
Comparative Genotypic Analysis of RAPD and RFLP Markers for Molecular Variation Detection of Methicillin-Resistant Staphylococcus aureus Clinical Isolates


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Phenotypic Identification of S. aureus
2.2. Phenotypic Recognition of MRSA
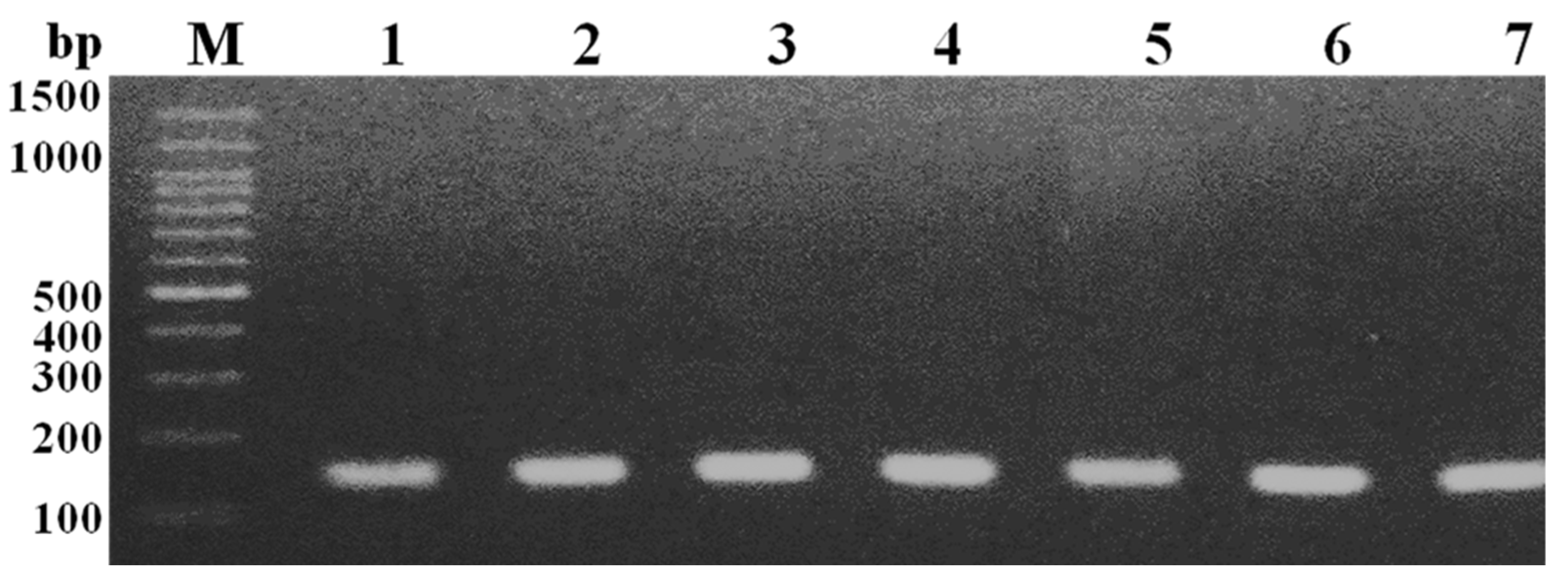
2.3. Genotypic Confirmation of MRSA
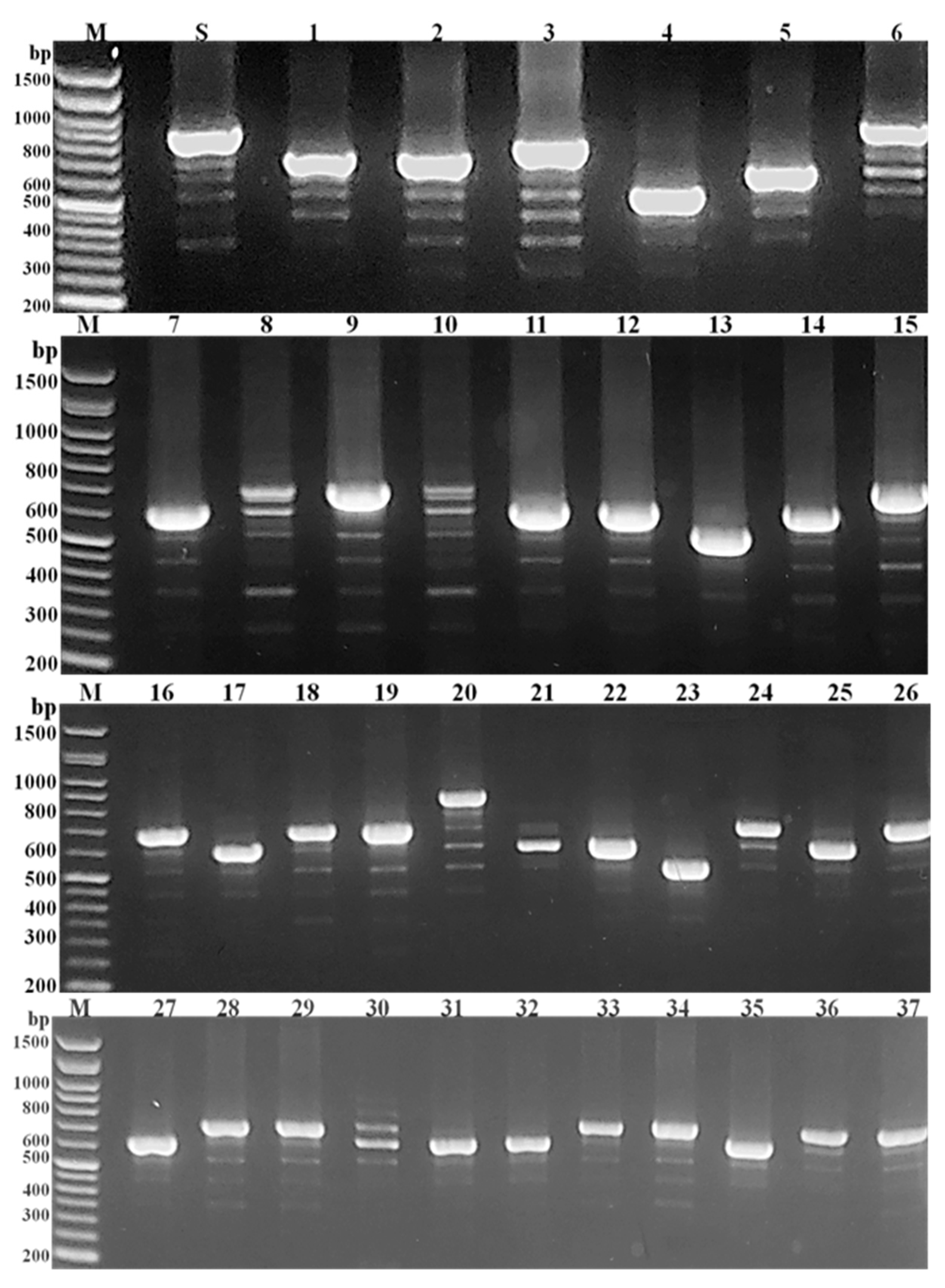
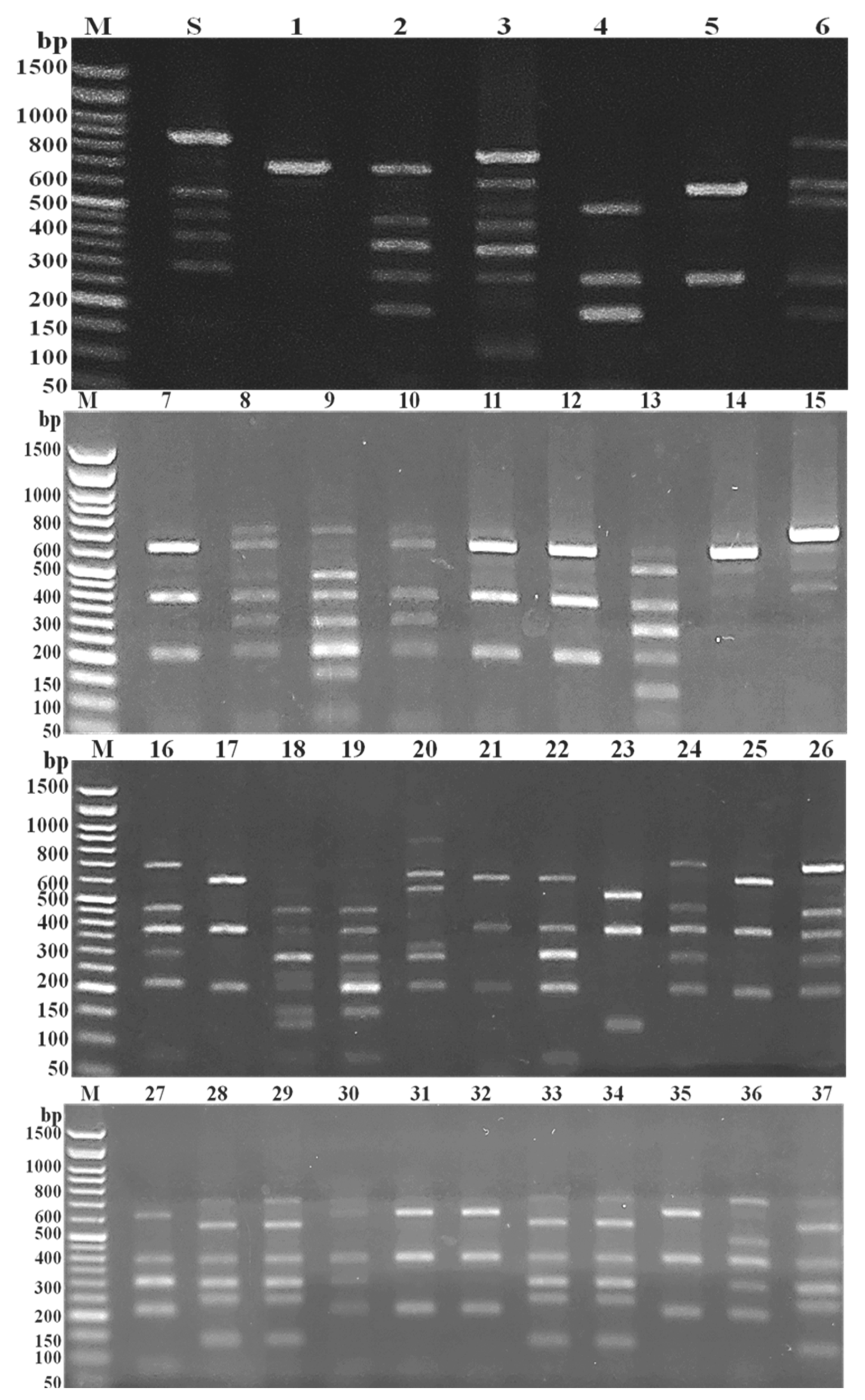
2.4. Coagulase (Coa) Gene Molecular Typing of MRSA Isolates
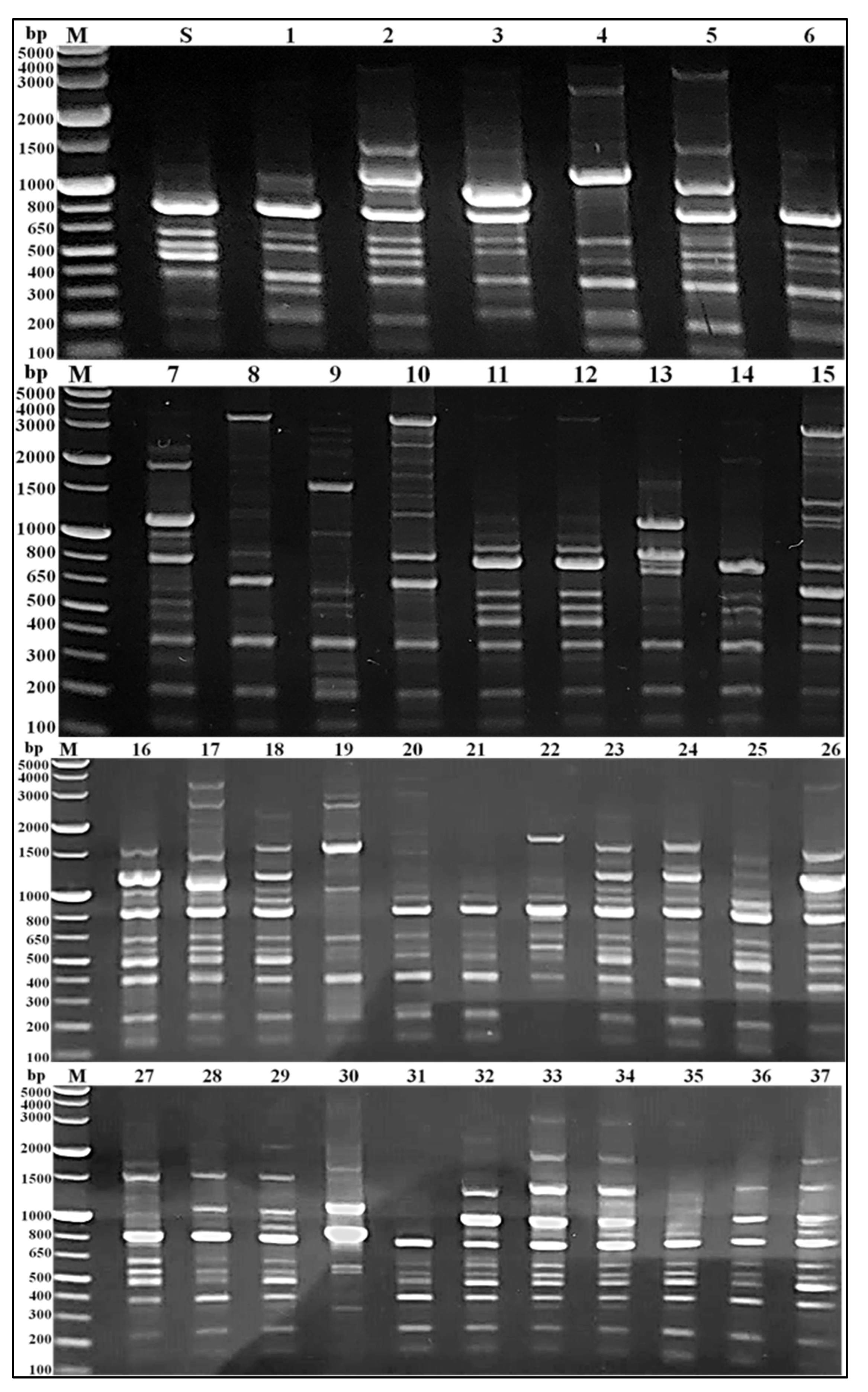
2.5. PCR-Based RAPD Fingerprinting of MRSA Isolates
2.6. Phylogenetic Analysis
2.7. Statistical Analysis
3. Results
3.1. Identification of MRSA
3.2. PCR-Based RFLP of Coa Gene
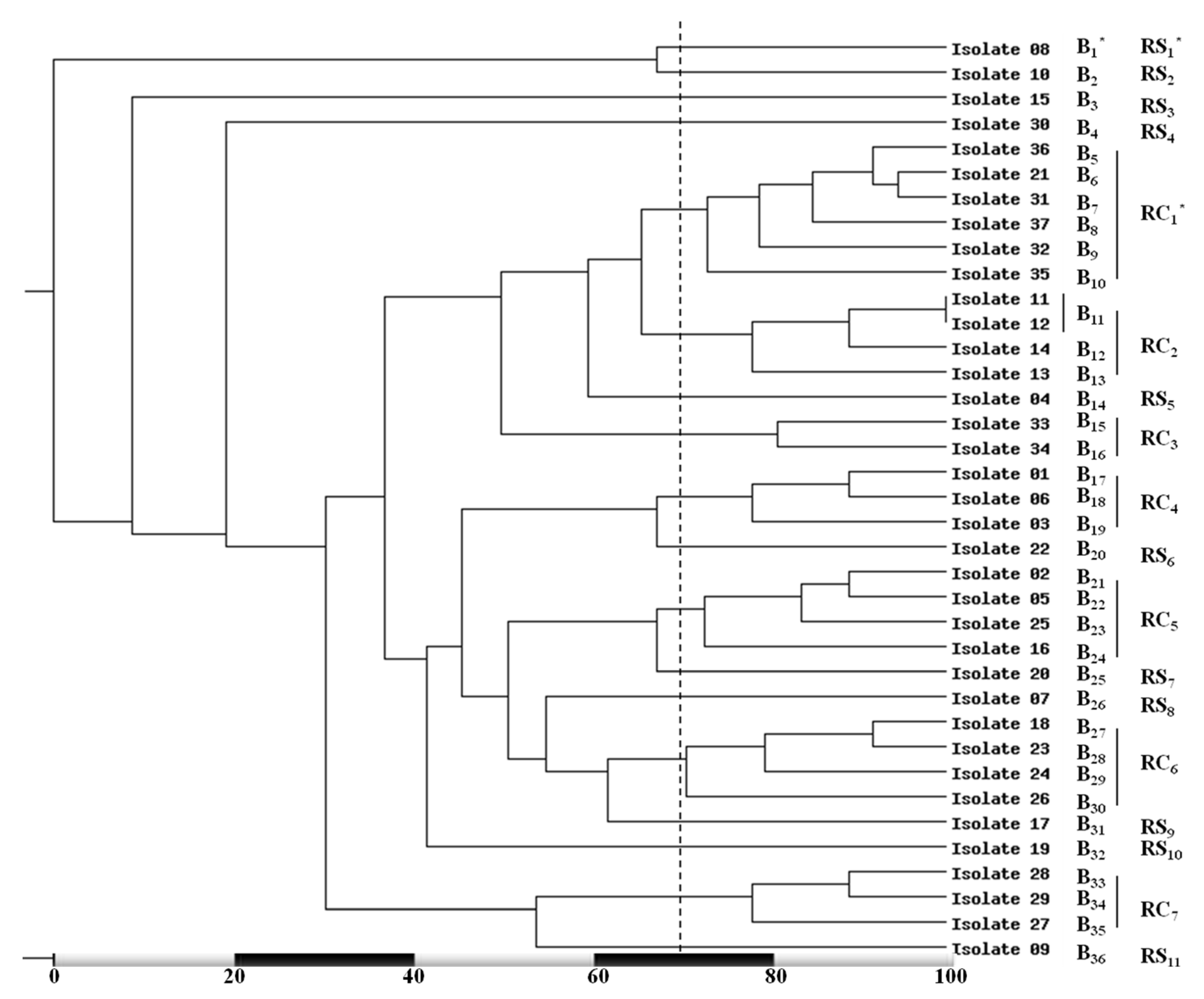
3.3. RAPD Fingerprinting of MRSA Isolates
3.4. Discrimination Index
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michalik, M.; Samet, A.; Podbielska-Kubera, A.; Savini, V.; Międzobrodzki, J.; Kosecka-Strojek, M. Coagulase-negative staphylococci (CoNS) as a significant etiological factor of laryngological infections: A review. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 26. [Google Scholar] [CrossRef] [PubMed]
- Salah, R.; Karmy, M.; Abdelraouf, A.; Kotb, S. Evaluation of the bactericidal effect of silver nanoparticles against Methicillin Resistant Staphylococcus aureus (MRSA) and Methicillin Sensitive Staphylococcus aureus (MSSA) strains isolated from mastitic milk of small ruminants and their surrounding environment in Aswan. J. Veter.-Med. Res. 2021, 27, 143–151. [Google Scholar] [CrossRef]
- Gajdács, M. The Continuing Threat of Methicillin-Resistant Staphylococcus aureus. Antibiotics 2019, 8, 52. [Google Scholar] [CrossRef] [PubMed]
- Liesenborghs, L.; Verhamme, P.; Vanassche, T. Staphylococcus aureus, master manipulator of the human hemostatic system. J. Thromb. Haemost. 2018, 16, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Abdeen, E.; Mousa, W.; Abdelsalam, S.; Heikal, H.; Shawish, R.; Nooruzzaman, M.; Soliman, M.; Batiha, G.; Hamad, A.; Abdeen, A. Prevalence and Characterization of Coagulase Positive Staphylococci from Food Products and Human Specimens in Egypt. Antibiotics 2021, 10, 75. [Google Scholar] [CrossRef] [PubMed]
- Tahmasbi, F.; Sheikhi, R.; Ashraf, A.; Mojtahedi, A. Biofilm formation and molecular characterization of methicillin-resistant Staphylococcus aureus strains isolated from the patients, personnel, air and environment of ICUs. Gene Rep. 2020, 20, 100736. [Google Scholar] [CrossRef]
- Stefańska, I.; Kwiecień, E.; Górzyńska, M.; Sałamaszyńska-Guz, A.; Rzewuska, M. RAPD-PCR-Based Fingerprinting Method as a Tool for Epidemiological Analysis of Trueperella pyogenes Infections. Pathogens 2022, 11, 562. [Google Scholar] [CrossRef]
- Dallal, M.M.S.; Khoramizadeh, M.R.; Amiri, S.A.; Yaraghi, A.A.S.; Fard, R.M.N. Coagulase gene polymorphism of Staphylococcus aureus isolates: A study on dairy food products and other foods in Tehran, Iran. Food Sci. Hum. Wellness 2016, 5, 186–190. [Google Scholar] [CrossRef]
- Javid, F.; Taku, A.; Bhat, M.A.; Badroo, G.A.; Mudasir, M.; Sofi, T.A. Molecular typing of Staphylococcus aureus based on coagulase gene. Veter.-World 2018, 11, 423–430. [Google Scholar] [CrossRef]
- Omar, N.Y.; Ali, H.A.S.; Harfoush, R.A.H.; El Khayat, E.H. Molecular Typing of Methicillin Resistant Staphylococcus aureus Clinical Isolates on the Basis of Protein A and Coagulase Gene Polymorphisms. Int. J. Microbiol. 2014, 2014, 650328. [Google Scholar] [CrossRef] [Green Version]
- Rahimi-Moghaddam, A.; Salmanzadeh-Ahrabi, S.; Tahereh, T.; Seifali, M.; Pourramezan, Z. Genotyping of Streptococcus Pyogenes Isolates using Optimized RAPD-PCR Protocol. Biol. J. Microorg. 2019, 8, 131–138. [Google Scholar]
- Abed, S.; Assie, A.; Abu-Elteen, K.; Dheeb, B.; Abu-Qatouseh, L. Molecular Characterization of Methicillin-Resistant and Methicillin- Susceptible Staphylococcus aureus Isolates Obtained from Human-Skin Samples in Iraq. Biomed. Pharmacol. J. 2020, 13, 737–746. [Google Scholar] [CrossRef]
- Fernandes Queiroga Moraes, G.; Cordeiro, L.V.; de Andrade Júnior, F.P. Main laboratory methods used for the isolation and identification of Staphylococcus spp. Rev. Colomb. Cienc. Químico-Farm. 2021, 50, 5–28. [Google Scholar]
- Madhavan, A.; Sachu, A.; Balakrishnan, A.; Vasudevan, A.; Balakrishnan, S.; Vasudevapanicker, J. Comparison of PCR and phenotypic methods for the detection of methicillin resistant Staphylococcus aureus. Iran. J. Microbiol. 2021, 13, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Fahim, K.M.; Khalf, M.A.M.; Nader, S.M.; Ismael, E. Impacts of Housing and Storage Environments on Physical Quality and the Potential Public Health Risks of Chicken Table Eggs. Adv. Anim. Veter.-Sci. 2021, 9, 1176–1189. [Google Scholar] [CrossRef]
- Mkhize, S.; Amoako, D.G.; Shobo, C.O.; Zishiri, O.T.; Bester, L.A. Genotypic and Phenotypic Characterizations of Methicillin-Resistant Staphylococcus aureus (MRSA) on Frequently Touched Sites from Public Hospitals in South Africa. Int. J. Microbiol. 2021, 2021, 6011045. [Google Scholar] [CrossRef]
- Sogut, M.U.; Bas, B.; Bilgin, M.; Sezener, M.G.; Findik, A. Molecular analysis and genotyping of methicillin-resistant Staphylococcus aureus strains isolated from different clinical sources. Niger. J. Clin. Pract. 2020, 23, 912. [Google Scholar] [CrossRef]
- Mahmoudi, H.; Arabestani, M.; Mousavi, S.F.; Alikhani, M.Y. Molecular analysis of the coagulase gene in clinical and nasal carrier isolates of methicillin-resistant Staphylococcus aureus by restriction fragment length polymorphism. J. Glob. Antimicrob. Resist. 2017, 8, 41–45. [Google Scholar] [CrossRef]
- Pacha, P.A.; Munoz, M.A.; González-Rocha, G.; Martín, I.S.; Quezada-Aguiluz, M.; Aguayo-Reyes, A.; Bello-Toledo, H.; Latorre, A.A. Molecular diversity of Staphylococcus aureus and the role of milking equipment adherences or biofilm as a source for bulk tank milk contamination. J. Dairy Sci. 2021, 104, 3522–3531. [Google Scholar] [CrossRef]
- Dhungel, S.; Rijal, K.R. Methicillin-Resistant Staphylococcus aureus (MRSA): Prevalence, Antimicrobial Susceptibility Pattern, and Detection of mecA Gene among Cardiac Patients from a Tertiary Care Heart Center in Kathmandu, Nepal. Infect. Dis. Res. Treat. 2021, 14, 11786337211037355. [Google Scholar] [CrossRef]
- Gharib, A.; Attia, A.; Bendary, M. Detection of the Coa Gene In Staphylococcus aureus From Different Sources By Polymerase Chain Reaction. Suez. Canal. Veter.-Med. J. SCVMJ 2013, 18, 167–177. [Google Scholar] [CrossRef]
- Bokharaei, N.M.; Dallal, M.S.; Pourmand, M.; Rajabi, Z. Antibiotic Resistance Pattern and Detection of mecA Gene in Staphylococcus aureus Isolated from Iranian Hamburger Samples. J. Food Qual. Hazards Control 2020, 7, 188–195. [Google Scholar] [CrossRef]
- Mohajeri, P.; Azizkhani, S.; Farahani, A.; Norozi, B. Genotyping of coa and aroA Genes of Methicillin-Resistant Staphylococcus aureus Strains Isolated From Nasal Samples in Western Iran. Jundishapur J. Microbiol. 2016, 9, e26460. [Google Scholar] [CrossRef] [PubMed]
- Abdulghany, H.M.; Khairy, R.M. The Frequency of Methicillin-Resistant Staphylococcus aureus and Coagulase Gene Polymorphism in Egypt. Int. J. Bacteriol. 2014, 2014, 680983. [Google Scholar] [CrossRef]
- A Al-Ruwaili, M. The coa, mec, and spa genes diversity among methicillin-resistant Staphylococcus aureus Strains from health-care workers and patients. Niger. J. Clin. Pract. 2018, 21, 1065–1074. [Google Scholar] [CrossRef]
- Naorem, R.S.; Urban, P.; Goswami, G.; Fekete, C. Characterization of methicillin-resistant Staphylococcus aureus through genomics approach. 3 Biotech 2020, 10, 401. [Google Scholar] [CrossRef]
- Al-Ajealy, B.A.; Al-Shukri, M.S.; Al-Jumaily, H.S. Detection of newly defined superantigenic toxin genes and coagulase gene polymorphism in Staphylococcus aureus isolates. Rev. Med. Microbiol. 2017, 28, 158–163. [Google Scholar] [CrossRef]
- Dendani, Z.C.; Bezille, P.; Arcangioli, M.-A. PCR and PCR-RFLP genotyping of Staphylococcus aureus coagulase gene: Convenience compared to pulse-field gel electrophoresis. Comp. Clin. Pathol. 2016, 25, 1061–1064. [Google Scholar] [CrossRef]
- Ibrahim, O.M.A.; Bilal, N.E.; Azoz, M.E.H.; Eltahir, H.B. Coagulase gene polymorphisms of Staphylococcus aureus isolates from patients at Kosti Teaching Hospital, Sudan. Access Microbiol. 2019, 1, e000026. [Google Scholar] [CrossRef]
- IL-Zengena, A.A.; Al-Taai, H.R.; Al-Dulaimi, A.A. Molecular and genetics analysis of coagulase (COA) gene polymorphism in clinical isolates of staphylococcus aureus by PCR-RFLP in patient of Baquba city, Iraq. Biochem. Cell. Arch. 2020, 20, 1513–1517. [Google Scholar]
- Khazaie, F.; Ahmadi, E. Bovine subclinical mastitis-associated methicillin-resistant Staphylococcus aureus, selective genotyping and antimicrobial susceptibility profile of the isolates in Kurdistan province of Iran. Iran. J. Microbiol. 2021, 13, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Mobasherizadeh, S.; Shojaei, H.; Havaei, S.A.; Mostafavizadeh, K.; Davoodabadi, F.; Khorvash, F.; Ataei, B.; Daei-Naser, A. Application of the Random Amplified Polymorphic DNA (RAPD) Fingerprinting to Analyze Genetic Variation in Community Associated-Methicillin Resistant Staphylococcus Aureus (CA-MRSA) Isolates in Iran. Glob. J. Health Sci. 2015, 8, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Halil, B.; Altanlar, N. Determination of Genetic Diversity and Similarity among Methicillin Resistant Staphylococcus aureus strains by RAPD-PCR. Turk. J. Sci. Health 2012, 3, 62–69. [Google Scholar]
- Hado, H.A.; Assafi, M.S. Molecular fingerprinting of methicillin resistant Staphylococcus aureus strains isolated from human and poultry in Duhok, Iraq. Iraqi J. Veter.-Sci. 2021, 35, 99–103. [Google Scholar] [CrossRef]
- Ye, Y.; Jiang, Q.; Wu, Q.; Zhang, J.; Lu, J.; Lin, L. The characterization and comparison of Staphylococcus aureus by antibiotic susceptibility testing, enterobacterial repetitive intergenic consensus–polymerase chain reaction, and random amplified polymorphic DNA–polymerase chain reaction. Foodborne Pathog. Dis. 2012, 9, 168–171. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elkady, F.M.; Al-Askar, A.A.; Tawab, A.A.; Alkherkhisy, M.M.; Arishi, A.A.; Hashem, A.H. Comparative Genotypic Analysis of RAPD and RFLP Markers for Molecular Variation Detection of Methicillin-Resistant Staphylococcus aureus Clinical Isolates. Medicina 2022, 58, 1245. https://doi.org/10.3390/medicina58091245
Elkady FM, Al-Askar AA, Tawab AA, Alkherkhisy MM, Arishi AA, Hashem AH. Comparative Genotypic Analysis of RAPD and RFLP Markers for Molecular Variation Detection of Methicillin-Resistant Staphylococcus aureus Clinical Isolates. Medicina. 2022; 58(9):1245. https://doi.org/10.3390/medicina58091245
Chicago/Turabian StyleElkady, Fathy M., Abdulaziz A. Al-Askar, Ahmed Abdel Tawab, Mohammad M. Alkherkhisy, Amr A. Arishi, and Amr H. Hashem. 2022. "Comparative Genotypic Analysis of RAPD and RFLP Markers for Molecular Variation Detection of Methicillin-Resistant Staphylococcus aureus Clinical Isolates" Medicina 58, no. 9: 1245. https://doi.org/10.3390/medicina58091245