
CD34—Structure, Functions and Relationship with Cancer Stem Cells
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Structure and Functions of CD34

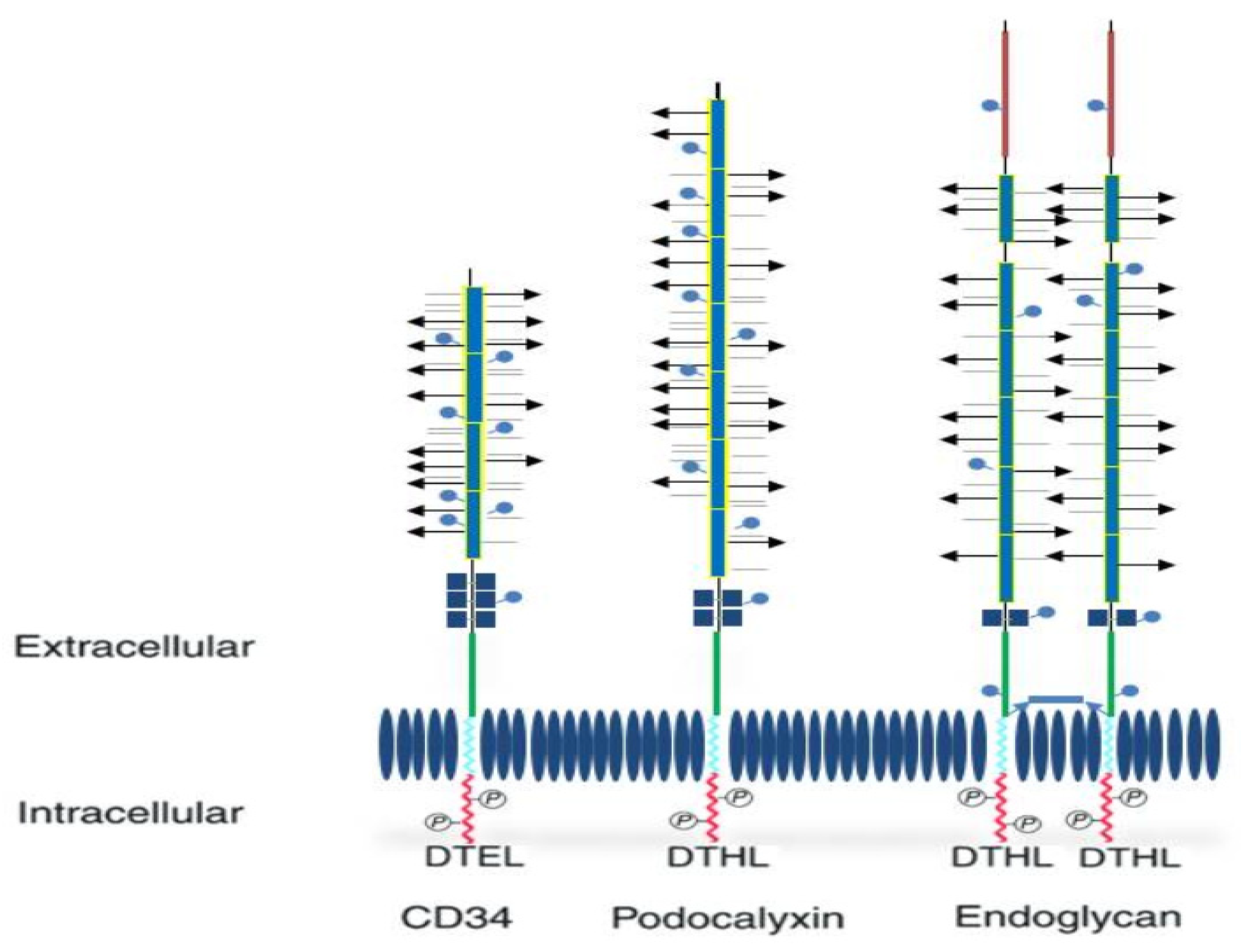{kind=link}
| Structural Characteristics | CD34 | Endoglycan | Podocalyxin |
|---|---|---|---|
| Mucin domain length | 120 amino acids | 350 amino acids | 250 amino acids |
| Cysteine globular domains | Three pairs | Single pair | Two pairs |
| Cysteine in juxtamembrane region | Absent | Unpaired—involved in its homodimerization | Absent |
| N-Linked glycosylation sites | Numerous | Numerous | Numerous |
| Extracellular effective size range (kDa) | 90–170 | 90–170 | 90–170 |
| Nonglycosylated N-terminal sequences | Absent | Present—high abundance of glutamic acid. | Absent |
| C-terminal binding pattern | Mildly modified—which has functional implications for intracellular ligand binding | Similar to podocalyxin | Similar to endoglycan |

- (a)
- Enhancing proliferation and blocking cell differentiation.
- (b)
- Enhance lymphocyte adhesion.
- (c)
- Cellular development
2.1. CD 34 and Hematopoietic Cells
2.2. CD 34 Multipotent Mesenchymal Stromal Cells (MSC)
2.3. CD 34 and Muscle Stem Cells
2.4. CD34 and Endothelial Cells
2.5. CD 34 and Cancer Stem Cells (CSCs)
2.6. CD34 in Clinical Applications
3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tindle, R.W.; Nichols, R.A.; Chan, L.; Campana, D.; Catovsky, D.; Birnie, G.D. A novel monoclonal antibody BI-3C5 recognises myeloblasts and non-B non-T lymphoblasts in acute leukaemias and CGL blast crises, and reacts with immature cells in normal bone marrow. Leuk. Res. 1985, 9, 1–9. [Google Scholar] [CrossRef]
- Civin, C.I.; Strauss, L.C.; Brovall, C.; Fackler, M.J.; Schwartz, J.F.; Shaper, J.H. Antigenic analysis of hematopoiesis. III. A hematopoietic progenitor cell surface antigen defined by a monoclonal antibody raised against KG-1a cells. J. Immunol. 1984, 133, 157–165. [Google Scholar] [CrossRef]
- Sidney, L.E.; Branch, M.J.; Dunphy, S.E.; Dua, H.S.; Hopkinson, A. Concise review: Evidence for CD34 as a common marker for diverse progenitors. Stem Cells 2014, 32, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Fina, L.; Molgaard, H.V.; Robertson, D.; Bradley, N.J.; Monaghan, P.; Delia, D.; Sutherland, D.R.; Baker, M.A.; Greaves, M.F. Expression of the CD34 gene in vascular endothelial cells. Blood 1990, 75, 2417–2426. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Greaves, M.F.; Molgaard, H.V. The gene encoding the stem cell antigen, CD34, is conserved in mouse and expressed in haemopoietic progenitor cell lines, brain, and embryonic fibroblasts. Int. Immunol. 1991, 3, 175–184. [Google Scholar] [CrossRef]
- Nielsen, J.S.; McNagny, K.M. Novel functions of the CD34 family. J. Cell Sci. 2008, 121, 3683–3692. [Google Scholar] [CrossRef]
- Baumheter, S.; Singer, M.S.; Henzel, W.; Hemmerich, S.; Renz, M.; Rosen, S.D.; Lasky, L.A. Binding of L-selectin to the vascular sialomucin CD34. Science 1993, 262, 436–438. [Google Scholar] [CrossRef]
- Baumhueter, S.; Dybdal, N.; Kyle, C.; Lasky, L.A. Global vascular expression of murine CD34, a sialomucin-like endothelial ligand for L-selectin. Blood 1994, 84, 2554–2565. [Google Scholar] [CrossRef]
- Doyonnas, R.; Nielsen, J.S.; Chelliah, S.; Drew, E.; Hara, T.; Miyajima, A.; McNagny, K.M. Podocalyxin is a CD34-related marker of murine hematopoietic stem cells and embryonic erythroid cells. Blood 2005, 105, 4170–4178. [Google Scholar] [CrossRef]
- Doyonnas, R.; Kershaw, D.B.; Duhme, C.; Merkens, H.; Chelliah, S.; Graf, T.; McNagny, K.M. Anuria, omphalocele, and perinatal lethality in mice lacking the CD34-related protein podocalyxin. J. Exp. Med. 2001, 194, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Kerjaschki, D.; Sharkey, D.J.; Farquhar, M.G. Identification and characterization of podocalyxin—The major sialoprotein of the renal glomerular epithelial cell. J. Cell Biol. 1984, 98, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Kelley, T.W.; Huntsman, D.; McNagny, K.M.; Roskelley, C.D.; Hsi, E.D. Podocalyxin: A marker of blasts in acute leukemia. Am. J. Clin. Pathol. 2005, 124, 134–142. [Google Scholar] [CrossRef]
- Casey, G.; Neville, P.J.; Liu, X.; Plummer, S.J.; Cicek, M.S.; Krumroy, L.M.; Curran, A.P.; McGreevy, M.R.; Catalona, W.J.; Klein, E.A.; et al. Podocalyxin variants and risk of prostate cancer and tumor aggressiveness. Hum. Mol. Genet. 2006, 15, 735–741. [Google Scholar] [CrossRef]
- Sassetti, C.; Van Zante, A.; Rosen, S.D. Identification of endoglycan, a member of the CD34/podocalyxin family of sialomucins. J. Biol. Chem. 2000, 275, 9001–9010. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, S.; Shenoy, S.P.; Bose, B. CD34 cells in somatic, regenerative and cancer stem cells: Developmental biology, cell therapy, and omics big data perspective. J. Cell. Biochem. 2020, 121, 3058–3069. [Google Scholar] [CrossRef] [PubMed]
- Dawood, S.; Austin, L.; Cristofanilli, M. Cancer stem cells: Implications for cancer therapy. Oncology 2014, 28, 1101–1107. [Google Scholar]
- Takeda, T. Podocyte cytoskeleton is connected to the integral membrane protein podocalyxin through Na+/H+-exchanger regulatory factor 2 and ezrin. Clin. Exp. Nephrol. 2003, 7, 260–269. [Google Scholar] [CrossRef]
- Hilkens, J.; Ligtenberg, M.J.; Vos, H.L.; Litvinov, S.V. Cell membrane-associated mucins and their adhesion-modulating property. Trends Biochem. Sci. 1992, 17, 359–363. [Google Scholar] [CrossRef]
- Sassetti, C.; Tangemann, K.; Singer, M.S.; Kershaw, D.B.; Rosen, S.D. Identification of podocalyxin-like protein as a high endothelial venule ligand for L-selectin: Parallels to CD34. J. Exp. Med. 1998, 187, 1965–1975. [Google Scholar] [CrossRef]
- He, X.Y.; Antao, V.P.; Basila, D.; Marx, J.C.; Davis, B.R. Isolation and molecular characterization of the human CD34 gene. Blood 1992, 79, 2296–2302. [Google Scholar] [CrossRef] [PubMed]
- Furness, S.G.; McNagny, K. Beyond mere markers: Functions for CD34 family of sialomucins in hematopoiesis. Immunol. Res. 2006, 34, 13–32. [Google Scholar] [CrossRef]
- Nielsen, J.S.; McNagny, K.M. CD34 is a key regulator of hematopoietic stem cell trafficking to bone marrow and mast cell progenitor trafficking in the periphery. Microcirculation 2009, 16, 487–496. [Google Scholar] [CrossRef]
- Krause, D.S.; Fackler, M.J.; Civin, C.I.; May, W.S. CD34: Structure, biology, and clinical utility. Blood 1996, 87, 1–13. [Google Scholar] [CrossRef]
- Cheng, J.; Baumhueter, S.; Cacalano, G.; Carver-Moore, K.; Thibodeaux, H.; Thomas, R.; Broxmeyer, H.E.; Cooper, S.; Hague, N.; Moore, M.; et al. Hematopoietic defects in mice lacking the sialomucin CD34. Blood 1996, 87, 479–490. [Google Scholar] [CrossRef]
- Nielsen, J.S.; McNagny, K.M. Influence of host irradiation on long-term engraftment by CD34-deficient hematopoietic stem cells. Blood 2007, 110, 1076–1077. [Google Scholar] [CrossRef] [PubMed]
- Drew, E.; Merzaban, J.S.; Seo, W.; Ziltener, H.J.; McNagny, K.M. CD34 and CD43 inhibit mast cell adhesion and are required for optimal mast cell reconstitution. Immunity 2005, 22, 43–57. [Google Scholar] [CrossRef]
- Lasky, L.A. Selectins: Interpreters of cell-specific carbohydrate information during inflammation. Science 1992, 258, 964–969. [Google Scholar] [CrossRef] [PubMed]
- Fieger, C.B.; Sassetti, C.M.; Rosen, S.D. Endoglycan, a member of the CD34 family, functions as an L-selectin ligand through modification with tyrosine sulfation and sialyl Lewis x. J. Biol. Chem. 2003, 278, 27390–27398. [Google Scholar] [CrossRef] [PubMed]
- Sarangapani, K.K.; Yago, T.; Klopocki, A.G.; Lawrence, M.B.; Fieger, C.B.; Rosen, S.D.; McEver, R.P.; Zhu, C. Low force decelerates L-selectin dissociation from P-selectin glycoprotein ligand-1 and endoglycan. J. Biol. Chem. 2004, 279, 2291–2298. [Google Scholar] [CrossRef]
- Healy, L.; May, G.; Gale, K.; Grosveld, F.; Greaves, M.; Enver, T. The stem cell antigen CD34 functions as a regulator of hemopoietic cell adhesion. Proc. Natl. Acad. Sci. USA 1995, 92, 12240–12244. [Google Scholar] [CrossRef]
- Larrucea, S.; Butta, N.; Arias-Salgado, E.G.; Alonso-Martin, S.; Ayuso, M.S.; Parrilla, R. Expression of podocalyxin enhances the adherence, migration, and intercellular communication of cells. Exp. Cell Res. 2008, 314, 2004–2015. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, E.; Dekan, G.; Miettinen, A.; Farquhar, M.G. Biogenesis of podocalyxin—The major glomerular sialoglycoprotein—In the newborn rat kidney. Eur. J. Cell Biol. 1989, 48, 313–326. [Google Scholar] [PubMed]
- Vitureira, N.; McNagny, K.; Soriano, E.; Burgaya, F. Pattern of expression of the podocalyxin gene in the mouse brain during development. Gene Expr. Patterns GEP 2005, 5, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Gao, S.Y.; Chen, J.; Saito, K.; Miyawaki, K.; Li, C.Y.; Pan, L.; Saito, S.; Terashita, T.; Matsuda, S. Process formation of the renal glomerular podocyte: Is there common molecular machinery for processes of podocytes and neurons? Anat. Sci. Int. 2004, 79, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, A.; Solin, M.L.; Reivinen, J.; Juvonen, E.; Vaisanen, R.; Holthofer, H. Podocalyxin in rat platelets and megakaryocytes. Am. J. Pathol. 1999, 154, 813–822. [Google Scholar] [CrossRef]
- Berenson, R.J.; Bensinger, W.I.; Hill, R.S.; Andrews, R.G.; Garcia-Lopez, J.; Kalamasz, D.F.; Still, B.J.; Spitzer, G.; Buckner, C.D.; Bernstein, I.D.; et al. Engraftment after infusion of CD34+ marrow cells in patients with breast cancer or neuroblastoma. Blood 1991, 77, 1717–1722. [Google Scholar] [CrossRef] [PubMed]
- Majeti, R.; Park, C.Y.; Weissman, I.L. Identification of a hierarchy of multipotent hematopoietic progenitors in human cord blood. Cell Stem Cell 2007, 1, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Huss, R. Isolation of primary and immortalized CD34-hematopoietic and mesenchymal stem cells from various sources. Stem Cells 2000, 18, 1–9. [Google Scholar] [CrossRef]
- Mao, Q.; Chu, S.; Ghanta, S.; Padbury, J.F.; De Paepe, M.E. Ex vivo expanded human cord blood-derived hematopoietic progenitor cells induce lung growth and alveolarization in injured newborn lungs. Respir. Res. 2013, 14, 37. [Google Scholar] [CrossRef]
- Jang, Y.Y.; Collector, M.I.; Baylin, S.B.; Diehl, A.M.; Sharkis, S.J. Hematopoietic stem cells convert into liver cells within days without fusion. Nat. Cell Biol. 2004, 6, 532–539. [Google Scholar] [CrossRef]
- da Silva Meirelles, L.; Chagastelles, P.C.; Nardi, N.B. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J. Cell Sci. 2006, 119, 2204–2213. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; Prockop, D.J. Concise review: Mesenchymal stem/multipotent stromal cells: The state of transdifferentiation and modes of tissue repair—Current views. Stem Cells 2007, 25, 2896–2902. [Google Scholar] [CrossRef]
- Lin, C.S.; Ning, H.; Lin, G.; Lue, T.F. Is CD34 truly a negative marker for mesenchymal stromal cells? Cytotherapy 2012, 14, 1159–1163. [Google Scholar] [CrossRef]
- Scherberich, A.; Di Maggio, N.D.; McNagny, K.M. A familiar stranger: CD34 expression and putative functions in SVF cells of adipose tissue. World J. Stem Cells 2013, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Simmons, P.J.; Torok-Storb, B. CD34 expression by stromal precursors in normal human adult bone marrow. Blood 1991, 78, 2848–2853. [Google Scholar] [CrossRef]
- Quirici, N.; Soligo, D.; Bossolasco, P.; Servida, F.; Lumini, C.; Deliliers, G.L. Isolation of bone marrow mesenchymal stem cells by anti-nerve growth factor receptor antibodies. Exp. Hematol. 2002, 30, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, G.A.; De Francesco, F.; Nicoletti, G.; Paino, F.; Desiderio, V.; Tirino, V.; D’Andrea, F. Human adipose CD34+ CD90+ stem cells and collagen scaffold constructs grafted in vivo fabricate loose connective and adipose tissues. J. Cell. Biochem. 2013, 114, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, F.; Tirino, V.; Desiderio, V.; Ferraro, G.; D’Andrea, F.; Giuliano, M.; Libondi, G.; Pirozzi, G.; De Rosa, A.; Papaccio, G. Human CD34/CD90 ASCs are capable of growing as sphere clusters, producing high levels of VEGF and forming capillaries. PLoS ONE 2009, 4, e6537. [Google Scholar] [CrossRef]
- Kopher, R.A.; Penchev, V.R.; Islam, M.S.; Hill, K.L.; Khosla, S.; Kaufman, D.S. Human embryonic stem cell-derived CD34+ cells function as MSC progenitor cells. Bone 2010, 47, 718–728. [Google Scholar] [CrossRef]
- Sinanan, A.C.; Hunt, N.P.; Lewis, M.P. Human adult craniofacial muscle-derived cells: Neural-cell adhesion-molecule (NCAM.; CD56)-expressing cells appear to contain multipotential stem cells. Biotechnol. Appl. Biochem. 2004, 40, 25–34. [Google Scholar] [CrossRef]
- Beauchamp, J.R.; Heslop, L.; Yu, D.S.; Tajbakhsh, S.; Kelly, R.G.; Wernig, A.; Buckingham, M.E.; Partridge, T.A.; Zammit, P.S. Expression of CD34 and Myf5 defines the majority of quiescent adult skeletal muscle satellite cells. J. Cell Biol. 2000, 151, 1221–1234. [Google Scholar] [CrossRef]
- Cossu, G.; Molinaro, M.; Pacifici, M. Differential response of satellite cells and embryonic myoblasts to a tumor promoter. Dev. Biol. 1983, 98, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Lecourt, S.; Marolleau, J.P.; Fromigue, O.; Vauchez, K.; Andriamanalijaona, R.; Ternaux, B.; Lacassagne, M.N.; Robert, I.; Boumediene, K.; Chereau, F.; et al. Characterization of distinct mesenchymal-like cell populations from human skeletal muscle in situ and in vitro. Exp. Cell Res. 2010, 316, 2513–2526. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Cao, B.; Crisan, M.; Sun, B.; Li, G.; Logar, A.; Yap, S.; Pollett, J.B.; Drowley, L.; Cassino, T.; et al. Prospective identification of myogenic endothelial cells in human skeletal muscle. Nat. Biotechnol. 2007, 25, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Dupas, T.; Rouaud, T.; Rouger, K.; Lieubeau, B.; Cario-Toumaniantz, C.; Fontaine-Perus, J.; Gardahaut, M.F.; Auda-Boucher, G. Fetal muscle contains different CD34+ cell subsets that distinctly differentiate into adipogenic, angiogenic and myogenic lineages. Stem Cell Res. 2011, 7, 230–243. [Google Scholar] [CrossRef] [PubMed]
- Brenes, R.A.; Bear, M.; Jadlowiec, C.; Goodwin, M.; Hashim, P.; Protack, C.D.; Ziegler, K.R.; Li, X.; Model, L.S.; Lv, W.; et al. Cell-based interventions for therapeutic angiogenesis: Review of potential cell sources. Vascular 2012, 20, 360–368. [Google Scholar] [CrossRef]
- Mackie, A.R.; Losordo, D.W. CD34-positive stem cells: In the treatment of heart and vascular disease in human beings. Tex. Heart Inst. J. 2011, 38, 474–485. [Google Scholar]
- Tondreau, T.; Meuleman, N.; Delforge, A.; Dejeneffe, M.; Leroy, R.; Massy, M.; Mortier, C.; Bron, D.; Lagneaux, L. Mesenchymal stem cells derived from CD133-positive cells in mobilized peripheral blood and cord blood: Proliferation, Oct4 expression, and plasticity. Stem Cells 2005, 23, 1105–1112. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kuroda, R.; Mifune, Y.; Kawamoto, A.; Shoji, T.; Miwa, M.; Asahara, T.; Kurosaka, M. Circulating endothelial/skeletal progenitor cells for bone regeneration and healing. Bone 2008, 43, 434–439. [Google Scholar] [CrossRef]
- Siemerink, M.J.; Klaassen, I.; Vogels, I.M.; Griffioen, A.W.; Van Noorden, C.J.; Schlingemann, R.O. CD34 marks angiogenic tip cells in human vascular endothelial cell cultures. Angiogenesis 2012, 15, 151–163. [Google Scholar] [CrossRef]
- Ye, G.; Qin, Y.; Lu, X.; Xu, X.; Xu, S.; Wu, C.; Wang, X.; Wang, S.; Pan, D. The association of renin-angiotensin system genes with the progression of hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2015, 459, 18–23. [Google Scholar] [CrossRef]
- Clarke, M.F.; Dick, J.E.; Dirks, P.B.; Eaves, C.J.; Jamieson, C.H.; Jones, D.L.; Visvader, J.; Weissman, I.L.; Wahl, G.M. Cancer stem cells—Perspectives on current status and future directions: AACR Workshop on cancer stem cells. Cancer Res. 2006, 66, 9339–9344. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Shi, P.; Zhao, G.; Xu, J.; Peng, W.; Zhang, J.; Zhang, G.; Wang, X.; Dong, Z.; Chen, F.; et al. Targeting cancer stem cell pathways for cancer therapy. Signal Transduct. Target. Ther. 2020, 5, 8. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, T.; Sirard, C.; Vormoor, J.; Murdoch, B.; Hoang, T.; Caceres-Cortes, J.; Minden, M.; Paterson, B.; Caligiuri, M.A.; Dick, J.E. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 1994, 367, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Nguyen, N.T.; Eun, J.R.; Zhang, Y.; Jung, Y.J.; Tschudy-Seney, B.; Trotsyuk, A.; Lam, A.; Ramsamooj, R.; Zhang, Y.; et al. Identification of cancer stem cell subpopulations of CD34(+) PLC/PRF/5 that result in three types of human liver carcinomas. Stem Cells Dev. 2015, 24, 1008–1021. [Google Scholar] [CrossRef]
- Yin, P.; Ono, M.; Moravek, M.B.; Coon, J.S.t.; Navarro, A.; Monsivais, D.; Dyson, M.T.; Druschitz, S.A.; Malpani, S.S.; Serna, V.A.; et al. Human uterine leiomyoma stem/progenitor cells expressing CD34 and CD49b initiate tumors in vivo. J. Clin. Endocrinol. Metab. 2015, 100, E601–E606. [Google Scholar] [CrossRef]
- Aravindan, N.; Somasundaram, D.B.; Herman, T.S.; Aravindan, S. Significance of hematopoietic surface antigen CD34 in neuroblastoma prognosis and the genetic landscape of CD34-expressing neuroblastoma CSCs. Cell Biol. Toxicol. 2021, 37, 461–478. [Google Scholar] [CrossRef]
- Kuroda, R.; Matsumoto, T.; Kawakami, Y.; Fukui, T.; Mifune, Y.; Kurosaka, M. Clinical impact of circulating CD34-positive cells on bone regeneration and healing. Tissue Eng. Part B Rev. 2014, 20, 190–199. [Google Scholar] [CrossRef]
- Berenson, R.J.; Andrews, R.G.; Bensinger, W.I.; Kalamasz, D.; Knitter, G.; Buckner, C.D.; Bernstein, I.D. Antigen CD34+ marrow cells engraft lethally irradiated baboons. J. Clin. Investig. 1988, 81, 951–955. [Google Scholar] [CrossRef]
- Lee, F.Y.; Chen, Y.L.; Sung, P.H.; Ma, M.C.; Pei, S.N.; Wu, C.J.; Yang, C.H.; Fu, M.; Ko, S.F.; Leu, S.; et al. Intracoronary Transfusion of Circulation-Derived CD34+ Cells Improves Left Ventricular Function in Patients With End-Stage Diffuse Coronary Artery Disease Unsuitable for Coronary Intervention. Crit. Care Med. 2015, 43, 2117–2132. [Google Scholar] [CrossRef]
- El-Badawy, A.; El-Badri, N. Clinical Efficacy of Stem Cell Therapy for Diabetes Mellitus: A Meta-Analysis. PLoS ONE 2016, 11, e0151938. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Nakamura, T.; Torimura, T.; Iwamoto, H.; Masuda, H.; Koga, H.; Abe, M.; Hashimoto, O.; Ueno, T.; Sata, M. Transplantation of endothelial progenitor cells ameliorates vascular dysfunction and portal hypertension in carbon tetrachloride-induced rat liver cirrhotic model. J. Gastroenterol. Hepatol. 2013, 28, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Quyyumi, A.A.; Vasquez, A.; Kereiakes, D.J.; Klapholz, M.; Schaer, G.L.; Abdel-Latif, A.; Frohwein, S.; Henry, T.D.; Schatz, R.A.; Dib, N.; et al. PreSERVE-AMI: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial of Intracoronary Administration of Autologous CD34+ Cells in Patients With Left Ventricular Dysfunction Post STEMI. Circ. Res. 2017, 120, 324–331. [Google Scholar] [CrossRef] [PubMed]
| Cell Type | Differentiation Potential | Morphology | Other Markers |
|---|---|---|---|
| Hematopoietic stem/progenitor cells |
|
|
|
| Multipotent stromal cells (MSCs) |
|
|
|
| Muscle stem cells |
|
|
|
| Fibrocytes |
|
|
|
| Endothelial cells |
|
|
|
| Interstitial cells |
|
|
|
| Ligand | Cell Type | Protein Bound | Interaction |
|---|---|---|---|
| L-selectin | High endothelial venules (HEV) | CD 34 Endoglycan Podocalyxin | Sialyl lewis-x carbohydrate dependent |
| NRERF-1 | Hematopoietic cells | Endoglycan Podocalyxin | C-terminal PDZ interaction |
| NHERF-2 | Podocyte | Podocalyxin | Terminal PDZ interaction |
| CRKL | Hematopoietic cells | CD 34 | Juxtamembrane |
| ERM | MDCK (Madin-Darby Canine Kidney) cell line | Podocalyxin | Juxtamembrane |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radu, P.; Zurzu, M.; Paic, V.; Bratucu, M.; Garofil, D.; Tigora, A.; Georgescu, V.; Prunoiu, V.; Pasnicu, C.; Popa, F.; et al. CD34—Structure, Functions and Relationship with Cancer Stem Cells. Medicina 2023, 59, 938. https://doi.org/10.3390/medicina59050938
Radu P, Zurzu M, Paic V, Bratucu M, Garofil D, Tigora A, Georgescu V, Prunoiu V, Pasnicu C, Popa F, et al. CD34—Structure, Functions and Relationship with Cancer Stem Cells. Medicina. 2023; 59(5):938. https://doi.org/10.3390/medicina59050938
Chicago/Turabian StyleRadu, Petru, Mihai Zurzu, Vlad Paic, Mircea Bratucu, Dragos Garofil, Anca Tigora, Valentin Georgescu, Virgiliu Prunoiu, Costin Pasnicu, Florian Popa, and et al. 2023. "CD34—Structure, Functions and Relationship with Cancer Stem Cells" Medicina 59, no. 5: 938. https://doi.org/10.3390/medicina59050938