
Marine-Derived Actinomycetes: Biodegradation of Plastics and Formation of PHA Bioplastics—A Circular Bioeconomy Approach

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Evaluation of Plastics Biodegradation Potential by Actinomycetes Using Plastics Emulsified Media Assays
2.2. Evaluation of Plastics Biodegradation by Actinomycetes Using Thin Plastic Films Assays
2.2.1. Thin Plastic Films’ Weight Loss
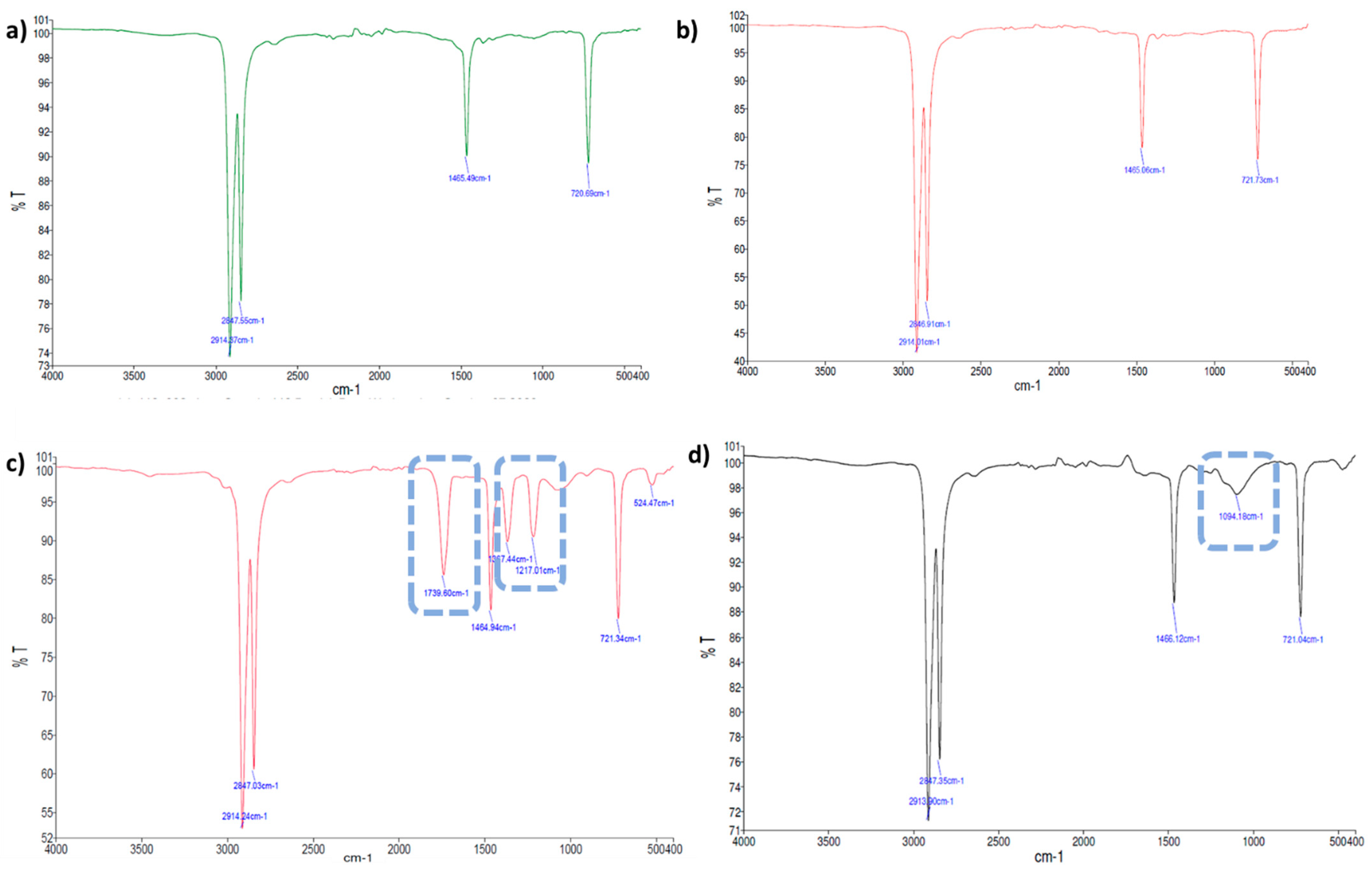
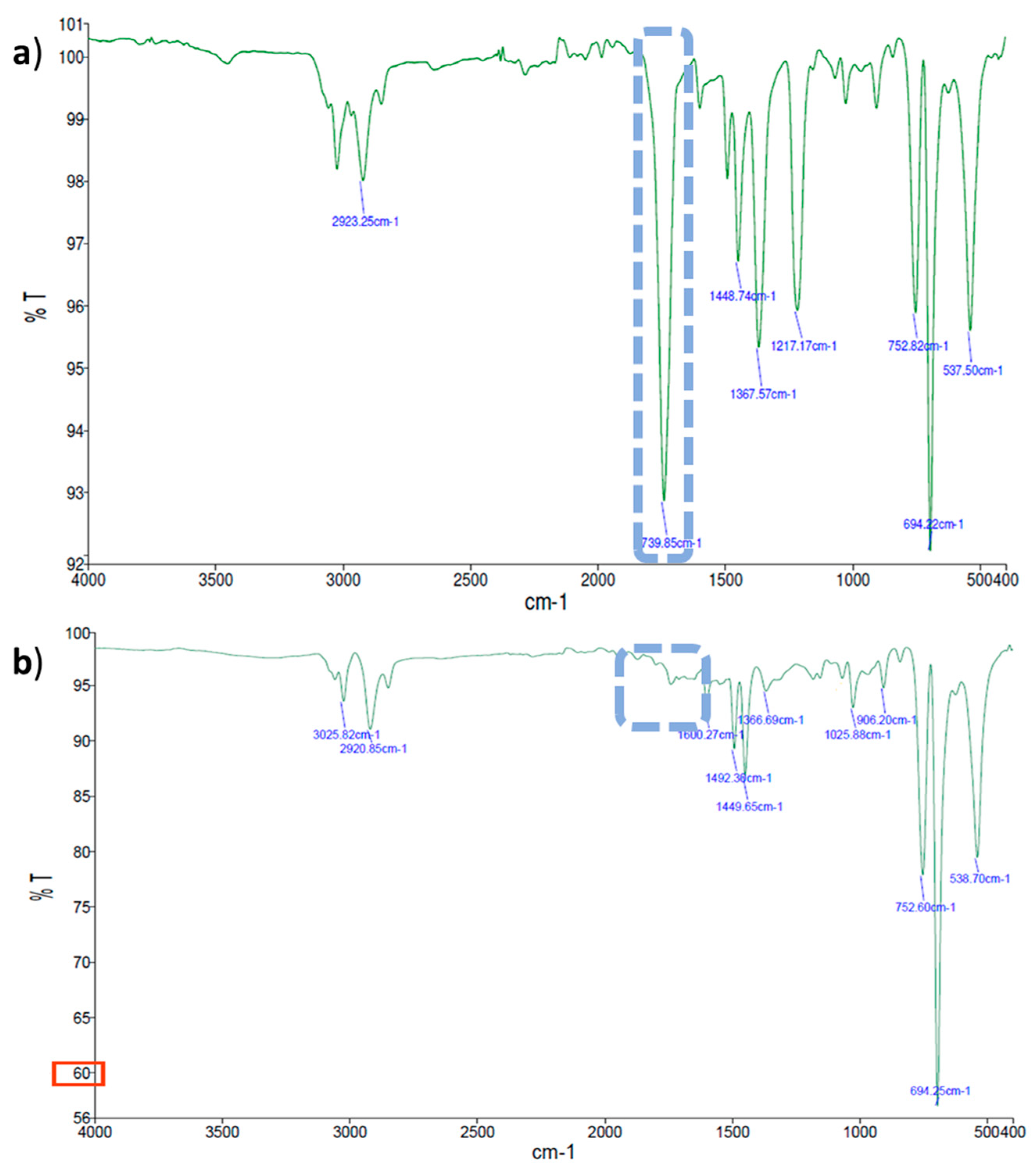
2.2.2. Evaluation of Thin Plastic Films Chemical Changes by FTIR-ATR Spectroscopy
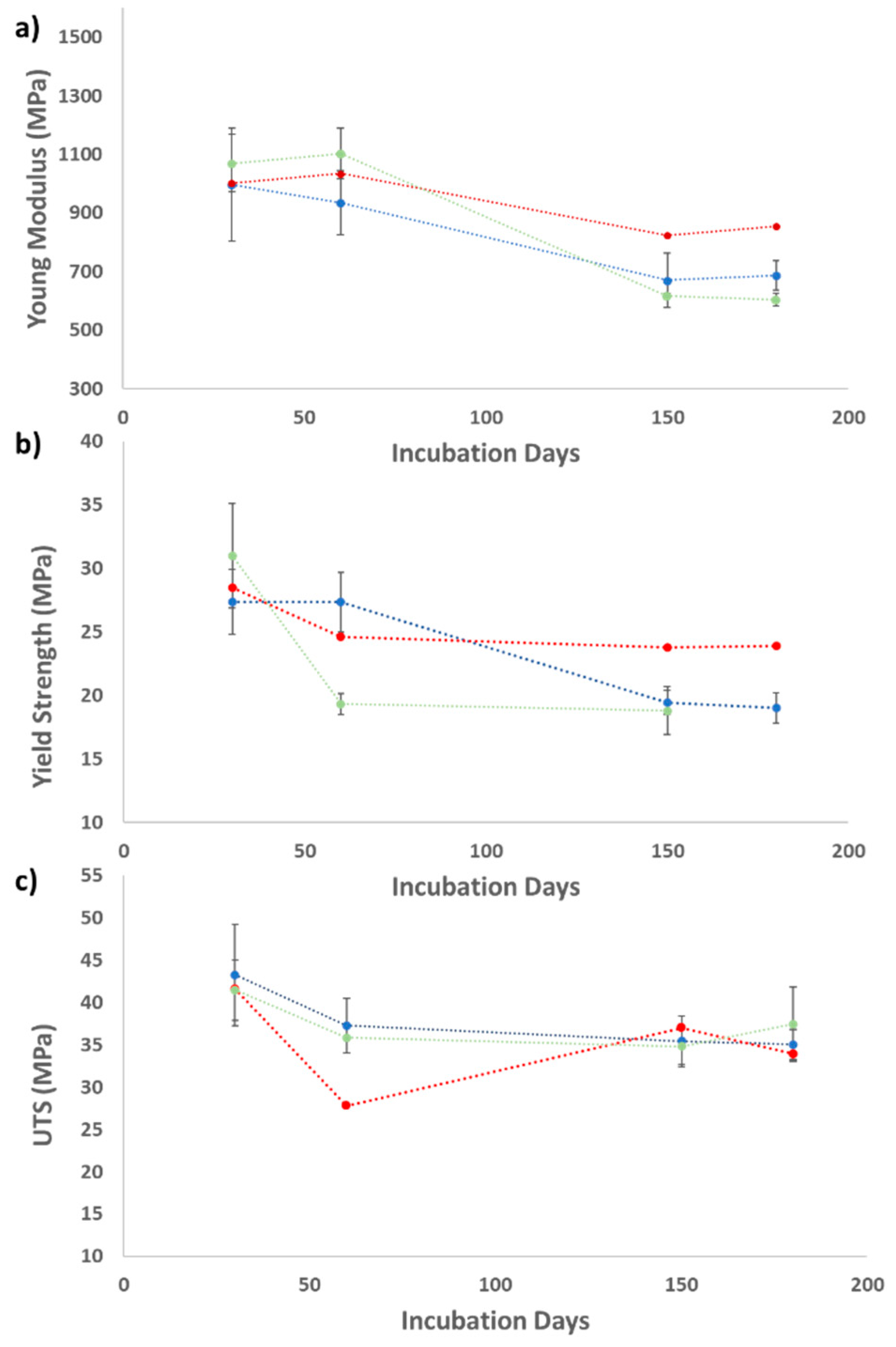
2.2.3. Mechanical Properties of the Thin Plastic Films
2.3. PHA Inclusions in the Actinomycetes Cells
2.4. LDPE Biodegradation and PHA Production by Actinomycetes
2.5. PS Biodegradation and PHA Production by Actinomycetes
2.6. PLA Biodegradation and PHA Production by Actinomycetes
3. Materials and Methods
3.1. Preparation of Polymer-Emulsified Media
3.2. Actinomycetes for Plastics Biodegradation Screening
3.3. Clear Zone Test for Plastics Biodegradation
3.4. Production and Preparation of Polymeric Thin Plastic Films
3.5. Thin Plastic Films Biodegradation Assays
3.6. Monitoring of Thin Plastic Films Biodegradation
3.6.1. Plastic Films Weight Loss
3.6.2. Fourier Transform Infrared Spectra of Plastic Films
3.6.3. Tensile Strength Tests
3.7. Evaluation of Bioplastic Production by Detection of PHA Inclusions in Actinomycete Cells
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hopewell, J.; Dvorak, R.; Kosior, E. Plastics recycling: Challenges and opportunities. Philos. Trans. R. Soc. 2009, 364, 2115–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savelli, H.; Beunen, R.; Kalz, M.; Ragas, A.; Belleghem, F.V. Solutions for global marine litter pollution. Curr. Opin. Environ. Sustain. 2017, 28, 90–99. [Google Scholar] [CrossRef] [Green Version]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Association of Plastics Manufacturers. P.E. Plastics—The Facts 2019. An Analysis of European Plastics Production, Demand and Waste Data. Available online: https://www.plasticseurope.org/application/files/9715/7129/9584/FINAL_web_version_Plastics_the_facts2019_14102019.pdf (accessed on 7 April 2022).
- Eriksen, M.; Lebreton, L.C.M.; Carson, H.S.; Thiel, M.; Moore, C.J.; Borerro, J.C.; Galgani, F.; Ryan, P.G. Plastic pollution in the World’s oceans: More than 5 trillion plastic pieces weighing over 250,000 tons afloat at sea. PLoS ONE 2014, 9, e111913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GESAMP. Sources, Fate and Effects of Microplastics in the Marine Environment: A Global Assessment; Kershaw, P.J., Ed.; International Maritime Organization: London, UK, 2015.
- Restrepo-Flórez, J.; Bassi, A.; Thompson, M.R. Microbial degradation and deterioration of polyethylene—A review. Int. Biodeterior. Biodegrad. 2014, 88, 83–90. [Google Scholar] [CrossRef]
- Andrady, A.L.; Hamid, S.H.; Hu, X.; Torikai, A. Effects of increased solar ultraviolet radiation on terrestrial ecosystems. J. Photochem. Photobiol. 1998, 46, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.; Belchior, A.; da Silva, V.D.; Rotter, A.; Petrovski, Z.; Almeida, P.L.; Lourenço, N.D.; Gaudêncio, S.P. Marine environmental plastic pollution: Mitigation by microorganism degradation and recycling valorization. Front. Mar. Sci. 2020, 7, 567126. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, C.G. Biodegradation of micro-polyethylene particles by bacterial colonization of a mixed microbial consortium isolated from a land fill site. Chemosphere 2019, 222, 527–533. [Google Scholar] [CrossRef]
- Raddadi, N.; Fava, F. Biodegradation of oil-based plastics in the environment: Existing knowledge and needs of research and innovation. Sci. Total Environ. 2019, 679, 148–158. [Google Scholar] [CrossRef]
- Chandra, R.; Rustgi, R. Biodegradable Polymers. Prog. Polym. Sci. 1998, 23, 1273–1335. [Google Scholar] [CrossRef]
- Amobonye, A.; Bhagwat, P.; Singh, S.; Pillai, S. Plastic biodegradation: Frontline microbes and their enzymes. Sci. Total Environ. 2020, 759, 143536. [Google Scholar] [CrossRef] [PubMed]
- Sivan, A.; Szanto, M.; Pavlov, V. Biofilm development of the polyethylene-degrading bacterium Rhodococcus ruber. Appl. Microb. Cell Physiol. 2006, 72, 346–352. [Google Scholar] [CrossRef]
- Farzi, A.; Dehnad, A.; Fotouhi, A.F. Biodegradation of polyethylene terephthalate waste using Streptomyces species and kinetic modeling of the process. Biocatal. Agric. Biotechnol. 2019, 17, 25–31. [Google Scholar] [CrossRef]
- Bonhomme, S.; Cuer, A.; Delort, A.; Lemaire, J.; Sancelme, M.; Scott, G. Environmental biodegradation of polyethylene. Polym. Degrad. Stab. 2003, 81, 441–452. [Google Scholar] [CrossRef]
- Usha, R.; Muthusamy, P.; Palaniswamy, M. Screening of polyethylene degrading microorganisms from garbage soil. Libyan Agric. Res. Cent. J. Int. 2011, 2, 200–204. [Google Scholar]
- Auta, H.S.; Emenike, C.U.; Jayanthi, B.; Fauziah, S.H. Growth kinetics and biodeterioration of polypropylene microplastics by Bacillus sp. and Rhodococcus sp. isolated from mangrove sediment. Mar. Pollut. Bull. 2018, 127, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Castro-Aguirre, E.; Iñiguez-Franco, F.; Samsudin, H.; Fang, X.; Auras, R. Poly(lactic acid)—Mass production, processing, industrial applications, and end of life. Adv. Drug Deliv. Rev. 2016, 107, 333–366. [Google Scholar] [CrossRef] [Green Version]
- Tokiwa, Y.; Jarerat, A. Biodegradation of poly(L-lactide). Biotechnol. Lett. 2004, 26, 771–777. [Google Scholar] [CrossRef]
- Jarerat, A.; Tokiwa, Y. Poly(L -lactide) degradation by Saccharothrix waywayandensis. Biotechnol. Lett. 2003, 25, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Jarerat, A.; Tokiwa, Y.; Tanaka, H. Poly(L-lactide) degradation by Kibdelosporangium aridum. Biotechnol. Lett. 2003, 25, 2035–2038. [Google Scholar] [CrossRef]
- Jeon, H.J.; Kim, M.N. Biodegradation of poly (L-lactide) (PLA) exposed to UV irradiation by a mesophilic bacterium. Int. Biodeterior. Biodegrad. 2013, 85, 289–293. [Google Scholar] [CrossRef]
- Butbunchu, N.; Pathom-aree, W. Actinobacteria as promising candidate for polylactic acid type bioplastic degradation. Front. Microbiol. 2019, 10, 2834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, T.A. Developments in Bioplastic Materials for Packaging Food, Beverages and Other Fast-Moving Consumer Goods; Woodhead Publising Limited: Sawston, UK, 2013; pp. 108–152. ISBN 9780857095039. [Google Scholar]
- Lambert, S.; Wagner, M. Environmental performance of bio-based and biodegradable plastics: The road ahead. Chem. Soc. Rev. 2017, 46, 6855–6871. [Google Scholar] [CrossRef] [PubMed]
- Meereboer, K.W.; Misra, M.; Mohanty, A.K. Review of recent advances in the biodegradability of polyhydroxyalkanoate (PHA) bioplastics and their composites. Green Chem. 2020, 22, 5519–5558. [Google Scholar] [CrossRef]
- Prieto-Davó, A.; Dias, T.; Gomes, S.E.; Rodrigues, S.; Parera-Valadez, Y.; Borralho, P.M.; Pereira, F.; Rodrigues, C.M.; Santos-Sanches, I.; Gaudêncio, S.P. The Madeira archipelago as a significant source of marine-derived actinomycete diversity with anticancer and antimicrobial potential. Front. Microbiol. 2016, 7, 1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto-Almeida, A.; Bauermeister, A.; Luppino, L.; Grilo, I.R.; Oliveira, J.; Sousa, J.R.; Petras, D.; Rodrigues, C.F.; Prieto-Davó, A.; Tasdemir, D.; et al. The diversity, metabolomics profiling, and the pharmacological potential of actinomycetes isolated from the Estremadura Spur pockmarks (Portugal). Mar. Drugs 2022, 20, 21. [Google Scholar] [CrossRef]
- Urbanek, A.K.; Rymowicz, W.; Mirończuk, A.M. Degradation of plastics and plastic-degrading bacteria in cold marine habitats. Appl. Microbiol. Biotechnol. 2018, 102, 7669–7678. [Google Scholar] [CrossRef] [Green Version]
- Pometto, A.L., III; Lee, B.; Johnson, K.E. Production of an extracellular polyethylene-degrading enzyme(s) by Streptomyces species. Appl. Environ. Microbiol. 1992, 58, 731–733. [Google Scholar] [CrossRef] [Green Version]
- Abraham, J.; Ghosh, E.; Mukherjee, P.; Gajendiran, A. Microbial degradation of low density polyethylene. Environ. Prog. Sustain. Energy 2017, 36, 147–154. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Pal, S.; Ray, S. Study of microbes having potentiality for biodegradation of plastics. Environ. Sci. Pollut. Res. Int. 2013, 20, 4339–4355. [Google Scholar] [CrossRef]
- Rosario, L.; Dell, E. Biodegradability of plastics testing in an undergraduate materials laboratory course. In Proceedings of the 2010 Annual Conference & Exposition, Louisville, KY, USA, 20–23 June 2010. [Google Scholar] [CrossRef]
- Hossain, U.H.; Muench, F.; Ensinger, W. A comparative study on degradation characteristics of fluoropolymers irradiated by high energy heavy ions. RSC Adv. 2014, 4, 50171–50179. [Google Scholar] [CrossRef]
- Ghatge, S.; Yang, Y.; Ahn, J.H.; Hur, H.-G. Biodegradation of polyethylene: A brief review. Appl. Biol. Chem. 2020, 63, 27. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J.; Jia, R.; Dun, Z.; Wang, B.; Hu, X.; Wang, Y. Selection and evaluation of microorganisms for biodegradation of agricultural plastic film. 3 Biotech 2018, 8, 308. [Google Scholar] [CrossRef] [PubMed]
- Ho, B.T.; Roberts, T.K.; Lucas, S. An overview on biodegradation of polystyrene andmodified polystyrene: The microbial approach. Crit. Rev. Biotechnol. 2018, 38, 308–320. [Google Scholar] [CrossRef]
- Kathiresan, K. Polythene and plastics-degrading microbes from the mangrove soil. Rev. Biol. Trop. 2003, 51, 629–634. [Google Scholar]
- Syranidou, E.; Karkanorachaki, K.; Amorotti, F.; Franchini, M.; Repouskou, E.; Kaliva, M.; Vamvakaki, M.; Kolvenbach, B.; Fava, F.; Corvini, P.F.X.; et al. Biodegradation of weathered polystyrene films in seawater microcosms. Sci. Rep. 2017, 7, 17991. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, A.K.; Vijayakumar, R.P. Studies on biological degradation of polystyrene by pure fungal cultures. Environ. Dev. Sustain. 2020, 22, 4495–4508. [Google Scholar] [CrossRef]
- Coates, J. Interpretation of infrared spectra, a practical approach. Encycl. Anal. Chem. 2006, 1–23. [Google Scholar] [CrossRef]
- Pretsch, E.; Buhlmann, P.; Badertscher, M. Structure Determination of Organic Compounds, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-3-540-93809-5. [Google Scholar]
- Gardette, M.; Perthue, A.; Gardette, J.; Janecska, T.; Földes, E.; Pukánszky, B.; Therias, S. Photo- and thermal-oxidation of polyethylene: Comparison of mechanisms and influence of unsaturation content. Polym. Degrad. Stab. 2013, 98, 2383–2390. [Google Scholar] [CrossRef] [Green Version]
- Abrusci, C.; Pablos, J.; Corrales, T.; López-Marín, J.; Marín, I.; Catalina, F. Biodegradation of photo-degraded mulching films based on polyethylenes and stearates of calcium and iron as pro-oxidant additives. Int. Biodeterior. Biodegrad. 2011, 65, 451–459. [Google Scholar] [CrossRef]
- Montazer, Z.; Habibi-Najafi, M.B.; Mohebbi, M.; Oromiehei, A. Microbial degradation of UV-pretreated low-density polyethylene films by novel polyethylene-degrading bacteria isolated from plastic-dump soil. J. Polym. Environ. 2018, 26, 3613–3625. [Google Scholar] [CrossRef]
- Fang, J.; Xuan, Y.; Li, Q. Preparation of polystyrene spheres in different particle sizes and assembly of the PS colloidal crystals. Sci. China Technol. Sci. 2010, 53, 3088–3093. [Google Scholar] [CrossRef]
- Atiq, N. Biodegradability of Synthetic Plastics Polystyrene and Styrofoam by Fungal Isolates. Ph.D. Thesis, Quaid-i-Azam University, Islamabad, Pakistan, 2011. [Google Scholar]
- Yousif, E.; Haddad, R. Photodegradation and photostabilization of polymers, especially polystyrene: Review. Springerplus 2013, 2, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yang, Y.; Wu, W.; Zhao, J.; Jiang, L. Evidence of polyethylene biodegradation by bacterial strains from the guts of plastic-eating waxworms. Environ. Sci. Technol. 2014, 48, 13776–13784. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Lee, H.M.; Jeon, E.; Yu, H.C.; Lee, S.; Li, J.; Kim, D.-H. Biodegradation of polystyrene by Pseudomonas sp. isolated from the gut of superworms. ChemRxiv 2019. [Google Scholar] [CrossRef]
- Pushpadass, H.A.; Weber, R.W.; Dumais, J.J.; Hanna, M.A. Biodegradation characteristics of starch-polystyrene loose-fill foams in a composting medium. Bioresour. Technol. 2010, 101, 7258–7264. [Google Scholar] [CrossRef] [PubMed]
- Mooney, A.; Ward, P.G.; O’Connor, K.E. Microbial degradation of styrene: Biochemistry, molecular genetics, and perspectives for biotechnological applications. Appl. Microbiol. Biotechnol. 2006, 72, 1–10. [Google Scholar] [CrossRef]
- Shimpi, N.; Borane, M.; Mishra, S.; Kadam, M. Biodegradation of polystyrene (PS)-poly (lactic acid) (PLA) nanocomposites using Pseudomonas aeruginosa. Macromol. Res. 2012, 20, 181–187. [Google Scholar] [CrossRef]
- Ho, T.B. Evaluation of Biodegradability of Polystyrene Materials in the Managed Landfill and Soil. Ph.D. Thesis, University of Newcastle, Callaghan, Australia, 2018. [Google Scholar]
- Auta, H.S.; Emenike, C.U.; Fauziah, S.H. Screening of Bacillus strains isolated from mangrove ecosystems in Peninsular Malaysia for microplastic degradation. Environ. Pollut. 2017, 231, 1552–1559. [Google Scholar] [CrossRef]
- Janczak, K.; Hrynkiewicz, K.; Znajewska, Z.; Dabrowska, G. Use of rhizosphere microorganisms in the biodegradation of PLA and PET polymers in compost soil. Int. Biodeterior. Biodegrad. 2018, 130, 65–75. [Google Scholar] [CrossRef]
- Oliveira, M.; Santos, E.; Araújo, A.; Fechine, G.J.M.; Machado, A.V.; Botelho, G. The role of shear and stabilizer on PLA degradation. Polym. Test. 2016, 51, 109–116. [Google Scholar] [CrossRef]
- Pattanasuttichonlakul, W.; Sombatsompop, N.; Prapagdee, B. Accelerating biodegradation of PLA using microbial consortium from dairy wastewater sludge combined with PLA-degrading bacterium. Int. Biodeterior. Biodegrad. 2018, 132, 74–83. [Google Scholar] [CrossRef]
- Lipsa, R.; Tudorachi, N.; Darie-Nita, R.N.; Oprică, L.; Vasile, C.; Chiriac, A. Biodegradation of poly(lactic acid) and some of its based systems with Trichoderma viride. Int. J. Biol. Macromol. 2016, 88, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Dilara, P.A.; Briassoulis, D. Standard testing methods for mechanical properties and degradation of low density polyethylene (LDPE) films used as greenhouse covering materials: A critical evaluation. Polym. Test. 1998, 17, 549–585. [Google Scholar] [CrossRef]
- Cosgrove, L.; McGeechan, P.L.; Robson, G.D.; Handley, P.S. Fungal communities associated with degradation of polyester polyurethane in soil. Appl. Environ. Microbiol. 2007, 73, 5817–5824. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Nadir, S.; Shah, Z.U.; Shah, A.A.; Karunarathna, S.C.; Xu, J.; Khan, A.; Munir, S.; Hasan, F. Biodegradation of polyester polyurethane by Aspergillus tubingensis. Environ. Pollut. 2017, 225, 469–480. [Google Scholar] [CrossRef]
- Engineers Edge Yield Strength (Mechanics) of Materials. Available online: https://www.engineersedge.com/material_science/yield_strength.htm (accessed on 2 October 2022).
- Engineers Edge Modulus of Elasticity. Young’s Modulus for Common Engineering Materials Table. Available online: https://www.engineersedge.com/manufacturing_spec/properties_of_metals_strength.htm (accessed on 2 October 2022).
- SD3D PLA Technical Data Sheet. Available online: https://www.sd3d.com/wp-content/uploads/2017/06/MaterialTDS-PLA_01.pdf (accessed on 10 October 2022).
- Reis, M.; Albuquerque, M.; Villano, M.; Majone, M. Mixed Culture Processes for Polyhydroxyalkanoate Production from Agro-Industrial Surplus/Wastes as Feedstocks, 2nd ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2011; Volume 6, ISBN 9780080885049. [Google Scholar]
- Matias, F.; Bonatto, D.; Padilla, G.; Rodrigues, M.F.D.A.; Henriques, J.A.P. Polyhydroxyalkanoates production by actinobacteria isolated from soil. Can. J. Microbiol. 2009, 55, 790–800. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Gautam, M.; Maurya, H.K. Recent development of biodegradation techniques of polymer. Int. J. Res. Granthaalayah 2018, 6, 414–452. [Google Scholar] [CrossRef]
- Sowmya, H.V.; Ramalingappa; Krishnappa, M.; Thippeswamy, B. Degradation of polyethylene by Trichoderma harzianum—SEM, FTIR, and NMR analyses. Environ. Monit. Assess. 2014, 186, 6577–6586. [Google Scholar] [CrossRef]
- Orr, I.G.; Hadar, Y.; Sivan, A. Colonization, biofilm formation and biodegradation of polyethylene by a strain of Rhodococcus ruber. Appl. Microbiol. Biotechnol. 2004, 65, 97–104. [Google Scholar] [CrossRef]
- Nowak, B.; Pajak, J.; Drozd-bratkowicz, M.; Rymarz, G. Microorganisms participating in the biodegradation of modified polyethylene films in different soils under laboratory conditions. Int. Biodeterior. Biodegrad. 2011, 65, 757–767. [Google Scholar] [CrossRef]
- Shao, H.; Chen, M.; Fei, X.; Zhang, R.; Zhong, Y.; Ni, W.; Tao, X.; He, X.; Zhang, E.; Yong, B.; et al. Complete genome sequence and characterization of a polyethylene biodegradation strain, Streptomyces albogriseolus LBX-2. Microorganisms 2019, 7, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soleimani, Z.; Gharavi, S.; Soudi, M.; Moosavi-Nejad, Z. A survey of intact low-density polyethylene film biodegradation by terrestrial actinobacterial species. Int. Microbiol. 2020, 24, 65–73. [Google Scholar] [CrossRef]
- Hakkarainen, M.; Albertsson, A. Environmental degradation of polyethylene. Adv. Polym. Sci. 2004, 169, 177–199. [Google Scholar] [CrossRef]
- Esmaeili, A.; Pourbabaee, A.A.; Alikhani, H.A.; Shabani, F.; Esmaeili, E. Biodegradation of low-density polyethylene (LDPE) by mixed culture of Lysinibacillus xylanilyticus and Aspergillus niger in soil. PLoS ONE 2013, 8, e71720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada-Onodera, K.; Mukumoto, H.; Katsuyaya, Y. Degradation of polyethylene by a fungus, Penicillium simplicissimum YK. Polym. Degrad. Stab. 2001, 72, 323–327. [Google Scholar] [CrossRef]
- Jakubowicz, I.; Yarahmadi, N.; Petersen, H. Evaluation of the rate of abiotic degradation of biodegradable polyethylene in various environments. Polym. Degrad. Stab. 2006, 91, 1556–1562. [Google Scholar] [CrossRef]
- Suresh, B.; Maruthamuthu, S.; Kannan, M.; Chandramohan, A. Mechanical and surface properties of low-density polyethylene film modified by photo-oxidation. Polym. J. 2011, 43, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Pometto, A.L., III; Fratzke, A.; Theodore, B.; Bailey, J. Biodegradation of degradable plastic polyethylene by Phanerochaete and Streptomyces species. Appl. Environ. Microbiol. 1991, 57, 678–685. [Google Scholar] [CrossRef] [Green Version]
- Inoue, D.; Suzuki, Y.; Uchida, T.; Morohoshi, J.; Sei, K. Polyhydroxyalkanoate production potential of heterotrophic bacteria in activated sludge. J. Biosci. Bioeng. 2016, 121, 47–51. [Google Scholar] [CrossRef]
- Valappil, S.P.; Boccaccini, A.R.; Bucke, C.; Roy, I. Polyhydroxyalkanoates in Gram-positive bacteria: Insights from the genera Bacillus and Streptomyces. Antonie Van Leeuwenhoek 2007, 91, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.S.; Ahmad, I.; Ishaq, M. Degradation study of used polystyrene with UV irradiation. Adv. Mater. Sci. 2017, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mor, R.; Sivan, A. Biofilm formation and partial biodegradation of polystyrene by the actinomycete Rhodococcus ruber. Biodegradation 2008, 19, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Yottakot, S.; Leelavatcharamas, V. Isolation and optimisation of polylactic acid (PLA) -packaging- degrading actinomycete for PLA-packaging degradation. Pertanika J. Trop. Agric. Sci. 2019, 42, 1111–1129. [Google Scholar]
- Suutari, M.; Lignell, U.; Hyvarinen, A.; Nevalainen, A. Media for cultivation of indoor Strptomycetes. J. Microbiol. Methods 2002, 51, 411–416. [Google Scholar] [CrossRef]
- Tang, Z.-L.; Kuo, T.-A.; Liu, H.-H. The study of the microbes degraded polystyrene. Adv. Technol. Innov. 2017, 2, 13–17. [Google Scholar]
- Jasso, G.C.F.; González-Ortiz, L.J.; Contreras, J.R.; Mensizábal, M.E. The degradation of high impact polystyrene with and without starch in concentrated activated sludge. Polym. Eng. Sci. 1998, 38, 863–869. [Google Scholar] [CrossRef]
- Kiatkamjornwong, S.; Sonsuk, M.; Wittayapichet, S.; Prasassarakich, P.; Vejjanukroh, P.-C. Degradation of styrene-g-cassava starch filled polystyrene plastics. Polym. Degrad. Stab. 1999, 66, 323–335. [Google Scholar] [CrossRef]
- Wältermann, M.; Steinbüchel, A. Neutral lipid bodies in prokaryotes: Recent insights into structure, formation, and relationship to eukaryotic lipid depots. J. Bacteriol. 2005, 187, 3607–3619. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, M.; Kumarasamy, S.; Kandasamy, G.; Kandasamy, S.; Narayanamoorthy, B.; Shanmugam, S.; Alharbi, S.A.; Almoallim, H.S.; Pugazhendhi, A. A novel insight into the fabrication of polyhydroxyalkanoates from actinobacteria Streptomyces toxytricini D2: Screening, optimization, and biopolymer characterization. J. Polym. Environ. 2022, 30, 2128–2141. [Google Scholar] [CrossRef]
- Franco-Correa, M.; Gómez-Méndez, D.; Castro-Medina, N.; Rendón-Ruiz, M. Polyhydroxyalkanoate of actinomycetes native from Colombian soils. Rev. Peru. Biol. 2009, 16, 115–118. [Google Scholar]
- Alvarez, H.M.; Kalscheuer, R.; Steinbüchel, A. Accumulation of storage lipids in species of Rhodococcus and Nocardia and effect of inhibitors and polyethylene glycol. Eur. J. Lipid Sci. Technol. 1997, 99, 239–246. [Google Scholar] [CrossRef]
- Kenny, S.T.; Runic, J.N.; Kaminsky, W.; Woods, T.; Babu, R.P.; Keely, C.M.; Blau, W.; O’Connor, K.E. Up-cycling of PET (polyethylene terephthalate) to the biodegradable plastic PHA (polyhydroxyalkanoate). Environ. Sci. Technol. 2008, 42, 7696–7701. [Google Scholar] [CrossRef] [PubMed]
- Radecka, I.; Irorere, V.; Jiang, G.; Hill, D.; Williams, C.; Adamus, G.; Kwiecień, M.; Marek, A.A.; Zawadiak, J.; Johnston, B.; et al. Oxidized polyethylene wax as a potential carbon source for PHA production. Materials 2016, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Pratt, D.B.; Waddell, G. Adaptation of marine bacteria to growth in media lacking sodium chloride. Nature 1959, 183, 1208–1209. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Linares, L.; Acquaviva, M.; Bertrand, J.-C.; Gauthier, M. Effect of sodium chloride concentration on growth and degradation of eicosane by the marine halotolerant bacterium Marinobacter hydrocarbonoclasticus. Syst. Appl. Microbiol. 1996, 19, 113–121. [Google Scholar] [CrossRef]
- Said, G.; Ahmad, F. Effects of salt concentration on the production of cytotoxic geodin from marine-derived fungus Aspergillus sp. Turk. J. Biochem. 2022, 47, 399–402. [Google Scholar] [CrossRef]
- Jarerat, A.; Pranamuda, H.; Tokiwa, Y. Poly(L-lactide)-degrading activity in various actinomycete. Macromol. Biosci. 2002, 2, 420–428. [Google Scholar] [CrossRef]
- Jarerat, A.; Tokiwa, Y.; Tanaka, H. Production of poly(L-lactide) -degrading enzyme by Amycolatopsis orientalis for biological recycling of poly(L -lactide). Appl. Microbiol. Biotechnol. 2006, 72, 726–731. [Google Scholar] [CrossRef]
- Bennur, T.; Kumar, A.R.; Zinjarde, S.; Javdekar, V. Nocardiopsis species: Incidence, ecological roles and adaptations. Microbiol. Res. 2015, 174, 33–47. [Google Scholar] [CrossRef]
- Walczak, M.; Brzezinska, M.S.; Sionkowska, A.; Michalska, M.; Jankiewicz, U.; Deja-Sikora, E. Biofilm formation on the surface of polylactide during its biodegradation in different environments. Colloids Surf. B Biointerfaces 2015, 136, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Sedničková, M.; Pekařová, S.; Kucharczyk, P.; Bočkaj, J.; Janigová, I.; Kleinová, A.; Jochec-Mošková, D.; Omaníková, L.; Perďochová, D.; Koutný, M.; et al. Changes of physical properties of PLA-based blends during early stage of biodegradation in compost. Int. J. Biol. Macromol. 2018, 113, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Janczak, K.; Dąbrowska, G.B.; Raszkowska-Kaczor, A.; Kaczor, D.; Hrynkiewicz, K.; Richert, A. Biodegradation of the plastics PLA and PET in cultivated soil with the participation of microorganisms and plants. Int. Biodeterior. Biodegrad. 2020, 155, 105087. [Google Scholar] [CrossRef]
- Uchida, H.; Nakajima-kambe, T.; Shigeno-Akutsu, Y.; Nomura, N.; Tokiwa, Y.; Nakahara, T. Properties of a bacterium which degrades solid poly(tetramethylene succinate)-co-adipate, a biodegradable plastic. FEMS Microbiol. Lett. 2000, 189, 25–29. [Google Scholar] [CrossRef]
- Urbanek, A.K.; Rymowicz, W.; Strzelecki, M.C.; Kociuba, W.; Franczak, Ł.; Mirończuk, A.M. Isolation and characterization of Arctic microorganisms decomposing bioplastics. AMB Express 2017, 7, 148. [Google Scholar] [CrossRef]
- Sodergard, A.; Stolt, M. Properties of lactic acid based polymers and their correlation with composition. Prog. Polym. Sci. 2002, 27, 1123–1163. [Google Scholar] [CrossRef]
- Scalenghe, R. Resource or waste? A perspective of plastics degradation in soil with a focus on end-of-life options. Heliyon 2018, 4, e00941. [Google Scholar] [CrossRef] [Green Version]
- Vey, E.; Miller, A.F.; Claybourn, M.; Saiani, A. In Vitro degradation of poly(lactic-co-glycolic) acid random copolymers. Macromol. Symp. 2007, 251, 81–87. [Google Scholar] [CrossRef]
- Ostle, A.G.; Holt, J.G. Nile Blue A as a fluorescent stain for poly-β-hydroxybutyrate. Appl. Environ. Microbiol. 1982, 44, 238–241. [Google Scholar] [CrossRef] [Green Version]
- Serafim, L.S.; Lemos, P.C.; Levantesi, C.; Tandoi, V.; Santos, H.; Reis, M.A.M. Methods for detection and visualization of intracellular polymers stored by polyphosphate-accumulating microorganisms. J. Microbiol. Methods 2002, 51, 1–18. [Google Scholar] [CrossRef]
- Aly, M.M.; Tork, S.; Qari, H.A.; Al-Seeni, M.N. Poly-β-hydroxy butyrate depolymerase from Streptomyces lydicus MM10, isolated from wastewater sample. Int. J. Agric. Biol. 2015, 17, 891–900. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polymer Type | UV Treatment | Time (Days) | Actinomycete Strains | Weight Loss (%) |
|---|---|---|---|---|
| LDPE | No | 180 | S. gougerotti | 0.50 ± 0.08 |
| M. matsumotoense | 0.31 ± 0.02 | |||
| Yes | 90 | S. gougerotti | 0.56 ± 0.12 | |
| M. matsumotoense | 0.22 ± 0.01 | |||
| PS | No | 180 | S. gougerotti | 0.17 ± 0.01 |
| M. matsumotoense | 0.04 ± 0.00 | |||
| Yes | 90 | S. gougerotti | 0.30 ± 0.11 | |
| M. matsumotoense | 0.17 ± 0.04 | |||
| PS with yeast extract | No | 180 | S. gougerotti | 0.67 ± 0.03 |
| Yes | 90 | 0.19 ± 0.04 | ||
| PLA | No | 60 | N. prasina | 0.64 ± 0.20 |
| Yes | 0.97 ± 0.07 | |||
| PLA with yeast extract | Yes | 60 | N. prasina | 1.27 ± 0.19 |
| Actinomycete Strains | Polymer Type | UV Treatment | Band Wave Number | Chemical Changes |
|---|---|---|---|---|
| Streptomyces gougerotti | LDPE | No | New bands at 1739, 1367, and 1217 cm−1 | Formation of carbonyl groups; primary and secondary alcohols |
| Yes | New band at 1096 cm−1 | Oxidized functional groups | ||
| PS | No | New bands at 3600 and 1740 cm−1 | Alcohols generation; formation of carbonyl groups | |
| Yes | New band at 1740 cm−1; Decrease band intensity at 1026, 748, and 694 cm−1 | Indication of oxidation; mono-substituted benzene rings | ||
| PS with yeast extract | No | New bands at 1740, 1216, 1367, and 3600 cm−1 (low intensity) | Formation of carbonyl groups; carboxylic acids, esters; and generation of alcohols | |
| Yes | Increased band intensity; disappearance of the band at 1740 cm−1 | Evidence of film oxidation | ||
| Micromonospora matsumotoense | LDPE | No | New band at 1740 cm−1 | Formation of carbonyl groups |
| Yes | New band at 1070 cm−1 | Chain oxidation | ||
| PS | No | Decrease bands intensity | Biodegradation of polymer chains | |
| Yes | Increased bands intensity | Indication of slight transformation of the chemical structure | ||
| Nocardiopsis prasina | PLA | No | Decrease bands intensity | Biodegradation of polymer chains |
| Yes | New bands at 1382, 1127, and 704 cm−1 | Chain oxidation | ||
| PLA with yeast extract | Yes | New bands at 1382, 1266, 1128, and 700 cm−1 | Formation of vinyl un-saturated groups; and chain oxidation |
| Polymer Type | UV Treatment | Time (Days) | Actinomycete Strains | Young Modulus Decrease (%) | Yield Strength Decrease (%) | Ultimate Tensile Strength Decrease (%) |
|---|---|---|---|---|---|---|
| LDPE | No | 180 | S. gougerotti | 31.1 | 30.6 | 19 |
| M. matsumotoense | 40.7 | 18.3 | 9.7 | |||
| Yes | 90 | S. gougerotti | 12.4 | 25.5 | 5.4 | |
| M. matsumotoense | 12.4 | 25.9 | 20.7 | |||
| PS | No | 180 | S. gougerotti | N.D. | N.D. | N.D. |
| M. matsumotoense | 31.8 | N.D. | N.D. | |||
| Yes | 90 | S. gougerotti | N.D. | 16.7 | 0.7 | |
| M. matsumotoense | 3.51 | N.D. | N.D. | |||
| PS with yeast extract | No | 180 | S. gougerotti | 58.8 | N.D. | N.D. |
| Yes | 90 | 56.4 | 0.2 | 3 | ||
| PLA | No | 60 | N. prasina | 18.1 | N.D. | N.D. |
| Yes | 2.43 | N.D. | N.D. | |||
| PLA with yeast extract | Yes | 60 | N. prasina | 27.4 | N.D. | N.D. |
| Polymer Type | Polymer Quantity (g) | Melting Point (°C) | Press Temperature (°C) | Time on Heat Press (min) |
|---|---|---|---|---|
| LDPE | 1 | 135 | 150 | 10 |
| PS | 1 | 240 | 260 | 5 |
| PLA | 1.5 | ~150 | 200 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, J.; Almeida, P.L.; Sobral, R.G.; Lourenço, N.D.; Gaudêncio, S.P. Marine-Derived Actinomycetes: Biodegradation of Plastics and Formation of PHA Bioplastics—A Circular Bioeconomy Approach. Mar. Drugs 2022, 20, 760. https://doi.org/10.3390/md20120760
Oliveira J, Almeida PL, Sobral RG, Lourenço ND, Gaudêncio SP. Marine-Derived Actinomycetes: Biodegradation of Plastics and Formation of PHA Bioplastics—A Circular Bioeconomy Approach. Marine Drugs. 2022; 20(12):760. https://doi.org/10.3390/md20120760
Chicago/Turabian StyleOliveira, Juliana, Pedro L. Almeida, Rita G. Sobral, Nídia D. Lourenço, and Susana P. Gaudêncio. 2022. "Marine-Derived Actinomycetes: Biodegradation of Plastics and Formation of PHA Bioplastics—A Circular Bioeconomy Approach" Marine Drugs 20, no. 12: 760. https://doi.org/10.3390/md20120760