
Phytochemical Investigation of Three Cystoseira Species and Their Larvicidal Activity Supported with In Silico Studies

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. UPLC-ESI-MS Analysis for Characterization of Cystoseira myrica, C. trinodis, and C. tamariscifolia Methanol Fraction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min) | Compound Name | [M − H]− (m/z) | Molecular Formula | Relative Amount (%) | Ref. | ||
|---|---|---|---|---|---|---|---|---|
| C. myrica | C. trinodis | C. tamariscifolia | ||||||
| 1 | 1.29 | Compound 7 | 273.00 | C18H26O2 | 8.10 | - | 7.65 | [41] |
| 2 | 1.56 | Myricetin | 317.05 | C15H10O8 | - | - | 9.99 | [48] |
| 3 | 1.75 | (2E, l0E)-1-hydroxy-6,13-diketo-7- methylene-3,11,15-trimethylhexadeca-2, l0, l4-triene | 317.00 | C20H30O3 | - | - | 3.45 | [42] |
| 4 | 2.04 | Luteolin-7- glucoside | 447.05 | C21H20O11 | - | - | 2.36 | [49] |
| 5 | 3.67 | Bifuhalol hexacetate | 517.00 | C24H22O13 | 5.83 | - | - | [50] |
| 6 | 4.60 | Quercetin | 301.00 | C15H10O7 | 0.36 | 0.18 | - | [51] |
| 7 | 5.05 | Cyanidin-3-O- glucoside | 485.05 | C21H21O11+ | 48.27 | 10.06 | - | [52] |
| 8 | 5.58 | 7,8-Methylenedioxycoumarin | 189.10 | C10H6O4 | - | - | 13.87 | [53] |
| 9 | 7.32 | Oxocrinol | 223.10 | C14H24O2 | - | - | 6.50 | [43] |
| 10 | 7.86 | Cystoseirol monoacetate | 397.05 | C22H36O5 | 1.29 | 0.49 | 1.21 | [44] |
| 11 | 8.73 | Apigenin | 269.20 | C15H10O5 | - | - | 1.40 | [54] |
| 12 | 23.68 | α-Linolenic acid | 277.00 | C18H30O2 | - | - | 4.79 | [55] |
2.2. Larvicidal Bioassay
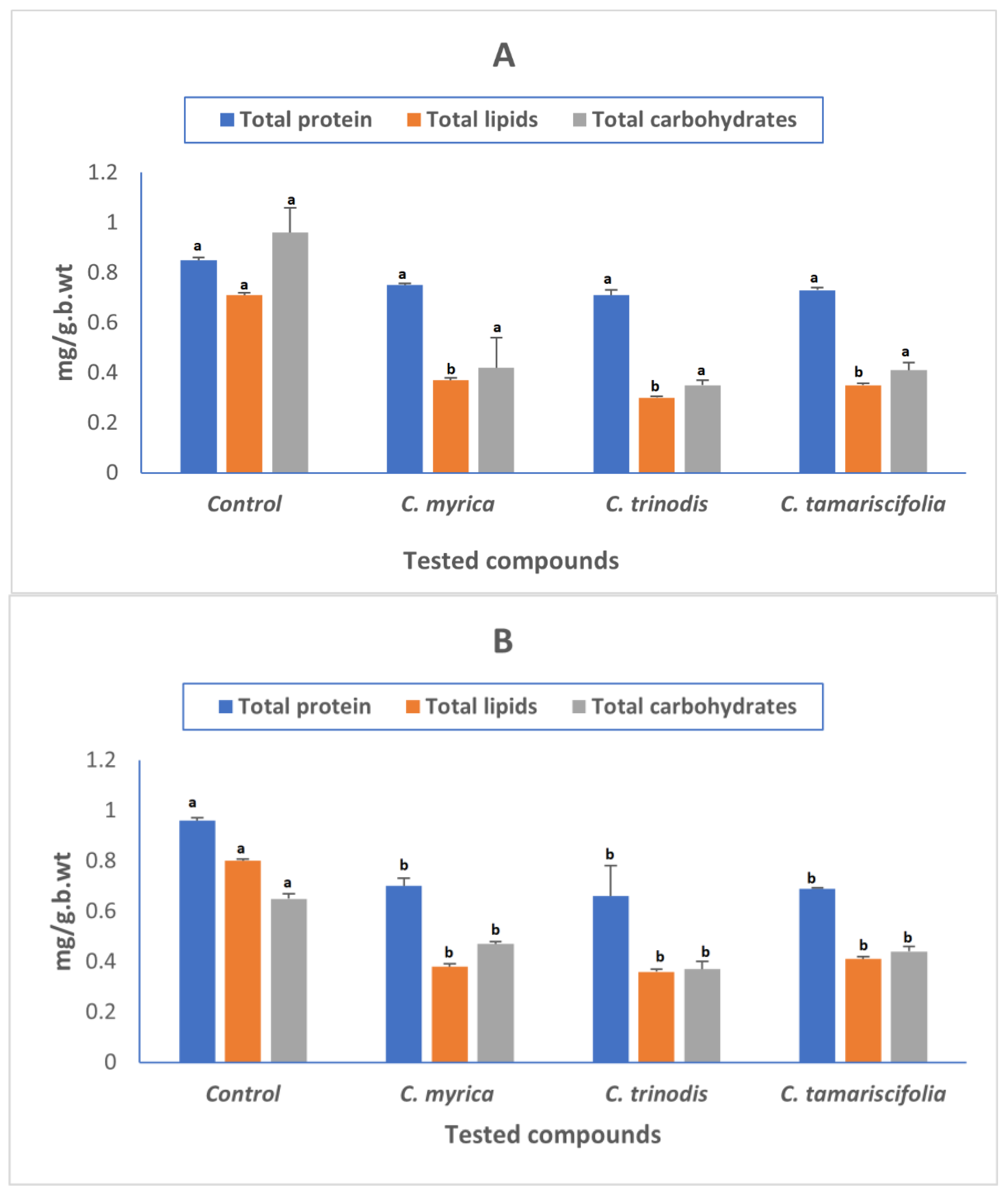
2.3. Biochemical Analysis
2.4. Determination of Total Proteins, Total Lipids, and Total Carbohydrates
2.5. In Silico Studies
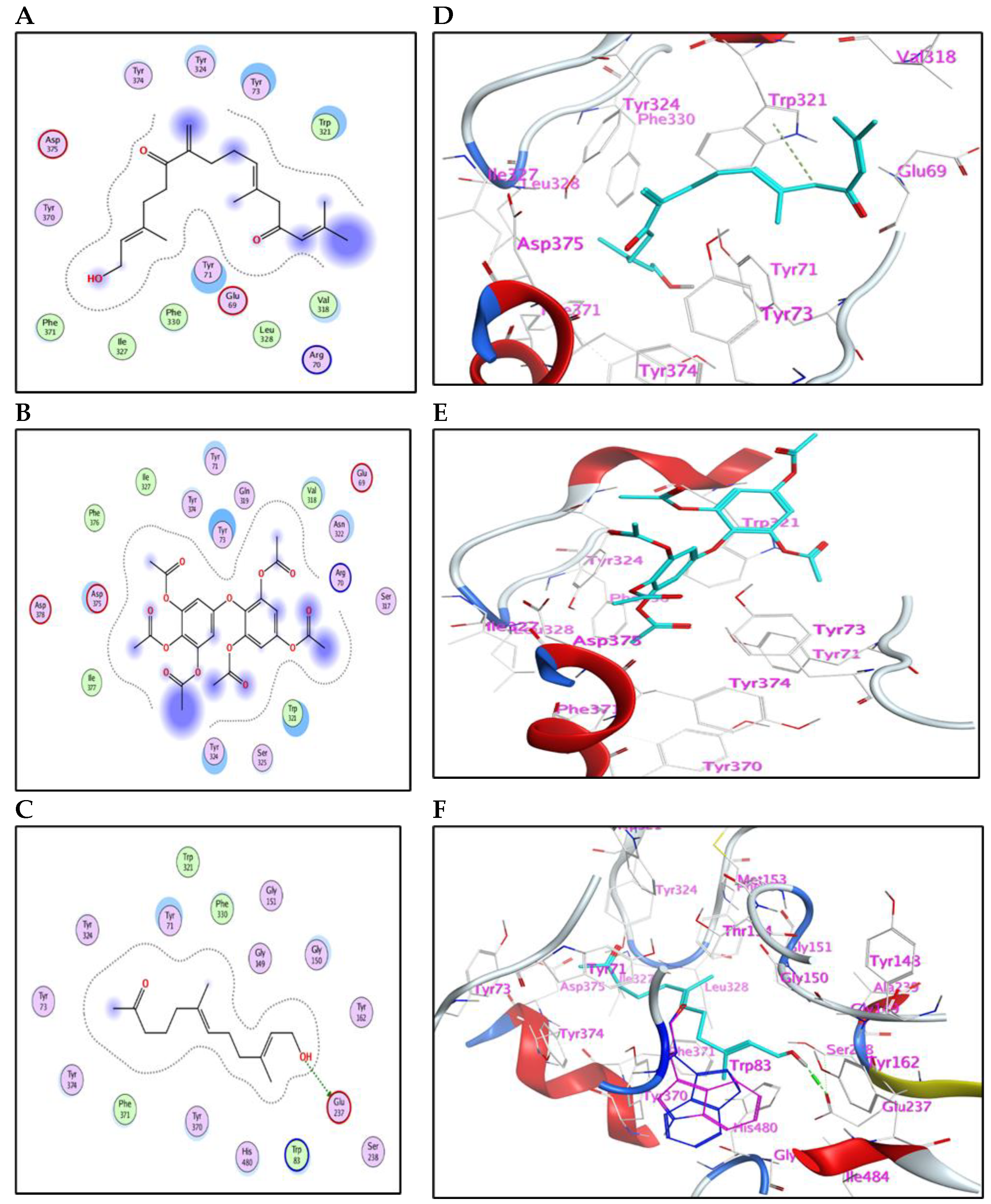
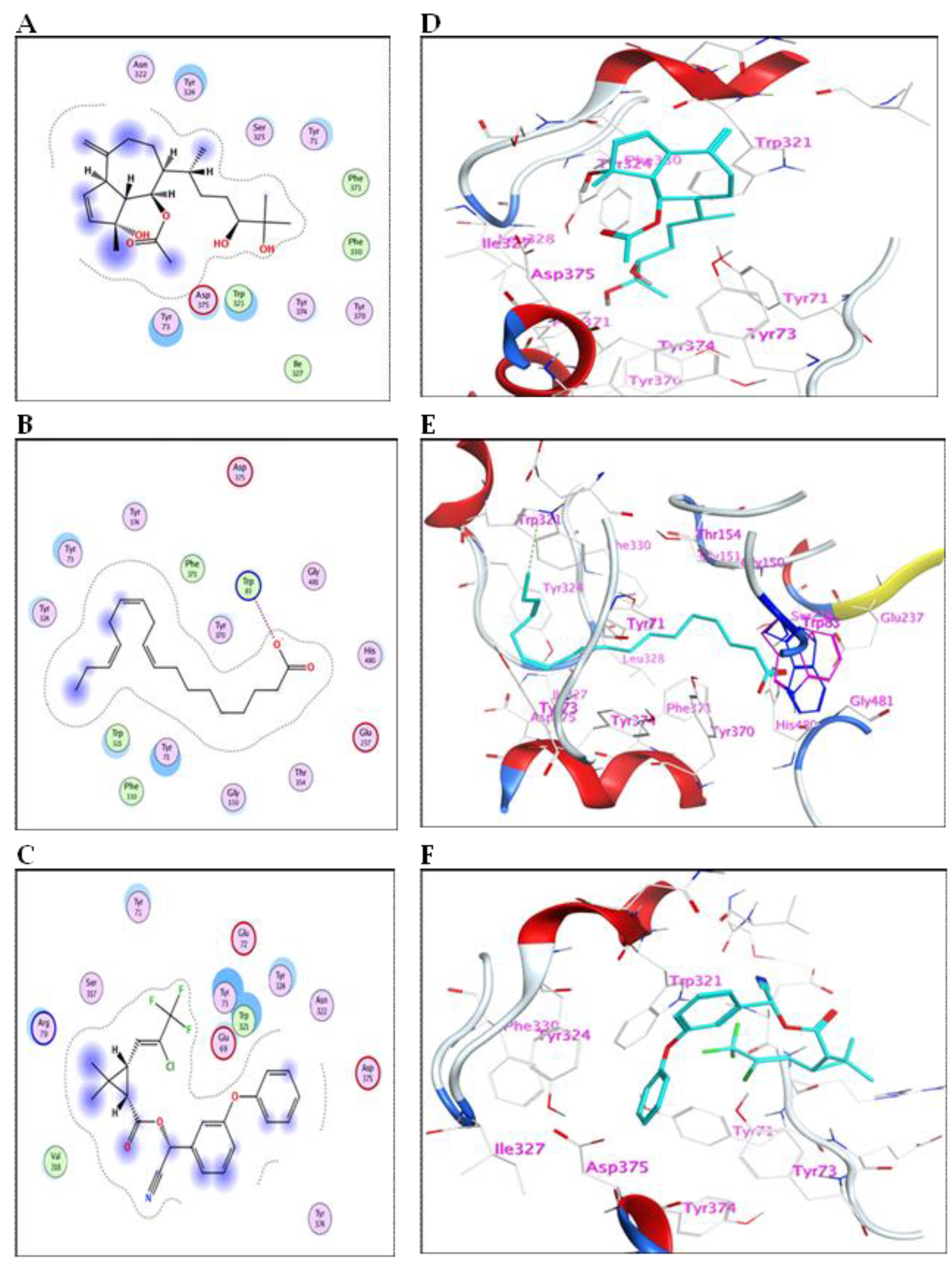
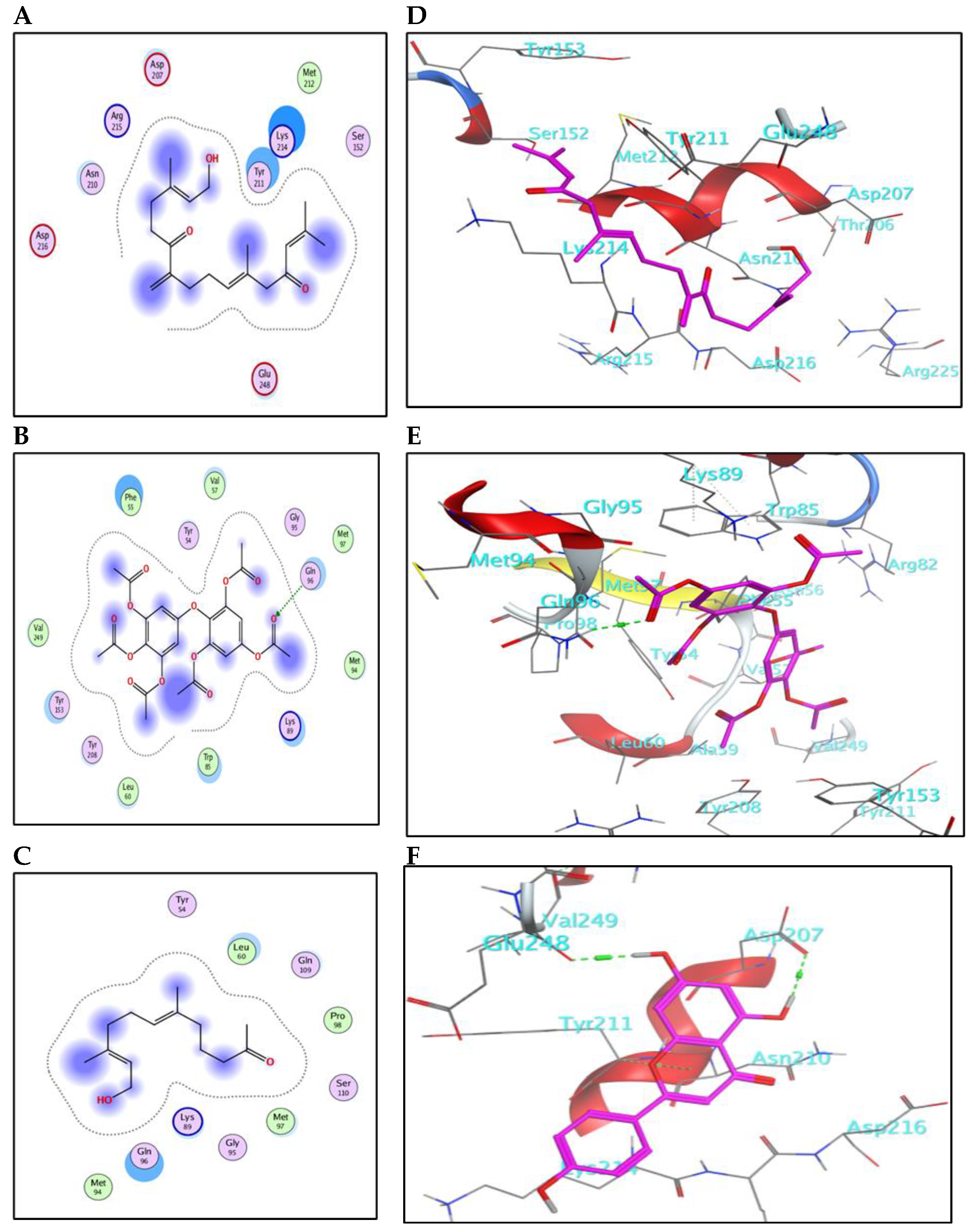
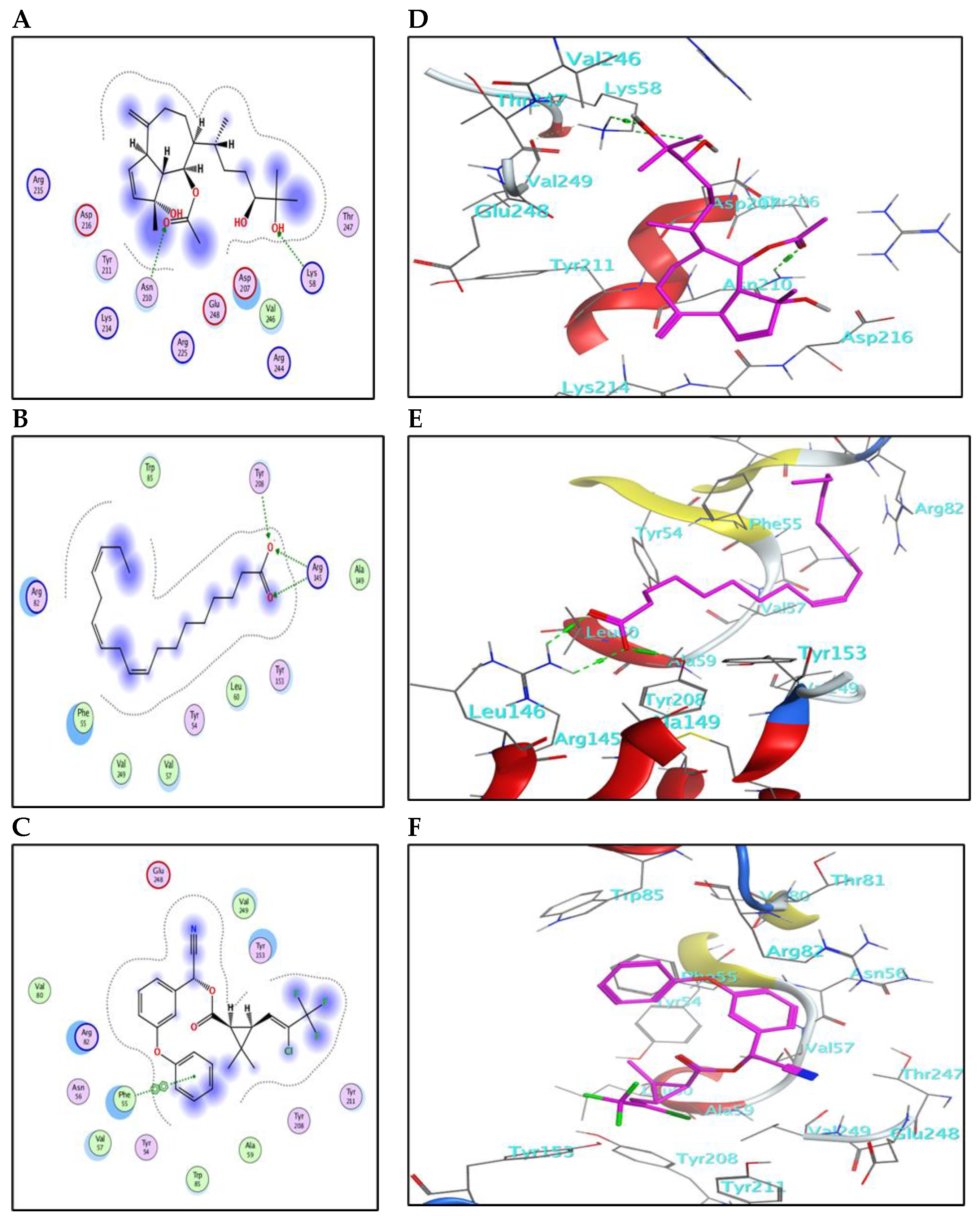
2.5.1. Binding Mode with AChE Enzyme
2.5.2. Binding Mode with GST Enzyme
3. Materials and Methods
3.1. Plant Material
3.2. Preparation of the Plant Extract
3.3. UPLC-ESI-MS Analysis
3.4. Insect Culture
3.5. Larvicidal Bioassay
3.6. Biochemical Assay
3.6.1. Estimation of Total Carbohydrates
3.6.2. Estimation of Total Lipids
3.6.3. Estimation of Total Proteins
3.7. Enzyme Activities
3.7.1. Glutathione S-transferase (GST)
3.7.2. Acetylcholine Esterase (AChE)
3.8. Statistical Analysis
3.9. In Silico Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kurane, I. The Effect of Global Warming on Infectious Diseases. Osong Public Health Res. Perspect. 2010, 1, 4–9. [Google Scholar] [CrossRef]
- Tibbetts, J. Driven to Extremes Health Effects of Climate Change. Environ. Health Perspect. 2007, 115, A196–A203. [Google Scholar] [CrossRef]
- Martinet, J.-P.; Ferté, H.; Failloux, A.-B.; Schaffner, F.; Depaquit, J. Mosquitoes of North-Western Europe as Potential Vectors of Arboviruses: A Review. Viruses 2019, 11, 1059. [Google Scholar] [CrossRef] [PubMed]
- Harbach, R.E. Culex pipiens: Species versus Species Complex Taxonomic History and Perspective. J. Am. Mosq. Control Assoc. 2012, 28, 10–23. [Google Scholar] [CrossRef]
- Pitasawat, B.; Champakaew, D.; Choochote, W.; Jitpakdi, A.; Chaithong, U.; Kanjanapothi, D.; Rattanachanpichai, E.; Tippawangkosol, P.; Riyong, D.; Tuetun, B.; et al. Aromatic Plant-Derived Essential Oil: An Alternative Larvicide for Mosquito Control. Fitoterapia 2007, 78, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, S.; Inbaneson, S.J.; Suganthi, P.; Gokulakrishnan, R.; Venkatesan, M. In Vitro Antiplasmodial Activity of Ethanolic Extracts of Seaweed Macroalgae against Plasmodium falciparum. Parasitol. Res. 2011, 108, 1411–1416. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Pavela, R.; Maggi, F.; Petrelli, R.; Nicoletti, M. Commentary: Making Green Pesticides Greener? The Potential of Plant Products for Nanosynthesis and Pest Control. J. Clust. Sci. 2017, 28, 3–10. [Google Scholar] [CrossRef]
- Paeporn, P.; Komalamisra, N.; Deesin, V.; Rongsriyam, Y.; Eshita, Y.; Thongrungkiat, S. Temephos Resistance in Two Forms of Aedes aegypti and Its Significance for the Resistance Mechanism. Southeast Asian J. Trop. Med. Public Health 2003, 34, 786–792. [Google Scholar]
- Abdel Haleem, D.R.; El Tablawy, N.H.; Ahmed Alkeridis, L.; Sayed, S.; Saad, A.M.; El-Saadony, M.T.; Farag, S.M. Screening and Evaluation of Different Algal Extracts and Prospects for Controlling the Disease Vector Mosquito Culex pipiens L. Saudi J. Biol. Sci. 2022, 29, 933–940. [Google Scholar] [CrossRef]
- Aly, S.H.; Elissawy, A.M.; Allam, A.E.; Farag, S.M.; Eldahshan, O.A.; Elshanawany, M.A.; Singab, A.N.B. New Quinolizidine Alkaloid and Insecticidal Activity of Sophora secundiflora and Sophora tomentosa against Culex pipiens (Diptera: Culicidae). Nat. Prod. Res. 2021, 36, 2722–2734. [Google Scholar] [CrossRef]
- Tantawy, A.H.; Farag, S.M.; Abdel-Haleem, D.R.; Mohamed, H.I. Facile Synthesis, Larvicidal Activity, Biological Effects, and Molecular Docking of Sulfonamide-Incorporating Quaternary Ammonium Iodides as Acetylcholinesterase Inhibitors against Culex pipiens L. Bioorg. Chem. 2022, 128, 106098. [Google Scholar] [CrossRef]
- Singab, A.N.B.; Mostafa, N.M.; Elkhawas, Y.A.; Al-Sayed, E.; Bishr, M.M.; Elissawy, A.M.; Elnaggar, M.S.; Fawzy, I.M.; Salama, O.M.; Tsai, Y.H.; et al. Cyclodepsipeptides: Isolation from Endophytic Fungi of Sarcophyton ehrenbergi and Verification of Their Larvicidal Activity via In-Vitro and In-Silico Studies. Mar. Drugs 2022, 20, 331. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, K. Biologically Active Compounds in Seaweed Extracts—The Prospects for the Application. Open Conf. Proc. J. 2012, 3, 20–28. [Google Scholar] [CrossRef]
- Dias, C.N.; Moraes, D.F.C. Essential Oils and Their Compounds as Aedes aegypti L. (Diptera: Culicidae) Larvicides: Review. Parasitol. Res. 2014, 113, 565–592. [Google Scholar] [CrossRef] [PubMed]
- El Sayed, K.A.; Dunbar, D.C.; Perry, T.L.; Wilkins, S.P.; Hamann, M.T.; Greenplate, J.T.; Wideman, M.A. Marine Natural Products as Prototype Insecticidal Agents. J. Agric. Food Chem. 1997, 45, 2735–2739. [Google Scholar] [CrossRef]
- Elbrense, H.; Gheda, S. Evaluation of the Insecticidal and Antifeedant Activities of Some Seaweed Extracts against the Egyptian Cotton Leaf Worm, Spodoptera littoralis, and the Lesser Grain Borer Rhyzopertha dominica. Egypt. J. Exp. Biol. 2021, 17, 1. [Google Scholar] [CrossRef]
- Fatima Manzelat, S.; Mohammed Mufarrah, A.; Ahmed Hasan, B.; Ali Hussain, N.; Shuqaiq, A.; Huraidha, A.; Qahma, A.; Birk, A. Macro Algae of the Red Sea from Jizan, Saudi Arabia. Phykos 2018, 48, 88–108. [Google Scholar] [CrossRef]
- Pati, M.P.; Sharma, S.D.; Nayak, L.; Panda, C.R. Uses of Seaweed and Its Application to Human Welfare: A Review. Int. J. Pharm. Pharm. Sci. 2016, 8, 12–20. [Google Scholar] [CrossRef]
- Stranska-Zachariasova, M.; Kurniatanty, I.; Gbelcova, H.; Jiru, M.; Rubert, J.; Nindhia, T.G.T.; D’Acunto, C.W.; Sumarsono, S.H.; Tan, M.I.; Hajslova, J.; et al. Bioprospecting of Turbinaria Macroalgae as a Potential Source of Health Protective Compounds. Chem. Biodivers. 2017, 14, e1600192. [Google Scholar] [CrossRef]
- Argandoña, V.; Del Pozo, T.; San-Martín, A.; Rovirosa, J. Insecticidal Activity of Plocamium cartilagineum Monoterpene. Boletín la Soc. Chil. Química 2000, 45, 371–376. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Mary Jeeva, Y. Eficacia Ninfcida y Ovicida de Una Alga Marina Sargassum tenerrimum (J. Agardh), Contra Dysdercus cingulatus (Fab.) (Pyrrhocoridae). Chil. J. Agric. Res. 2012, 72, 152–156. [Google Scholar] [CrossRef] [Green Version]
- Salvador-Neto, O.; Gomes, S.A.; Soares, A.R.; Da Silva Machado, F.L.; Samuels, R.I.; Fonseca, R.N.D.; Souza-Menezes, J.; Da Cunha Moraes, J.L.; Campos, E.; Mury, F.B.; et al. Larvicidal Potential of the Halogenated Sesquiterpene (+)-Obtusol, Isolated from the Alga Laurencia dendroidea J. Agardh (Ceramiales: Rhodomelaceae), against the Dengue Vector Mosquito Aedes aegypti (Linnaeus) (Diptera: Culicidae). Mar. Drugs 2016, 14, 20. [Google Scholar] [CrossRef]
- Yu, K.-X.; Jantan, I.; Ahmad, R.; Wong, C.-L. The Major Bioactive Components of Seaweeds and Their Mosquitocidal Potential. Parasitol. Res. 2014, 113, 3121–3141. [Google Scholar] [CrossRef] [PubMed]
- Abbassy, M. Insecticidal and Fungicidal Activity of Ulva lactuca Linnaeus (Chlorophyta) Extracts and Their Fractions. Annu. Res. Rev. Biol. 2014, 4, 2252–2262. [Google Scholar] [CrossRef]
- Manilal, A.; Selvin, J.; Thajuddin, N.; Sujith, S.; Panikkar, M.V.N.; Idhayadhulla, A.; Kumar, R.S. Biopotentials of Marine Alga, Lobophora variegate Collected from the South Indian Littoral. Thalassas 2012, 28, 47–54. [Google Scholar]
- Ishii, T.; Nagamine, T.; Nguyen, B.C.Q.; Tawata, S. Insecticidal and Repellent Activities of Laurinterol from the Okinawan Red Alga Laurencia nidifica. Rec. Nat. Prod. 2017, 11, 63–68. [Google Scholar]
- González-Castro, A.L.; Muñoz-Ochoa, M.; Hernández-Carmona, G.; López-Vivas, J.M. Evaluation of Seaweed Extracts for the Control of the Asian Citrus Psyllid diaphorina citri. J. Appl. Phycol. 2019, 31, 3815–3821. [Google Scholar] [CrossRef]
- Elbanna, S.; Hegazi, M. Screening of Some Seaweeds Species from South Sinai, Red Sea as Potential Bioinsecticides against Mosquito Larvae, Culex pipiens. Egypt. Acad. J. Biol. Sci. A Entomol. 2011, 4, 21–30. [Google Scholar] [CrossRef]
- Saber, A.A.; Hamed, S.M.; Abdel-Rahim, E.F.M.; Cantonati, M. Insecticidal Prospects of Algal and Cyanobacterial Extracts against the Cotton Leafworm Spodoptera littoralis. Vie Milieu 2018, 68, 199–212. [Google Scholar]
- Banaigs, B.; Francisco, C.; Gonzalez, E.; Fenical, W. Diterpenoid Metabolites from the Marine Alga Cystoseira elegans. Tetrahedron 1983, 39, 629–638. [Google Scholar] [CrossRef]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Delattre, C.; Molinié, R.; Petit, E.; Elboutachfaiti, R.; Nikolova, M.; Iliev, I.; Murdjeva, M.; et al. Structural Characterization and In Vivo Anti-Inflammatory Activity of Fucoidan from Cystoseira crinita (Desf.) Borry. Mar. Drugs 2022, 20, 714. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, G.; Fontana, M.; Asnaghi, V.; Chiantore, M.; Mirata, S.; Salis, A.; Damonte, G.; Scarfì, S. The Remarkable Antioxidant and Anti-Inflammatory Potential of the Extracts of the Brown Alga Cystoseira amentacea Var. Stricta. Mar. Drugs 2021, 19, 2. [Google Scholar] [CrossRef] [PubMed]
- Zbakh, H.; Zubía, E.; de los Reyes, C.; Calderón-Montaño, J.M.; López-Lázaro, M.; Motilva, V. Meroterpenoids from the Brown Alga Cystoseira usneoides as Potential Anti-Inflammatory and Lung Anticancer Agents. Mar. Drugs 2020, 18, 207. [Google Scholar] [CrossRef]
- Čagalj, M.; Skroza, D.; Razola-Díaz, M.D.C.; Verardo, V.; Bassi, D.; Frleta, R.; Mekinić, I.G.; Tabanelli, G.; Šimat, V. Variations in the Composition, Antioxidant and Antimicrobial Activities of Cystoseira compressa during Seasonal Growth. Mar. Drugs 2022, 20, 64. [Google Scholar] [CrossRef] [PubMed]
- Saber, F.R.; Aly, S.H.; Khallaf, M.A.; El-Nashar, H.A.S.; Fahmy, N.M.; El-Shazly, M.; Radha, R.; Prakash, S.; Kumar, M.; Taha, D.; et al. Hyphaene thebaica (Areceaeae) as a Promising Functional Food: Extraction, Analytical Techniques, Bioactivity, Food, and Industrial Applications. Food Anal. Methods 2022. [Google Scholar] [CrossRef]
- El-Nashar, H.A.S.; Eldehna, W.M.; Al-Rashood, S.T.; Alharbi, A.; Eskandrani, R.O.; Aly, S.H. GC/MS Analysis of Essential Oil and Enzyme Inhibitory Activities of Syzygium cumini (Pamposia) Grown in Docking Studies. Molecules 2021, 26, 6984. [Google Scholar] [CrossRef]
- El-Nashar, H.A.S.; Aly, S.H.; Ahmadi, A.; El-Shazly, M. The Impact of Polyphenolics in the Management of Breast Cancer: Mechanistic Aspects and Recent Patents. Recent Pat. Anticancer Drug Discov. 2021, 17, 358–379. [Google Scholar] [CrossRef]
- Aly, S.H.; Eldahshan, O.A.; Al-rashood, S.T.; Binjubair, F.A.; El Hassab, M.A.; Eldehna, W.M.; Acqua, S.D.; Zengin, G. Chemical Constituents, Antioxidant, and Enzyme Inhibitory Activities Supported by In-Silico Study of n-Hexane Extract and Essential Oil of Guava Leaves. Molecules 2022, 27, 8979. [Google Scholar] [CrossRef]
- Ads, E.N.; Hassan, S.I.; Rajendrasozhan, S.; Hetta, M.H.; Aly, S.H.; Ali, M.A. Isolation, Structure Elucidation and Antimicrobial Evaluation of Natural Pentacyclic Triterpenoids and Phytochemical Investigation of Different Fractions of Ziziphus spina-christi (L.) Stem Bark Using LCHRMS Analysis. Molecules 2022, 27, 1805. [Google Scholar] [CrossRef]
- Aly, S.H.; El-hassab, M.A.; Elhady, S.S.; Gad, H.A. Comparative Metabolic Study of Tamarindus Indica L.’s Various Organs Based on GC / MS Analysis, In Silico and In Vitro Anti-Inflammatory and Wound Healing Activities. Plants 2022, 12, 87. [Google Scholar] [CrossRef]
- Mokrini, R.; Ben Mesaoud, M.; Daoudi, M.; Hellio, C.; Maréchal, J.P.; El Hattab, M.; Ortalo-Magné, A.; Piovetti, L.; Culioli, G. Meroditerpenoids and Derivatives from the Brown Alga Cystoseira baccata and Their Antifouling Properties. J. Nat. Prod. 2008, 71, 1806–1811. [Google Scholar] [CrossRef]
- Amico, V.; Oriente, G.; Piattelli, M.; Ruberto, G.; Tringali, C. Novel Acyclic Diterpenes from the Brown Alga Cystoseira crinita. Phytochemistry 1981, 20, 1085–1088. [Google Scholar] [CrossRef]
- Fattorusso, E.; Magno, S.; Mayol, L.; Santacroce, C.; Sica, D.; Amico, V.; Oriente, G.; Piattelli, M.; Tringali, C. Oxocrinol and Crinitol, Novel Linear Terpenoids from the Brown Alga Cystoseira crinita. Tetrahedron Lett. 1976, 17, 937–940. [Google Scholar] [CrossRef]
- Ayyad, S.E.N.; Abdel-Halim, O.B.; Shier, W.T.; Hoye, T.R. Cytotoxic Hydroazulene Diterpenes from the Brown Alga Cystoseira myrica. Zeitschrift Fur Naturforsch.—Sect. C J. Biosci. 2003, 58, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Oliyaei, N.; Moosavi-Nasab, M. Ultrasound-Assisted Extraction of Fucoxanthin from Sargassum angustifolium and Cystoseira indica Brown Algae. J. Food Process. Preserv. 2021, 45, e15929. [Google Scholar] [CrossRef]
- Bharadwaj, S.; Sheeja, L.; Lakshmi, D.; Parveen, S.K. 1 H NMR Analysis and Bioautography Screening of Methanol Extract of Sargassum wightii by Chromatographic Separation. Res. J. Pharm. Technol. 2017, 10, 473. [Google Scholar] [CrossRef]
- El-Khateeb, A.; Hamed, E.; Ibrahim, F.; Hamed, S. Eco-Friendly Synthesis of Selenium and Zinc Nanoparticles with Biocompatible Sargassum latifolium Algae Extract in Preservation of Edible Oils. J. Food Dairy Sci. 2019, 10, 141–146. [Google Scholar] [CrossRef]
- Sun, J.; Liang, F.; Bin, Y.; Li, P.; Duan, C. Screening Non-Colored Phenolics in Red Wines Using Liquid Chromatography/Ultraviolet and Mass Spectrometry/Mass Spectrometry Libraries. Molecules 2007, 12, 679–693. [Google Scholar] [CrossRef]
- Savarese, M.; De Marco, E.; Sacchi, R. Food Chemistry Characterization of Phenolic Extracts from Olives (Olea europaea Cv. Pisciottana) by Electrospray Ionization Mass Spectrometry. Food Chem. 2007, 105, 761–770. [Google Scholar] [CrossRef]
- Glombitza, K.W.; Rosener, H.U.; Müller, D. Bifuhalol und Diphlorethol aus Cystoseira tamariscifolia. Phytochemistry 1975, 14, 1115–1116. [Google Scholar] [CrossRef]
- Faheem, S.A.; Saeed, N.M.; El-Naga, R.N.; Ayoub, I.M.; Azab, S.S. Hepatoprotective Effect of Cranberry nutraceutical Extract in Non-Alcoholic Fatty Liver Model in Rats: Impact on Insulin Resistance and Nrf-2 Expression. Front. Pharmacol. 2020, 11, 218. [Google Scholar] [CrossRef]
- Abdelghffar, E.A.; El-Nashar, H.A.S.; Al-Mohammadi, A.G.A.; Eldahshan, O.A. Orange Fruit (Citrus sinensis) Peel Extract Attenuates Chemotherapy-Induced Toxicity in Male Rats. Food Funct. 2021, 12, 9443–9455. [Google Scholar] [CrossRef]
- Koul, S.K.; Dhar, K.L.; Thakur, R.S. A New Coumarin Glucoside from Prangos pabularia. Phytochemistry 1979, 18, 1762–1763. [Google Scholar] [CrossRef]
- Emam, M.; Abdel-Haleem, D.R.; Salem, M.M.; Abdel-Hafez, L.J.M.; Latif, R.R.A.; Farag, S.M.; Sobeh, M.; Raey, M.A. El Phytochemical Profiling of Lavandula coronopifolia poir. Aerial Parts Extract and Its Larvicidal, Antibacterial, and Antibiofilm Activity against Pseudomonas aeruginosa. Molecules 2021, 26, 1710. [Google Scholar] [CrossRef] [PubMed]
- Segarra, G.; Jáuregui, O.; Casanova, E.; Trillas, I. Simultaneous Quantitative LC-ESI-MS/MS Analyses of Salicylic Acid and Jasmonic Acid in Crude Extracts of Cucumis sativus under Biotic Stress. Phytochemistry 2006, 67, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Minicante, S.A.; Carlin, S.; Stocco, M.; Sfriso, A.; Capelli, G.; Montarsi, F. Preliminary Results on the Efficacy of Macroalgal Extracts Against Larvae of Aedes albopictus. J. Am. Mosq. Control Assoc. 2017, 33, 352–354. [Google Scholar] [CrossRef] [PubMed]
- Fouda, A.; Eid, A.M.; Abdel-Rahman, M.A.; EL-Belely, E.F.; Awad, M.A.; Hassan, S.E.D.; AL-Faifi, Z.E.; Hamza, M.F. Enhanced Antimicrobial, Cytotoxicity, Larvicidal, and Repellence Activities of Brown Algae, Cystoseira crinita-Mediated Green Synthesis of Magnesium Oxide Nanoparticles. Front. Bioeng. Biotechnol. 2022, 10, 849921. [Google Scholar] [CrossRef]
- Waliwitiya, R.; Nicholson, R.A.; Kennedy, C.J.; Lowenberger, C.A. The Synergistic Effects of Insecticidal Essential Oils and Piperonyl Butoxide on Biotransformational Enzyme Activities in Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Boily, M.; Sarrasin, B.; Deblois, C.; Aras, P.; Chagnon, M. Acetylcholinesterase in Honey Bees (Apis mellifera) Exposed to Neonicotinoids, Atrazine and Glyphosate: Laboratory and Field Experiments. Environ. Sci. Pollut. Res. Int. 2013, 20, 5603–5614. [Google Scholar] [CrossRef]
- Rajashekar, Y.; Raghavendra, A.; Bakthavatsalam, N. Acetylcholinesterase Inhibition by Biofumigant (Coumaran) from Leaves of Lantana camara in Stored Grain and Household Insect Pests. Biomed Res. Int. 2014, 2014, 187019. [Google Scholar] [CrossRef]
- Huang, C.S.; Lii, C.K.; Lin, A.H.; Yeh, Y.W.; Yao, H.T.; Li, C.C.; Wang, T.S.; Chen, H.W. Protection by Chrysin, Apigenin, and Luteolin against Oxidative Stress Is Mediated by the Nrf2-Dependent up-Regulation of Heme Oxygenase 1 and Glutamate Cysteine Ligase in Rat Primary Hepatocytes. Arch. Toxicol. 2013, 87, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Farag, S.M.; Hussein, M.A.; Hafez, S.E.; Khaled, A.S.; Kamel, O.M.; Zyaan, O.H. Larvicidal, Biological, and Histopathological Alterations Induced by Pomegranate Peel Extract, Punica granatum against Culex pipiens L. (Diptera: Culicidae). Egypt. J. Aquat. Biol. Fish. 2021, 25, 139–161. [Google Scholar] [CrossRef]
- Ibraheem, I.B.M.; Alharbi, R.M.; Abdel-Raouf, N.; Al-Enazi, N.M. Contributions to the Study of the Marine Algae Inhabiting Umluj Seashore, Red Sea. Beni-Suef Univ. J. Basic Appl. Sci. 2014, 3, 278–285. [Google Scholar] [CrossRef]
- Aly, S.H.; Elissawy, A.M.; Fayez, A.M.; Eldahshan, O.A.; Elshanawany, M.A.; Singab, A.N.B. Neuroprotective Effects of Sophora secundiflora, Sophora tomentosa Leaves and Formononetin on Scopolamine-Induced Dementia. Nat. Prod. Res. 2020, 35, 5848–5852. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; He, H.; Li, P.; Zhu, J.; Xiong, M. Identification and Quantification of Atractylenolide I and Atractylenolide III in Rhizoma Atractylodes Macrocephala by Liquid Chromatography-Ion Trap Mass Spectrometry. Biomed. Chromatogr. 2013, 27, 699–707. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Huang, Y.; Wen, X.; Wu, Y.; Zhao, Y.; Ni, Y. Extraction, Purification, and Hydrolysis Behavior of Apigenin-7-O-Glucoside from Chrysanthemum morifolium Tea. Molecules 2018, 23, 2933. [Google Scholar] [CrossRef]
- Elhawary, E.A.; Mostafa, N.M.; Shehata, A.Z.I.; Labib, R.M.; Singab, A.N.B. Comparative Study of Selected Rosa Varieties’ Metabolites through UPLC-ESI-MS/MS, Chemometrics and Investigation of Their Insecticidal Activity against Culex pipiens L. Jordan J. Pharm. Sci. 2021, 14, 417–433. [Google Scholar]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. 1925. J. Am. Mosq. Control Assoc. 1987, 3, 302–303. [Google Scholar]
- Saad, A.M.; El-Saadony, M.T.; Mohamed, A.S.; Ahmed, A.I.; Sitohy, M.Z. Impact of Cucumber Pomace Fortification on the Nutritional, Sensorial and Technological Quality of Soft Wheat Flour-Based Noodles. Int. J. Food Sci. Technol. 2021, 56, 3255–3268. [Google Scholar] [CrossRef]
- Knight, J.A.; Anderson, S.; Rawle, J.M. Chemical Basis of the Sulfo-Phospho-Vanillin Reaction for Estimating Total Serum Lipids. Clin. Chem. 1972, 18, 199–202. [Google Scholar] [CrossRef]
- Wojciechowska, M.; Stepnowski, P.; Gołębiowski, M. Identification and Quantitative Analysis of Lipids and Other Organic Compounds Contained in Eggs of Colorado Potato Beetle (Leptinotarsa decemlineata). J. Plant Dis. Prot. 2019, 126, 379–384. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kao, C.-W.; Bakshi, M.; Sherameti, I.; Dong, S.; Reichelt, M.; Oelmüller, R.; Yeh, K.-W. A Chinese Cabbage (Brassica campetris subsp. Chinensis) τ-Type Glutathione-S-Transferase Stimulates Arabidopsis Development and Primes against Abiotic and Biotic Stress. Plant Mol. Biol. 2016, 92, 643–659. [Google Scholar] [CrossRef]
- Simpson, D.R.; Bull, D.L.; Lindquist, D.A. A Semimicrotechnique for the Estimation of Cholinesterase Activity in Boll Weevils1. Ann. Entomol. Soc. Am. 1964, 57, 367–371. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1971. [Google Scholar]
- Nachon, F.; Rosenberry, T.L.; Silman, I.; Sussman, J.L. A Second Look at the Crystal Structures of Drosophila melanogaster Acetylcholinesterase in Complex with Tacrine Derivatives Provides Insights Concerning Catalytic Intermediates and the Design of Specific Insecticides. Molecules 2020, 25, 1198. [Google Scholar] [CrossRef]
- Agianian, B.; Tucker, P.A.; Schouten, A.; Leonard, K.; Bullard, B.; Gros, P. Structure of a Drosophila Sigma Class Glutathione S-Transferase Reveals a Novel Active Site Topography Suited for Lipid Peroxidation Products. J. Mol. Biol. 2003, 326, 151–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, H.; Kidachi, Y.; Kamiie, K.; Noshita, T.; Umetsu, H. Structural Insight into the Ligand-Receptor Interaction between Glycyrrhetinic Acid (GA) and the High-Mobility Group Protein B1 (HMGB1)-DNA Complex. Bioinformation 2012, 8, 1147–1153. [Google Scholar] [CrossRef]
- Aziz, M.; Ejaz, S.A.; Tamam, N.; Siddique, F.; Riaz, N.; Qais, F.A.; Chtita, S.; Iqbal, J. Identification of Potent Inhibitors of NEK7 Protein Using a Comprehensive Computational Approach. Sci. Rep. 2022, 12, 6404. [Google Scholar] [CrossRef]
- Morris, G.M.; Lim-Wilby, M. Molecular Docking. In Methods in Molecular Biology; Kukol, A., Ed.; Humana Press: Totowa, NJ, USA, 2008; Volume 443, pp. 365–382. ISBN 9781588298645. [Google Scholar]






| Conc. (ppm) | Percentage of Mortalities % | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| C. myrica | C. trinodis | C. tamariscifolia | |||||||
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |
| 0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| 50 | 18.66 | 26.66 | 34.66 | 10.66 | 18.33 | 22.66 | 5.3 | 13.33 | 21.33 |
| 100 | 44 | 52 | 60 | 24 | 36 | 41.33 | 21.33 | 32 | 44 |
| 150 | 60 | 68 | 72 | 52 | 60 | 69.33 | 60 | 68 | 76 |
| 200 | 86.66 | 80 | 92 | 74.66 | 82.66 | 88 | 76 | 82.66 | 88 |
| 250 | 90.66 | 94.66 | 98.66 | 81.33 | 90.66 | 92 | 90.66 | 93.33 | 97.33 |
| LC25 | 64.51 (54.4–73.5) | 51.99 (41.5–61.3) | 44.74 (17.9–51.3) | 95.02 (85–104) | 75.17 (65.7–83.6) | 60.27 (51–68.5) | 86.96 (75.6–97) | 68.65 (40.3–82) | 60.33 (50.5–69) |
| LC50 | 105.06 (94.8–115) | 91.09 (80–101.5) | 75.63 (42.8–94.7) | 135.08 (125.6–145) | 113.42 (104–122.7) | 94.66 (85.3–103.7) | 138.71 (127.1–151) | 111.51 (80.8–141) | 98.15 (88.1–107.8) |
| LC95 | 345.06 (292–432.7) | 357.67 (294.9–468.6) | 272.09 (234.8–638.2) | 318.54 (281–375.3) | 309.33 (269.7–370.5) | 284.62 (245.6–346.3) | 433.21 (360.6–558) | 364.01 (306.7–758.8) | 321.55 (273.5–399.9) |
| * Slope ± SE | 3.19 ± 0.28 | 2.77 ± 0.26 | 2.96 ± 0.27 | 4.41 ± 0.36 | 3.78 ± 0.30 | 3.44 ± 0.29 | 3.33 ± 0.29 | 3.20 ± 0.28 | 3.19 ± 0.28 |
| * X2 | 7.38 | 5.89 | 10.62 | 5.46 | 7.43 | 5.57 | 6.16 | 8.43 | 7.69 |
| p-value | 0.06 | 0.11 | 0.0139 | 0.14 | 0.06 | 0.13 | 0.10 | 0.04 | 0.05 |
| Sample Tested | GST (UX103/g.b.wt) | AChE (ug AchBr/min/g.b.wt) | ||
|---|---|---|---|---|
| 24 h | 48 h | 24 h | 48 h | |
| Untreated | 0.86 ± 0.03 a | 0.78 ± 0.06 a | 7.23 ± 0.26 a | 7.60 ± 0.37 a |
| C. myrica | 1.69 ± 0.005 b | 1.52 ± 0.08 b | 5.69 ± 0.4 b | 6.23 ± 0.33 b |
| C. trinodis | 1.70 ± 0.01 b | 1.54 ± 0.06 b | 6.16 ± 0.8 b | 6.26 ± 0.34 b |
| C. tamariscifolia | 1.72 ± 0.01 b | 1.56 ± 0.05 b | 6.26 ± 0.6 a | 6.36 ± 0.27 b |
| Code | Compound Name | AChE Interaction | GST Interaction | ||
|---|---|---|---|---|---|
| BE a (∆G) | RMSD b Refine | BE (∆G) | RMSD b Refine | ||
| 1 | Compound 7 | −5.02 | 1.34 | −5.22 | 0.96 |
| 2 | Myricetin | −6.17 | 1.41 | −5.67 | 1.78 |
| 3 | (2E, l0E)-1-hydroxy-6,13-diketo-7-methylene-3,11,15-trimethyl- hexadeca-2,l0,l4-triene | −7.18 | 1.97 | −5.75 | 1.22 |
| 4 | Luteolin-7-O-glucoside | −6.98 | 1.61 | −6.36 | 1.18 |
| 5 | Bifuhalol hexacetate | −7.26 | 1.84 | −6.51 | 1.66 |
| 7 | Cyanidin-3-O-glucoside | −6.80 | 1.17 | −6.21 | 1.32 |
| 8 | 7,8-Methylenedioxycoumarin | −4.98 | 1.19 | −4.42 | 1.72 |
| 9 | Oxocrinol | −7.28 | 1.77 | −5.23 | 1.42 |
| 10 | Cystoseirol monoacetate | −6.86 | 1.84 | −6.06 | 1.65 |
| 11 | Apigenin | −5.59 | 0.51 | −5.18 | 1.29 |
| 12 | α-Linolenic acid | −7.97 | 1.89 | −5.71 | 1.79 |
| - | λ-Cyhalothrin | −6.57 | 1.16 | −5.86 | 1.31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aly, S.H.; Elissawy, A.M.; Salah, D.; Alfuhaid, N.A.; Zyaan, O.H.; Mohamed, H.I.; Singab, A.N.B.; Farag, S.M. Phytochemical Investigation of Three Cystoseira Species and Their Larvicidal Activity Supported with In Silico Studies. Mar. Drugs 2023, 21, 117. https://doi.org/10.3390/md21020117
Aly SH, Elissawy AM, Salah D, Alfuhaid NA, Zyaan OH, Mohamed HI, Singab ANB, Farag SM. Phytochemical Investigation of Three Cystoseira Species and Their Larvicidal Activity Supported with In Silico Studies. Marine Drugs. 2023; 21(2):117. https://doi.org/10.3390/md21020117
Chicago/Turabian StyleAly, Shaza H., Ahmed M. Elissawy, Dina Salah, Nawal Abdulaziz Alfuhaid, Ola H. Zyaan, Hany I. Mohamed, Abdel Nasser B. Singab, and Shaimaa M. Farag. 2023. "Phytochemical Investigation of Three Cystoseira Species and Their Larvicidal Activity Supported with In Silico Studies" Marine Drugs 21, no. 2: 117. https://doi.org/10.3390/md21020117