitory effects on porcine pancreatic elastase with IC50 values of 123 nM, 210 nM and 120 nM, respectively [38].
Peptolide (45) and kyanamide (65) possess a 3-amino-6-hydroxy-2-piperidone (Ahp) residue, and the former could selectively inhibit trypsin over elastase and chymotrypsin with an IC50 value of 2.4 mM [39,40]. Jizanpeptins A−E (50–54), possessing an Ahp residue in a typical micropeptin motif, showed the specific inhibition of the serine protease trypsin in vitro and were cytotoxic to HeLa cervical and NCI-H460 lung cancer cell lines [41]. Structure–activity relationship (SAR) studies and X-ray cocrystal structure analysis suggested that compound 55 has similar activity, comparable to the clinically approved elastase inhibitor sivelestat in short-term assays and superior sustained activity in longer-term assays [42,43]. Trikoveramides A–C (62–64) are members of the kulolide superfamily [44]. Largamides A–G (66–72) are characterized by the unusual occurrence of a senecioic acid unit, and compounds 69-72 exhibited a strong inhibitory effect on chymotrypsin [45]. Loggerpeptins A–C (73–75) were reported as new Ahp-containing cyclic depsipeptides and displayed an inhibitory effect on the cleavage of the elastase substrate CD40 [46]. Molassamide (76) was the first depsipeptide reported from the marine cyanobacterial genus Dichothrix and has protease inhibitory activity [47]. Odoamide (77) showed potent cytotoxicity against HeLa S3 human cervical cancer cells with an IC50 value of 26.3 nM [48]. Tutuilamides A–C (78–80) are characterized by the presence of several unusual residues, including Ahp, 2-amino-2-butenoic acid and vinyl chloride [49].
2.2.2. Cyclohexadepsipeptide
The marine cyanobacterium Moorena producens is the most important producer of cyclic hexadepsipeptides (82–103, Figure 4) [33,50,51,52,53,54,55]. The cytotoxicity-guided fractionation of a strain of M. producens collected from Papua New Guinea led to the isolation of aurilides B (86) and C (87), which showed excellent in vitro cytotoxicity toward NCI-H460 human lung tumor and the neuro-2a mouse neuroblastoma cell lines [51]. Guineamides D (88) and F (89) contain α-amino or α-hydroxy carboxylic acid residues and were isolated from a Papua New Guinea collection of M. producens [32]. Palmyramide A (90) features an unusual arrangement of three amino acids and three hydroxy acids; one of the hydroxy acids is the rare 2,2-dimethyl-3-hydroxyhexanoic acid (Dmhha) unit. This compound showed sodium-channel-blocking activity in neuro-2a cells and cytotoxic activity in H-460 human lung carcinoma cells [50]. Trungapeptins A–C (91–93) containing a 3-hydroxy-2-methyl-7-octynoic acid (Hmoya) residue were isolated and characterized from M. producens collected from Trung (Thailand) [52]. Veraguamides A-G (94–100) are characterized by the presence of an invariant proline residue, multiple N-methylated amino acids, an α-hydroxy acid and a C8-polyketide-derived β-hydroxy acid moiety with a characteristic terminus that is either an alkynyl bromide, alkyne or vinyl group. These metabolites showed moderate to weak cytotoxic activity against HT29 colorectal adenocarcinoma and HeLa cervical carcinoma cell lines [53].
2.2.3. Cycloheptadepsipeptide
As of the end of 2022, as many as 20 cyclic heptadepsipeptides (104–123, Figure 5) had been isolated and characterized from marine cyanobacteria, including Lyngbya, Leptolyngbya, Okeania, Dichothrix, Symploca and Rivularia [32]. Compounds 104–110 were derived from several Lyngbya spp., and compounds 105–107 displayed significant antimalarial properties and potent cytotoxic activities against P388 murine leukemia cell lines. Kohamamides A–C (112–114), containing a Leu residue adjacent to a Pro residue, belong to the kulolide superfamily [54]. Lagunamide D (115) exhibited low-nanomolar antiproliferative activity against A549 human lung adenocarcinoma cells, while its structural transformation from a 26-membered macrocycle to a 24-membered ring structure led to a 9.6-fold decrease in activity [55]. Pemukainalides A (116) had cytotoxicity against the MOLT-4 leukemia cell line with an IC50 value of 5.6 μM [56]. Viequeamides A-F (118–123) are a family of 2,2-dimethyl-3-hydroxy-7-octanic acid-containing cyclic depsipeptides, and compound 118 was found to be highly toxic to H460 human lung cancer cells (IC50 = 60 ± 10 nM) [57].
2.2.4. Thiazole-Containing Cyclodepsipeptides
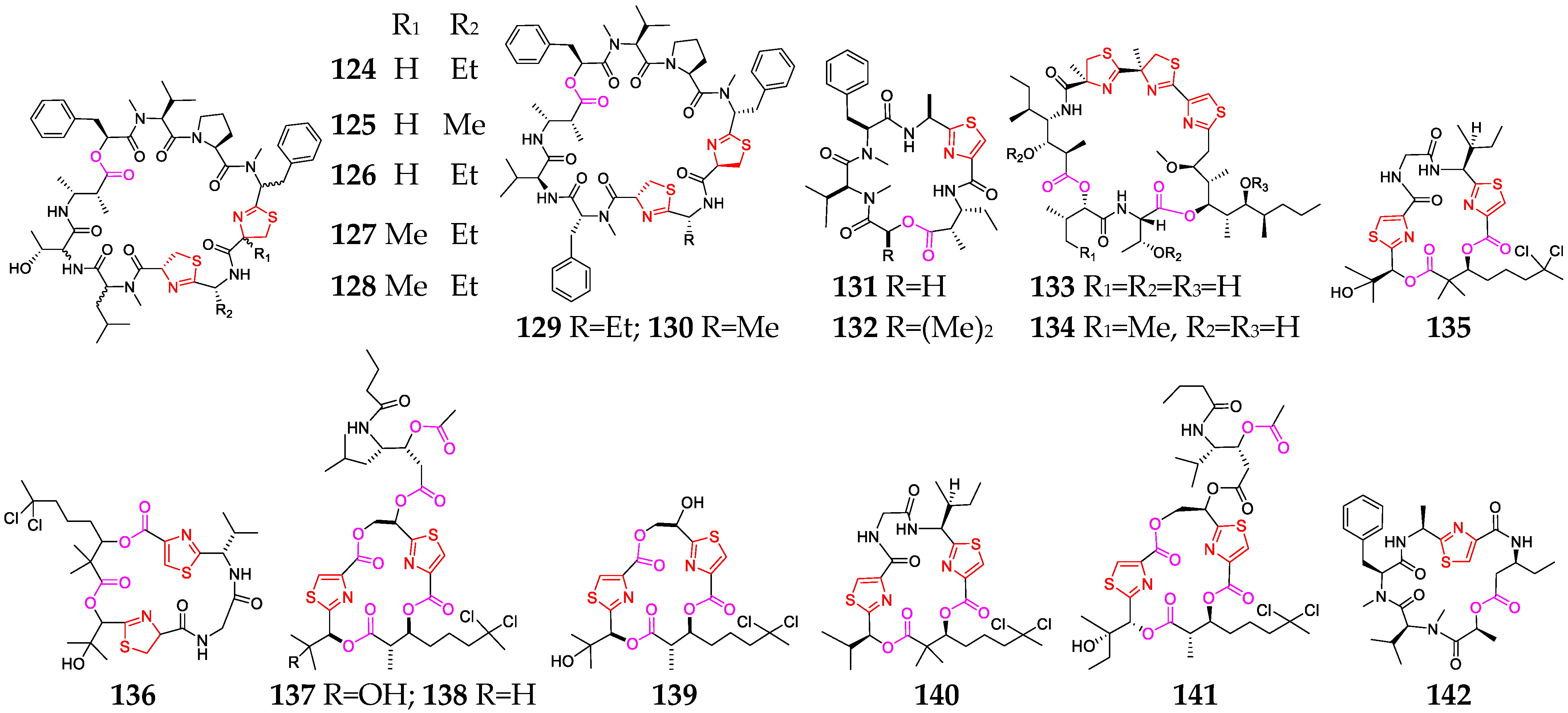
Nineteen thiazole-containing cyclic depsipeptides (124–142, Figure 6) were discovered in three marine cyanobacteria: Lyngbya, Leptolyngbya and Phormidium [11,28,55,56,57,58,59,60]. Grassypeptolides A-G (124–130) are a group of closely related bis-thiazoline-containing cyclic depsipeptides. SAR analyses indicated that the ethyl substituent in 124 is changed to a methyl substituent in 125, and its cytoactivity was only slightly reduced (3~4-fold), whereas the inversion of the Phe unit flanking the bis-thiazoline moiety resulted in 16~23-fold greater potency [58]. Compounds 127 and 128 showed significant cytotoxicity against HeLa and mouse neuro-2a blastoma cells [59], while 129 and 130 had moderate inhibitory activity against the transcription factor AP-1 (IC50 = 5.2 and 6.0 μM, respectively) [60]. Guineamides B (132) possessed moderate cytotoxicity against a mouse neuroblastoma cell line with an IC50 value of 15 µM [32]. Hoiamides A (133) and B (134) belong to the unique hoiamide structural class [15,61]. Compound 133 showed a potent inhibitory effect on [3H] batrachotoxin binding to voltage-gated sodium channels (IC50 = 92.8 nM) and activated sodium influx (EC50 = 2.31 μM) in mouse neocortical neurons, while 134 could stimulate sodium influx and suppressed spontaneous Ca2+ oscillations with EC50 values of 3.9 µM and 79.8 nM, respectively. Lyngbyabellin A (135) was shown to be a potent disrupter of the cellular microfilament network, and lyngbyabellin B (136) displayed potent toxicity toward brine shrimp and Candida albicans [62], while compounds 137 and 138 exhibited good cytotoxicity against NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cell lines, with LC50 values between 0.2 and 4.8 mM. Obyanamide (142), derived from a variety of L. confervoides, was cytotoxic against KB cells, with an IC50 value of 0.58 µg/mL [63].
2.2.5. Other Cyclodepsipeptides
Almost twenty other cyclodepsipeptides (143–161, Figure 7) have been obtained from Lyngbya [61,62,63,64,65,66], Symploca [67,68,69] and Okeania [70]. Guineamide G (143) showed potent brine shrimp toxicity and significant cytotoxicity against a mouse neuroblastoma cell line, with an LC50 value of 2.7 µM [64]. Desmethoxymajusculamide C (145) demonstrated potent and selective anti-solid-tumor activity against the HCT-116 human colon carcinoma cell line, with an IC50 value of 20 nM via the disruption of cellular microfilament networks [65]. Homodolastatin 16 (146), containing a 4-phenylvaline (dolaphenvaline, Dpv) moiety and a rare 2,2-dimethyl-3-hydroxyhexanoic acid (Dmhha) unit, shares higher homology with the potential anticancer agent dolastatin 16 [66]. Lyngbyastatin 3 (148) possesses two unusual amino acid residues, 3-amino-2-methylhexanoic acid (Amha) and 4-amino-2,2-dimethyl-3-oxopentanoic acid units (Ibu), and is a potent disrupter of cellular microfilament networks [67,68]. The four cytotoxic depsipeptides wewakpeptins A-D (150–153) represent an unusual arrangement of amino and hydroxy acid subunits and possess a bis-ester, a Dhoya or 2,2-dimethyl-3-hydroxyoctanoic acid (Dhoaa) residue, and a diprolyl group reminiscent of dolastatin 15 [69]. Malevamide E (156) had a potent inhibitory effect on Ca2+ release-activated Ca2+ (CRAC) channels [70]. Triproamide (157) contains the rare 4-phenylvaline (dolaphenvaline, Dpv) and a β-amino acid, dolamethylleucine (Dml), originally discovered in dolastatin 16 [56]. Companeramides A (158) and B (159) showed high nanomolar in vitro antiplasmodial activity [71], and hapalosin (160) displayed multidrug-resistance-reversing activity [72]. Urumamide (161) is a novel chymotrypsin inhibitor with a b-amino acid from a marine cyanobacterium Okeania sp. [73].
3. Marine Sponges
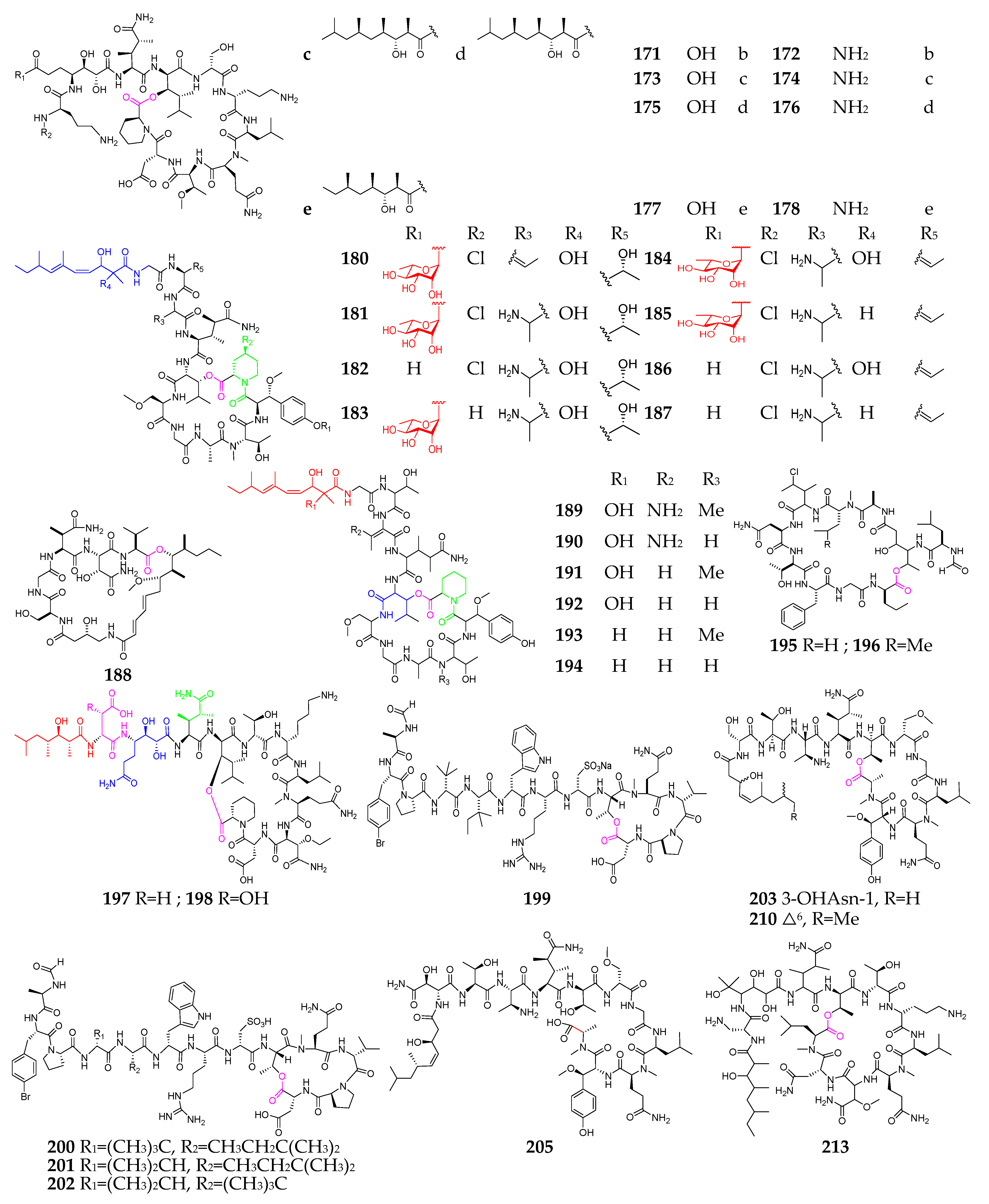
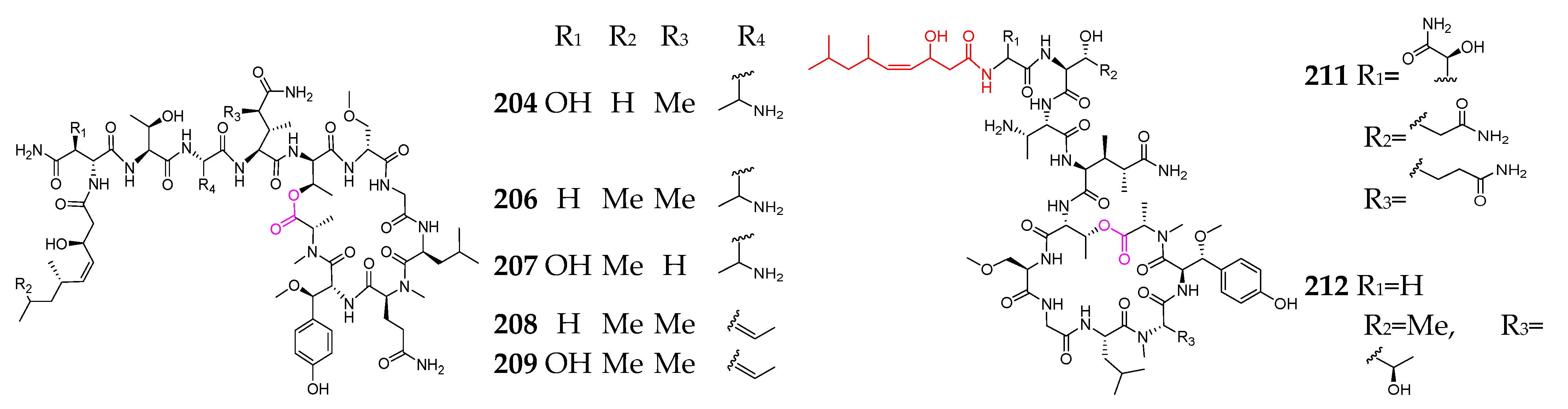
Marine sponges are well known as prolific sources of biologically natural products and are the second largest group of producers of marine-derived depsipeptides (162–212, Figure 8) [71,72,73,74,75,76,77,78,79,80,81,82,83,84]. With respect to the genus and species, however, there are no striking features about these sponges. Callipeltin A (162), obtained from a shallow water sponge of the genus Callipelta, exhibited a protective effect on cells infected with human immunodeficiency (HIV) virus [74]. Callipeltins N (164) and O (165) showed significant cytotoxicity against A2058, HT-29, MCF-7 and MRC-5 cell lines, with an IC50 value of 0.16 µM [75]. Cyclolithistide A (166) was discovered as a novel antifungal cyclodepsipeptide containing the unique amino acids 4-amino-3,5-dihydroxyhexanoic acid, formyl-leucine and chloroisoleucine [76]. Daedophamide (167) displayed strong cytotoxic activity against a panel of four human tumor cell lines with GI50 values in the submicromolar range [77]. Gunungamide A (168), produced by an Indonesian sponge Discodermia sp., possesses an unusual chloropyrrol ring [78]. Homophymine analogs (169–178) featuring new polyketide-derived end groups displayed potent antiproliferative activity (IC50 in the nM range) against a panel of human cancer cell lines [79,80]. Microspinosamide (179) was the first naturally occurring cyclodepsipeptide containing a ɑ-hydroxy-p-bromophenylalanine residue and inhibited the cytopathic effect of HIV-1 infection in an XTT-based in vitro assay, with an EC50 value of approximately 0.2 µg/mL [81]. Mirabamides A–H (180–187) have two new residues, 4-chlorohomoproline and β-methoxytyrosine 4′-O-α-L-rhamnopyranoside, along with a rare N-terminal aliphatic hydroxy acid, and were shown to potently inhibit HIV-1 fusion [82]. Papuamides A–F (189–194) were the first marine-derived depsipeptides reported to contain 3-hydroxyleucine and homoproline residues, as well as a previously undescribed 2,3-dihydroxy-2,6,8-trimethyldeca-(4Z,6E)-dienoic acid moiety. Both 189 and 190 inhibited the infection of human T-lymphoblastoid cells with HIV-1RF in vitro, with an EC50 of 4 ng/mL, and 189 was also cytotoxic against a panel of human cancer cell lines, with a mean IC50 value of 75 ng/mL [83]. Pipecolidepsins A and B (197 and 198) contain unusual residues, including 2-amino-3-hydroxy-4,5-dimethylhexanoic acid, 3-ethoxyasparagine,3,4-dimethylglutamine,4,7-diamino-2,3-dihydroxy-7-oxoheptanoic acid and 3-hydroxyaspartic acid, as well as a terminal 3-hydroxy-2,4,6-trimethylheptanoic acid residue [84]. Polydiscamides B–D (200–202) were the first examples of nonendogenous human SNSR agonists [85]. Two new HIV inhibitory depsipeptides, stellettapeptins A (211) and B (212), were the first peptides reported to contain a 3-hydroxy-6,8-dimethylnon-4-(Z)-enoic acid moiety [86]. Theopapuamide (213) was strongly cytotoxic against CEM-TART and HCT-116 cell lines, with EC50 values of 0.5 and 0.9 µM, respectively [87].
4. Marine Mollusks
At this point in time, a total of thirty depsipeptides (214–243, Figure 9) have been discovered in marine mollusks, including Dolabella, Elysia, Philinopsis and Onchidium [85,86,87,88,89,90,91,92,93,94]. Aurilide (214) was reported as a new 26-membered cyclodepsipeptide and displayed potent cytotoxicity against HeLa S3 cells, with an IC50 of 0.011 μg/mL [88]. Dolastatin D (215) is a cytotoxic cyclodepsipeptide possessing a novel β-amino acid (2R,3R)-3-amino-2-methylbutanoic acid residue [89]. Two 35-membered depsipeptides, dolastatin G (218) and nordolastatin G (219), exhibited cytotoxicity against HeLa S3 cells, with IC50 values of 1.0 and 5.3 µg/mL, respectively [90]. Dolastatin 14 (220), derived from the Indian Ocean sea hare Dolabella auricularia, was shown to be a novel cytostatic (PS EDm 0.022 μg/mL) agent [91]. Kahalalides Z1 (231) and Z2 (232) displayed potent antifungal properties and strong anticancer activities [92].
Two cytotoxic cyclodepsipeptides (233 and 234) containing two unusual amino acids, 4-phenylvaline and 3-amino-2-methylhexanoic acid, were purified from the cephalaspidean mollusk Philinopsis speciosa [93,94]. Kulolide, a cyclic depsipeptide, was isolated from a cephalaspidean mollusk, Philinopsis speciosa Pease [95], and kulolide (235), possessing a rare Dhoya residue, displayed a strong cytotoxic effect on L-1210 leukemia cells and P388 murine leukemia cells, with IC50 values of 0.7 and 2.1 µg/mL, respectively [95]. Onchidin A (241) was structurally determined to have a new β-amino acid, 3-amino-2-methyloct-7-ynoic acid (Amo) [96], while its analog onchidin B (242) contains two 2-hydroxy-3-methylpentanoic acid (Hmp) moieties and two 3-hydroxy-2-methyloct-7-ynoic acid (Hymo) units [97].
5. Marine Fungi
A growing body of evidence has indicated that marine fungi are important sources of depsipeptides, and the genus Fusarium is the most common producer. To date, all marine-fungus-derived depsipeptides (244–259, Figure 10) are cyclic and have a wide array of biological properties [95,96,97,98,99,100]. Enniatin G (245) has inhibitory activity against Heps 7402, with an ED50 value of 12 μg/mL [98]. Two cyclohexadepsipeptides, fusarihexin A (246) and fusarihexin B (247), are manufactured by the marine mangrove endophytic fungus Fusarium sp. R5 and exhibited stronger inhibitory activity against the plant pathogenic fungi C. gloeosporioides, C. musae and F. oxysporum than carbendazim, which is widely used as an agricultural and horticultural fungicide worldwide [99]. HA 23 (250) was reported as a novel cyclodepsipeptide containing a 14-carbon polyketide unit, a substituted tyrosine and pipecolinic acid [100]. Sansalvamide (254) exhibited selective in vitro cytotoxicity toward COLO 205 colon and SK-MEL-2 melanoma cancer cell lines [101]. W493 A-D (255-258) possess the unique residue 3-hydroxy-4-methyl-tetradecanoic acid (Hmta) and exhibited moderate activity against Cladosporium cladosporiodes and weak antitumor activity against the human ovarian cancer cell line A2780 [102,103]. Compound 261 had significant cytotoxicity in the NCI-60 cell line panel (median GI50 = 9.1μM), with highly enhanced selectivity against the CNS cancer cell line SF-268 (GI50 = 6.5 nM) and the renal cancer cell line RXF 393 (GI50 ≤ 5.0 nM) [75].
6. Marine Bacteria
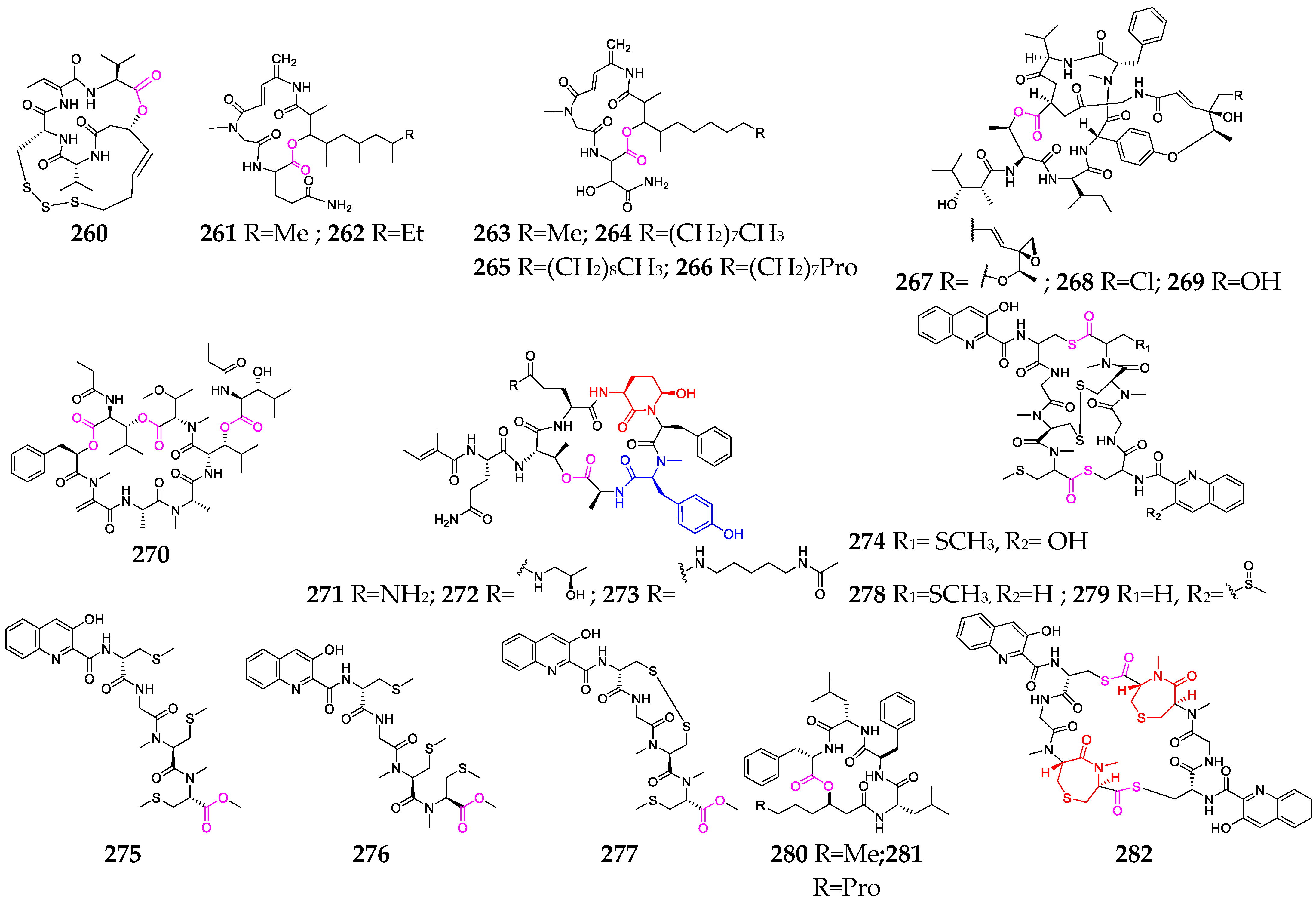
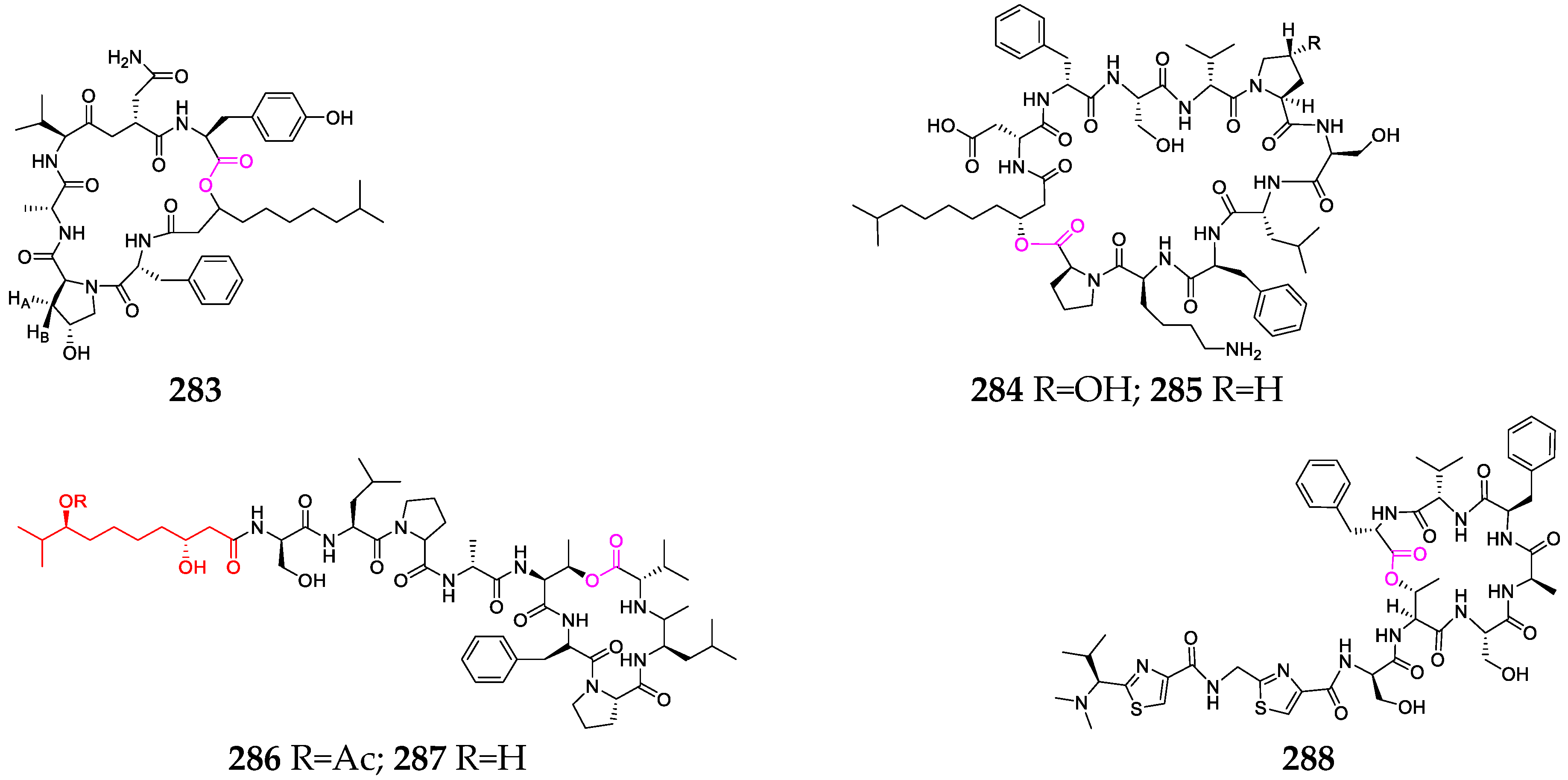
Over twenty depsipeptides (260–282, Figure 11) have been identified in marine bacterial genera, including Mciromonospora, Streptomyces, Chromobacterium, Verrucosispora and Photobacterium [104]. Chromopeptide A (260) was investigated as a novel bicyclic depsipeptide and was found to suppress the proliferation of HL-60, K-562 and Ramos cells, with average IC50 values of 7.7, 7.0 and 16.5 nmol/L, respectively [105]. Rakicidins G–I (264–266), containing a long aliphatic chain without terminal methyl branching, were found to be 18.2~20.3-fold and 7.4~8.7-fold more cytotoxic under hypoxic than under normoxic conditions toward PANC-1 and HCT-8, respectively, and exhibited potent antibacterial effects against Gram-positive anaerobic bacteria [80]. Salinamides A (267) and B (268), sharing a rigid bicyclic hexadepsipeptide core with two esters and an aromatic ether link, showed significant topical anti-inflammatory activity [81]. Streptopeptolins A–C (271–273), containing the unusual amino acids Ahp and N-methyl tyrosine, were the first cyanopeptolin-type peptides isolated from S. olivochromogenes strain NBRC 3561 and demonstrated potent inhibitory activities against chymotrypsin [106]. Thiocoraline (274), produced by Micromonospora sp. strain L-13-ACM2-092, showed potent cytotoxic activity against P-388, A-549 and MEL-28 cell lines, as well as strong antimicrobial activity against Gram-positive bacteria by binding to supercoiling DNA and inhibiting RNA synthesis. Unnarmicins A (280) and C (281) selectively inhibited the growth of two strains belonging to the genus Pseudovibrio, one of the most prevalent genera in the marine environment [107]. Verrucosamide (282), composed of two rare seven-membered 1,4-thiazepane rings, was shown to have moderate cytotoxicity against MDA-MB-468 and COLO 205, with LD50 values of 1.26 µM and 1.4 µM, respectively [108].
7. Marine Algae
To date, a half dozen depsipeptides (283–288, Figure 12) have been obtained from two marine macroalgae, Bryopsis and Derbesia [108]. Mebamamides A (286) and B (287) were reported as new lipopeptides with four D-amino acid residues and a 3,8-dihydroxy-9-methyldecanoic acid residue, but exhibited no growth inhibitory activity against HeLa and HL60 cells at 10 μM [109].
8. Conclusions and Perspectives
In summary, as many as 288 depsipeptides have been discovered in marine organisms, including cyanobacteria, sponges, mollusks, bacteria, fungi and algae, among which marine cyanobacteria are the largest group of producers. Most of these substances are formed by closing the loops of their terminal amino acids. It is very exciting that a large number of marine-derived cyclodepsipeptides display potent cytotoxic effects since they have absolute advantages in structural rigidity, biochemical stability, binding affinity and membrane permeability, which greatly improve their anticancer activity [110], such as the hormones or hormone analogs oxytocin [111], octreotide [112] and vasopressin [113], the antibiotics vancomycin [114], daptomycin [115] and polymyxin B [116] and the immunosuppressant cyclosporine [117]. Therefore, the discovery of novel marine cyclodepsipeptides for new drug development has been attractive to academic researchers and pharmaceutical companies. In the past decade, however, the number of new marine depsipeptides has been greatly reduced, as almost all accessible marine organisms have been collected and chemically studied. Fortunately, marine microorganisms (such as Fusarium, Mciromonospora, Streptomyces) have been shown to be a rich and unexploited source of bioactive natural products due to vast species richness and the biosynthetic potential of secondary metabolites, especially those of symbiotic microbes in marine sponges, mollusks, tunicates, macroalgae and mangroves. Therefore, more efforts should be made toward strain separation and chemical research using classical methods (e.g., strain cultivation and fermentation, chromatographic and spectroscopic techniques) and advanced approaches (e.g., metabolomics, genome mining and engineering).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/md21020120/s1, Table S1: Detail information for marine cyanobacterium-derived depsipeptides (1-161); Table S2. Detail information for marine sponge-derived depsipeptides (162–213); Table S3: Detail information for marine mollusk-derived depsipeptides (214–243); Table S4 Detail information for marine fungus-derived depsipeptides (244–259); Table S5: Detail information for marine bacterium-derived depsipeptides (260–282); Table S6. Detail information for marine algae-derived depsipeptides (283–288).
Author Contributions
Conceptualization and funding acquisition, H.Z.; project administration, M.Z. and H.Z.; writing—original draft, M.Z., J.T. and S.X.; writing—review and editing, H.Z. and X.B. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support from the National Key Research and Development Program of China (2022YFC2804203), the National Natural Science Foundation of China (41776139) and the General Research Project of Education Department of Zhejiang Province (Y202147473) is greatly appreciated.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Papon, N.; Copp, B.R.; Courdavault, V. Marine drugs: Biology, pipelines, current and future prospects for production. Biotechnol. Adv. 2022, 54, 0734–9750. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Asgher, M.; Sher, F.; Hussain, S.M.; Nazish, N.; Joshi, N.; Sharma, A.; Parra-Saldívar, R.; Bilal, M.; Iqbal, H.M.N. Exploring marine as a rich source of bioactive peptides: Challenges and opportunities from marine pharmacology. Mar. Drugs 2022, 20, 208. [Google Scholar] [CrossRef]
- Alonzo, D.A.; Schmeing, T.M. Biosynthesis of depsipeptides, or Depsi: The peptides with varied generations. Protein Sci. 2020, 29, 2316–2347. [Google Scholar] [CrossRef] [PubMed]
- Bachy, E.; Camus, V.; Thieblemont, C.; Sibon, D.; Casasnovas, R.O.; Ysebaert, L.; Damaj, G.; Guidez, S.; Pica, G.M.; Kim, W.S.; et al. Romidepsin Plus CHOP Versus CHOP in Patients with Previously Untreated Peripheral T-Cell Lymphoma: Results of the Ro-CHOP Phase III Study (Conducted by LYSA). J. Clin. Oncol. 2022, 40, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Falchi, L.; Ma, H.; Klein, S.; Lue, J.K.; Montanari, F.; Marchi, E.; Deng, C.; Kim, H.A.; Rada, A.; Jacob, A.T.; et al. Combined oral 5-azacytidine and romidepsin are highly effective in patients with PTCL: A multicenter phase 2 study. Blood 2021, 137, 2161–2170. [Google Scholar] [CrossRef]
- Leisch, M.; Egle, A.; Greil, R. Plitidepsin: A potential new treatment for relapsed/refractory multiple myeloma. Future Oncol. 2019, 15, 109–120. [Google Scholar] [CrossRef]
- Broggini, M.; Marchini, S.V.; Galliera, E.; Borsotti, P.; Taraboletti, G.; Erba, E.; Sironi, M.; Jimeno, J.; Faircloth, G.T.; Giavazzi, R.; et al. Aplidine, a new anticancer agent of marine origin, inhibits vascular endothelial growth factor (VEGF) secretion and blocks VEGF-VEGFR-1 (flt-1) autocrine loop in human leukemia cells MOLT-4. Leukemia 2003, 17, 52–59. [Google Scholar] [CrossRef]
- Miguel-Lillo, B.; Valenzuela, B.; Peris-Ribera, J.E.; Soto-Matos, A.; Pérez-Ruixo, J.J. Population pharmacokinetics of kahalalide F in advanced cancer patients. Cancer Chemother. Pharm. 2015, 76, 365–374. [Google Scholar] [CrossRef]
- Wyer, S.; Townsend, D.M.; Ye, Z.; Kourtidis, A.; Choo, Y.M.; de Barros, A.L.B.; Donia, M.S.; Hamann, M.T. Recent advances and limitations in the application of kahalalides for the control of cancer. Biomed. Pharmacother. 2022, 148, 112676. [Google Scholar] [CrossRef]
- Heath, E.I.; Weise, A.; Vaishampayan, U.; Danforth, D.; Ungerleider, R.S.; Urata, Y. Phase Ia dose escalation study of OBP-801, a cyclic depsipeptide class I histone deacetylase inhibitor, in patients with advanced solid tumors. Investig. New Drugs 2022, 40, 300–307. [Google Scholar] [CrossRef]
- Gao, F.-x.; Tian, M. Research progress on the depsipeptides. Chin. J. Antibiot. 2018, 43, 777–785. [Google Scholar]
- Rangel, M.; de Santana, C.J.; Pinheiro, A.; Dos Anjos, L.; Barth, T.; Pires, O.R.; Fontes, W.; Castro, M.S. Marine depsipeptides as promising pharmacotherapeutic agents. Curr. Protein Pept. Sci. 2017, 18, 72–91. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Zhang, J.R.; He, S.; Yan, X.J. New peptides isolated from marine cyanobacteria, an overview over the past decade. Mar. Drugs 2017, 15, 132. [Google Scholar] [CrossRef] [PubMed]
- Al-Awadhi, F.H.; Law, B.K.; Paul, V.J.; Luesch, H. Grassystatins D–F, potent aspartic protease inhibitors from marine cyanobacteria as potential antimetastatic agents targeting invasive breast cancer. J. Nat. Prod. 2017, 80, 2969–2986. [Google Scholar] [CrossRef]
- Choi, H.; Pereira, A.R.; Cao, Z.; Shuman, C.F.; Engene, N.; Byrum, T.; Matainaho, T.; Murray, T.F.; Mangoni, A.; Gerwick, W.H. The hoiamides, structurally intriguing neurotoxic lipopeptides from Papua New Guinea marine cyanobacteria. J. Nat. Prod. 2010, 73, 1411–1421. [Google Scholar] [CrossRef]
- Malloy, K.L.; Choi, H.; Fiorilla, C.; Valeriote, F.A.; Matainaho, T.; Gerwick, W.H. Hoiamide D, a marine cyanobacteria-derived inhibitor of p53/MDM2 interaction. Bioorg. Med. Chem. Lett. 2012, 22, 683–688. [Google Scholar] [CrossRef]
- Horgen, F.D.; Kazmierski, E.B.; Westenburg, H.E.; Yoshida, W.Y.; Scheuer, P.J. Malevamide D: Isolation and structure determination of an isodolastatin H analogue from the marine cyanobacterium Symploca hydnoides. J. Nat. Prod. 2002, 65, 487–491. [Google Scholar] [CrossRef]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H. Symplostatin 3, a new dolastatin 10 analogue from the marine cyanobacterium Symploca sp. VP452. J. Nat. Prod. 2002, 65, 16–20. [Google Scholar] [CrossRef]
- Williams, P.G.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Tasiamide, a cytotoxic peptide from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2002, 65, 1336–1339. [Google Scholar] [CrossRef]
- Williams, P.G.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. The isolation and structure elucidation of Tasiamide B, a 4-amino-3-hydroxy-5-phenylpentanoic acid containing peptide from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2003, 66, 1006–1009. [Google Scholar] [CrossRef]
- Mevers, E.; Haeckl, F.P.; Boudreau, P.D.; Byrum, T.; Dorrestein, P.C.; Valeriote, F.A.; Gerwick, W.H. Lipopeptides from the tropical marine cyanobacterium Symploca sp. J. Nat. Prod. 2014, 77, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Al-Awadhi, F.H.; Ratnayake, R.; Paul, V.J.; Luesch, H. Tasiamide F, a potent inhibitor of cathepsins D and E from a marine cyanobacterium. Bioorg. Med. Chem. 2016, 24, 3276–3282. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, Y.; Iwasaki, A.; Sumimoto, S.; Matsubara, T.; Sato, T.; Suenaga, K. Izenamides A and B, statine-containing depsipeptides, and an analogue from a marine cyanobacterium. J. Nat. Prod. 2018, 81, 1673–1681. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.C.; Eksioglu, E.A.; Liu, C.; Paul, V.J.; Luesch, H. Grassystatins A–C from marine cyanobacteria, potent cathepsin E inhibitors that reduce antigen presentation. J. Med. Chem. 2009, 52, 5732–5747. [Google Scholar] [CrossRef]
- Iwasaki, A.; Ohno, O.; Sumimoto, S.; Suda, S.; Suenaga, K. Maedamide, a novel chymotrypsin inhibitor from a marine cyanobacterial assemblage of Lyngbya sp. Tetrahedron Lett. 2014, 55, 4126–4128. [Google Scholar] [CrossRef]
- Williams, P.G.; Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Continuing studies on the cyanobacterium Lyngbya sp.: Isolation and structure determination of 15-norlyngbyapeptin A and lyngbyabellin D. J. Nat. Prod. 2003, 66, 595–598. [Google Scholar] [CrossRef]
- Petitbois, J.G.; Casalme, L.O.; Lopez, J.A.V.; Alarif, W.M.; Abdel-Lateff, A.; Al-Lihaibi, S.S.; Yoshimura, E.; Nogata, Y.; Umezawa, T.; Matsuda, F.; et al. Serinolamides and Lyngbyabellins from an Okeania sp. Cyanobacterium Collected from the Red Sea. J. Nat. Prod. 2017, 80, 2708–2715. [Google Scholar] [CrossRef]
- Linington, R.G.; Clark, B.R.; Trimble, E.E.; Almanza, A.; Ureña, L.D.; Kyle, D.E.; Gerwick, W.H. Antimalarial peptides from marine cyanobacteria: Isolation and structural elucidation of gallinamide A. J. Nat. Prod. 2009, 72, 14–17. [Google Scholar] [CrossRef]
- Mevers, E.; Liu, W.-T.; Engene, N.; Mohimani, H.; Byrum, T.; Pevzner, P.A.; Dorrestein, P.C.; Spadafora, C.; Gerwick, W.H. Cytotoxic veraguamides, alkynyl bromide-containing cyclic depsipeptides from the marine cyanobacterium cf. Oscillatoria margaritifera. J. Nat. Prod. 2011, 74, 928–936. [Google Scholar] [CrossRef]
- Rubio, B.K.; Parrish, S.M.; Yoshida, W.; Schupp, P.J.; Schils, T.; Williams, P.G. Depsipeptides from a guamanian marine cyanobacterium, Lyngbya bouillonii, with selective inhibition of serine proteases. Tetrahedron Lett. 2010, 51, 6718–6721. [Google Scholar] [CrossRef]
- Gunasekera, S.P.; Owle, C.S.; Montaser, R.; Luesch, H.; Paul, V.J. Malyngamide 3 and cocosamides A and B from the marine cyanobacterium Lyngbya majuscula from cocos lagoon, guam. J. Nat. Prod. 2011, 74, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T.; Sitachitta, N.; Gerwick, W.H. The guineamides, novel cyclic depsipeptides from a papua new guinea collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2003, 66, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Taori, K.; Paul, V.J.; Luesch, H. Kempopeptins A and B, serine protease inhibitors with different selectivity profiles from a marine cyanobacterium, Lyngbya sp. J. Nat. Prod. 2008, 71, 1625–1629. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Salvador, L.A.; Law, B.K.; Paul, V.J.; Luesch, H. Kempopeptin C, a novel marine-derived serine protease inhibitor targeting invasive breast cancer. Mar. Drugs 2017, 15, 290. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Sumimoto, S.; Ohno, O.; Suda, S.; Suenaga, K. Kurahamide, a cyclic depsipeptide analog of dolastatin 13 from a marine cyanobacterial assemblage of Lyngbya sp. Bull. Chem. Soc. Jpn. 2014, 87, 609–613. [Google Scholar] [CrossRef]
- Matthew, S.; Ross, C.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatin 4, a dolastatin 13 analogue with elastase and chymotrypsin inhibitory activity from the marine cyanobacterium Lyngbya confervoides. J. Nat. Prod. 2007, 70, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Taori, K.; Matthew, S.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatins 5-7, potent elastase inhibitors from floridian marine cyanobacteria, Lyngbya spp. J. Nat. Prod. 2007, 70, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.C.; Taori, K.; Paul, V.J.; Luesch, H. Lyngbyastatins 8-10, elastase inhibitors with cyclic depsipeptide scaffolds isolated from the marine cyanobacterium Lyngbya semiplena. Mar. Drugs 2009, 7, 528–538. [Google Scholar] [CrossRef]
- Matthew, S.; Ross, C.; Paul, V.J.; Luesch, H. Pompanopeptins A and B, new cyclic peptides from the marine cyanobacterium Lyngbya confervoides. Tetrahedron 2008, 64, 4081–4089. [Google Scholar] [CrossRef]
- Ozaki, K.; Iwasaki, A.; Suenaga, K.; Teruya, T. Kyanamide, a new Ahp-containing depsipeptide from marine cyanobacterium Caldora penicillata. Tetrahedron 2019, 75, 3382–3386. [Google Scholar] [CrossRef]
- Gallegos, D.A.; Sauri, J.; Cohen, R.D.; Wan, X.; Videau, P.; Vallota-Eastman, A.O.; Shaala, L.A.; Youssef, D.T.A.; Williamson, R.T.; Martin, G.E.; et al. Jizanpeptins, cyanobacterial protease inhibitors from a Symploca sp. cyanobacterium collected in the red sea. J. Nat. Prod. 2018, 81, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, G.G.; Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Nagle, D.G.; Paul, V.J. Symplostatin 2: A dolastatin 13 analogue from the marine cyanobacterium Symploca hydnoides. J. Nat. Prod. 1999, 62, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Salvador, L.A.; Taori, K.; Biggs, J.S.; Jakoncic, J.; Ostrov, D.A.; Paul, V.J.; Luesch, H. Potent elastase inhibitors from cyanobacteria: Structural basis and mechanisms mediating cytoprotective and anti-inflammatory effects in bronchial epithelial cells. J. Med. Chem. 2013, 56, 1276–1290. [Google Scholar] [CrossRef] [PubMed]
- Phyo, M.Y.; Katermeran, N.P.; Goh, J.X.; Tan, L.T. Trikoveramides A-C, cyclic depsipeptides from the marine cyanobacterium Symploca hydnoides. Phytochemistry 2021, 190, 112879. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Bewley, C.A. Largamides A-H, unusual cyclic peptides from the marine cyanobacterium Oscillatoria sp. J. Org. Chem. 2006, 71, 6898–6907. [Google Scholar] [CrossRef] [PubMed]
- Al-Awadhi, F.H.; Paul, V.J.; Luesch, H. Structural diversity and anticancer activity of marine-derived elastase inhibitors: Key features and mechanisms mediating the antimetastatic effects in invasive breast cancer. ChemBioChem 2018, 19, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Miller, M.W.; Kwan, J.C.; Luesch, H.; Paul, V.J. Molassamide, a depsipeptide serine protease inhibitor from the marine cyanobacterium Dichothrix utahensis. J. Nat. Prod. 2010, 73, 459–462. [Google Scholar] [CrossRef]
- Sueyoshi, K.; Kaneda, M.; Sumimoto, S.; Oishi, S.; Fujii, N.; Suenaga, K.; Teruya, T. Odoamide, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Okeania sp. Tetrahedron 2016, 72, 5472–5478. [Google Scholar] [CrossRef]
- Keller, L.; Canuto, K.M.; Liu, C.; Suzuki, B.M.; Almaliti, J.; Sikandar, A.; Naman, C.B.; Glukhov, E.; Luo, D.; Duggan, B.M.; et al. Tutuilamides A-C: Vinyl-chloride-containing cyclodepsipeptides from marine cyanobacteria with potent elastase inhibitory properties. ACS Chem. Biol. 2020, 15, 751–757. [Google Scholar] [CrossRef]
- Taniguchi, M.; Nunnery, J.K.; Engene, N.; Esquenazi, E.; Byrum, T.; Dorrestein, P.C.; Gerwick, W.H. Palmyramide A, a cyclic depsipeptide from a Palmyra Atoll collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 393–398. [Google Scholar] [CrossRef]
- Han, B.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Aurilides B and C, cancer cell toxins from a papua new guinea collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006, 69, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Bunyajetpong, S.; Yoshida, W.Y.; Sitachitta, N.; Kaya, K. Trungapeptins A-C, cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006, 69, 1539–1542. [Google Scholar] [CrossRef] [PubMed]
- Salvador, L.A.; Biggs, J.S.; Paul, V.J.; Luesch, H. Veraguamides A-G, cyclic hexadepsipeptides from a dolastatin 16-producing cyanobacterium Symploca cf. hydnoides from Guam. J. Nat. Prod. 2011, 74, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Shiota, I.; Sumimoto, S.; Matsubara, T.; Sato, T.; Suenaga, K. Kohamamides A, B, and C, cyclic depsipeptides from an Okeania sp. marine cyanobacterium. J. Nat. Prod. 2017, 80, 1948–1952. [Google Scholar] [CrossRef]
- Luo, D.; Putra, M.Y.; Ye, T.; Paul, V.J.; Luesch, H. Isolation, structure elucidation and biological evaluation of Lagunamide D: A new cytotoxic macrocyclic depsipeptide from marine cyanobacteria. Mar. Drugs 2019, 17, 83. [Google Scholar] [CrossRef]
- Phyo, M.Y.; Goh, J.X.; Tan, L.T. Triproamide and Pemukainalides, cyclic depsipeptides from the marine cyanobacterium Symploca hydnoides. J. Nat. Prod. 2022, 85, 485–492. [Google Scholar] [CrossRef]
- Boudreau, P.D.; Byrum, T.; Liu, W.T.; Dorrestein, P.C.; Gerwick, W.H. Viequeamide A, a cytotoxic member of the kulolide superfamily of cyclic depsipeptides from a marine button cyanobacterium. J. Nat. Prod. 2012, 75, 1560–1570. [Google Scholar] [CrossRef]
- Kwan, J.C.; Ratnayake, R.; Abboud, K.A.; Paul, V.J.; Luesch, H. Grassypeptolides A-C, cytotoxic bis-thiazoline containing marine cyclodepsipeptides. J. Org. Chem. 2010, 75, 8012–8023. [Google Scholar] [CrossRef]
- Thornburg, C.C.; Thimmaiah, M.; Shaala, L.A.; Hau, A.M.; Malmo, J.M.; Ishmael, J.E.; Youssef, D.T.; McPhail, K.L. Cyclic depsipeptides, grassypeptolides D and E and Ibu-epidemethoxylyngbyastatin 3, from a Red Sea Leptolyngbya cyanobacterium. J. Nat. Prod. 2011, 74, 1677–1685. [Google Scholar] [CrossRef]
- Popplewell, W.L.; Ratnayake, R.; Wilson, J.A.; Beutler, J.A.; Colburn, N.H.; Henrich, C.J.; McMahon, J.B.; McKee, T.C. Grassypeptolides F and G, cyanobacterial peptides from Lyngbya majuscula. J. Nat. Prod. 2011, 74, 1686–1691. [Google Scholar] [CrossRef]
- Pereira, A.; Cao, Z.; Murray, T.F.; Gerwick, W.H. Hoiamide a, a sodium channel activator of unusual architecture from a consortium of two papua new Guinea cyanobacteria. Chem. Biol. 2009, 16, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; McPhail, K.L.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Isolation and structure of five lyngbyabellin derivatives from a papua new guinea collection of the marine cyanobacterium Lyngbya majuscula. Tetrahedron 2005, 61, 11723–11729. [Google Scholar] [CrossRef]
- Williams, P.G.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Isolation and structure determination of obyanamide, a novel cytotoxic cyclic depsipeptide from the marine cyanobacterium Lyngbya confervoides. J. Nat. Prod. 2002, 65, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Gross, H.; McPhail, K.L.; Goeger, D.; Maier, C.S.; Gerwick, W.H. Wewakamide A and guineamide G, cyclic depsipeptides from the marine cyanobacteria Lyngbya semiplena and Lyngbya majuscula. J. Microbiol. Biotechnol. 2011, 21, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.L.; Nogle, L.M.; Media, J.; Valeriote, F.A.; Mooberry, S.L.; Gerwick, W.H. Desmethoxymajusculamide C, a cyanobacterial depsipeptide with potent cytotoxicity in both cyclic and ring-opened forms. J. Nat. Prod. 2009, 72, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Davies-Coleman, M.T.; Dzeha, T.M.; Gray, C.A.; Hess, S.; Pannell, L.K.; Hendricks, D.T.; Arendse, C.E. Isolation of homodolastatin 16, a new cyclic depsipeptide from a kenyan collection of Lyngbya majuscula. J. Nat. Prod. 2003, 66, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, G.G.; Yoshida, W.Y.; Moore, R.E.; Nagle, D.G.; Park, P.U.; Biggs, J.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H.; Valeriote, F.A. Isolation, structure determination, and biological activity of dolastatin 12 and lyngbyastatin 1 from Lyngbya majuscula/Schizothrix calcicola cyanobacterial assemblages. J. Nat. Prod. 1998, 61, 1221–1225. [Google Scholar] [CrossRef]
- Williams, P.G.; Moore, R.E.; Paul, V.J. Isolation and structure determination of lyngbyastatin 3, a lyngbyastatin 1 homologue from the marine cyanobacterium Lyngbya majuscula. Determination of the configuration of the 4-amino-2,2-dimethyl-3-oxopentanoic acid unit in majusculamide C, dolastatin 12, lyngbyastatin 1, and lyngbyastatin 3 from cyanobacteria. J. Nat. Prod. 2003, 66, 1356–1363. [Google Scholar]
- Han, B.; Goeger, D.; Maier, C.S.; Gerwick, W.H. The wewakpeptins, cyclic depsipeptides from a papua new guinea collection of the marine cyanobacterium Lyngbya semiplena. J. Org. Chem. 2005, 70, 3133–3139. [Google Scholar] [CrossRef]
- Adams, B.; Porzgen, P.; Pittman, E.; Yoshida, W.Y.; Westenburg, H.E.; Horgen, F.D. Isolation and structure determination of malevamide E, a dolastatin 14 analogue, from the marine cyanobacterium symploca laete-viridis. J. Nat. Prod. 2008, 71, 750–754. [Google Scholar] [CrossRef]
- Vining, O.B.; Medina, R.A.; Mitchell, E.A.; Videau, P.; Li, D.; Serrill, J.D.; Kelly, J.X.; Gerwick, W.H.; Proteau, P.J.; Ishmael, J.E.; et al. Depsipeptide companeramides from a Panamanian marine cyanobacterium associated with the coibamide producer. J. Nat. Prod. 2015, 78, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Stratmann, K.; Burgoyne, D.L.; Moore, R.E.; Patterson, G.M.L.; Smith, C.D. Hapalosin, a cyanobacterial cyclic depsipeptide with multidrug-resistance reversing activity. J. Org. Chem. 2002, 59, 7219–7226. [Google Scholar] [CrossRef]
- Kanamori, Y.; Iwasaki, A.; Sumimoto, S.; Suenaga, K. Urumamide, a novel chymotrypsin inhibitor with a β-amino acid from a marine cyanobacterium Okeania sp. Tetrahedron Lett. 2016, 57, 4213–4216. [Google Scholar] [CrossRef]
- Zampella, A.; D’Auria, M.V.; Paloma, L.G.; Casapullo, A.; Minale, L.; Debitus, C.; Henin, Y. Callipeltin A, an anti-HIV cyclic depsipeptide from the new caledonian lithistida sponge Callipelta sp. J. Am. Chem. Soc. 1996, 118, 6202–6209. [Google Scholar] [CrossRef]
- Stierhof, M.; Hansen, K.Ø.; Sharma, M.; Feussner, K.; Subko, K.; Díaz-Rullo, F.F.; Isaksson, J.; Pérez-Victoria, I.; Clarke, D.; Hansen, E.; et al. New cytotoxic callipeltins from the Solomon Island marine sponge Asteropus sp. Tetrahedron 2016, 72, 6929–6934. [Google Scholar] [CrossRef]
- Clark, D.P.; Carroll, J.; Naylor, S.; Crews, P. An antifungal cyclodepsipeptide, Cyclolithistide A, from the sponge Theonella swinhoei. J. Org. Chem. 1998, 63, 8757–8764. [Google Scholar] [CrossRef]
- Urda, C.; Fernandez, R.; Rodriguez, J.; Perez, M.; Jimenez, C.; Cuevas, C. Daedophamide, a cytotoxic cyclodepsipeptide from a Daedalopelta sp. sponge collected in indonesia. J. Nat. Prod. 2017, 80, 3054–3059. [Google Scholar] [CrossRef] [PubMed]
- Tarazona, G.; Fernández, R.; Cruz, P.G.; Pérez, M.; Rodríguez, J.; Jiménez, C.; Cuevas, C. Combining JBCA and Marfey’s methodology to determine the absolute configuration of threonines: The case of gunungamide A, a new cyclic depsipeptide containing chloropyrrole from the sponge Discodermia sp. Org. Chem. Front. 2019, 6, 15–21. [Google Scholar] [CrossRef]
- Zampella, A.; Sepe, V.; Luciano, P.; Bellotta, F.; Monti, M.C.; D’Auria, M.V.; Jepsen, T.; Petek, S.; Adeline, M.T.; Laprevote, O.; et al. Homophymine A, an anti-HIV cyclodepsipeptide from the sponge Homophymia sp. J. Org. Chem. 2008, 73, 5319–5327. [Google Scholar] [CrossRef]
- Zampella, A.; Sepe, V.; Bellotta, F.; Luciano, P.; D’Auria, M.V.; Cresteil, T.; Debitus, C.; Petek, S.; Poupat, C.; Ahond, A. Homophymines B-E and A1-E1, a family of bioactive cyclodepsipeptides from the sponge Homophymia sp. Org. Biomol. Chem. 2009, 7, 4037–4044. [Google Scholar] [CrossRef]
- Rashid, M.A.; Gustafson, K.R.; Cartner, L.K.; Shigematsu, N.; Pannell, L.K.; Boyd, M.R. Microspinosamide, a new HIV-inhibitory cyclic depsipeptide from the marine sponge Sidonops microspinosa. J. Nat. Prod. 2001, 64, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Gustchina, E.; Baker, H.L.; Kelly, M.; Bewley, C.A. Mirabamides A-D, depsipeptides from the sponge Siliquariaspongia mirabilis that inhibit HIV-1 fusion. J. Nat. Prod. 2007, 70, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Ford, P.W.; Gustafson, K.R.; McKee, T.C.; Shigematsu, N.; Maurizi, L.K.; Pannell, L.K.; Williams, D.E.; Dilip de Silva, E.; Lassota, P.; Allen, T.M.; et al. Papuamides A−D, HIV-inhibitory and cytotoxic depsipeptides from the sponges Theonella mirabilis and Theonella swinhoei collected in papua new guinea. J. Am. Chem. Soc. 1999, 121, 5899–5909. [Google Scholar] [CrossRef]
- Coello, L.; Reyes, F.; Martin, M.J.; Cuevas, C.; Fernandez, R. Isolation and structures of pipecolidepsins A and B, cytotoxic cyclic depsipeptides from the madagascan sponge Homophymia lamellosa. J. Nat. Prod. 2014, 77, 298–303. [Google Scholar] [CrossRef]
- Feng, Y.; Carroll, A.R.; Pass, D.M.; Archbold, J.K.; Avery, V.M.; Quinn, R.J. Polydiscamides B-D from a marine sponge Ircinia sp. as potent human sensory neuron-specific G protein coupled receptor agonists. J. Nat. Prod. 2008, 71, 8–11. [Google Scholar] [CrossRef]
- Shin, H.J.; Rashid, M.A.; Cartner, L.K.; Bokesch, H.R.; Wilson, J.A.; McMahon, J.B.; Gustafson, K.R. Stellettapeptins A and B, HIV-inhibitory cyclic depsipeptides from the marine sponge Stelletta sp. Tetrahedron Lett. 2015, 56, 4215–4219. [Google Scholar] [CrossRef]
- Ratnayake, A.S.; Bugni, T.S.; Feng, X.; Harper, M.K.; Skalicky, J.J.; Mohammed, K.A.; Andjelic, C.D.; Barrows, L.R.; Ireland, C.M. Theopapuamide, a cyclic depsipeptide from a Papua New Guinea lithistid sponge Theonella swinhoei. J. Nat. Prod. 2006, 69, 1582–1586. [Google Scholar] [CrossRef]
- Suenaga, K.; Mutou, T.; Shibata, T.; Itoh, T.; Fujita, T.; Takada, N.; Hayamizu, K.; Takagi, M.; Irifune, T.; Kigoshi, H.; et al. Aurilide, a cytotoxic depsipeptide from the sea hare Dolabella auricularia: Isolation, structure determination, synthesis, and biological activity. Tetrahedron 2004, 60, 8509–8527. [Google Scholar] [CrossRef]
- Sone, H.; Nemoto, T.; Ishiwata, H.; Ojika, M.; Yamada, K. Isolation, structure, and synthesis of dolastatin D, a cytotoxic cyclic depsipeptide from the sea hare Dolabella abricularia. Tetrahedron Lett. 1993, 34, 8449–8452. [Google Scholar] [CrossRef]
- Mutou, T.; Kondo, T.; Ojika, M.; Yamada, K. Isolation and stereostructures of dolastatin G and nordolastatin G, cytotoxic 35-membered cyclodepsipeptides from the Japanese sea hare Dolabella auricularia. J. Org. Chem. 1996, 61, 6340–6345. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Dufresne, C.; Bates, R.B.; Schmidt, J.M.; Cerny, R.L.; Kizu, H. Antineoplastic agents. 190. isolation and structure of the cyclodepsipeptide dolastatin 14. J. Org. Chem 2002, 55, 2989–2990. [Google Scholar] [CrossRef]
- Ciavatta, M.L.; Devi, P.; Carbone, M.; Mathieu, V.; Kiss, R.; Casapullo, A.; Gavagnin, M. Kahalalide F analogues from the mucous secretion of Indian sacoglossan mollusc Elysia ornata. Tetrahedron 2016, 72, 625–631. [Google Scholar] [CrossRef]
- Kimura, J.; Takada, Y.; Inayoshi, T.; Nakao, Y.; Goetz, G.; Yoshida, W.Y.; Scheuer, P.J. Kulokekahilide-1, a cytotoxic depsipeptide from the cephalaspidean mollusk Philinopsis speciosa. J. Org. Chem. 2002, 67, 1760–1767. [Google Scholar] [CrossRef]
- Nakao, Y.; Yoshida, W.Y.; Takada, Y.; Kimura, J.; Yang, L.; Mooberry, S.L.; Scheuer, P.J. Kulokekahilide-2, a cytotoxic depsipeptide from a cephalaspidean mollusk Philinopsis speciosa. J. Nat. Prod. 2004, 67, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Nakao, Y.; Yoshida, W.Y.; Szabo, C.M.; Baker, B.J.; Scheuer, P.J. More peptides and other diverse constituents of the marine mollusk Philinopsis speciosa. J. Org. Chem. 1998, 63, 3272–3280. [Google Scholar] [CrossRef]
- Rodríguez, J.; Fernández, R.; Quiñoá, E.; Riguera, R.; Debitus, C.; Bouchet, P. Onchidin: A cytotoxic depsipeptide with C2 symmetry from a marine mollusc. Tetrahedron Lett. 1994, 35, 9239–9242. [Google Scholar] [CrossRef]
- Fernández, R.; Rodríguez, J.; Quiñoá, E.; Riguera, R.; Muñoz, L.; Fernández-Suárez, M.; Debitus, C. Onchidin B: A new cyclodepsipeptide from the mollusc Onchidium sp. J. Am. Chem. Soc. 1996, 118, 11635–11643. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, J.; Wu, X.; Zhou, S.; Vrijmoed, L.L.P.; Jones, E.B.G. A novel compound Enniatin G from the mangrove fungus Halosarpheia sp. (strain #732) from the south China sea. Aust. J. Chem. 2002, 55, 225–227. [Google Scholar]
- Zhu, X.; Zhong, Y.; Xie, Z.; Wu, M.; Hu, Z.; Ding, W.; Li, C. Fusarihexins A and B: Novel cyclic hexadepsipeptides from the mangrove endophytic fungus Fusarium sp. R5 with antifungal activities. Planta Med. 2018, 84, 1355–1362. [Google Scholar] [CrossRef]
- Feng, Y.; Blunt, J.W.; Cole, A.L.; Munro, M.H. A novel cyclodepsipeptide, HA23, from a Fusarium sp. Org. Lett. 2002, 4, 2095–2096. [Google Scholar] [CrossRef]
- Belofsky, G.N.; Jensen, P.R.; Fenical, W. Sansalvamide: A new cytotoxic cyclic depsipeptide produced by a marine fungus of the genus Fusarium. Tetrahedron Lett. 1999, 40, 2913–2916. [Google Scholar] [CrossRef]
- Nihei, K.; Itoh, H.; Hashimoto, K.; Miyairi, K.; Okuno, T. Antifungal cyclodepsipeptides, W493 A and B, from Fusarium sp.: Isolation and structural determination. Biosci. Biotechnol. Biochem. 1998, 62, 858–863. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.; Daletos, G.; Lin, W.; Proksch, P. Two new cyclic depsipeptides from the endophytic fungus Fusarium sp. Nat. Prod. Commun. 2015, 10, 1667–1670. [Google Scholar] [CrossRef] [PubMed]
- Haygood, M.G. Light organ symbioses in fishes. Crit. Rev. Microbiol. 1993, 19, 191–216. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, X.; Zhang, H.; Sun, J.; Zheng, L.; Liu, H.; Wang, J.; Shen, A.; Geng, M.; Guo, Y. Chromopeptide A, a highly cytotoxic depsipeptide from the marine sediment-derived bacterium Chromobacterium sp. HS-13-94. Acta. Pharm. Sin. B 2015, 5, 62–66. [Google Scholar] [CrossRef]
- Kodani, S.; Komaki, H.; Hemmi, H.; Miyake, Y.; Kaweewan, I.; Dohra, H. Streptopeptolin, a cyanopeptolin-type peptide from Streptomyces olivochromogenes. ACS. Omega 2018, 3, 8104–8110. [Google Scholar] [CrossRef]
- Oku, N.; Kawabata, K.; Adachi, K.; Katsuta, A.; Shizuri, Y. Unnarmicins A and C, new antibacterial depsipeptides produced by marine bacterium Photobacterium sp. MBIC06485. J. Antibiot. 2008, 61, 11–17. [Google Scholar] [CrossRef]
- Nair, V.; Kim, M.C.; Golen, J.A.; Rheingold, A.L.; Castro, G.A.; Jensen, P.R.; Fenical, W. Verrucosamide, a cytotoxic 1,4-thiazepane-containing thiodepsipeptide from a marine-derived Actinomycete. Mar. Drugs 2020, 18, 549. [Google Scholar] [CrossRef]
- Iwasaki, A.; Ohno, O.; Sumimoto, S.; Matsubara, T.; Shimada, S.; Sato, T.; Suenaga, K. Mebamamides A and B, cyclic lipopeptides isolated from the Green Alga Derbesia marina. J. Nat. Prod. 2015, 78, 901–908. [Google Scholar] [CrossRef]
- Zhang, J.N.; Xia, Y.X.; Zhang, H.J. Natural cyclopeptides as anticancer agents in the last 20 Years. Int. J. Mol. Sci. 2021, 22, 3973. [Google Scholar] [CrossRef]
- Love, T.M. Oxytocin, motivation and the role of dopamine. Pharmacol. Biochem. Behav. 2014, 119, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Battershill, P.E.; Clissold, S.P. Octreotide. A review of its pharmacodynamic and pharmacokinetic properties, and therapeutic potential in conditions associated with excessive peptide secretion. Drugs 1989, 38, 658–702. [Google Scholar] [CrossRef] [PubMed]
- Kam, P.C.; Williams, S.; Yoong, F.F. Vasopressin and terlipressin: Pharmacology and its clinical relevance. Anaesthesia 2004, 59, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, M.P. Vancomycin. Mayo Clin. Proc. 1991, 66, 1165–1170. [Google Scholar] [CrossRef]
- Heidary, M.; Khosravi, A.D.; Khoshnood, S.; Nasiri, M.J.; Soleimani, S.; Goudarzi, M. Daptomycin. J. Antimicrob. Chemother. 2018, 73, 1–11. [Google Scholar] [CrossRef]
- Tran, T.B.; Velkov, T.; Nation, R.L.; Forrest, A.; Tsuji, B.T.; Bergen, P.J.; Li, J. Pharmacokinetics/pharmacodynamics of colistin and polymyxin B: Are we there yet? Int. J. Antimicrob. Agents 2016, 48, 592–597. [Google Scholar] [CrossRef]
- Barshes, N.R.; Goodpastor, S.E.; Goss, J.A. Pharmacologic immunosuppression. Front. Biosci. 2004, 9, 411–420. [Google Scholar] [CrossRef]
Figure 1.
Percentage distribution of depsipeptides from marine organisms.

Figure 2.
Marine-cyanobacterium-derived linear lipopeptides (1–27).

Figure 3.
Marine-cyanobacterium-derived cyclopentadepsipeptides (28–81).


Figure 4.
Marine-cyanobacterium-derived cyclohexadepsipeptides (82-103).

Figure 5.
Marine-cyanobacterium-derived cycloheptadepsipeptides (104–123).

Figure 6.
Marine-cyanobacterium-derived thiazole-containing cyclodepsipeptides (124–142).

Figure 7.
Marine-cyanobacterium-derived other cyclic depsipeptides (143–161).

Figure 8.
Marine-sponge-derived depsipeptides (162–213).



Figure 9.
Marine-mollusk-derived depsipeptides (214–243).

Figure 10.
Marine-fungus-derived depsipeptides (244–259).

Figure 11.
Marine-bacterium-derived depsipeptides (260–282).

Figure 12.
Marine-alga-derived depsipeptides (283–288).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zeng, M.; Tao, J.; Xu, S.; Bai, X.; Zhang, H. Marine Organisms as a Prolific Source of Bioactive Depsipeptides. Mar. Drugs 2023, 21, 120. https://doi.org/10.3390/md21020120
AMA Style
Zeng M, Tao J, Xu S, Bai X, Zhang H. Marine Organisms as a Prolific Source of Bioactive Depsipeptides. Marine Drugs. 2023; 21(2):120. https://doi.org/10.3390/md21020120
Chicago/Turabian StyleZeng, Mingyuan, Jianyun Tao, Shuang Xu, Xuelian Bai, and Huawei Zhang. 2023. "Marine Organisms as a Prolific Source of Bioactive Depsipeptides" Marine Drugs 21, no. 2: 120. https://doi.org/10.3390/md21020120
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.