
Changes in the Global Epidemiology of Type 1 Diabetes in an Evolving Landscape of Environmental Factors: Causes, Challenges, and Opportunities

{kind=link}
Abstract
:1. Introduction
2. Rationale and Search Strategy
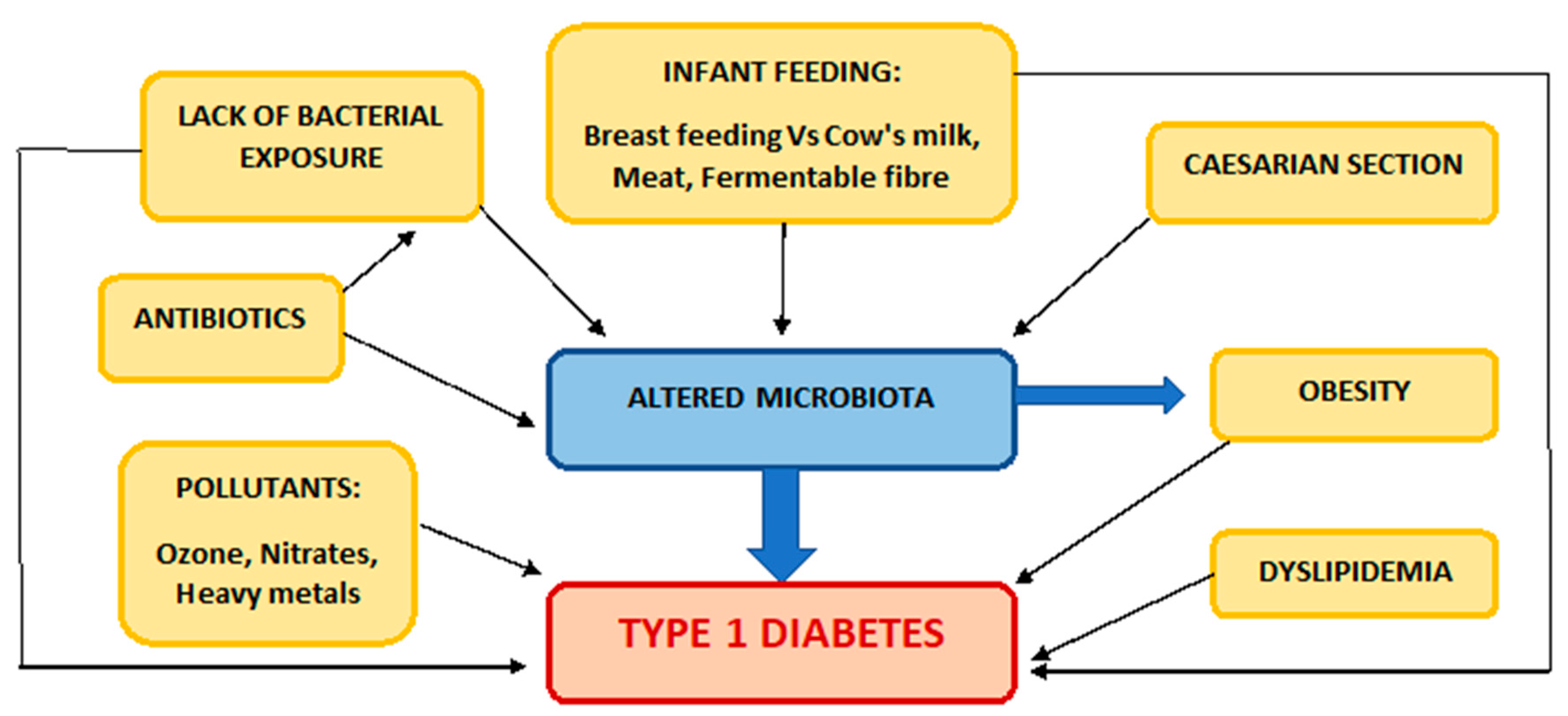
3. Lifestyle Factors
4. Early Life Factors
5. Microbial Exposure and the Hygiene Hypothesis
6. Pollution
7. Ecological and Geographical Factors
8. Gut under the Microscope: The Role of the Microbiome
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gregory, G.; Robinson, T.; Linklater, S.; Wang, F.; Colagiuri, S.; Beaufort, C.; Donaghue, K.; Magliano, D.; Maniam, J.; Orchard, T.; et al. Global incidence, prevalence, and mortality of type 1 diabetes in 2021 with projection to 2040: A modelling study. Lancet Diabetes Endocrinol. 2022, 10, 741–760. [Google Scholar] [CrossRef]
- Snell-Bergeon, J.K.; Maahs, D.M. Diabetes: Elevated risk of mortality in type 1 diabetes mellitus. Nat. Rev. Endocrinol. 2015, 11, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Reutens, A.T. Epidemiology of diabetic kidney disease. Med. Clin. N. Am. 2013, 97, 1–18. [Google Scholar] [CrossRef]
- Nielsen, H.B.; Ovesen, L.L.; Mortensen, L.H.; Lau, C.J.; Joensen, L.E. Type 1 diabetes, quality of life, occupational status and education level—A comparative population-based study. Diabetes Res. Clin. Pract. 2016, 121, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Sharif, K.; Watad, A.; Coplan, L.; Amital, H.; Shoenfeld, Y.; Afek, A. Psychological stress and type 1 diabetes mellitus: What is the link? Expert Rev. Clin. Immunol. 2018, 14, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ren, Z.H.; Qiang, H.; Wu, J.; Shen, M.; Zhang, L.; Lyu, J. Trends in the incidence of diabetes mellitus: Results from the Global Burden of Disease Study 2017 and implications for diabetes mellitus prevention. BMC Public Health 2020, 20, 1415. [Google Scholar] [CrossRef]
- Buzzetti, R.; Zampetti, S.; Pozzilli, P. Impact of obesity on the increasing incidence of type 1 diabetes. Diabetes Obes. Metab. 2020, 22, 1009–1013. [Google Scholar] [CrossRef]
- Ludvigsson, J. Why diabetes incidence increases—A unifying theory. Ann. N. Y. Acad. Sci. 2006, 1079, 374–382. [Google Scholar] [CrossRef]
- Kibirige, M.; Metcalf, B.; Renuka, R.; Wilkin, T.J. Testing the accelerator hypothesis: The relationship between body mass and age at diagnosis of type 1 diabetes. Diabetes Care. 2003, 26, 2865–2870. [Google Scholar] [CrossRef] [Green Version]
- Betts, P.; Mulligan, J.; Ward, P.; Smith, B.; Wilkin, T. Increasing body weight predicts the earlier onset of insulin-dependant diabetes in childhood: Testing the ‘accelerator hypothesis’ (2). Diabet. Med. 2005, 22, 144–151. [Google Scholar] [CrossRef]
- Knerr, I.; Wolf, J.; Reinehr, T.; Stachow, R.; Grabert, M.; Schober, E.; Rascher, W.; Holl, R.W. DPV Scientific Initiative of Germany and Austria. The ‘accelerator hypothesis’: Relationship between weight, height, body mass index and age at diagnosis in a large cohort of 9,248 German and Austrian children with type 1 diabetes mellitus. Diabetologia 2005, 48, 2501–2504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyppönen, E.; Virtanen, S.M.; Kenward, M.G.; Knip, M.; Akerblom, H.K.; Childhood Diabetes in Finland Study Group. Obesity, increased linear growth, and risk of type 1 diabetes in children. Diabetes Care 2000, 23, 1755–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, M.A.; Donath, S.; Cameron, F.J. Major increase in Type 1 diabetes: No support for the accelerator hypothesis. Diabet. Med. 2007, 24, 920–923. [Google Scholar] [CrossRef]
- Vehik, K.; Hamman, R.F.; Lezotte, D.; Norris, J.M.; Klingensmith, G.J.; Dabelea, D. Childhood growth and age at diagnosis with Type 1 diabetes in Colorado young people. Diabet. Med. 2009, 26, 961–967. [Google Scholar] [CrossRef]
- Viner, R.M.; Hindmarsh, P.C.; Taylor, B.; Cole, T.J. Childhood body mass index (BMI), breastfeeding and risk of Type 1 diabetes: Findings from a longitudinal national birth cohort. Diabet. Med. 2008, 25, 1056–1061. [Google Scholar] [CrossRef]
- Ljungkrantz, M.; Ludvigsson, J.; Samuelsson, U. Type 1 diabetes: Increased height and weight gains in early childhood. Pediatr. Diabetes 2008, 9 Pt 2, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Knip, M.; Reunanen, A.; Virtanen, S.M.; Nuutinen, M.; Viikari, J.; Akerblom, H.K. Does the secular increase in body mass in children contribute to the increasing incidence of type 1 diabetes? Pediatr. Diabetes 2008, 9 Pt 2, 46–49. [Google Scholar] [CrossRef]
- Vella, S.; Bezzina Sultana, M.; Fava, S. Association of cholesterol and lifestyle markers with type 1 diabetes incidence rates at a population level. J. Public Health 2017, 39, 542–548. [Google Scholar]
- Rydgren, T.; Vaarala, O.; Sandler, S. Simvastatin protects against multiple low-dose streptozotocin-induced type 1 diabetes in CD-1 mice and recurrence of disease in nonobese diabetic mice. J. Pharmacol. Exp. Ther. 2007, 323, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Strom, A.; Kolb, H.; Martin, S.; Herder, C.; Simon, M.-C.; Koenig, W.; Heise, T.; Heinemann, L.; Roden, M.; Schloot, N.C.; et al. DIATOR Study Group. Improved preservation of residual beta cell function by atorvastatin in patients with recent onset type 1 diabetes and high CRP levels (DIATOR trial). PLoS ONE 2012, 7, e33108. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Paulsson, G.; Stemme, S.; Hansson, G.K. Hypercholesterolemia is associated with a T helper (Th) 1/Th2 switch of the autoimmune response in atherosclerotic apo E-knockout mice. J. Clin. Investig. 1998, 101, 1717–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cnop, M.; Hannaert, J.C.; Grupping, A.Y.; Pipeleers, D.G. Low density lipoprotein can cause death of islet beta-cells by its cellular uptake and oxidative modification. Endocrinology 2002, 143, 3449–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, L.; Xu, L.; Lin, Y.; Peng, Y. Statins and beta-cell function. Med. Sci. Monit. 2010, 16, HY1-2. [Google Scholar]
- Boljat, A.; Gunjača, I.; Konstantinović, I.; Vidan, N.; Perica, V.B.; Pehlić, M.; Škrabić, V.; Zemunik, T. Environmental risk factors for type 1 diabetes mellitus development. Exp. Clin. Endocrinol. Diabetes 2017, 125, 563–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muntoni, S.; Mereu, R.; Atzori, L.; Mereu, A.; Galassi, S.; Corda, S.; Frongia, P.; Angius, E.; Pusceddu, P.; Contu, P.; et al. High meat consumption is associated with type 1 diabetes mellitus in a Sardinian case-control study. Acta Diabetol. 2013, 50, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Syrjälä, E.; Nevalainen, J.; Peltonen, J.; Takkinen, H.M.; Hakola, L.; Åkerlund, M.; Veijola, R.; Ilonen, J.; Toppari, J.; Knip, M.; et al. A joint modeling approach for childhood meat, fish and egg consumption and the risk of advanced islet autoimmunity. Sci. Rep. 2019, 9, 7760. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Lopera, N.; Pineda-Trujillo, N.; Diaz-Valencia, P.A. Correlating the global increase in type 1 diabetes incidence across age groups with national economic prosperity: A systematic review. World J. Diabetes 2019, 10, 560–580. [Google Scholar] [CrossRef]
- Manuwald, U.; Heinke, P.; Salzsieder, E.; Hegewald, J.; Schoffer, O.; Kugler, J.; Kapellen, T.M.; Kiess, W.; Rothe, U. Incidence trends of type 1 diabetes before and after the reunification in children up to 14 years of age in Saxony, Eastern German. PLoS ONE 2017, 12, e0183665. [Google Scholar] [CrossRef] [Green Version]
- Onkamo, P.; Vaananen, S.; Karvonen, M.; Tuomilehto, J. Worldwide increase of type 1 diabetes Fanalysis of the data on published incidence trends. Diabetologia 1999, 42, 1395–1403. [Google Scholar] [CrossRef] [Green Version]
- Wilkin, T. The accelerator hypothesis: A review of the evidence for insulin resistance as the basis for type I as well as type II diabetes. Int. J. Obes. 2009, 33, 716–726. [Google Scholar] [CrossRef] [Green Version]
- Devendra, D.; Liu, E.; Eisenbarth, G.S. Type 1 diabetes: Recent developments. BMJ 2004, 27, 750–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gale, E.A. To boldly go--or to go too boldly? The accelerator hypothesis revisited. Diabetologia 2007, 50, 1571–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blom, L.; Persson, L.A.; Dahlquist, G. A high linear growth is associated with an increased risk of childhood diabetes mellitus. Diabetologia 1992, 35, 528–533. [Google Scholar] [CrossRef] [Green Version]
- Wasyl-Nawrot, B.; Wójcik, M.; Nazim, J.; Skupień, J.; Starzyk, J.B. Increased Incidence of Type 1 Diabetes in Children and No Change in the Age of Diagnosis and BMI-SDS at the Onset—Is the Accelerator Hypothesis not Working? J. Clin. Res. Pediatr. Endocrinol. 2020, 12, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Cardwell, C.R.; Stene, L.C.; Joner, G.; Cinek, O.; Svensson, J.; Goldacre, M.J.; Parslow, R.C.; Pozzilli, P.; Brigis, G.; Stoyanov, D.; et al. Caesarean section is associated with an increased risk of childhood-onset type 1 diabetes mellitus: A meta-analysis of observational studies. Diabetologia 2008, 51, 726–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanoey, J.; Gulati, A.; Patterson, C.; Becher, H. Risk of type 1 diabetes in the offspring born through elective or non-elective caesarean section in comparison to vaginal delivery: A meta-analysis of observational studies. Curr. Diab. Rep. 2019, 19, 124. [Google Scholar] [CrossRef] [PubMed]
- Waernbaum, I.; Dahlquist, G.; Lind, T. Perinatal risk factors for type 1 diabetes revisited: A population-based register study. Diabetologia 2019, 62, 1173–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Caesarean Section Rates Continue to Rise, Amid Growing Inequalities in Access: WHO. 2021. Available online: https://www.who.int/news/item/16-06-2021-caesarean-section-rates-continue-to-rise-amid-growing-inequalities-in-access-who (accessed on 1 February 2023).
- Lund-Blix, N.A.; Dydensborg Sander, S.; Størdal, K.; Andersen, A.-M.N.; Rønningen, K.S.; Joner, G.; Skrivarhaug, T.; Njølstad, P.R.; Husby, S.; Stene, L.C. Infant feeding and risk of type 1 diabetes in two large scandinavian birth cohorts. Diabetes Care 2017, 40, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Güngör, D.; Nadaud, P.; LaPergola, C.C.; Dreibelbis, C.; Wong, Y.P.; Terry, N.; Abrams, S.A.; Beker, L.; Jacobovits, T.; Järvinen, K.M. Infant milk-feeding practices and diabetes outcomes in offspring: A systematic review. Am. J. Clin. Nutr. 2019, 109 (Suppl. S7), 817S–837S. [Google Scholar] [CrossRef]
- Knip, M.; Åkerblom, H.K.; Becker, D.; Dosch, H.-M.; Dupre, J.; Fraser, W.; Howard, N.; Ilonen, J.; Krischer, J.P.; Kordonouri, O.; et al. TRIGR Study Group. Hydrolyzed infant formula and early β-cell autoimmunity: A randomized clinical trial. JAMA 2014, 311, 2279–2287. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, M.; Andersen, Z.J.; Nordsborg, R.B.; Becker, T.; Tjønneland, A.; Overvad, K.; Raaschou-Nielsen, O. Long-term exposure to road traffic noise and incident diabetes: A cohort study. Environ. Health Perspect. 2013, 121, 217–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Van’t Land, B.; Engen, P.A.; Naqib, A.; Green, S.J.; Nato, A.; Leusink-Muis, T.; Garssen, J.; Keshavarzian, A.; Stahl, B.; et al. Human milk oligosaccharides protect against the development of autoimmune diabetes in NOD-mice. Sci. Rep. 2018, 8, 3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rook, G.A. Hygiene hypothesis and autoimmune diseases. Clin. Rev. Allergy Immunol. 2012, 42, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Bach, J.F. The hygiene hypothesis in autoimmunity: The role of pathogens and commensals. Nat. Rev. Immunol. 2018, 18, 105–120. [Google Scholar] [CrossRef]
- Abela, A.G.; Fava, S. Association of incidence of type 1 diabetes with mortality from infectious disease and with antibiotic susceptibility at a country level. Acta Diabetol. 2013, 50, 859–865. [Google Scholar] [CrossRef]
- EURODIAB Substudy 2 Study Group. Infections and vaccinations as risk factors for childhood type I (insulin-dependent) diabetes mellitus: A multicentre case-control investigation. Diabetologia 2000, 43, 47–53. [Google Scholar] [CrossRef] [Green Version]
- de Goffau, M.C.; Luopajärvi, K.; Knip, M.; Ilonen, J.; Ruohtula, T.; Härkönen, T.; Orivuori, L.; Hakala, S.; Welling, G.W.; Harmsen, H.J.; et al. Fecal microbiotacomposition differs between children with β-cell autoimmunity and those without. Diabetes 2013, 62, 1238–1244. [Google Scholar] [CrossRef] [Green Version]
- Boursi, B.; Mamtani, R.; Haynes, K.; Yang, Y.X. The effect of past antibiotic exposure on diabetes risk. Eur. J. Endocrinol. 2015, 172, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Hathout, E.H.; Beeson, W.L.; Ischander, M.; Rao, R.; Mace, J.W. Air pollution and type 1 diabetes in children. Pediatr. Diabetes 2006, 7, 81–87. [Google Scholar] [CrossRef]
- Malmqvist, E.; Larsson, H.E.; Jönsson, I.; Rignell-Hydbom, A.; Ivarsson, S.-A.; Tinnerberg, H.; Stroh, E.; Rittner, R.; Jakobsson, K.; Swietlicki, E.; et al. Maternal exposure to air pollution and type 1 diabetes-Accounting for genetic factors. Environ. Res. 2015, 140, 268–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elten, M.; Donelle, J.; Lima, I.; Burnett, R.T.; Weichenthal, S.; Stieb, D.M.; Hystad, P.; van Donkelaar, A.; Chen, H.; Paul, L.A.; et al. Ambient air pollution and incidence of early-onset paediatric type 1 diabetes: A retrospective population-based cohort study. Environ. Res. 2020, 184, 109291. [Google Scholar] [CrossRef] [PubMed]
- Jantzen, K.; Jensen, A.; Kermanizadeh, A.; Elholm, G.; Sigsgaard, T.; Møller, P.; Roursgaard, M.; Loft, S. Inhalation of house dust and ozone alters systemic levels of endothelial progenitor cells, oxidative stress, and inflammation in elderly subjects. Toxicol. Sci. 2018, 163, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Lodovici, M.; Bigagli, E. Oxidative stress and air pollution exposure. J. Toxicol. 2011, 2011, 487074. [Google Scholar] [CrossRef]
- Kurai, J.; Onuma, K.; Sano, H.; Okada, F.; Watanabe, M. Ozone augments interleukin-8 production induced by ambient particulate matter. Genes Environ. 2018, 40, 14. [Google Scholar] [CrossRef]
- Holland, N.; Davé, V.; Venkat, S.; Wong, H.; Donde, A.; Balmes, J.; Arjomandi, M. Ozone inhalation leads to a dose-dependent increase of cytogenetic damage in human lymphocytes. Environ. Mol. Mutagen. 2015, 56, 378–387. [Google Scholar] [CrossRef] [Green Version]
- Vella, R.E.; Pillon, N.J.; Zarrouki, B.; Croze, M.L.; Koppe, L.; Guichardant, M.; Pesenti, S.; Chauvin, M.-A.; Rieusset, J.; Géloën, A.; et al. Ozone exposure triggers insulin resistance through muscle c-Jun N-terminal kinase activation. Diabetes 2015, 64, 1011–1024. [Google Scholar] [CrossRef] [Green Version]
- Tsai, D.H.; Riediker, M.; Berchet, A.; Paccaud, F.; Waeber, G.; Vollenweider, P.; Bochud, M. Effects of short- and long-term exposures to particulate matter on inflammatory marker levels in the general population. Environ. Sci. Pollut. Res. Int. 2019, 26, 19697–19704. [Google Scholar] [CrossRef]
- Bengalli, R.; Molteni, E.; Longhin, E.; Refsnes, M.; Camatini, M.; Gualtieri, M. Release of IL-1 β triggered by Milan summer PM10:molecular pathways involved in the cytokine release. BioMed Res. Int. 2013, 2013, 158093. [Google Scholar] [CrossRef] [Green Version]
- Gruzieva, O.; Merid, S.K.; Gref, A.; Gajulapuri, A.; Lemonnier, N.; Ballereau, S.; Gigante, B.; Kere, J.; Auffray, C.; Melén, E.; et al. Exposure to traffic-related air pollution and serum inflammatory cytokines in children. Environ. Health Perspect. 2017, 125, 067007. [Google Scholar] [CrossRef] [Green Version]
- Perret, J.L.; Bowatte, G.; Lodge, C.J.; Knibbs, L.D.; Gurrin, L.C.; Kandane-Rathnayake, R.; Johns, D.P.; Lowe, A.J.; Burgess, J.A.; Thompson, B.R.; et al. The dose-response association between nitrogen dioxide exposure and serum interleukin-6 concentrations. Int. J. Mol. Sci. 2017, 18, 1015. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Huang, G.; Nagy, T.; Guo, T.L. Bisphenol A alteration of type 1 diabetes in non-obese diabetic (NOD) female mice is dependent on window of exposure. Arch. Toxicol. 2019, 93, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Cetkovic-Cvrlje, M.; Olson, M.; Schindler, B.; Gong, H.K. Exposure to DDT metabolite p,p′-DDE increases autoimmune type 1 diabetes incidence in NOD mouse model. J. Immunotoxicol. 2016, 13, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Bagenstose, L.M.; Salgame, P.; Monestier, M. Cytokine regulation of a rodent model of mercuric chloride-induced autoimmunity. Environ. Health Perspect. 1999, 107 (Suppl. S5), 807–810. [Google Scholar] [PubMed]
- Ludvigsson, J.; Andersson-White, P.; Guerrero-Bosagna, C. Toxic metals in cord blood and later development of Type 1 diabetes. Pediatr. Dimens. 2019, 4. [Google Scholar] [CrossRef]
- Chen, Y.L.; Huang, Y.C.; Qiao, Y.C.; Ling, W.; Pan, Y.H.; Geng, L.J.; Xiao, J.L.; Zhang, X.X.; Zhao, H.L. Climates on incidence of childhood type 1 diabetes mellitus in 72 countries. Sci. Rep. 2017, 7, 12810. [Google Scholar] [CrossRef] [Green Version]
- Koufakis, T.; Karras, S.N.; Mustafa, O.G.; Zebekakis, P.; Kotsa, K. The Effects of High Altitude on Glucose Homeostasis, Metabolic Control, and Other Diabetes-Related Parameters: From Animal Studies to Real Life. High Alt. Med. Biol. 2019, 20, 1–11. [Google Scholar] [CrossRef]
- Mobasseri, M.; Shirmohammadi, M.; Amiri, T.; Vahed, N.; Hosseini Fard, H.; Ghojazadeh, M. Prevalence and incidence of type 1 diabetes in the world: A systematic review and meta-analysis. Health Promot. Perspect. 2020, 10, 98–115. [Google Scholar] [CrossRef]
- Kanta, A.; Lyka, E.; Koufakis, T.; Zebekakis, P.; Kotsa, K. Prevention strategies for type 1 diabetes: A story of promising efforts and unmet expectations. Hormones 2020, 19, 453–465. [Google Scholar] [CrossRef]
- Muntoni, S.; Fonte, M.T.; Stoduto, S.; Marietti, G.; Bizzarri, C.; Crinò, A.; Ciampalini, P.; Multari, G.; Suppa, M.A.; Matteoli, M.C.; et al. Incidence of insulindependent diabetes mellitus among Sardinian heritage children born in the Lazio region, Italy. Lancet 1997, 349, 160–162. [Google Scholar] [CrossRef]
- Dorman, J.; McCarthy, B.J.; O’Leary, L.A.; Koehler, A.N. Risk factors for insulin-dependent diabetes. In Diabetes in America, 2nd ed.; NIH Publication No. 95-1468; National Institute of Health: Bethesda, MD, USA, 1995; pp. 165–177. [Google Scholar]
- Dorman, J.S. (Ed.) Standardization of Epidemiologic Studies of Host Susceptibility; Plenum Press: New York, NY, USA, 1994. [Google Scholar]
- Serrano-Rìos, M.; Goday, A.; Martìnez Larrad, T. Migrant populations and the incidence of Type 1 diabetes mellitus: An overview of the literature with a focus on the Spanish-heritage countries in Latin America. Diabetes/Metab. Res. Rev. 1999, 15, 113–132. [Google Scholar] [CrossRef]
- Zheng, P.; Li, Z.; Zhou, Z. Gut microbiome in type 1 diabetes: A comprehensive review. Diabetes Metab. Res. Rev. 2018, 34, e3043. [Google Scholar] [CrossRef]
- Petersen, C.; Round, J.L. Defining dysbiosis and its influence on host immunity and disease. Cell. Microbiol. 2014, 16, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Tai, N.; Peng, J.; Liu, F.; Gulden, E.; Hu, Y.; Zhang, X.; Chen, L.; Wong, F.S.; Wen, L. Microbial antigen mimics activate diabetogenic CD8 T cells in NOD mice. J. Exp. Med. 2016, 213, 2129–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hevia, A.; Milani, C.; López, P.; Cuervo, A.; Arboleya, S.; Duranti, S.; Turroni, F.; González, S.; Suárez, A.; Gueimonde, M.; et al. Intestinal dysbiosis associated with systemic lupus erythematosus. mBio 2014, 5, e01548-14. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.T.; Davis-Richardson, A.G.; Giongo, A.; Gano, K.A.; Crabb, D.B.; Mukherjee, N.; Casella, G.; Drew, J.C.; Ilonen, J.; Knip, M.; et al. Gut microbiome metagenomics analysis suggests a functional model for the development of autoimmunity for type 1 diabetes. PLoS ONE 2011, 6, e25792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maffeis, C.; Martina, A.; Corradi, M.; Quarella, S.; Nori, N.; Torriani, S.; Plebani, M.; Contreas, G.; Felis, G.E. Association between intestinal permeability and faecal microbiota composition in Italian children with beta cell autoimmunity at risk for type 1 diabetes. Diabetes Metab. Res. Rev. 2016, 32, 700–709. [Google Scholar] [CrossRef]
- Ruiz, V.E.; Battaglia, T.; Kurtz, Z.D.; Bijnens, L.; Ou, A.; Engstrand, I.; Zheng, X.; Iizumi, T.; Mullins, B.J.; Müller, C.L.; et al. A single early-in-life macrolide course has lasting effects on murine microbial network topology and immunity. Nat. Commun. 2017, 8, 518. [Google Scholar] [CrossRef] [Green Version]
- Valladares, R.; Sankar, D.; Li, N.; Williams, E.; Lai, K.K.; Abdelgeliel, A.S.; Gonzalez, C.F.; Wasserfall, C.H.; Larkin, J.; Schatz, D.; et al. Lactobacillus johnsonii N6.2 mitigates the development of type 1 diabetes in BB-DP rats. PLoS ONE 2010, 5, e10507. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.R.; Françozo, M.C.; de Oliveira, G.G.; Ignacio, A.; Castoldi, A.; Zamboni, D.S.; Ramos, S.G.; Câmara, N.O.; de Zoete, M.R.; Palm, N.W.; et al. Gut microbiota translocation to the pancreatic lymph nodes triggers NOD2 activation and contributes to T1D onset. J. Exp. Med. 2016, 213, 1223–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, C.H.; Krych, L.; Nielsen, D.S.; Vogensen, F.K.; Hansen, L.H.; Sørensen, S.J.; Buschard, K.; Hansen, A.K. Early life treatment with vancomycin propagates Akkermansia muciniphila and reduces diabetes incidence in the NOD mouse. Diabetologia 2012, 55, 2285–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marietta, E.V.; Gomez, A.M.; Yeoman, C.; Tilahun, A.Y.; Clark, C.R.; Luckey, D.H.; Murray, J.A.; White, B.A.; Kudva, Y.C.; Rajagopalan, G. Low incidence of spontaneous type 1 diabetes in non-obese diabetic mice raised on gluten-free diets is associated with changes in the intestinal microbiome. PLoS ONE 2013, 8, e78687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Pan, L.L.; Niu, W.; Fang, X.; Liang, W.; Li, J.; Li, H.; Pan, X.; Chen, W.; Zhang, H.; et al. Modulation of Gut Microbiota by Low Methoxyl Pectin Attenuates Type 1 Diabetes in Non-obese Diabetic Mice. Front. Immunol. 2019, 10, 1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Heck, J.I.P.; Gacesa, R.; Stienstra, R.; Fu, J.; Zhernakova, A.; Harmsen, H.J.M.; Weersma, R.K.; Joosten, L.A.B.; Tack, C.J. The Gut Microbiome Composition Is Altered in Long-standing Type 1 Diabetes and Associates With Glycemic Control and Disease-Related Complications. Diabetes Care 2022, 45, 2084–2094. [Google Scholar] [CrossRef]
- de Groot, P.; Nikolic, T.; Pellegrini, S.; Sordi, V.; Imangaliyev, S.; Rampanelli, E.; Hanssen, N.; Attaye, I.; Bakker, G.; Duinkerken, G.; et al. Faecal microbiota transplantation halts progression of human new-onset type 1 diabetes in a randomised controlled trial. Gut 2021, 70, 92–105. [Google Scholar] [CrossRef]
- Ho, N.T.; Li, F.; Lee-Sarwar, K.A.; Tun, H.M.; Brown, B.P.; Pannaraj, P.S.; Bender, J.M.; Azad, M.B.; Thompson, A.L.; Weiss, S.T.; et al. Meta-analysis of effects of exclusive breastfeeding on infant gut microbiota across populations. Nat. Commun. 2018, 9, 4169. [Google Scholar] [CrossRef] [Green Version]
- Sadauskaite-Kuehne, V.; Ludvigsson, J.; Padaiga, Z.; Jasinskiene, E.; Samuelsson, U. Longer breastfeeding is an independent protective factor against development of type 1 diabetes mellitus in childhood. Diabetes Metab. Res. Rev. 2004, 20, 150–157. [Google Scholar] [CrossRef]
- Uusitalo, U.; Liu, X.; Yang, J.; Aronsson, C.A.; Hummel, S.; Butterworth, M.; Lernmark, Å.; Rewers, M.; Hagopian, W.; She, J.X.; et al. Association of Early Exposure of Probiotics and Islet Autoimmunity in the TEDDY Study. JAMA Pediatr. 2016, 170, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.; Nicolucci, A.C.; Virtanen, H.; Schick, A.; Meddings, J.; Reimer, R.A.; Huang, C. Effect of Prebiotic on Microbiota, Intestinal Permeability, and Glycemic Control in Children With Type 1 Diabetes. J. Clin. Endocrinol. Metab. 2019, 104, 4427–4440. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Sun, L.; Zhang, S.; Zhao, X.; Gang, X.; Wang, G. Evaluating the Causal Role of Gut Microbiota in Type 1 Diabetes and Its Possible Pathogenic Mechanisms. Front. Endocrinol. 2020, 11, 125. [Google Scholar] [CrossRef]
- Lee, H.; Ko, G. Effect of metformin on metabolic improvement and gut microbiota. Appl. Environ. Microbiol. 2014, 80, 5935–5943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kihl, P.; Krych, L.; Buschard, K.; Wesley, J.D.; Kot, W.; Hansen, A.K.; Nielsen, D.S.; von Herrath, M.G. Oral insulin does not alter gut microbiota composition of NOD mice. Diabetes Metab. Res. Rev. 2018, 34, e3010. [Google Scholar] [CrossRef] [PubMed]
- Waldron-Lynch, F.; Henegariu, O.; Deng, S.; Preston-Hurlburt, P.; Tooley, J.; Flavell, R.; Herold, K.C. Teplizumab induces human gut-tropic regulatory cells in humanized mice and patients. Sci. Transl. Med. 2012, 4, 118ra12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis-Richardson, A.G.; Ardissone, A.N.; Dias, R.; Simell, V.; Leonard, M.T.; Kemppainen, K.M.; Drew, J.C.; Schatz, D.; Atkinson, M.A.; Kolaczkowski, B.; et al. Bacteroides dorei dominates gut microbiome prior to autoimmunity in Finnish children at high risk for type 1 diabetes. Front. Microbiol. 2014, 5, 678. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogrotis, I.; Koufakis, T.; Kotsa, K. Changes in the Global Epidemiology of Type 1 Diabetes in an Evolving Landscape of Environmental Factors: Causes, Challenges, and Opportunities. Medicina 2023, 59, 668. https://doi.org/10.3390/medicina59040668
Ogrotis I, Koufakis T, Kotsa K. Changes in the Global Epidemiology of Type 1 Diabetes in an Evolving Landscape of Environmental Factors: Causes, Challenges, and Opportunities. Medicina. 2023; 59(4):668. https://doi.org/10.3390/medicina59040668
Chicago/Turabian StyleOgrotis, Ioannis, Theocharis Koufakis, and Kalliopi Kotsa. 2023. "Changes in the Global Epidemiology of Type 1 Diabetes in an Evolving Landscape of Environmental Factors: Causes, Challenges, and Opportunities" Medicina 59, no. 4: 668. https://doi.org/10.3390/medicina59040668