
Cystitis Induces Altered CREB Expression Related with Micturition Reflex

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Induction of E. coli Cystitis
2.3. Microscopic Examination
2.4. Study Groups
2.5. Perfusion and Tissue Harvesting
2.6. Immunohistochemistry for p-CREB
2.7. Assessment of IR-Stained DRG Cells
2.8. Data Analysis
3. Results
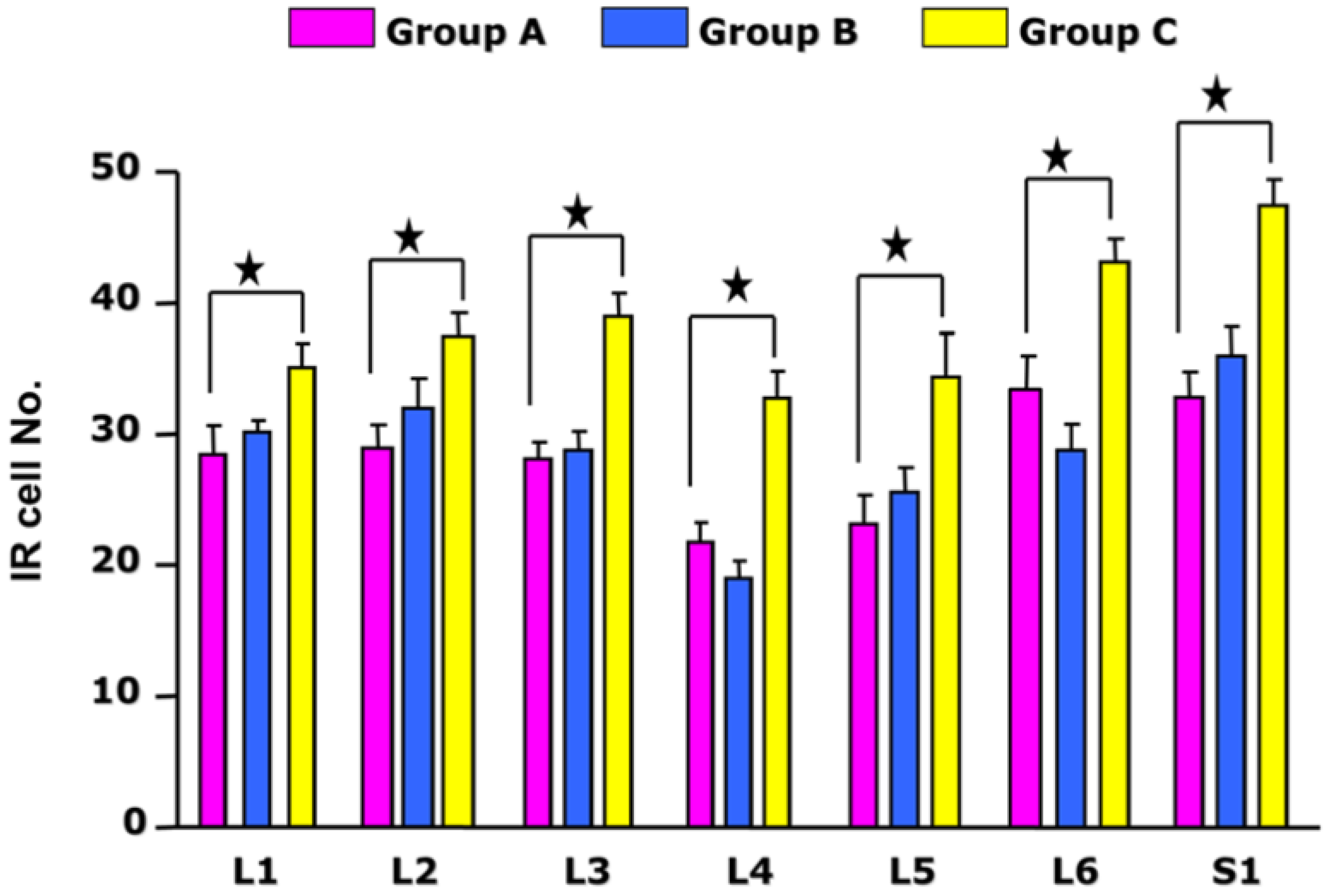
3.1. Quantitative Analysis of p-CREB in Each Segment of the DRG
3.2. Time Series Analysis of p-CREB IR in DRG with E. coli Cystitis
3.3. Changes in p-CREB IR in Bladder Afferent Cells with or without E. coli Cystitis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lowe, E.M.; Anand, P.; Terenghi, G.; Williams-Chestnut, R.E.; Sinicropi, D.V.; Osborne, J.L. Increased nerve growth factor levels in the urinary bladder of women with idiopathic sensory urgency and interstitial cystitis. Br. J. Urol. 1997, 79, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.C.; Fraser, M.O.; Yu, Y.; Chancellor, M.B.; de Groat, W.C.; Yoshimura, N. The role of bladder afferent pathways in bladder hyperactivity induced by the intravesical administration of nerve growth factor. J. Urol. 2001, 165, 975–979. [Google Scholar] [CrossRef]
- Vizzard, M.A. Changes in urinary bladder neurotrophic factor mRNA and NGF protein following urinary bladder dysfunction. Exp. Neurol. 2000, 161, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.Y.; Vizzard, M.A. Up-regulation of phosphorylated CREB but not c-Jun in bladder afferent neurons in dorsal root ganglia after cystitis. J. Comp. Neurol. 2004, 469, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Hawley, R.J.; Scheibe, R.J.; Wagner, J.A. NGF induces the expression of the VGF gene through a cAMP response element. J. Neurosci. 1992, 12, 2573–2581. [Google Scholar] [CrossRef] [PubMed]
- Riccio, A.; Ahn, S.; Davenport, C.M.; Blendy, J.A.; Ginty, D.D. Mediation by a CREB family transcription factor of NGF-dependent survival of sympathetic neurons. Science 1999, 286, 2358–2361. [Google Scholar] [CrossRef]
- Lee, D.G.; Cho, J.J.; Park, H.K.; Kim, D.K.; Kim, J.I.; Chang, S.G.; Lee, S.J. Preventive effects of hyaluronic acid on Escherichia coli-induced urinary tract infection in rat. Urology 2010, 75, 949–954. [Google Scholar] [CrossRef]
- Donovan, M.K.; Winternitz, S.R.; Wyss, J.M. An analysis of the sensory innervation of the urinary system in the rat. Brain Res. Bull. 1983, 11, 321–324. [Google Scholar] [CrossRef]
- Keast, J.R.; De Groat, W.C. Segmental distribution and peptide content of primary afferent neurons innervating the urogenital organs and colon of male rats. J. Comp. Neurol. 1992, 319, 615–623. [Google Scholar] [CrossRef]
- Qiao, L.Y.; Vizzard, M.A. Spinal cord injury-induced expression of TrkA, TrkB, phosphorylated CREB, and c-Jun in rat lumbosacral dorsal root ganglia. J. Comp. Neurol. 2005, 482, 142–154. [Google Scholar] [CrossRef]
- Vizzard, M.A. Alterations in neuropeptide expression in lumbosacral bladder pathways following chronic cystitis. J. Chem. Neuroanat. 2001, 21, 125–138. [Google Scholar] [CrossRef]
- Vizzard, M.A.; Erdman, S.L.; de Groat, W.C. Increased expression of neuronal nitric oxide synthase in bladder afferent pathways following chronic bladder irritation. J. Comp. Neurol. 1996, 370, 191–202. [Google Scholar] [CrossRef]
- Qiao, L.Y.; Vizzard, M.A. Cystitis-induced upregulation of tyrosine kinase (TrkA, TrkB) receptor expression and phosphorylation in rat micturition pathways. J. Comp. Neurol. 2002, 454, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Chancellor, M.B.; Yoshimura, N. Treatment of interstitial cystitis. Urology 2004, 63, 85–92. [Google Scholar] [CrossRef]
- Yoshimura, N.; de Groat, W.C. Increased excitability of afferent neurons innervating rat urinary bladder after chronic bladder inflammation. J. Neurosci. 1999, 19, 4644–4653. [Google Scholar] [CrossRef] [PubMed]
- Keay, S. Cell signaling in interstitial cystitis/painful bladder syndrome. Cell. Signal. 2008, 20, 2174–2179. [Google Scholar] [CrossRef]
- Ogawa, T.; Homma, T.; Igawa, Y.; Seki, S.; Ishizuka, O.; Imamura, T.; Akahane, S.; Homma, Y.; Nishizawa, O. CXCR3 binding chemokine and TNFSF14 over expression in bladder urothelium of patients with ulcerative interstitial cystitis. J. Urol. 2010, 183, 1206–1212. [Google Scholar] [CrossRef]
- Yoshimura, N.; Oguchi, T.; Yokoyama, H.; Funahashi, Y.; Yoshikawa, S.; Sugino, Y.; Kawamorita, N.; Kashyap, M.P.; Chancellor, M.B.; Tyagi, P.; et al. Bladder afferent hyperexcitability in bladder pain syndrome/interstitial cystitis. Int. J. Urol. 2014, 21 (Suppl. S1), 18–25. [Google Scholar] [CrossRef]
- Lishnak, T.S.; Vizzard, M.A. Phosphorylation of CREB in thoracolumbar spinal neurons and dorsal root ganglia after renal artery occlusion in rat. Auton. Neurosci. 2001, 94, 62–73. [Google Scholar] [CrossRef]
- Jenkins, R.; McMahon, S.B.; Bond, A.B.; Hunt, S.P. Expression of c-Jun as a response to dorsal root and peripheral nerve section in damaged and adjacent intact primary sensory neurons in the rat. Eur. J. Neurosci. 1993, 5, 751–759. [Google Scholar] [CrossRef]
- Kenney, A.M.; Kocsis, J.D. Peripheral axotomy induces long-term c-Jun amino-terminal kinase-1 activation and activator protein-1 binding activity by c-Jun and junD in adult rat dorsal root ganglia In vivo. J. Neurosci. 1998, 18, 1318–1328. [Google Scholar] [CrossRef] [PubMed]
- Broude, E.; McAtee, M.; Kelley, M.S.; Bregman, B.S. c-Jun expression in adult rat dorsal root ganglion neurons: Differential response after central or peripheral axotomy. Exp. Neurol. 1997, 148, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.R.; Rupp, F. Phosphorylation of transcription factor CREB in rat spinal cord after formalin-induced hyperalgesia: Relationship to c-fos induction. J. Neurosci. 1997, 17, 1776–1785. [Google Scholar] [CrossRef] [PubMed]
- Messersmith, D.J.; Kim, D.J.; Iadarola, M.J. Transcription factor regulation of prodynorphin gene expression following rat hindpaw inflammation. Brain Res. Mol. Brain Res. 1998, 53, 260–269. [Google Scholar] [CrossRef]
- De Leon, M.; Nahin, R.L.; Molina, C.A.; de Leon, D.D.; Ruda, M.A. Comparison of c-jun, junB, and junD mRNA expression and protein in the rat dorsal root ganglia following sciatic nerve transection. J. Neurosci. Res. 1995, 42, 391–401. [Google Scholar] [CrossRef]
- Steers, W.D.; de Groat, W.C. Effect of bladder outlet obstruction on micturition reflex pathways in the rat. J. Urol. 1988, 140, 864–871. [Google Scholar] [CrossRef]
- Kay, J.C.; Xia, C.M.; Liu, M.; Shen, S.; Yu, S.J.; Chung, C.; Qiao, L.Y. Endogenous PI3K/Akt and NMDAR act independently in the regulation of CREB activity in lumbosacral spinal cord in cystitis. Exp. Neurol. 2013, 250, 366–375. [Google Scholar] [CrossRef]
- Zhang, X.; Yao, J.; Gao, K.; Chi, Y.; Mitsui, T.; Ihara, T.; Sawada, N.; Kamiyama, M.; Fan, J.; Takeda, M. AMPK Suppresses Connexin43 Expression in the Bladder and Ameliorates Voiding Dysfunction in Cyclophosphamide-induced Mouse Cystitis. Sci. Rep. 2016, 6, 19708. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| DRG | Control (Mean ± SD) | Acute Cystitis (Mean ± SD) | Chronic Cystitis (Mean ± SD) |
|---|---|---|---|
| L1 | 28.69 ± 8.85 | 31.3 ± 4.42 | 33.9 ± 1.40 |
| L2 | 29.44 ± 8.41 | 32 ± 8.85 | 36.82 ± 1.49 |
| L3 | 28.33 ± 7.83 | 28.79 ± 5.86 | 38.15 ± 1.69 |
| L4 | 22.81 ± 5.04 | 20.17 ± 6.05 | 31.79 ± 2.07 |
| L5 | 23.85 ± 7.19 | 25.64 ± 7.26 | 33.25 ± 2.17 |
| L6 | 32.22 ± 7.33 | 29.14 ± 6.59 | 41.94 ± 1.94 |
| S1 | 31.4 ± 9.20 | 36 ± 8.49 | 42.57 ± 2.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, T.; Lee, D.-G. Cystitis Induces Altered CREB Expression Related with Micturition Reflex. Medicina 2022, 58, 1210. https://doi.org/10.3390/medicina58091210
Choi T, Lee D-G. Cystitis Induces Altered CREB Expression Related with Micturition Reflex. Medicina. 2022; 58(9):1210. https://doi.org/10.3390/medicina58091210
Chicago/Turabian StyleChoi, Taesoo, and Dong-Gi Lee. 2022. "Cystitis Induces Altered CREB Expression Related with Micturition Reflex" Medicina 58, no. 9: 1210. https://doi.org/10.3390/medicina58091210