
Evaluating the Magnolol Anticancer Potential in MKN-45 Gastric Cancer Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagent Preparation
2.2. Cell Culture Protocol
2.3. MTT Assay
2.4. Study Design
2.5. Cell Proliferation Assay
2.6. Colony Formation Analysis
2.7. Real-Time PCR
2.8. Statistical Analysis
3. Results
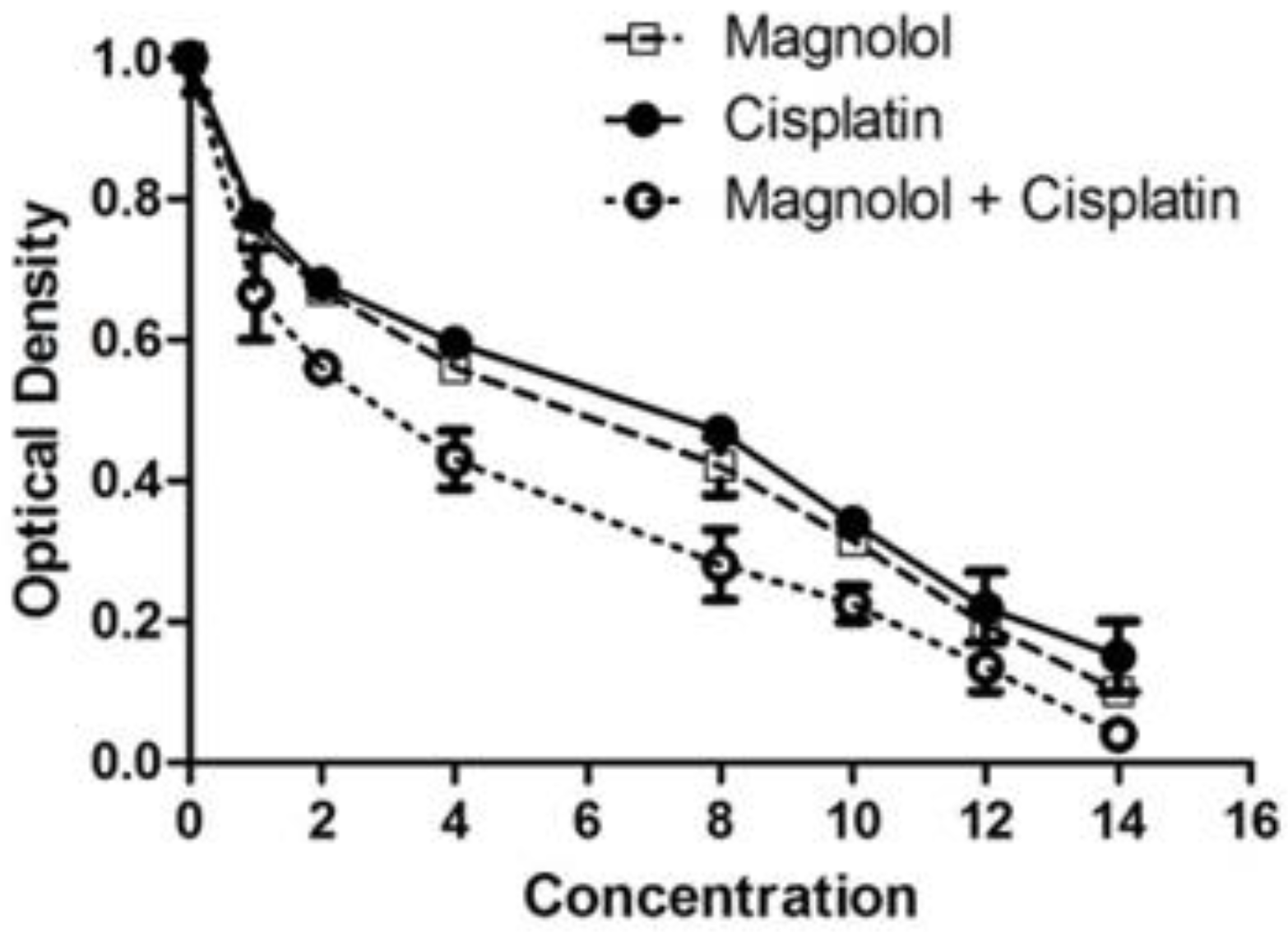
3.1. The Effects of Magnolol and/or Cisplatin on MKN-45 Cells’ Viability
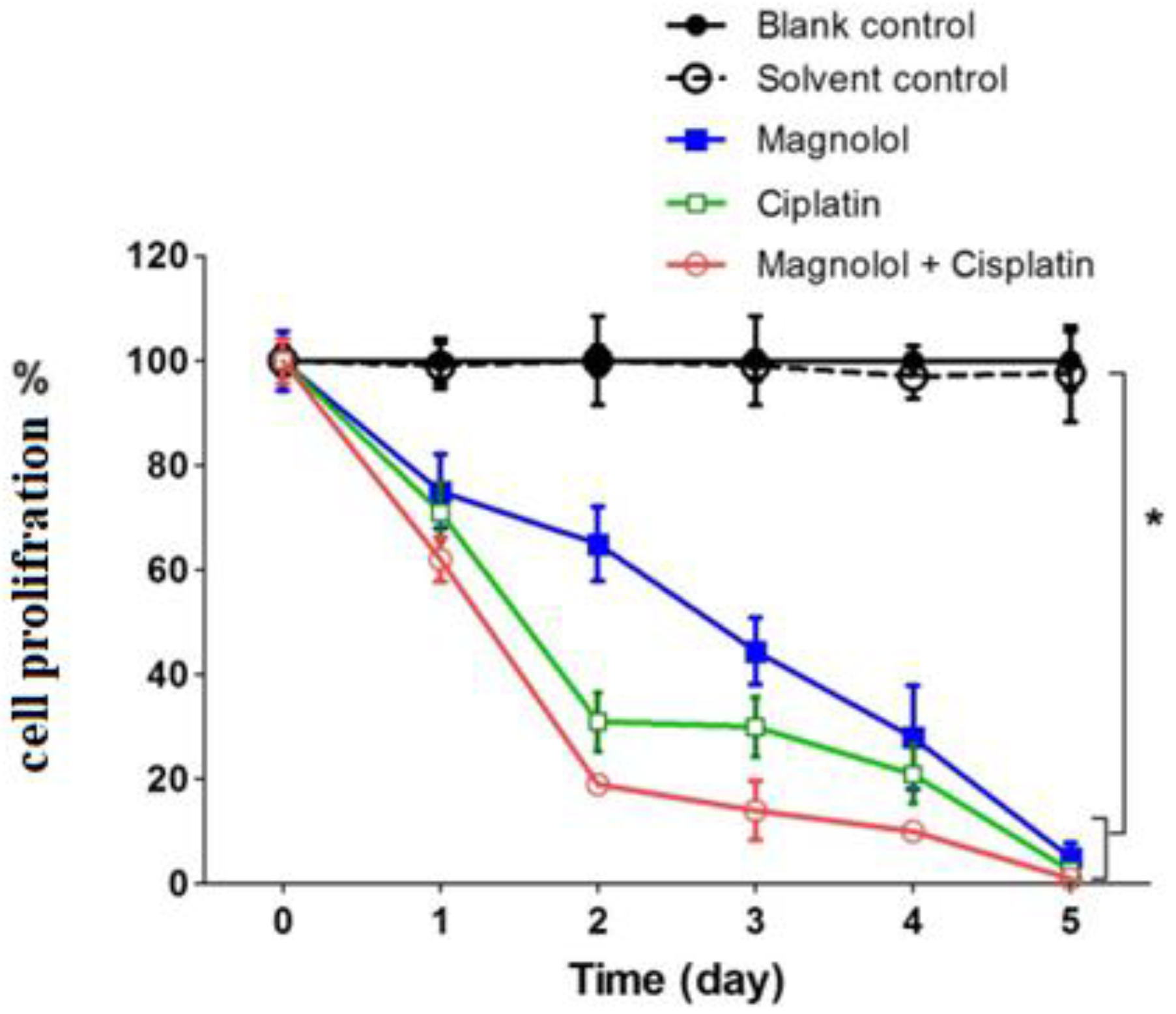
3.2. The Effects of Magnolol and/or Cisplatin on MKN-45 Cells’ Proliferation
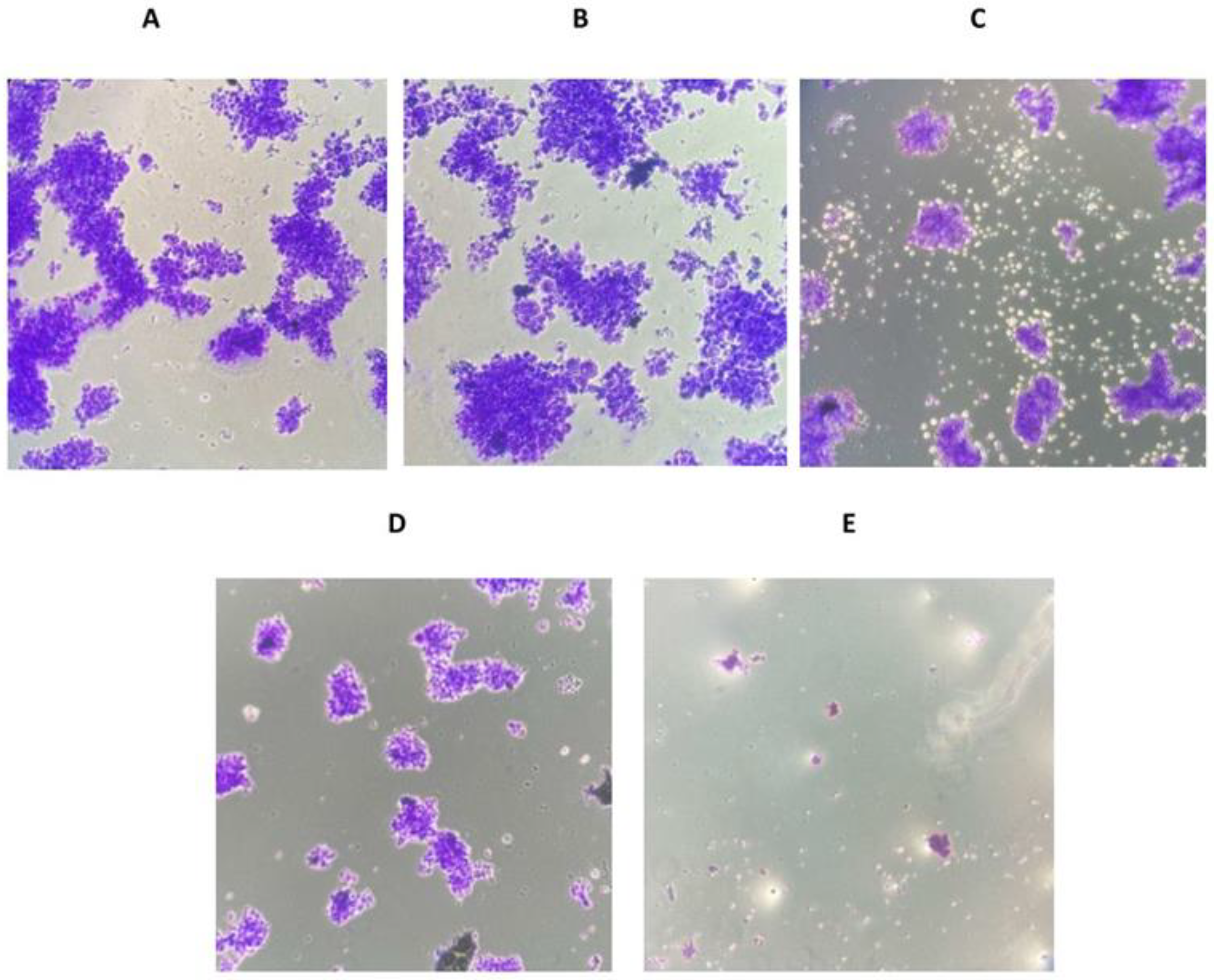
3.3. The Effects of Magnolol and/or Cisplatin on the Ability of MKN-45 Cells to Form Colonies
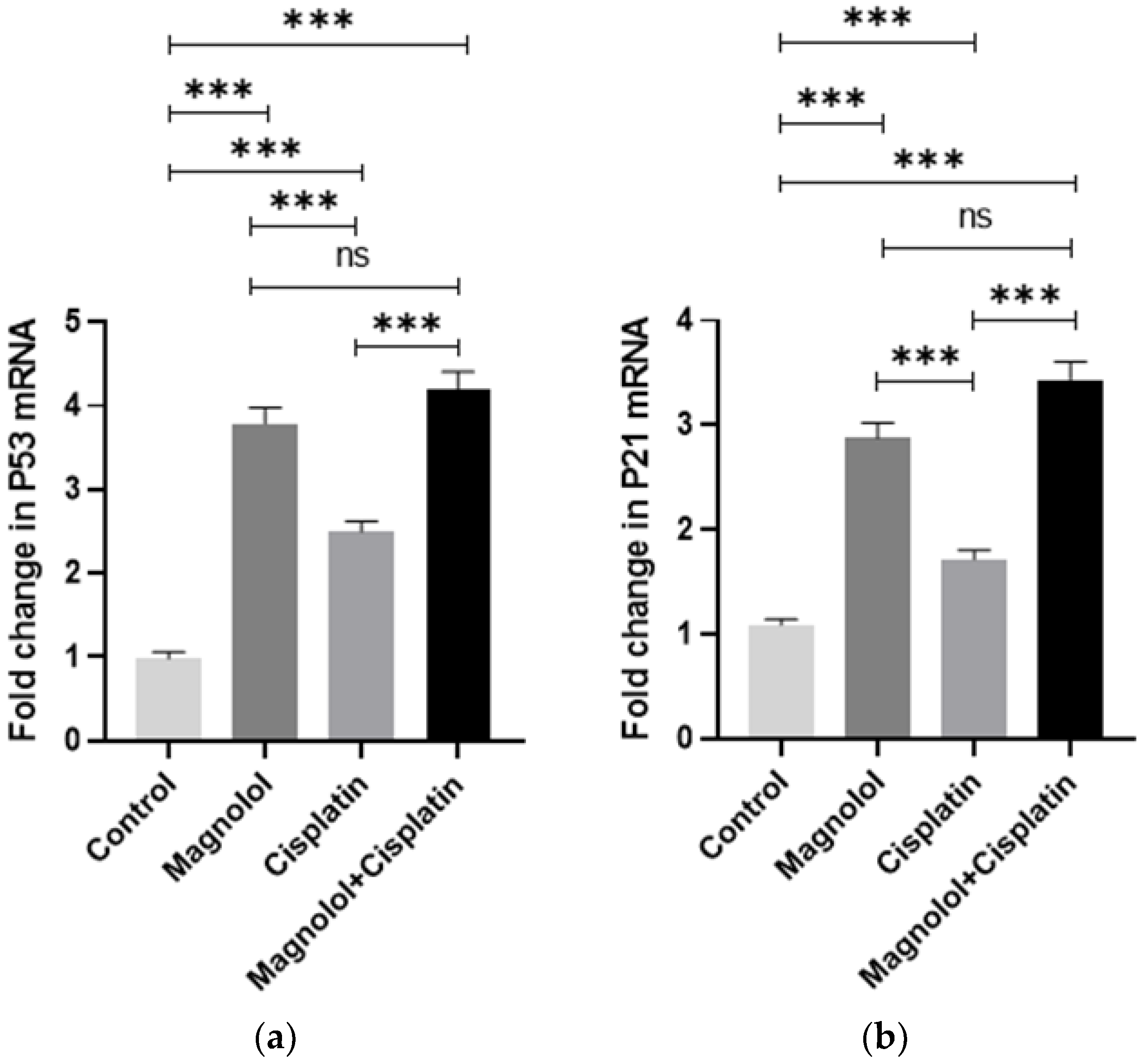
3.4. The Effects of Magnolol and/or Cisplatin on the Expression of Apoptosis-Dependent Genes
3.5. The Effects of Magnolol and/or Cisplatin on the Expression of Cell Cycle Regulator Genes
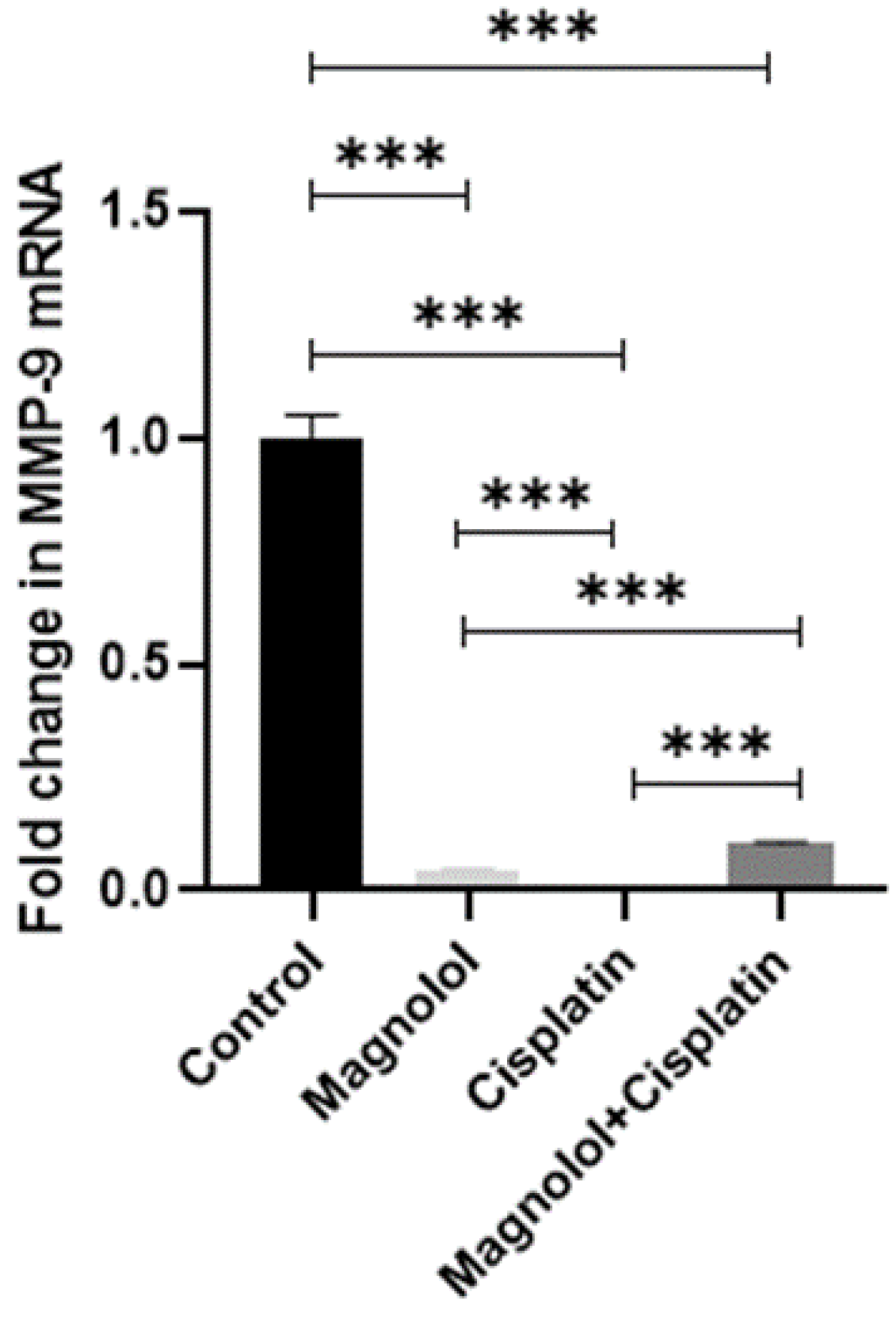
3.6. The Effects of Magnolol and/or Cisplatin on the Expression of Extracellular Matrix Remodeling Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, Y.; Shi, L.; He, X.; Luo, Y. Gastrointestinal cancers in China, the USA, and Europe. Gastroenterol. Rep. 2021, 9, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Ilic, M.; Ilic, I. Epidemiology of stomach cancer. World J. Gastroenterol. 2022, 28, 1187. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Li, N.; Luo, C.-Y.; Cai, J.; Lu, M.; Zhang, Y.-H.; Chen, H.-D.; Dai, M. Colorectal cancer incidence and mortality: The current status, temporal trends and their attributable risk factors in 60 countries in 2000–2019. Chin. Med. J. 2021, 134, 1941–1951. [Google Scholar] [CrossRef]
- Joshi, S.S.; Badgwell, B.D. Current treatment and recent progress in gastric cancer. CA Cancer J. Clin. 2021, 71, 264–279. [Google Scholar] [CrossRef]
- Lin, Z.; Cai, M.; Zhang, P.; Li, G.; Liu, T.; Li, X.; Cai, K.; Nie, X.; Wang, J.; Liu, J. Phase II, single-arm trial of preoperative short-course radiotherapy followed by chemotherapy and camrelizumab in locally advanced rectal cancer. J. Immunother. Cancer 2021, 9. [Google Scholar] [CrossRef]
- Lehtomäki, K.; Mustonen, H.; Kellokumpu-Lehtinen, P.-L.; Joensuu, H.; Hermunen, K.; Soveri, L.-M.; Boisen, M.K.; Dehlendorff, C.; Johansen, J.S.; Haglund, C. Lead time and prognostic role of serum CEA, CA19-9, IL-6, CRP, and YKL-40 after adjuvant chemotherapy in colorectal cancer. Cancers 2021, 13, 3892. [Google Scholar] [CrossRef]
- Wu, L.; Cai, S.; Deng, Y.; Zhang, Z.; Zhou, X.; Su, Y.; Xu, D. PD-1/PD-L1 enhanced cisplatin resistance in gastric cancer through PI3K/AKT mediated P-gp expression. Int. Immunopharmacol. 2021, 94, 107443. [Google Scholar] [CrossRef]
- Araújo, D.; Cabral, I.; Vale, N.; Amorim, I. Canine Gastric Cancer: Current Treatment Approaches. Vet. Sci. 2022, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Shetty, N.P.; Prabhakaran, M.; Srivastava, A.K. Pleiotropic nature of curcumin in targeting multiple apoptotic-mediated factors and related strategies to treat gastric cancer: A review. Phytother. Res. 2021, 35, 5397–5416. [Google Scholar] [CrossRef]
- Kanno, Y.; Chen, C.-Y.; Lee, H.-L.; Chiou, J.-F.; Chen, Y.-J. Molecular mechanisms of chemotherapy resistance in head and neck cancers. Front. Oncol. 2021, 11, 640392. [Google Scholar] [CrossRef]
- Liu, J.; Li, J.; Sun, Z.; Duan, Y.; Wang, F.; Wei, G.; Yang, J.-H. Bcl-2-associated transcription factor 1 Ser290 phosphorylation mediates DNA damage response and regulates radiosensitivity in gastric cancer. J. Transl. Med. 2021, 19, 339. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.S. A systematic review on multi-nutritional and phytopharmacological importance of Perilla frutescens. Int. J. Green Pharm. 2022, 16. [Google Scholar] [CrossRef]
- She, J.; Gu, T.; Pang, X.; Liu, Y.; Tang, L.; Zhou, X. Natural Products Targeting Liver X Receptors or Farnesoid X Receptor. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef]
- Mukherjee, S.; Dutta, A.; Chakraborty, A. External modulators and redox homeostasis: Scenario in radiation-induced bystander cells. Mutat. Res. Rev. Mutat. Res. 2021, 787, 108368. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, Y.; Sun, Z.; Ren, S.; Liu, M.; Wang, G.; Yang, J. Microglia in depression: An overview of microglia in the pathogenesis and treatment of depression. J. Neuroinflamm. 2022, 19, 132. [Google Scholar] [CrossRef]
- Yang, W.; Huang, G. Extraction methods and activities of natural glucans. Trends Food Sci. Technol. 2021, 112, 50–57. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhang, L.; Hou, X. Potential roles and molecular mechanisms of phytochemicals against cancer. Food Funct. 2022, 13, 9208–9225. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, L.; Guan, X.; Qin, J.-J. Inhibiting STAT3 Signaling Pathway by Natural Products for Cancer Prevention and Therapy: In Vitro and In Vivo Activity and Mechanisms of Action. Pharmacol. Res. 2022, 106357. [Google Scholar] [CrossRef]
- Zhu, D.; Gu, X.; Lin, Z.; Yu, D.; Wang, J. High expression of PSMC2 promotes gallbladder cancer through regulation of GNG4 and predicts poor prognosis. Oncogenesis 2021, 10, 43. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Wu, D.; Li, S.; Wang, C.; Han, Z.; Wang, J.; Wang, K.; Yang, Z.; Wei, Z. Magnolol prevents acute alcoholic liver damage by activating PI3K/Nrf2/PPARγ and inhibiting NLRP3 signaling pathway. Front. Pharmacol. 2019, 10, 1459. [Google Scholar] [CrossRef] [Green Version]
- Asif, P.J.; Longobardi, C.; Hahne, M.; Medema, J.P. The role of cancer-associated fibroblasts in cancer invasion and metastasis. Cancers 2021, 13, 4720. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cao, W.; Zhang, B.; Liu, Y.; Wang, Z.; Wu, Y.; Yu, X.; Zhang, X.; Ming, P.; Zhou, G. The natural compound magnolol inhibits invasion and exhibits potential in human breast cancer therapy. Sci. Rep. 2013, 3, 3098. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Hsu, F.-T.; Chen, W.-L.; Chen, J.-H. Induction of apoptosis, inhibition of MCL-1, and VEGF-A expression are associated with the anti-cancer efficacy of magnolol combined with regorafenib in hepatocellular carcinoma. Cancers 2021, 13, 2066. [Google Scholar] [CrossRef] [PubMed]
- Ranaware, A.M.; Banik, K.; Deshpande, V.; Padmavathi, G.; Roy, N.K.; Sethi, G.; Fan, L.; Kumar, A.P.; Kunnumakkara, A.B. Magnolol: A neolignan from the magnolia family for the prevention and treatment of cancer. Int. J. Mol. Sci. 2018, 19, 2362. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.-Y.; Yu, C.-C.; Huang, C.-C.; Liao, Y.-W.; Hsieh, P.-L.; Chu, P.-M.; Yu, C.-H.; Lin, S.-S. Magnolol inhibits cancer stemness and IL-6/Stat3 signaling in oral carcinomas. J. Formos. Med. Assoc. 2022, 121, 51–57. [Google Scholar] [CrossRef]
- Bijani, F.; Zabihi, E.; Bijani, A.; Nouri, H.R.; Nafarzadeh, S.; Seyedmajidi, M. Evaluation of apoptotic effect of crocin, cisplatin, and their combination in human oral squamous cell carcinoma cell line HN5. Dent. Res. J. 2021, 18, 70. [Google Scholar]
- Baharuddin, P.; Satar, N.; Fakiruddin, K.S.; Zakaria, N.; Lim, M.N.; Yusoff, N.M.; Zakaria, Z.; Yahaya, B.H. Curcumin improves the efficacy of cisplatin by targeting cancer stem-like cells through p21 and cyclin D1-mediated tumour cell inhibition in non-small cell lung cancer cell lines. Oncol. Rep. 2016, 35, 13–25. [Google Scholar] [CrossRef]
- Samie, K.A.; Dayer, D.; Eshkiki, Z.S. Human Colon Cancer HT29 Cell Line Treatment with High-Dose LAscorbic Acid Results to Reduced Angiogenic Proteins Expression and Elevated Pro-apoptotic Proteins Expression. In Current Molecular Medicine; Bentham Science Publishers: Bussum, The Netherlands, 2023. [Google Scholar]
- Dayer, D.; Tabandeh, M.R.; Kazemi, M. The radio-sensitizing effect of pharmacological concentration of ascorbic acid on human pancreatic Cancer cells. Anti-Cancer Agents Med. Chem. 2020, 20, 1927–1932. [Google Scholar] [CrossRef]
- Lee, K.-W.; Chung, I.-J.; Ryu, M.-H.; Park, Y.I.; Nam, B.-H.; Oh, H.-S.; Lee, K.H.; Han, H.S.; Seo, B.-G.; Jo, J.-C. Multicenter phase III trial of S-1 and cisplatin versus S-1 and oxaliplatin combination chemotherapy for first-line treatment of advanced gastric cancer (SOPP trial). Gastric Cancer 2021, 24, 156–167. [Google Scholar] [CrossRef]
- Ongnok, B.; Chattipakorn, N.; Chattipakorn, S.C. Doxorubicin and cisplatin induced cognitive impairment: The possible mechanisms and interventions. Exp. Neurol. 2020, 324, 113118. [Google Scholar] [CrossRef]
- Spirina, L.V.; Avgustinovich, A.V.; Afanas’ ev, S.G.; Cheremisina, O.V.; Volkov, M.Y.; Choynzonov, E.L.; Gorbunov, A.K.; Usynin, E.A. Molecular mechanism of resistance to chemotherapy in gastric cancers, the role of autophagy. Curr. Drug Targets 2020, 21, 713–721. [Google Scholar] [CrossRef]
- Peng, W.; Zhang, F.; Wang, Z.; Li, D.; He, Y.; Ning, Z.; Sheng, L.; Wang, J.; Xia, X.; Yu, C. Large scale, multicenter, prospective study of apatinib in advanced gastric cancer: A real-world study from China. Cancer Manag. Res. 2020, 12, 6977. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Yao, C.; Zhu, J.; Xie, Y.; Ye, X.-Y.; Bai, R.; Xie, T. Anti-tumor drug discovery based on natural product β-elemene: Anti-tumor mechanisms and structural modification. Molecules 2021, 26, 1499. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, K.; Ding, X.; Tang, H.; Xu, Z. Magnolol inhibits growth and induces apoptosis in esophagus cancer KYSE-150 cell lines via the MAP kinase pathway. J. Thorac. Dis. 2019, 11, 3030. [Google Scholar] [CrossRef] [PubMed]
- Rasul, A.; Yu, B.; Khan, M.; Zhang, K.; Iqbal, F.; Ma, T.; Yang, H. Magnolol, a natural compound, induces apoptosis of SGC-7901 human gastric adenocarcinoma cells via the mitochondrial and PI3K/Akt signaling pathways. Int. J. Oncol. 2012, 40, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Hyun, S.; Kim, M.S.; Song, Y.S.; Bak, Y.; Ham, S.Y.; Lee, D.H.; Hong, J.; Yoon, D.Y. Peroxisome proliferator-activated receptor-gamma agonist 4-O-methylhonokiol induces apoptosis by triggering the intrinsic apoptosis pathway and inhibiting the PI3K/Akt survival pathway in SiHa human cervical cancer cells. J. Microbiol. Biotechnol. 2015, 25, 334–342. [Google Scholar] [CrossRef]
- Ong, C.P.; Lee, W.L.; Tang, Y.Q.; Yap, W.H. Honokiol: A review of its anticancer potential and mechanisms. Cancers 2019, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.-q.; Fan, L.-y.; Yang, G.-l.; Guo, W.-H.; Hou, W.-l.; Chen, L.-j.; Wei, Y.-q. Improved therapeutic effectiveness by combining liposomal honokiol with cisplatin in lung cancer model. BMC Cancer 2008, 8, 242. [Google Scholar] [CrossRef]
- Park, J.B.; Lee, M.S.; Cha, E.Y.; Lee, J.S.; Sul, J.Y.; Song, I.S.; Kim, J.Y. Magnolol-induced apoptosis in HCT-116 colon cancer cells is associated with the AMP-activated protein kinase signaling pathway. Biol. Pharm. Bull. 2012, 35, 1614–1620. [Google Scholar] [CrossRef]
- Rasul, A.; Ding, C.; Li, X.; Khan, M.; Yi, F.; Ali, M.; Ma, T. Dracorhodin perchlorate inhibits PI3K/Akt and NF-κB activation, up-regulates the expression of p53, and enhances apoptosis. Apoptosis 2012, 17, 1104–1119. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, L.; Yi, T.; Xu, J.; Wang, J.; Qin, J.-J.; Chen, Q.; Yip, K.-M.; Pan, Y.; Hong, P. Synergistic effects of autophagy/mitophagy inhibitors and magnolol promote apoptosis and antitumor efficacy. Acta Pharm. Sin. B 2021, 11, 3966–3982. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.-R.; Chong, I.-W.; Chen, Y.-H.; Hwang, J.-J.; Yin, W.-H.; Chen, H.-L.; Chou, S.-H.; Chiu, C.-C.; Liu, P.-L. Magnolol induces apoptosis via caspase-independent pathways in non-small cell lung cancer cells. Arch. Pharmacal Res. 2014, 37, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.D.; Rubenstein, J.N.; Eggener, S.E.; Kozlowski, J.M. The p53 tumor suppressor gene and nuclear protein: Basic science review and relevance in the management of bladder cancer. J. Urol. 2003, 169, 1219–1228. [Google Scholar] [CrossRef] [PubMed]
- Shaltiel, I.A.; Aprelia, M.; Saurin, A.T.; Chowdhury, D.; Kops, G.J.; Voest, E.E.; Medema, R.H. Distinct phosphatases antagonize the p53 response in different phases of the cell cycle. Proc. Natl. Acad. Sci. USA 2014, 111, 7313–7318. [Google Scholar] [CrossRef]
- Tamura, G. Alterations of tumor suppressor and tumor-related genes in the development and progression of gastric cancer. World J. Gastroenterol. WJG 2006, 12, 192. [Google Scholar] [CrossRef]
- Lagger, G.; Doetzlhofer, A.; Schuettengruber, B.; Haidweger, E.; Simboeck, E.; Tischler, J.; Chiocca, S.; Suske, G.; Rotheneder, H.; Wintersberger, E. The tumor suppressor p53 and histone deacetylase 1 are antagonistic regulators of the cyclin-dependent kinase inhibitor p21/WAF1/CIP1 gene. Mol. Cell. Biol. 2003, 23, 2669–2679. [Google Scholar] [CrossRef]
- Nagaki, M.; Sugiyama, A.; Naiki, T.; Ohsawa, Y.; Moriwaki, H. Control of cyclins, cyclin-dependent kinase inhibitors, p21 and p27, and cell cycle progression in rat hepatocytes by extracellular matrix. J. Hepatol. 2000, 32, 488–496. [Google Scholar] [CrossRef]
- Zeestraten, E.C.; Benard, A.; Reimers, M.S.; Schouten, P.C.; Liefers, G.J.; Van de Velde, C.J.; Kuppen, P.J. The prognostic value of the apoptosis pathway in colorectal cancer: A review of the literature on biomarkers identified by immunohistochemistry. Biomark. Cancer 2013, 5, BIC-S11475. [Google Scholar] [CrossRef]
- Hsu, Y.F.; Lee, T.S.; Lin, S.Y.; Hsu, S.P.; Juan, S.H.; Hsu, Y.H.; Zhong, W.B.; Lee, W.S. Involvement of Ras/Raf-1/ERK actions in the magnolol-induced upregulation of p21 and cell-cycle arrest in colon cancer cells. Mol. Carcinog. Publ. Coop. Univ. Tex. MD Cancer Cent. 2007, 46, 275–283. [Google Scholar] [CrossRef]
- Shen, J.; Ma, H.; Zhang, T.; Liu, H.; Yu, L.; Li, G.; Li, H.; Hu, M. Magnolol inhibits the growth of non-small cell lung cancer via inhibiting microtubule polymerization. Cell. Physiol. Biochem. 2017, 42, 1789–1801. [Google Scholar] [CrossRef]
- He, Q.; Liu, Q.; Chen, Y.; Meng, J.; Zou, L. Long-zhi decoction medicated serum promotes angiogenesis in human umbilical vein endothelial cells based on autophagy. Evid.-Based Complement. Altern. Med. 2018, 2018, 6857398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Bi, Y.; Yang, C.; Yang, J.; Jiang, Y.; Meng, F.; Yu, B.; Khan, M.; Ma, T.; Yang, H. Magnolol induces apoptosis in MCF-7 human breast cancer cells through G2/M phase arrest and caspase-independent pathway. Die Pharm.-Int. J. Pharm. Sci. 2013, 68, 755–762. [Google Scholar]
- Jiao, L.; Bi, L.; Lu, Y.; Wang, Q.; Gong, Y.; Shi, J.; Xu, L. Cancer chemoprevention and therapy using chinese herbal medicine. Biol. Proced. Online 2018, 20, 1. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Jang, Y.S.; Min, S.Y.; Song, J.Y. Overexpression of MMP-9 and HIF-1α in breast cancer cells under hypoxic conditions. J. Breast Cancer 2011, 14, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Huang, S.; Yin, X.; Zan, Y.; Guo, Y.; Han, L. Quercetin suppresses the mobility of breast cancer by suppressing glycolysis through Akt-mTOR pathway mediated autophagy induction. Life Sci. 2018, 208, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xue, Z.; Yasin, A.; He, Y.; Chai, Y.; Li, J.; Zhang, K. Colorectal Cancer and Adjacent Normal Mucosa Differ in Apoptotic and Inflammatory Protein Expression. Eng. Regen. 2022, 2, 279–287. [Google Scholar] [CrossRef]
- Jin, T.; Li, D.; Yang, T.; Liu, F.; Kong, J.; Zhou, Y. PTPN1 promotes the progression of glioma by activating the MAPK/ERK and PI3K/AKT pathways and is associated with poor patient survival. Oncol. Rep. 2019, 42, 717–725. [Google Scholar] [CrossRef]
- Pezzuto, A.; Carico, E. Role of HIF-1 in cancer progression: Novel insights. A review. Curr. Mol. Med. 2018, 18, 343–351. [Google Scholar] [CrossRef]
- Patra, K.; Jana, S.; Sarkar, A.; Mandal, D.P.; Bhattacharjee, S. The inhibition of hypoxia-induced angiogenesis and metastasis by cinnamaldehyde is mediated by decreasing HIF-1α protein synthesis via PI3K/Akt pathway. Biofactors 2019, 45, 401–415. [Google Scholar] [CrossRef]
- Nagase, H.; Ikeda, K.; Sakai, Y. Inhibitory effect of magnolol and honokiol from Magnolia obovata on human fibrosarcoma HT-1080 invasiveness in vitro. Planta Med. 2001, 67, 705–708. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence |
|---|---|
| β-actin-hum-F | CAGCCTCAAGATCATCAGCAATG |
| β-actin-hum-R | CATGAGTCCTTCCACGATACCA |
| Bax-hum-F | AAGAAGCTGAGCGAGTGTCT |
| Bax-hum-R | TGCCGTCAGAAAACATGTCAG |
| MMP-9-hum-F | TAAGGAGTACTCGACCTGTACCA |
| MMP-9-hum-R | GAGGAACAAACTGTATCCTTGGTC |
| Bcl-2-hum-F | GGATGCCTTTGTGGAACTG |
| Bcl-2-hum-R | CAGCCAGGAGAAATCAAACAG |
| P53-hum-F | CAGACCTATGGAAACTACTTCCTG |
| P53-hum-R | ATTCTGGGAGCTTCATCTGGA |
| P21-hum-F | ATGTGGACCTGTCACTGTCTT |
| P21-hum-R | CGTTTGGAGTGGTAGAAATCTGTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naghashpour, M.; Dayer, D.; Karami, H.; Naghashpour, M.; Moghadam, M.T.; Haeri, S.M.J.; Suzuki, K. Evaluating the Magnolol Anticancer Potential in MKN-45 Gastric Cancer Cells. Medicina 2023, 59, 286. https://doi.org/10.3390/medicina59020286
Naghashpour M, Dayer D, Karami H, Naghashpour M, Moghadam MT, Haeri SMJ, Suzuki K. Evaluating the Magnolol Anticancer Potential in MKN-45 Gastric Cancer Cells. Medicina. 2023; 59(2):286. https://doi.org/10.3390/medicina59020286
Chicago/Turabian StyleNaghashpour, Mahsa, Dian Dayer, Hadi Karami, Mahshid Naghashpour, Mahin Taheri Moghadam, Seyed Mohammad Jafar Haeri, and Katsuhiko Suzuki. 2023. "Evaluating the Magnolol Anticancer Potential in MKN-45 Gastric Cancer Cells" Medicina 59, no. 2: 286. https://doi.org/10.3390/medicina59020286